Introduction
Recent studies have demonstrated that
pro-inflammatory mediators and pancreatic acinar cell apoptosis are
involved acute pancreatitis (AP) (1–3).
Apoptosis is characterized by distinct morphological and
biochemical changes, including cytoplasmic shrinkage, retention of
organelles, nuclear pycnosis, chromatin condensation and
margination, in addition to retention of integrity of the plasma
membranes, and thus no leakage of cellular contents to the
extracellular environment (4).
This programmed cell death is initiated in cells in response to
either extracellular cues via stimulation of the death receptors
(DRs), often referred to as the extrinsic pathway, or to a variety
of intracellular stress conditions that involve mitochondria and is
known as the intrinsic pathway (5). Certain members of the family of
cysteine aspartases (caspases) function as part of the apoptotic
pathway, and they are divided into initiators (caspases-2, −8, −9
and −10) and executioners (caspase-3, −6 and −7) (6,7).
Caspase-3 is an executioner caspase, activated by the intrinsic and
extrinsic pathways, which regulates the initial apoptotic process
(8,9). The B-cell lymphoma 2 (Bcl-2) family
is another functional component in the apoptosis pathway and it is
divided into pro-apoptotic [Bcl-2-like protein 4 (Bax), Bcl-2
homologous antagonist/killer and Bcl-2 related ovarian killer] and
anti-apoptotic [B-cell lymphoma 2 (Bcl-2), Bcl-2-extra large,
Bcl-2-like protein 2, myeloid cell leukemia sequence 1 and
Bcl-2-related protein A1] groups (10). The outcome for a cell in receipt of
an apoptotic stimulus is hypothesized to depend partly on the ratio
of pro- and anti-apoptotic members of the Bcl-2 family (11). Therefore, cellular injury and
apoptotic markers (cleaved caspase-3, Bcl-2 and Bax) were used in
the present study to determine the degree of caerulein
(CAE)-induced pancreatic cell apoptosis.
Previous studies have demonstrated that the classic
angiotensin-converting enzyme-angiotensin II type 1 receptor
(ACE-Ang II-AT1) axis activity increases and may aggravate the
development of pancreatitis (12–14).
Studies have demonstrated that ACE2-Ang-(1–7)-Mas
[ACE2-Ang-(1–7)-Mas] axis, the non-classical and the
depressor renin-Ang system (RAS) pathway, possess adverse effects
in numerous diseases and may be therapeutically useful (15,16).
However, little is known about the potential roles of
ACE2-Ang-(1–7)-Mas in apoptosis, and its precise
method of regulation in any organ system. Various studies have
demonstrated that Ang-(1–7) regulates alveolar epithelial cell
survival by inhibiting c-Jun N-terminal kinase phosphorylation and
apoptosis through the Ang-(1–7)
receptor Mas (17–20). Gaddam et al (12) identified that the Ang II to
Ang-(1–7) ratio increased in the pancreas,
suggesting dysregulation of RAS in AP as evidenced by altered Ang
II/Ang-(1–7) levels induced by the imbalance of
ACE/ACE2 activity. The purpose of the present study was to
determine whether the activation of the ACE2-Ang-(1–7)-Mas
axis possesses similarly positive effects on the digestive system,
particularly in pancreatic inflammatory disease, as it exhibits its
important function in the cardiovascular system.
Materials and methods
Cell culture
Mouse pancreatic acinar cancer cells (MPC-83; Cancer
Institute & Hospital, Chinese Academy of Medical Sciences,
Beijing, China) were cultured in RPMI-1640 medium (Hyclone; GE
Healthcare Life Sciences, Logan, UT, USA) supplemented with 10% FBS
(Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA), 100 U/ml
penicillin and 100 mg/ml streptomycin. Cells were incubated at 37°C
in a humidified atmosphere containing 5% CO2. All
experiments were conducted with RPMI-1640 medium, with or without
10 nmol/l CAE, for varying time periods. Cells were cultured in
culture bottles and petri dishes.
Cells in culture bottles were stimulated with 10
nmol/l CAE (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) for the
indicated time periods for AP (21), and the cells were divided into 4
groups: A, control; B, CAE; C, CAE + Ang-(1–7); and
D, CAE + A779 groups. Group A represented the normal growth of
MPC-83 cells without stimulation. Group B cells were harvested at
2, 6, 12, 24 and 48 h subsequent to being induced by 10 nmol/l CAE.
Groups C and D, based on the dynamic test outcomes of Group B, it
was identified that the expression of the cleaved caspase-3 protein
level reached its peak in cells stimulated by CAE at 24 h, and the
ratio of Bax and Bcl-2 reached its maximum at 6 h and remained
relatively high at 24 h however declined over time. Considering the
above, the MPC-83 cells in the CAE 24 h group (MPC-83 cells of the
CAE group harvested at 24 h) were selected for experimentation for
groups C and D. The cells were mock pretreated or pretreated with
different concentrations of Ang-(1–7) or
A779 (10−7, 10−6 or 10−5 mol/l)
for 30 min then were stimulated with 10 nmol/l CAE for 24 h.
Finally, cells were harvested for western blotting.
The cultivation steps of the cells in the petri
dishes were as described above. The harvested cells were seeded
onto glass coverslips in 6-well plates at 1×105
cells/well for immunocytochemistry, with 3 wells for each group.
These cells were used for immunofluorescence.
Immunofluorescence
Immunofluorescence staining was used to detect the
presence of ACE2, Mas receptor, caspase-3, Bax and Bcl-2 in MPC-83
cells. Briefly, MPC-83 cells were rinsed in PBS (pH 7.4; 138 mM
NaCl, 2.67 mM KCl, 8.10 mM Na2HPO4, 1.47 mM
KH2PO4) and fixed in 4% (v/v)
paraformaldehyde (40 min, 37°C). Following fixation, the cells were
blocked with 1% bovine serum albumin (Sigma-Aldrich; Merck KGaA)
for 30 min at 37°C. Then the cells were incubated with the
following primary antibodies: Anti-ACE2 (1:200; cat no. ab59351;
Abcam, Cambridge, MA, USA), anti-Mas receptor (1:100; cat no.
AAR-013; Alomone Labs, Jerusalem, Israel), anti-caspase-3 (1:100;
cat no. 9662; Cell Signaling Technology, Inc., Danvers, MA, USA),
anti-Bax (1:100; cat no. bs-0127R) and anti-Bcl-2 (1:100; cat no.
bs-0032R) (both from BIOSS, Beijing, China) at 4°C overnight. Goat
anti-rabbit fluorescein isothiocyanate (green fluorescent; 1:100;
cat no. bsF-0295G; BIOSS) probes were used as secondary antibody
and the cells were incubated with the secondary antibody for 40 min
at 37°C. Then the cell nucleus was stained with DAPI (1:1,000;
Santa Cruz Biotechnology, Inc., Dallas, TX, USA) for 5 min at 37°C.
Immunofluorescence was performed by visualization under a motorized
inverted system fluorescence microscope (Olympus IX81; Olympus
Corporation, Tokyo, Japan). Cells incubated with normal rabbit
serum (Sigma-Aldrich; Merck KGaA) instead of a primary antibody
served as a negative control.
Western blotting of ACE2, Mas
receptor, cleaved caspase-3, Bax and Bcl-2
Cells were washed three times with cold PBS (4°C),
followed by lysing the cells on ice with lysis buffer [pH 7.5; 150
mM NaCl, 50 mM Tris, 1 mM phenylmethylsulfonyl fluoride, 1 mM
Na3VO4, 1% NP-40, 0.1% SDS, 0.5% deoxycholic acid, 1% protease
inhibitor cocktail (BIOSS)] for 30 min. The total protein
concentration was determined with a bicinchoninic acid protein
assay kit (Pierce; Thermo Fisher Scientific, Inc.). Then the
protein (25 µg) was separated by 10% SDS-PAGE gel electrophoresis
and transferred onto PVDF membranes, which were blocked with 5%
non-fat dried milk for 2 h at 37°C and incubated overnight at 4°C
with rabbit anti-mouse ACE2 monoclonal antibody at 1:400 and mouse
β-actin monoclonal antibody at 1:500 (cat no. ab3280; Abcam),
rabbit anti-mouse Mas receptor polyclonal antibody at 1:200
(Alomone Labs) and rabbit anti-mouse caspase-3 monoclonal antibody
at 1:800 (Cell Signaling Technology, Inc.), and rabbit anti-mouse
Bax and Bcl-2 polyclonal antibody (1:200; BIOSS). All the
antibodies were diluted in TBS containing 0.05% Tween-20 (TBST).
Subsequent to washing three times with TBST, the membranes were
subsequently incubated for 1 h at room temperature with
peroxidase-conjugated goat anti-rabbit (1:8,000; cat no. sc-2004)
or goat anti-mouse (1:8,000; cat no. sc-2031) (both from Santa Cruz
Biotechnology, Inc.) secondary antibodies. The membranes were
washed three times with TBST and enhanced chemiluminescence (Merck
KGaA) was used to detect immunoreactive protein bands. Blots were
semi-quantified by densitometric analysis using Image Lab software
version 3.0 (Bio-Rad Laboratories, Inc., Hercules, CA, USA).
Statistical analysis
All experiments were repeated at least 3 times
independently. Data were expressed as the mean ± standard
deviation. Results were analyzed using SPSS software version 13.0
(SPSS, Inc., Chicago, IL, USA). The statistical significance of the
differences between groups was assessed using two-way analysis of
variance followed by post hoc Newman-Keuls test for multiple
comparisons. P<0.05 was considered to indicate a statistically
significant difference.
Results
Localization of ACE2, Mas receptor,
caspase-3, Bax, and Bcl-2 in MPC-83 cells
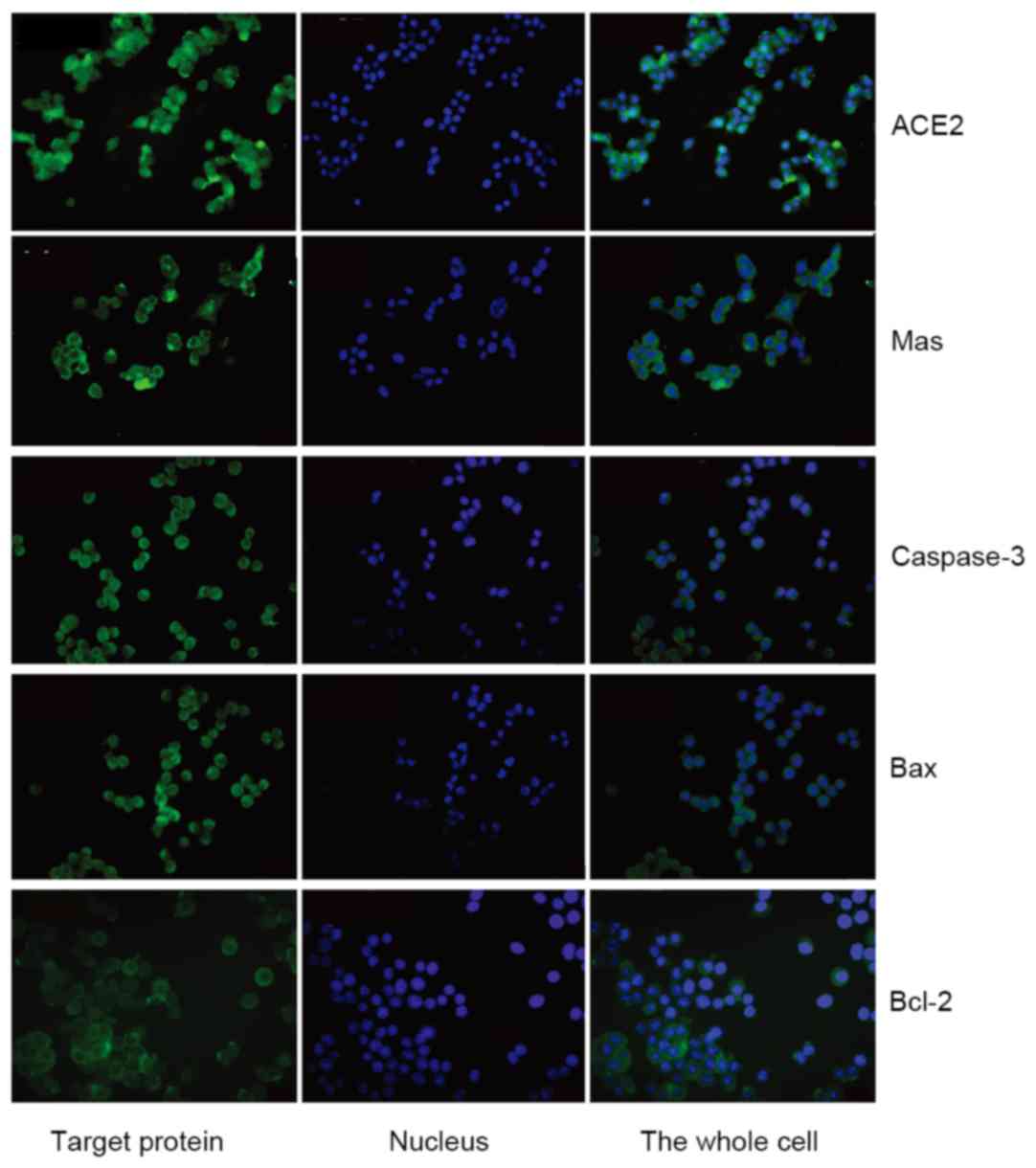
Because of its remarkable specificity, the
immunofluorescence assay is a valuable method for making a definite
diagnosis of the localization of ACE2, Mas receptor, caspase-3, Bax
and Bcl-2 in MPC-83 cells. Immunofluorescence demonstrated that
ACE2, caspase-3, Bax and Bcl-2 were present predominantly in the
cytoplasm, while Mas receptor was identified predominantly in the
membrane of the MPC-83 cells (Fig.
1).
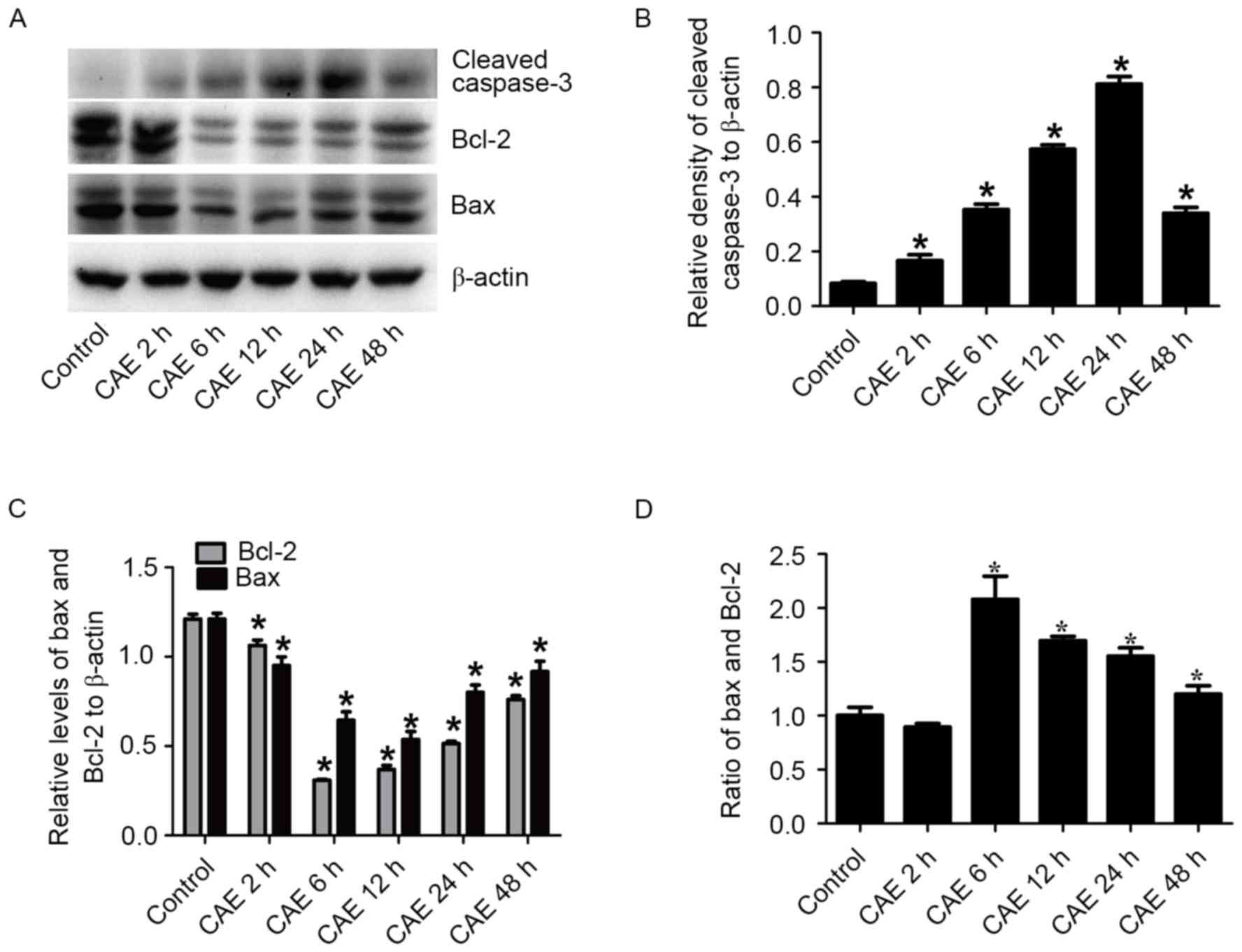
Dynamic protein levels of the cleaved
caspase-3, Bax and Bcl-2 in CAE group
Western blotting analysis indicated that exposure to
10 nmol/l CAE promoted cleaved caspase-3 expression, however
suppressed Bax and Bcl-2 protein levels (P<0.05; Fig. 2A-C). It exhibited a significant
evident increasing trend of the cleaved caspase-3 protein level
(P<0.05) and reached the highest value at 24 h (Fig. 2B). The ratio of Bax and Bcl-2
increased in the CAE group compared with the control group and
reached the highest value at 6 h (from 1.002±0.076 to 2.079±0.216;
P<0.05) then declined over time (Fig. 3D).
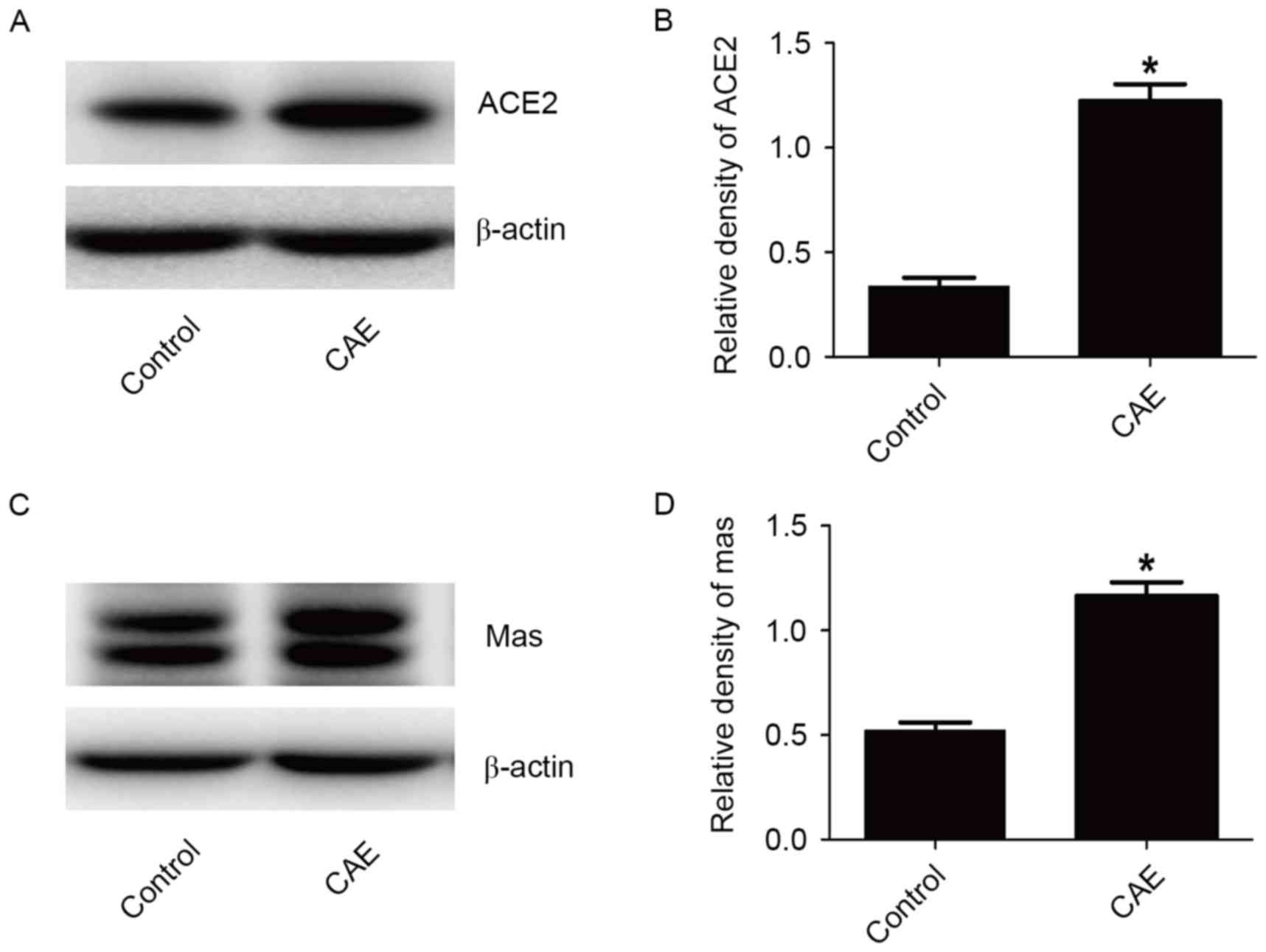
Protein profiles of ACE2, Mas receptor
in CAE 24 h group
ACE2 and Mas receptor protein expression in normal
MPC-83 cells was determined by western blotting, and quantified
with Image Lab software version 3.0 (Fig. 3; Bio-Rad Laboratories, Inc.). ACE2
and Mas receptor protein levels were significantly increased (from
0.340±0.067 to 1.222±0.142, P<0.05, and from 0.521±0.068 to
1.165±0.112, P<0.05) in the CAE 24 h group compared with the
control group.
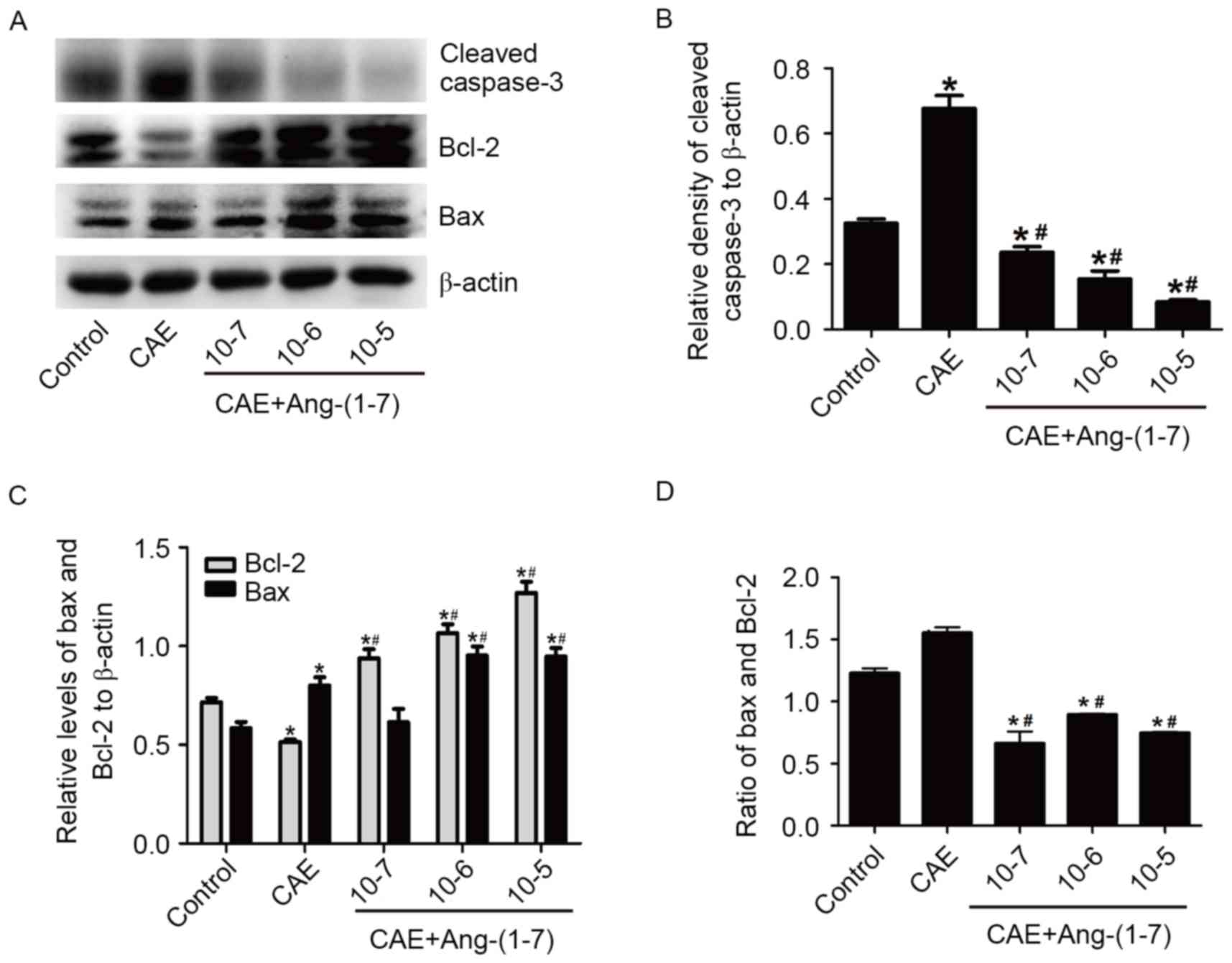
Ang-(1–7)
effects on the levels of the cleaved caspase-3, Bax and Bcl-2
To determine whether Ang-(1–7) has
positive effects on apoptosis, MPC-83 cells of the CAE group were
preincubated with different concentrations of Ang-(1–7)
(10−7, 10−6 and 10−5 mol/l) for 30
min. Soon afterwards the cleaved caspase-3, Bax and Bcl-2 protein
levels were detected. As expected, the cleaved caspase-3 protein
levels reduced in a dose-dependent manner and reached the lowest
level in the Ang-(1–7) 10−5 mol/l group compared
with the control and CAE groups (P<0.05; Fig. 4B). Fig. 4C demonstrates that Bcl-2 and Bax
had dose-dependent-rises (P<0.05). It is notable that the ratio
of Bax to Bcl-2 has a descending trend compared with the CAE and
the control group, suggesting that Ang-(1–7) had
anti-apoptotic potential (Fig.
4D).
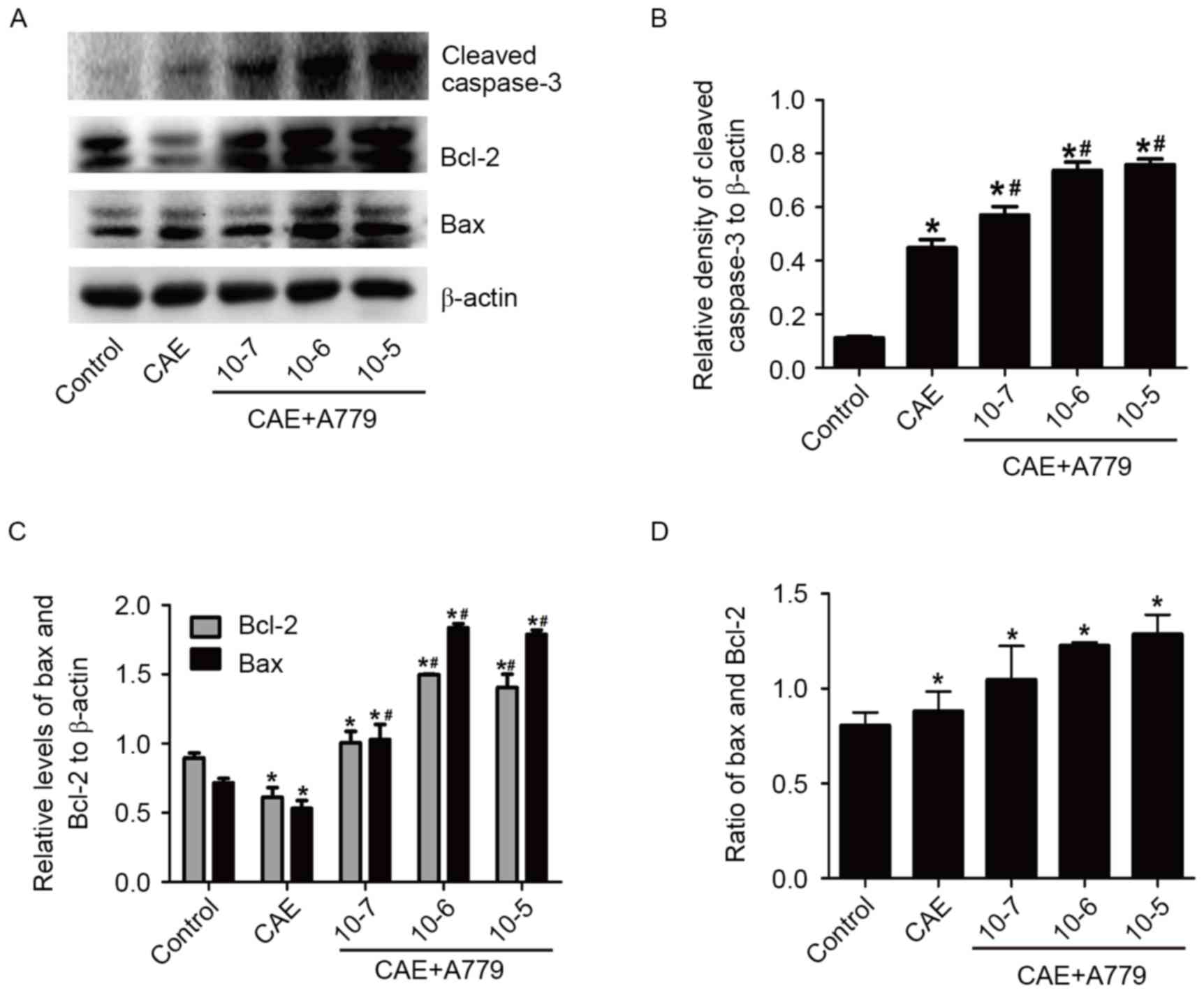
A779 influences the expressing
abilities of the cleaved caspase-3, Bax and Bcl-2
To investigate the possible protective effects of
ACE2-Ang-(1–7)-Mas axis on CAE-induced pancreatic
acinar cell apoptosis, the cells were pretreated with different
concentrations of A779 (10−7, 10−6 and
10−5 mol/l) for 30 min (Fig. 5). The A779 10−6 mol/l
and 10−5 mol/l groups demonstrated a significant
increase of the cleaved caspase-3 protein level compared with the
CAE group (P<0.05). In Fig. 5C and
D, it can be observed that while Bcl-2 and Bax increased in
A779 group, the increase in the level of pro-apoptotic factor Bax
was particularly pronounced compared with the anti-apoptotic factor
Bcl-2. The ratio of them increased dose-dependently from
0.880±0.180 (CAE group) to 1.285±0.179 (A779 10−5
group), P<0.05. The results demonstrated that A779,
Ang-(1–7) antagonist, possessed the potential to
promote apoptosis.
Discussion
In the present study, immunofluorescence assays
identified the presence of apoptotic factors in MPC-83 cells,
including caspase-3 (particularly cleaved caspase-3), pro-apoptotic
Bax and anti-apoptotic Bcl-2. It was observed that CAE upregulated
the expression of cleaved caspase-3, which is an executioner in
apoptosis. The ratio of Bax to Bcl-2, that is of pro-apoptotic to
anti-apoptotic proteins, significantly decreased in the
Ang-(1–7) group and increased in the A779 group.
These results suggested that Ang-(1–7) may
serve a distinct protective role in pancreatic acinar cell
apoptosis in the process of AP.
The ACE2-Ang-(1–7)-Mas
axis mediates a protective effect against the classic axis ACE-Ang
II-AT1 in cardiovascular disease, arthritis, liver fibrosis and
other diseases (22). Although
there are a number of potential pathways in the formation of
Ang-(1–7), it is suggested that that the
conversion of Ang II to Ang-(1–7) by
ACE2 is the primary physiological pathway for Ang-(1–7)
production (23). ACE2 may serve
an important counter regulatory role in the RAS pathway, protecting
cells from Ang II-mediated injury by inhibiting its production,
improving degradation rate and accelerating its conversion to
Ang-(1–7) (24).
Therefore, it was hypothesized that the upregulation of ACE2 and
Mas in AP MPC-83 cells may represent a protective response against
the functions of ACE and Ang II.
A previous study demonstrated that ACE2 and Mas
receptor protein levels were upregulated in CAE-induced pancreatic
acinar cells (25). We also
previously identified that ACE and ACE2 increased in acinar cells
in SAP mouse models, the imbalance between ACE and ACE2 possibly
contributing to pancreas injury by intensifying the severity of
pancreatitis (26). In the present
study, the ACE2-Ang-(1–7-Mas axis (the non-classical RAS
axis)-associated proteins of ACE2 and Mas receptor level
demonstrated a significant increase in CAE-induced pancreatic
acinar cells and Ang-(1–7) was able to curb cleaved caspase-3
protein expression and increase the anti-apoptotic factor Bcl-2
faster than the pro-apoptotic factor Bax, suggesting that
Ang-(1–7) possessed anti-apoptotic potential.
A779 was able to improve cleaved caspase-3 protein level and
enabled the pro-apoptotic factor Bax to increase more than the
anti-apoptotic factor Bcl-2. The results demonstrated that
Ang-(1–7) possessed the potential to inhibit
apoptosis and protect the pancreas from damage. These data,
together with a previous study (27), demonstrated that exogenous
Ang-(1–7) may affect the balance of Ang-(1–7)/Ang
II in addition to ACE2/ACE in AP. This is also consistent with
previous animal experiments; that identified that ulinastatin
significantly ameliorated pancreatic pathology through upregulating
ACE2 and Ang (1–7) in addition to the serum biochemistry
indicators of CAE-induced SAP mice (28). The re-establishment of the balance
of RAS through upregulating Ang-(1–7) and
ACE2 expression, which can ameliorate apoptosis, may possess
clinical therapeutic potential.
The observations of the current study may provide
evidence for the design of specific and powerful anti-apoptotic
agents that function by attenuating AP. The vast array of molecules
involved in apoptotic and necrotic pathways offer a number of
attractive therapeutic target sites. Considering the possibly
important role of Ang-(1–7) in pancreatic acinar cell apoptosis,
efforts should be made to exploit clinical strategies targeted to
the regulatory axis by taking adoptable methods including
upregulating the expression of ACE2 and/or Ang-(1–7).
Acknowledgements
The present study was supported by the Beijing
Natural Science Foundation (2014; grant no. 7142044) and the
National Natural Science Foundation of China (2016; grant no.
81571933).
Glossary
Abbreviations
Abbreviations:
|
ACE
|
angiotensin-converting enzyme
|
|
ACE2
|
angiotensin-converting enzyme 2
|
|
ACE2-Ang-(1–7)-Mas
|
ACE2-angiotensin-(1–7)-Mas
|
|
ACE-Ang II-AT1
|
angiotensin-converting
enzyme-angiotensin II type 1 receptor
|
|
AP
|
acute pancreatitis
|
|
CAE
|
caerulein
|
|
DRs
|
death receptors
|
|
MPC-83
|
mouse pancreatic acinar cancer
cells
|
|
RAS
|
renin-angiotensin system
|
References
|
1
|
Liu B, Huang J and Zhang B: Nobiletin
protects against murine l-arginine-induced acute pancreatitis in
association with downregulating p38MAPK and AKT. Biomed
Pharmacother. 81:104–110. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Fu Q, Qin T, Chen L, Liu CJ, Zhang X, Wang
YZ, Hu MX, Chu HY and Zhang HW: miR-29a up-regulation in AR42J
cells contributes to apoptosis via targeting TNFRSF1A gene. World J
Gastroenterol. 22:4881–4890. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Cai Y, Shen Y, Xu G, Tao R, Yuan W, Huang
Z and Zhang D: TRAM1 protects AR42J cells from caerulein-induced
acute pancreatitis through ER stress-apoptosis pathway. In Vitro
Cell Dev Biol Anim. 52:530–536. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yuan J, Liu Y, Tan T, Guha S, Gukovsky I,
Gukovskaya A and Pandol SJ: Protein kinase d regulates cell death
pathways in experimental pancreatitis. Front Physiol. 3:602012.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ashour AE, Ahmed AF, Kumar A, Zoheir KM,
Aboul-Soud MA, Ahmad SF, Attia SM, Abd-Allah AR, Cheryan VT and
Rishi AK: Thymoquinone inhibits growth of human medulloblastoma
cells by inducing oxidative stress and caspase-dependent apoptosis
while suppressing NF-κB signaling and IL-8 expression. Mol Cell
Biochem. 416:141–155. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chen K, Chu BZ, Liu F, Li B, Gao CM, Li
LL, Sun QS, Shen ZF and Jiang YY: New benzimidazole acridine
derivative induces human colon cancer cell apoptosis in vitro via
the ROS-JNK signaling pathway. Acta Pharmacol Sin. 36:1074–1084.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Mareninova OA, Sung KF, Hong P, Lugea A,
Pandol SJ, Gukovsky I and Gukovskaya AS: Cell death in
pancreatitis: Caspases protect from necrotizing pancreatitis. J
Biol Chem. 281:3370–3381. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Nuche-Berenguer B, Ramos-Álvarez I and
Jensen RT: Src kinases play a novel dual role in acute pancreatitis
affecting severity but no role in stimulated enzyme secretion. Am J
Physiol Gastrointest Liver Physiol. 310:G1015–G1027. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kobayashi C, Yaegaki K, Calenic B,
Ishkitiev N, Imai T, Ii H, Aoyama I, Kobayashi H, Izumi Y and
Haapasalo M: Hydrogen sulfide causes apoptosis in human pulp stem
cells. J Endod. 37:479–484. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ondroušková E and Vojtěšek B: Programmed
cell death in cancer cells. Klin Onkol. 27:(Suppl 1). S7–S14.
2014.(In Czech). View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Abogresha NM, Greish SM, Abdelaziz EZ and
Khalil WF: Remote effect of kidneyischemia-reperfusion injury on
pancreas: Role of oxidative stress and mitochondrial apoptosis.
Arch Med Sci. 12:252–262. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gaddam RR, Ang AD, Badiei A, Chambers ST
and Bhatia M: Alteration of the renin-angiotensin system in
caerulein induced acute pancreatitis in the mouse. Pancreatology.
15:647–653. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ibiş M, Yüksel O, Yilmaz G, Köklü S,
Yilmaz FM, Ertuğrul I, Uçar E and Altiparmak ME: Serum angiotensin
converting enzyme levels in pancreatic diseases.
Hepatogastroenterology. 55:1814–1817. 2008.PubMed/NCBI
|
|
14
|
Ulmasov B, Xu Z, Talkad V, Oshima K and
Neuschwander-Tetri BA: Angiotensin II signaling through the AT1a
and AT1b receptors does not have a role in the development of
cerulein-induced chronic pancreatitis in the mouse. Am J Physiol
Gastrointest Liver Physiol. 299:G70–G80. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Colafella KM, Hilliard LM and Denton KM:
Epochs in the depressor/pressor balance of the renin-angiotensin
system. Clin Sci (Lond). 130:761–771. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang J, Liu R, Qi H, Wang Y, Cui L, Wen Y,
Li H and Yin C: The ACE2-angiotensin-(1–7)-Mas axis protects
against pancreatic cell damage in cell culture. Pancreas.
44:266–272. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Gopallawa I and Uhal BD:
Angiotensin-(1–7)/mas inhibits apoptosis in alveolar epithelial
cells through upregulation of MAP kinase phosphatase-2. Am J
Physiol Lung Cell Mol Physiol. 310:L240–L248. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Uhal BD, Li X, Xue A, Gao X and
Abdul-Hafez A: Regulation of alveolar epithelial cell survival by
the ACE-2/angiotensin 1–7/Mas axis. Am J Physiol Lung Cell Mol
Physiol. 301:L269–L274. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Uhal BD, Nguyen H, Dang M, Gopallawa I,
Jiang J, Dang V, Ono S and Morimoto K: Abrogation of ER
stress-induced apoptosis of alveolar epithelial cells by
angiotensin 1–7. Am J Physiol Lung Cell Mol Physiol. 305:L33–L41.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ma X, Xu D, Ai Y, Zhao S, Zhang L, Ming G
and Liu Z: Angiotensin-(1–7)/Mas signaling inhibits
lipopolysaccharide-induced ADAM17 shedding activity and apoptosis
in alveolar epithelial cells. Pharmacology. 97:63–71. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yu JH, Lim JW and Kim H: Altered gene
expression in cerulein-stimulated pancreatic acinar cells:
Pathologic mechanism of acute pancreatitis. Korean J Physiol
Pharmacol. 13:409–416. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
da Silveira KD, Coelho FM, Vieira AT,
Sachs D, Barroso LC, Costa VV, Bretas TL, Bader M, de Sousa LP, da
Silva TA, et al: Anti-inflammatory effects of the activation of the
angiotensin-(1–7) receptor, MAS, in experimental models of
arthritis. J Immunol. 185:5569–5576. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Gallagher PE and Tallant EA: Inhibition of
human lung cancer cell growth by angiotensin-(1–7). Carcinogenesis.
25:2045–2052. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ohishi M, Yamamoto K and Rakugi H:
Angiotensin (1–7) and other angiotensin peptides. Curr Pharm Des.
19:3060–3064. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang Y, Wang J, Liu R, Qi H, Wen Y, Sun F
and Yin C: Severe acute pancreatitis is associated with
upregulation of the ACE2-angiotensin-(1–7)-Mas axis and promotes
increased circulating angiotensin-(1–7). Pancreatology. 12:451–457.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Liu R, Qi H, Wang J, Wang Y, Cui L, Wen Y
and Yin C: Angiotensin-converting enzyme (ACE and ACE2) imbalance
correlates with the severity of cerulein-induced acute pancreatitis
in mice. Exp Physiol. 99:651–663. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wang J, Liu R, Qi H, Wang Y, Cui L, Wen Y,
Li H and Yin C: The ACE2-angiotensin-(1–7)-Mas axis protects
against pancreatic cell damage in cell culture. Pancreas.
44:266–272. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Liu R, Qi H, Wang J, Wang Y, Cui L, Wen Y
and Yin C: Ulinastatin activates the renin-angiotensin system to
ameliorate the pathophysiology of severe acute pancreatitis. J
Gastroenterol Hepatol. 29:1328–1337. 2014. View Article : Google Scholar : PubMed/NCBI
|