Introduction
Cerebrovascular disorders are the third leading
cause of mortality worldwide, and the majority of affected patients
exhibit cerebral ischemia (1). The
recovery of blood flow to injured tissue has been demonstrated to
be the most effective therapeutic strategy to relieve the clinical
symptoms of cerebral ischemia, however, cerebral
ischemia-reperfusion (CIR) injury can occur, which is a more
serious clinical outcome. CIR injury presents a major medical
challenge and requires extensive investigation, as there are
currently few neuroprotective treatments (2,3).
Increasing evidence shows that oxidative stress is
crucial in facilitating neuronal death during CIR. During CIR
injury, abnormal mitochondrial activity produces high
concentrations of reactive oxygen species (ROS), and leads to cell
damage and eventual apoptosis or necrosis (4). Therefore, scavenging ROS may improve
the outcomes of CIR injury in humans. Although several agents,
including certain peptides and proteins, have shown promising in
vitro activities, disadvantages, including hydrophobicity,
antigenicity and large molecular size, impede the delivery of these
substances to brain tissues and thereby limit their therapeutic
benefits (5,6).
Deuterohemin His peptide-6 (DhHP-6;
Dh-β-AHTVEK-NH2), is a novel microperoxidase mimetic with a
molecular weight of 1,230 Da. It has been demonstrated that DhHP-6
can enter cells and can exhibit high enzyme-activity to scavenge
free radicals effectively (7,8).
Furthermore, DhHP-6 has been observed to increase survival rates in
Caenorhabditis elegans by promoting the elimination of
oxidative stress (9,10). The protective effects of DhHP-6 as
a scavenger of ROS have also been observed in myocardial
ischemia-reperfusion injury. The authors have observed that DhHP-6
also protects PC12 cells against H2O2-induced
oxidative stress injury (Yang et al, unpublished data).
However, whether DhHP-6 exhibits protective effects on CIR injury
in vivo remains to be elucidated. The aim of the present
study was to investigate the function of DhHP-6 and its underlying
mechanisms using a CIR rat model.
Materials and methods
Materials
DhHP-6 was provided by Dr Li Wei of the Life Science
College at Jilin University (Changchun, China) and had a purity of
>99%. Primary polyclonal antibodies against B-cell lymphoma-2
(Bcl-2; cat. no. ab59348), cleaved caspase-3 (cat. no. ab2302),
cytochrome c (cat. no. ab90529), Bcl-2-associated X protein (Bax;
cat. no. ab77566) and p-Akt (cat. no. ab8932) were purchased from
Abcam (Cambridge, UK). The biotinylated secondary antibodies (cat.
no. sc2357) were purchased from Santa Cruz Biotechnology, Inc.
(Santa Cruz, CA, USA). The catalase (CAT; cat. no. A007-1-1),
glutathione peroxidase (GSH-Px; cat. no. A005), superoxide
dismutase (SOD; cat. no. A001-3) and malondialdehyde (MDA; cat. no.
A003-1) assay kits were purchased from Jianchen Bioengineering
Institute (Nanjing, China). The TUNEL kit was acquired from Maixin
Bioengineering Institute (Fuzhou, China).
Animals
Male Wistar rats (6–8 weeks of age; 250–280 g) were
obtained from the Center of Laboratory Animal Science of Jilin
University. The rats were maintained in an animal house at a
temperature of 22±2°C in a 12-h light/dark cycle. The animals were
provided with food and water ad libitum. The protocol used in the
present study strictly adhered to the rules of the Jilin University
Animal Care and Use Committee for all procedures and followed the
guidelines outlined in the Principles of Laboratory Nursing of
Animals (11).
Experimental groups and drug
administration
A total of 80 male rats were used in the present
study. The rats were divided equally into two groups. One group was
used to measure neurological deficits and infarct area, and the
other group was used to evaluate ROS levels, MDA content, and
activities of CAT, SOD and GSH-Px. This group was also used for
performing TUNEL staining and western blot analysis. In each group,
40 rats were randomly separated into four subgroups (n=10):
Sham-operated group (Sham), ischemia-reperfusion group (I/R),
I/R+DhHP-6 (1 mg/kg) group (I/R+DH) and I/R+DhHP-6 (0.1 mg/kg)
group (I/R+DL). DhHP-6 was administered intraperitoneally at the
onset of reperfusion and it was only given once. The animals in the
sham and I/R groups were treated with saline in parallel.
Establishment of focal cerebral
ischemia-reperfusion
The occlusion and reperfusion model of the middle
cerebral artery (MCA) in rats was performed using the Longa method,
which has been described previously (12). In brief, 20% urethane (5 ml/kg) was
used to anesthetize the animals. The left common carotid artery,
the internal carotid artery (ICA) and the external carotid artery
were carefully exposed. A 3-0 monofilament nylon suture was
inserted into the left ICA, which was inserted ~18.0 mm from the
carotid artery bifurcation to obstruct the MCA. At 2 h post-MCAO,
reperfusion was performed by withdrawal of the nylon suture. During
surgery, the rectal temperature of the animals was maintained at
37.0±0.5°C. The animals in the sham group underwent the same
surgical procedure as those in the other animal groups, but did not
receive the nylon suture.
Neurological deficit assessment
Behavioral tests were performed 24 h following MCAO
by two investigators who were blinded to the experimental groups.
The scoring system was as follows: 0, no manifestation of
neurological dysfunction; 1, failure to fully stretch the forepaw
contralateral to the injured side, and failure to stretch and turn
the trunk towards the injured side when held by the tail; 2,
spinning motion toward the uninjured side; 3, failure to bear
weight on the injured side; and 4, inability to walk normally. When
the neurological deficit scores were higher, the disorders in
exercise behavior were more severe (13,14).
Evaluation of infarct area
The brains of the rats anesthetized with 20%
urethane (5 ml/kg i.p.) were removed and immediately placed on a
frozen surface for 5 min. Each brain was then cut into five
2-mm-thick coronal slices and stained at 37°C for 30 min in 2%
triphenyltetrazolium chloride (Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany). Infarct volume was detected using image
analysis software (Image Pro plus 6.0 system; Media Cybernetics,
Inc., Rockville, MD, USA). To avoid data errors caused by brain
edema, the following formula was used to evaluate infarct area:
Infarct volume (%) = (contralateral volume - ipsilateral
non-infarct volume)/contralateral volume.
Assessment of brain edema
The wet-dry method was used to assess brain edema.
Briefly, fresh tissue was weighed immediately following isolation
(termed wet weight). The tissues were then dried at 100°C in the
oven for 24 h, followed by weighing again (termed dry weight). The
percentage of brain water content was evaluated using the following
formula: Brain water content (%) = (wet weight - dry weight)/wet
weight × 100% (1).
Evaluation of ROS levels in the
brain
The tissue samples from the ischemic cerebral cortex
were homogenized in cold isolation buffer containing 0.25 M
sucrose, 10 mM Tris-HCl, 0.5 M EDTA and 250 µg/ml BSA (pH 7.1). The
cell fragments and nuclei were centrifuged at 4,000 × g for 3 min
at 4°C and then at 1,000 × g for 5 min at 4°C. The supernatants
were then centrifuged again for 15 min at 10,000 × g at 4°C.
Mitochondria were suspended in the isolation buffer. Mitochondria
isolated from different groups (0.5 mg protein, measured with
biocinchoninic acid protein assay kit) were incubated with 10 µM
DCFH-DA (Beyotime Institute of Biotechnology, Shanghai, China) at
37°C for 15 min in the dark. The levels of intracellular ROS were
evaluated using flow cytometry (BD Biosciences, Franklin Lakes, NJ,
USA) at an emission wavelength of 525 nm and an excitation
wavelength of 488 nm (15,16).
Measurement of MDA content and
activities of SOD, CAT and GSH-Px
Tissue samples of the ischemic cortex were
homogenized in ice-cold saline and then centrifuged at 2,000 × g
for 15 min at 4°C to obtain 10% homogenates. The concentration of
MDA was determined using an assay kit (Jiancheng Biotechnology
Institute, Nanjing, China) according to the manufacturer's
protocol. The activities of CAT, SOD and GSH-Px were also
determined using kits (Jiancheng Biotechnology Institute). The
protein concentrations were evaluated using a bicinchoninic acid
protein assay kit (Thermo Fisher Scientific, Inc., Walham, MA,
USA).
TUNEL staining
TUNEL-staining was performed using an in situ
apoptosis detection kit (Maixin Biotechnology, Fuzhou, China)
according to the manufacturer's protocol. The tissue sections were
incubated with proteinase K (20 µg/ml) for 15 min and 0.3%
H2O2 for 30 min at 37°C. The sections were
then treated with terminal deoxyribonucleotidy ltransferase enzyme
at 37°C for 1 h and peroxidase-conjugated antibody for at 37°C for
30 min. To terminate the reaction, the sections were incubated in a
reaction termination buffer for 5 min at room temperature. The
sections were visualized following staining with diaminobenzidinete
trahydrochloride. Apoptotic cells exhibited brown particles in the
nuclei. A total of five visual fields in the cerebral cortex were
analyzed from each section. TUNEL-positive cells were counted using
an optical microscope (Nikon Ti; Nikon Corporation, Tokyo, Japan).
The total number of cells and the number of TUNEL-positive cells
were assessed in each field. The percentage of TUNEL-positive cells
was calculated as follows: TUNEL-positive cells (%) = number of
positive cells/number of total cells × 100% (17).
Western blot analysis
Protein samples were prepared from the ischemic
cortex by homogenization in ice-cold lysate buffer (Beyotime
Institute of Biotechnology). The mitochondrial and cytosolic
proteins were prepared as previously described (18). To detect the expression of Bax,
cytochrome c and Bcl-2, the mitochondrial and cytosolic protein
extracts were evaluated independently. The expression of cleaved
caspase-3 was detected using the total protein lysates. The total
protein samples (60 µg) were separated on 12% SDS polyacrylamide
gels and were transferred onto polyvinylidenedifluoride membranes.
The membranes were blocked with 5% BSA for 2 h at room temperature,
and then were incubated overnight at 4°C with rabbit polyclonal
antibodies Bcl-2 (1:1,000), Bax (1:1,000), cytochrome c (1:1,000),
cleaved caspase-3 (1:500) and p-Akt (1:500) antibodies, (all from
Santa Cruz Biotechnology, Inc.). The membranes were then incubated
with secondary antibody (goat anti-rabbit; 1:1,000; Santa Cruz
Biotechnology, Inc.) for 2 h at room temperature. The
immunoreactive bands were visualized using enhanced
chemiluminescence. Images of the protein bands were captured using
an imaging densitometer and were quantified using image analysis
software (Quantity One; version 4.62; Bio-Rad Laboratories, Inc.,
Hercules, CA, USA). The protein values were normalized to those of
β-actin in the same lane.
Statistical analysis
All values are presented as the mean ± standard
deviation. Significant differences between groups were performed
using one-way analysis of variance followed by the Tukey's post hoc
test. GraphPad Prism statistical software (version 5.0; GraphPad
Software, Inc., La Jolla, CA, USA) was used for statistical
analysis. P<0.05 was considered to indicate a statistically
significant difference.
Results
DhHP-6 reduces infarct volume
MCAO produced a well-defined infarct, which included
the cortex and striatum (Fig. 1A and
B). In the I/R group, 2 h of cerebral ischemia and 22 h of
reperfusion caused infarction, which involved 35.28±4.13% of the
ipsilateral cerebral hemisphere. The infarct volume in the I/R+DH
group was significantly reduced to 23.56±3.47% (P<0.05). The
I/R+DL group also exhibited a reduced infarct volume, however, this
was not a significant difference (P>0.05).
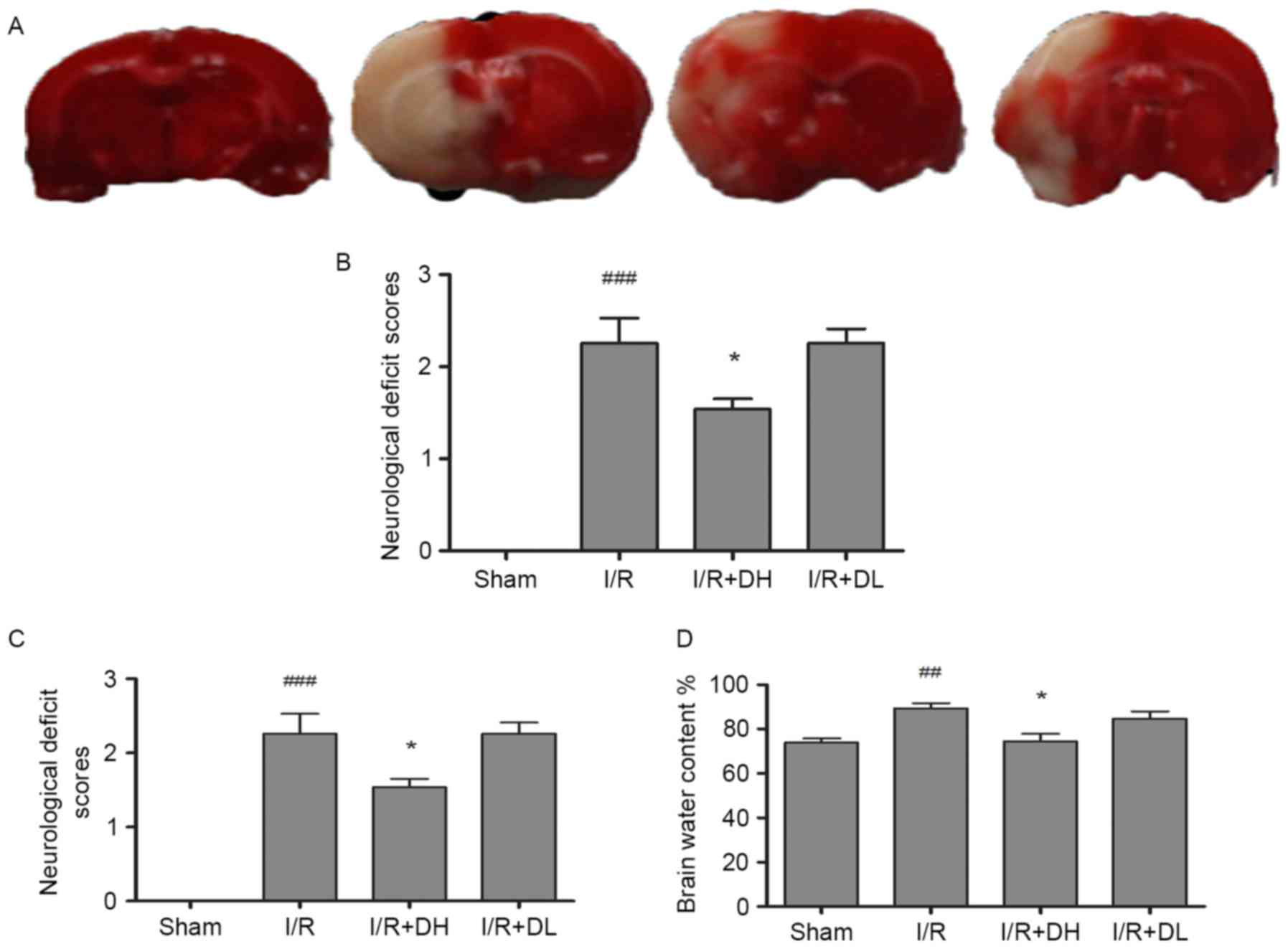 | Figure 1.Effect of DhHP-6 on cerebral ischemia
induced by 2 h of middle cerebral artery occlusion and 22 h of
reperfusion. (A) Triphenyltetrazolium chloride staining of coronal
brain sections 22 h following reperfusion. Infarct brain tissues
appeared unstained. Tissues in the I/R+DH group, but not the I/R+DL
group, demonstrated significant reductions in (B) infarct volume,
(C) neurological deficits and (D) brain edema, in a dose-dependent
manner. Data are shown as the mean ± standard deviation. One-way
analysis of variance and Tukey's post hoc test were performed.
###P<0.001 and ##P<0.01, vs. Sham
group; *P<0.05, vs. I/R group (n=10). I/R, ischemia/reperfusion;
DhHP-6, deuterohemin His peptide-6; DH, 1 mg/kg/day DhHP-6; DL, 0.1
mg/kg/day DhHP-6. |
DhHP-6 decreases neurological
dysfunction
Neurological symptoms were evaluated 22 h following
reperfusion (Fig. 1C). Normal
reflexes were present in all animals in the sham group (score 0),
where as animals in the I/R group presented with severe
neurological deficits (2.34±0.15; P<0.001). Compared with the
I/R group, the neurological deficit scores were markedly reduced to
1.57±0.28 in the I/R+DH group (P<0.05), but did not differ
significantly from those in the I/R+DL group (P>0.05).
DhHP-6 decreases brain edema
As shown in Fig.
1D, MCAO led to an increase in brain water content in the right
hemisphere of the I/R group, however, the level of brain water
content was significantly reduced in the I/R+DH group (P<0.05)
in comparison.
DhHP-6 reduces the oxidative stress
caused by MCAO
DhHP-6 acts as a free radical scavenger (9). Therefore, the present study aimed to
determine whether the levels of free radicals and oxidative stress
were reduced by DhHP-6 treatment following I/R. To evaluate this,
the levels of ROS and MDA, which is a lipid peroxidation product
caused by oxidative stress, were examined. The activities of
antioxidant enzymes, including CAT, SOD and GSH-Px, were also
measured.
The ROS level was determined as the percentage of
fluorescence intensity, compared with that in the sham group
(Fig. 2A). The mitochondrial DCF
fluorescence of the I/R group exceeded that of the shamgroup
(P<0.01). The levels of ROS in the mitochondria were decrease by
21.34% in the I/R+DH group. Compared with the I/R group, the I/R+DH
group showed a significant decrease in mitochondrial ROS
(P<0.05).
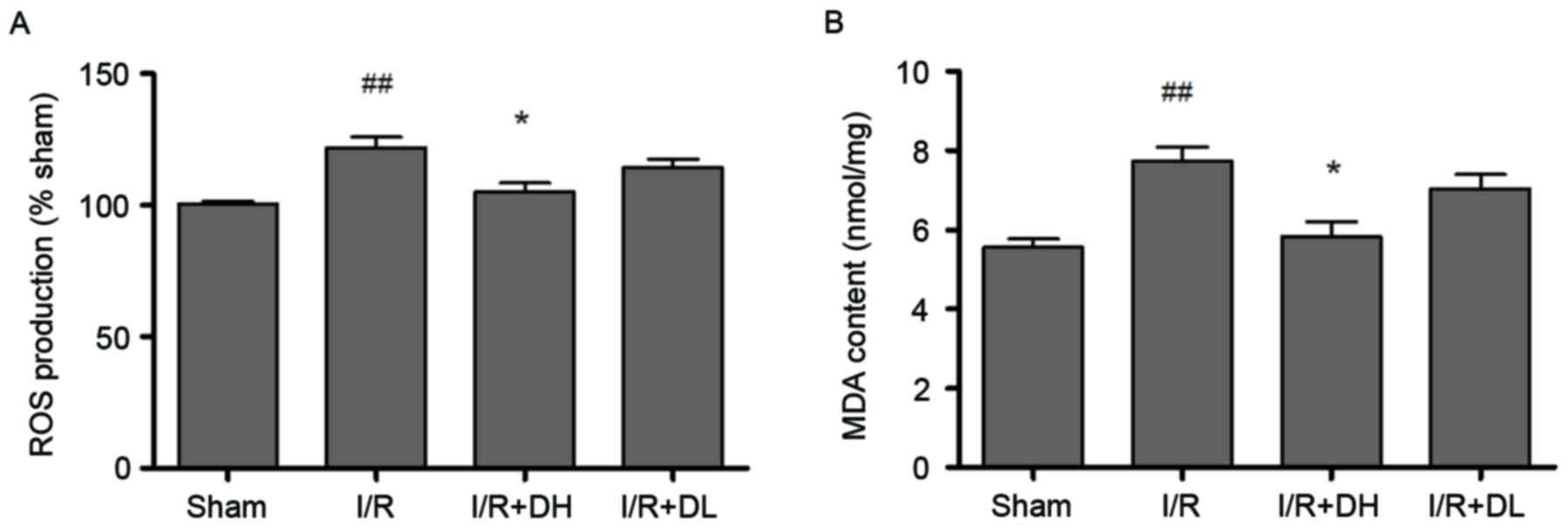 | Figure 2.Effect of DhHP-6 on oxidative stress
induced by 2 h of middle cerebral artery occlusion and 22 h of
reperfusion. (A) ROS levels in brain mitochondria are expressed as
the percentage of fluorescence intensity relative to that in the
Sham group. DhHP-6 attenuated ROS formation in a dose-dependent
manner. Compared with the I/R group, the ROS level was
significantly reduced in the I/R+DH group. (B) MDA content was
significantly reduced in the I/R+DH group, compared with that in
the I/R group. Data are shown as the mean ± standard deviation.
One-way analysis of variance and Tukey's post hoc test were
performed. ##P<0.01, vs. Sham group; *P<0.05, vs.
I/R group (n=10). I/R, ischemia/reperfusion; DhHP-6, deuterohemin
His peptide-6; DH, 1 mg/kg/day DhHP-6; DL, 0.1 mg/kg/day DhHP-6,
ROS, reactive oxygen species; MDA, malondialdehyde. |
As exhibited in Fig.
2B, I/R increased the formation of MDA. Compared with the level
in the I/R group, MDA was significantly reduced to 5.40±0.96
nmol/mg (P<0.05) in the I/R+DH group.
As demonstrated in Table I, CIR injury resulted in a
significant decrease in the activities of CAT, GSH-Px and SOD.
Treatment with DhHP-6 appeared to elevate the activities of these
antioxidant enzymes; however, no significant differences were
observed between the I/R group and the DhHP-6 treatment group
(P>0.05).
 | Table I.Effect of DhHP-6 on antioxidant
enzyme activities following 22 h of 12 reperfusion. |
Table I.
Effect of DhHP-6 on antioxidant
enzyme activities following 22 h of 12 reperfusion.
| Group | Dose (mg/kg) | CAT (U/mg
protein) | SOD (U/mg
protein) | GSH-Px (U/mg
protein) |
|---|
| Sham | – | 65.46±9.25 | 11.58±2.75 | 46.32±5.45 |
| I/R | – |
28.38±7.62a |
5.90±2.19b |
27.04±4.24b |
| I/R+DH | 1 |
35.69±6.81c |
8.58±2.23c |
35.96±5.08c |
| I/R+DL | 0.1 | 30.10±5.36 | 6.03±1.91 | 26.76±4.15 |
Together, these data indicated that DhHP-6 reduced
oxidative stress, however, the effect was not mediated by enhancing
the activities of endogenous CAT, GSH-Px or SOD antioxidant
enzymes.
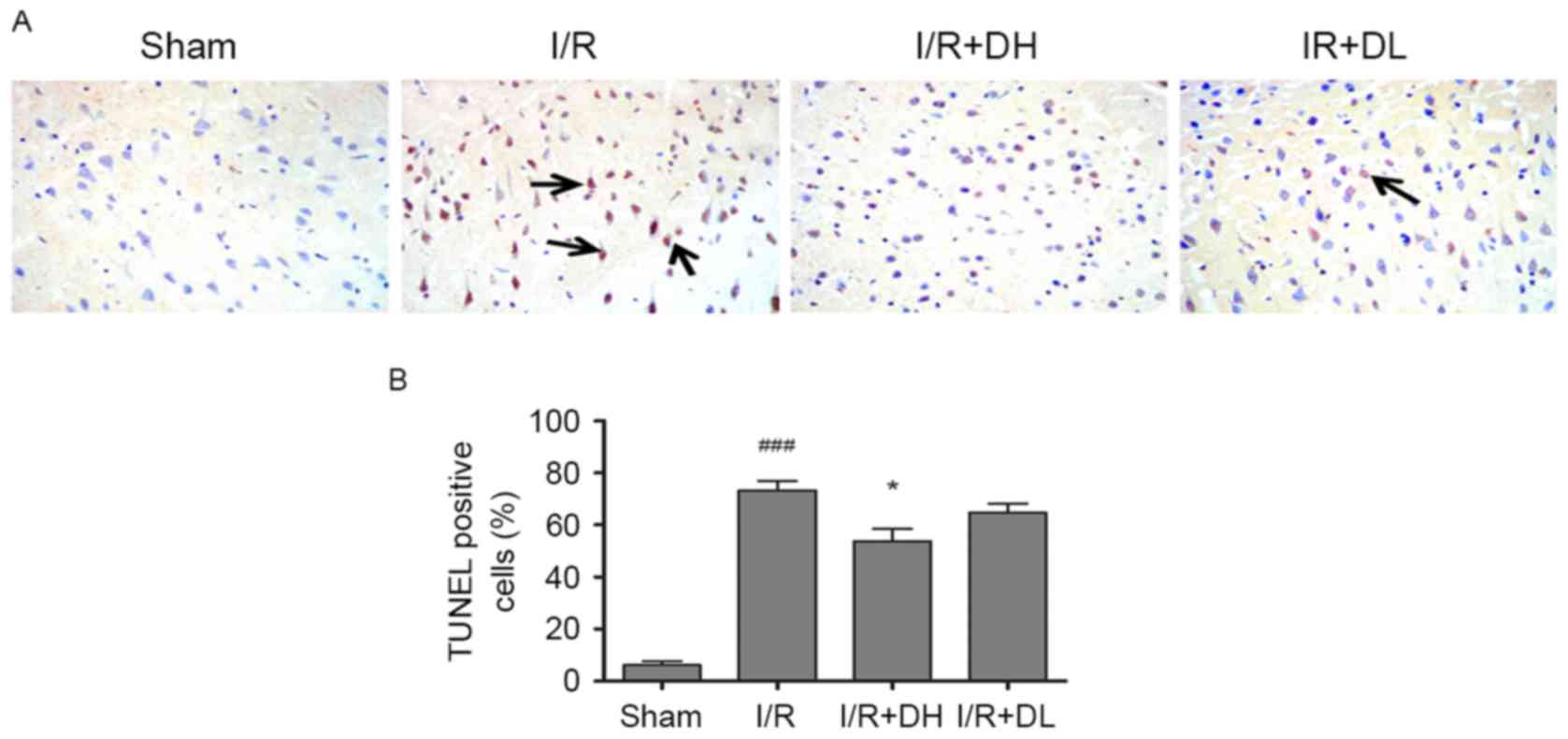
To examine the effects of DhHP-6 on apoptosis,
apoptotic cells were identified using TUNEL staining in the
parietal cortex 24 h post-MCAO. There were few TUNEL-positive
nuclei in the brain sections from animals in the sham group;
however, the number of TUNEL-positive cells was significantly
increased in the I/R group. DhHP-6 prevented the increase in
TUNEL-positive cells. As shown in Fig.
3A and B, the proportion of TUNEL-positive cells was
significantly reduced from 73.5±6.09% in the I/R group to
52.83±5.89% in the I/R+DH group (P<0.05).
DhHP-6 increases the Bcl-2/Bax
ratio
Bcl-2 is a key mitochondrial protein, which
contributes to cell survival. In the present study, the levels of
Bcl-2 were reduced in the I/R group, compared with those in the
sham group 24 h post-MCAO (Fig.
4). However, the important apoptotic protein, Bax, was
increased significantly in the I/R group. The ratio of Bcl-2/Bax
was decreased significantly following I/R (P<0.001), however,
there was an increase in the Bcl-2/Bax ratio when DhHP-6 was
administered at a dose of 1 mg/kg (P<0.05). These data indicated
that maintaining the balance between Bcl-2 and Bax was dependent on
DhHP-6.
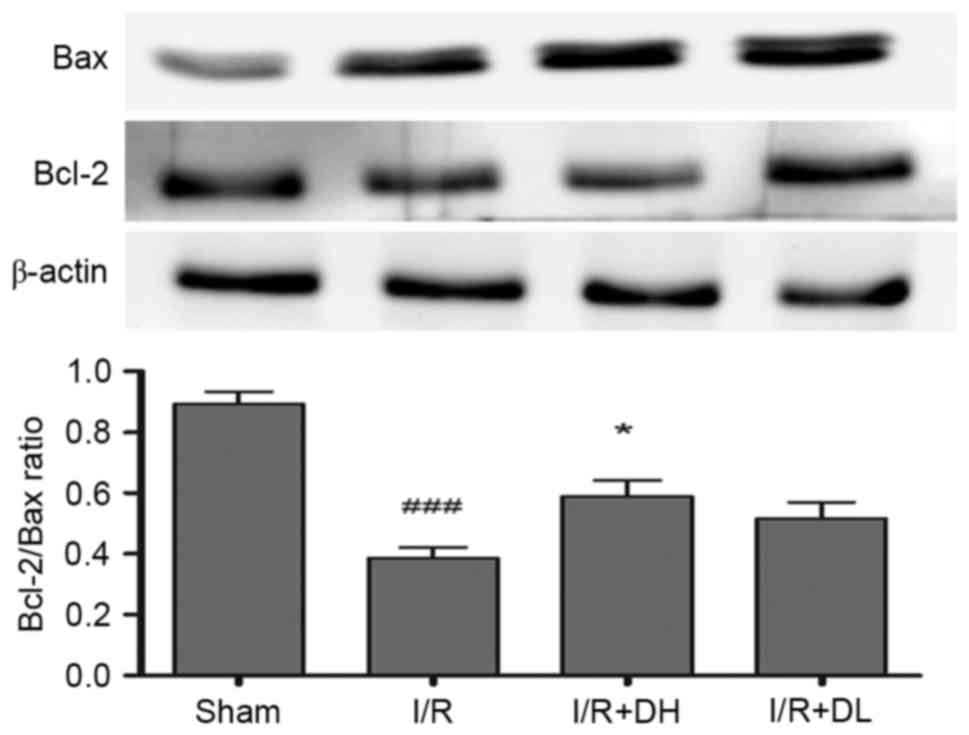 | Figure 4.Effects of DhHP-6 on expression levels
of Bcl-2 and Bax in the rat brain 22 h post-reperfusion. The
protein expression levels of Bcl-2 and Bax in subcellular fractions
were examined using western blot analysis. Scanning and
quantification of the intensity of protein bands were performed
using image analysis software. Representative bands from the Sham,
I/R, and DhHP-6 (I/R+DH and I/R+DL)-treated groups, and the
corresponding β-actin bands (loading control) are demonstrated.
Middle cerebral artery occlusion mediated a decrease in Bcl-2, and
an increase in Bax led to a significantly decreased Bcl-2/Bax ratio
in mitochondria, compared with the ratio in the Sham group.
Compared with the I/R group, the ratio of Bcl-2/Bax in mitochondria
was significantly higher in the I/R+DH group. Data are shown as the
mean ± standard deviation. One-way analysis of variance and Tukey's
post hoc test were used. ###P<0.001, vs. Sham group;
*P<0.05, vs. I/R group (n=3). DhHP-6, deuterohemin His
peptide-6; DH, 1 mg/kg/day DhHP-6; DL, 0.1 mg/kg/day DhHP-6; Bcl-2,
B-cell lymphoma 2; Bax, Bcl-2-associated X protein. |
DhHP-6 inhibits the activation of
caspase-3 and release of cytochrome c
As demonstrated in Fig.
5A, in the I/R group, an increased level of cytochrome c was
released into the cytoplasm from the mitochondria, and this effect
was significantly reduced by treatment with DhHP-6 (P<0.05). The
levels of cleaved caspase-3 were also investigated. DhHP-6 reduced
the expression of cleaved caspase-3. These effects were significant
in the I/R+DH group (P<0.05; Fig.
5B).
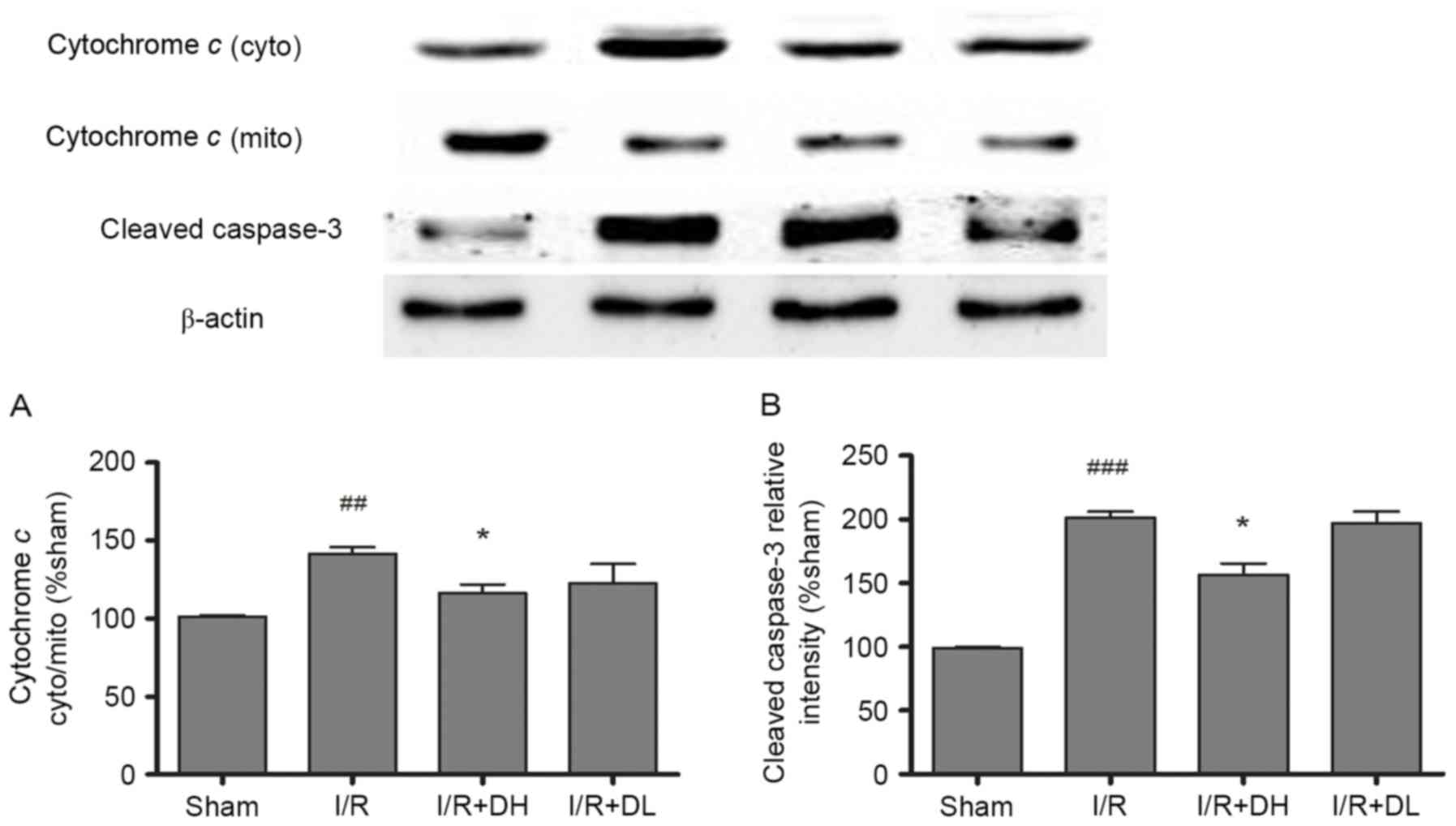 | Figure 5.Effects of DhHP-6 on the rate of
cytochrome c release and the activation of caspase-3 22 h
post-reperfusion. The contents of cytochrome c in the subcellular
fractions were examined using western blot analysis. (A) The ratio
of cytochrome c content in the cytosolic and mitochondrial
fractions was increased significantly in the I/R group, but the
ratio was decreased markedly in the I/R+DH group. (B) Additionally,
the expression level of cleaved caspase-3 was increased in the I/R
group at 22 h post-reperfusion, but significantly decreased in the
I/R+DH group, compared with the I/R group. Data are shown as the
mean ± standard deviation. One-way analysis of variance and Tukey's
post hoc test were performed. ###P<0.001 and
##P<0.01, vs. Sham group; *P<0.05, vs. I/R group
(n=3). DhHP-6, deuterohemin His peptide-6; DH, 1 mg/kg/day DhHP-6;
DL, 0.1 mg/kg/day DhHP-6; mito, mitochondria; cyto, cytosol; I/R,
ischemia/reperfusion. |
DhHP-6 increases the expression of
p-Akt/Akt
As demonstrated in Fig.
6, compared with the I/R group, treatment with DhHP-6 (1 mg/kg)
led to a significant increase in the expression of p-Akt/Akt
(P<0.05).
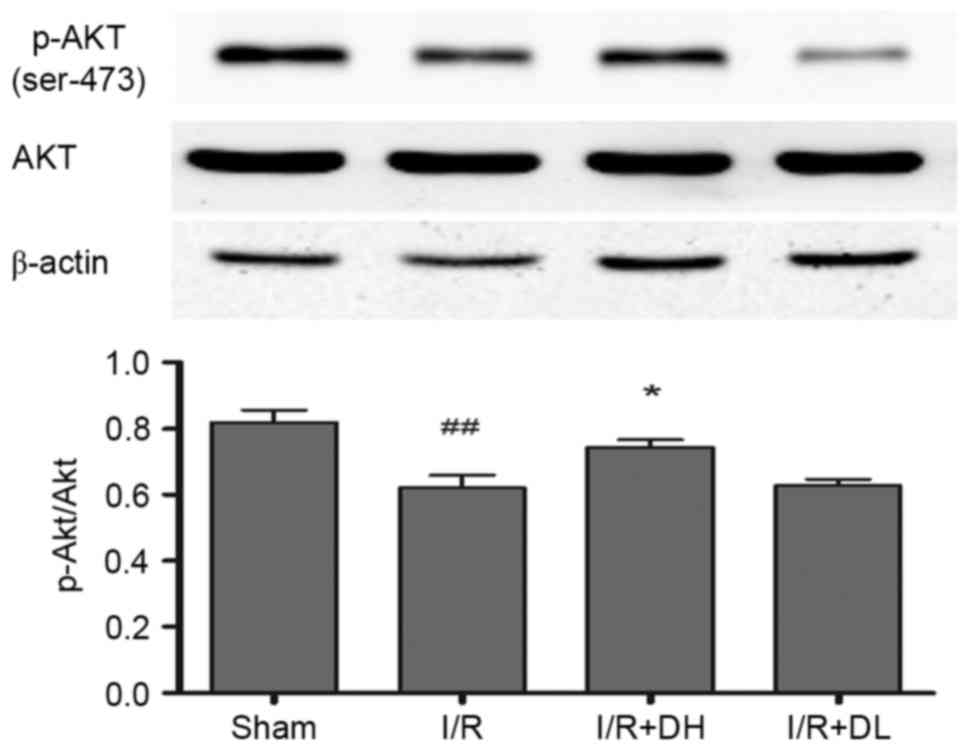 | Figure 6.Effect of DhHP-6 on p-Akt/Akt 22 h
post-reperfusion. To determine whether the activation of Akt
contributes to DhHP-6-dependent protection against apoptosis
induced by I/R injury, levels of p-Akt and Akt were examined using
western blot analysis. The histograms demonstrate the level of
p-Akt relative to that of Akt. Compared with the I/R group, the
p-Akt/Akt ratio was significantly increased in the I/R+DH group.
Data are shown as the mean ± standard deviation. One-way analysis
of variance and Tukey's post hoc test were performed.
##P<0.01, vs. Sham group; *P<0.05, vs. I/R group
(n=3). DhHP-6, deuterohemin His peptide-6; DH, 1 mg/kg/day DhHP-6;
DL, 0.1 mg/kg/day DhHP-6; p-, phosphorylated; I/R,
ischemia/reperfusion; Akt, AKT serine/threonine kinase. |
Discussion
In the present study, the function of DhHP-6 in
focal CIR injury was investigated in a rat model. The
administration of DhHP-6 at 1 mg/kg was sufficient to reduce
neurological deficits, brain edema and infarct volumes; however, a
dose of 0.1 mg/kg did not exert protective effects. These data
indicated that the beneficial effects of DhHP-6 occur in a
dose-dependent manner and also suggested that the effective dose
may be between 1 and 0.1 mg/kg. These results indicate a potential
novel application for DhHP-6 as a potent neuroprotectant in
cerebral ischemic disorder (19,20).
During I/R, ROS generation is enhanced and
antioxidant defenses in brain tissues are weakened; therefore, an
imbalance between oxidants and antioxidants occurs. The brain
isvulnerable to oxidative stress as it contains high levels of
unsaturated fatty acids, which can be oxidized leading to lipid
peroxidation (21,22). ROS are produced primarily by
mitochondria and are generated in I/R injury. DhHP-6 is a novel
free radical scavenger and is synthesized as a microperoxidase
mimetic. It was previously demonstrated that DhHP-6 produces
antioxidant effects in Caenorhabditis elegans (9). Another study showed that DhHP-6
treatment caused complete reduction in reactive oxygen (10). In the present study, the levels of
ROS in mitochondria were markedly increased following I/R injury,
and this increase was significantly attenuated by DhHP-6 treatment.
It was shown that oxidative injury to cell membrane lipids was
caused by free radicals, which can produce MDA. The level of MDA in
the I/R group was 2.18-fold higher, compared with that of the sham
group and was decreased significantly in the DhHP-6 group (Table I). The human body has different
mechanisms to decrease the impact of oxidative injury. The primary
defenses against oxidative injury are antioxidant enzymes,
including GSH-Px, SOD and CAT. However, the antioxidant effects of
DhHP-6 in the present study were not mediated through increasing
the activities of endogenous GSH-Px, SOD or CAT enzymes (Table I) (22,23).
The uncontrolled accumulation of ROS can disturb
mitochondrial function and lead to cell apoptosis. Mitochondria are
important in caspase-dependent and caspase-independent apoptotic
processes. Mitochondria can induce the opening of the mitochondrial
permeability transition pore and thereby lead to the release of
cytochrome c. The Bcl-2 family of proteins, which are
expressed on outer mitochondrial membranes, are also critical in
mitochondria-mediated apoptosis (24). In the present study, Bax was
identified in cytosolic brain tissue extracts. When Bax is
activated, it translocates to mitochondria, where it then
stimulates mitochondria to release cytochrome c. Bcl-2 can
inhibit the activation of Bax (25,26),
however, the present study showed that significantly higher levels
of Bax were expressed in the mitochondria of rat brains subjected
to MCAO; this effect was attenuated by treatment with DhHP-6.
Therefore, DhHP-6 inhibited the release of cytochrome c
andinhibited apoptosis by ameliorating the CIR-induced degradation
of the Bcl-2/Bax ratio, and by retaining the normal balance between
Bcl-2 and Bax. In the present study, it was also observed that
TUNEL-positive cells increased in the cortex 10-fold following 22 h
of brain reperfusion. This increase was significantly reduced in
the I/R+DH group. Therefore, these results indicated that DhHP-6
inhibited the mitochondria-initiated apoptotic pathway and thereby
improved neuronal survival (27,28).
The activation of phosphoinositide 3-kinase (PI3K)
and its downstream factor, Akt has been demonstrated to prevent
apoptosis and promote cell survival. Increasing evidence has
indicated that ROS-induced apoptosis correlates with the PI3K/Akt
pathway in SH-SY5Y cells (29). In
focal CIR injury models, p-Akt is usually dephosphorylated at
Ser473. It has been demonstrated that, following activation, Akt
can promote cell survival and subsequently inactivate
apoptosis-inducing factors, including GSK3β (30). A previous study also demonstrated
that GSK3β exerts its pro-apoptotic effects by regulating Bax
localization to mitochondria (31,32).
The results of the present study indicated thatDhHP-6 markedly
enhanced the phosphorylation of Akt in rats following 2 h of MCAO
and 22 h of reperfusion, compared with the results observed for the
I/R group.
Taken together, the findings of the present study
demonstrated that DhHP-6 reduced the levels of brain edema, infarct
volumes and neurological deficits in CIR. The protective mechanisms
of DhHP-6 in CIR injury included scavenging of radicals and
inhibiting the mitochondrial apoptotic-signaling pathway, which may
involve activation of the PI3K/Akt signaling pathway. Therefore,
the beneficial effects of DhHP-6 on CIR investigated in the present
study indicated that DhHP-6 may be a promising therapeutic
candidate for protecting against cell death resulting from ischemic
damage.
Acknowledgements
The authors would like to thank Dr Li Wei of the
Life Science College at Jilin University for the provision of
DhHP-6.
References
|
1
|
Abramov AY, Scorziello A and Duchen MR:
Three distinct mechanisms generate oxygen free radicals in neurons
and contribute to cell death during anoxia and reoxygenation. J
Neurosci. 27:1129–1138. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Marlangue C Charriaut, Remolleau S,
Zouaoui D Aggoun and Ben-Ari Y: Apoptosis and programmed cell
death: A role in cerebral ischemia. Biomed Pharmacother.
52:264–269. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Chan PH: Reactive oxygen radicals in
signaling and damage in the ischemic brain. J Cereb Blood Flow
Metab. 21:2–14. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Allen CL and Bayraktutan U: Oxidative
stress and its role in the pathogenesis of ischaemic stroke. Int J
Stroke. 4:461–470. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Egleton RD and Davis TP: Bioavailability
and transport of peptides and peptide drugs into the brain.
Peptides. 18:1431–1439. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Navaratna D, Guo S, Arai K and Lo EH:
Mechanisms and targets for angiogenic therapy after stroke. Cell
Adh Migr. 3:216–223. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Dong QG, Zhang Y, Wang MS, Feng J, Zhang
HH, Wu YG, Gu TJ, Yu XH, Jiang CL, Chen Y, et al: Improvement of
enzymatic stability and intestinal permeability of
deuterohemin-peptide conjugates by specific multi-site
N-methylation. Amino Acids. 43:2431–2441. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang H, Sun Y, Guo W, Fang C, Fawcett JP,
Li W, Gao Y, Yang Y and Gu J: Determination of a
deuterohemin-peptide conjugate in rat plasma by liquid
chromatography-tandem mass spectrometry and application to a
preclinical pharmacokinetic study. J Pharm Biomed Anal. 98:401–406.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Huang L, Li P, Wang G, Guan S, Sun X and
Wang L: DhHP-6 extends lifespan of Caenorhabditis elegans by
enhancing nuclear translocation and transcriptional activity of
DAF-16. Free Radic Res. 47:316–324. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Guan S, Li P, Luo J, Li Y, Huang L, Wang
G, Zhu L, Fan H, Li W and Wang L: A deuterohemin peptide extends
lifespan and increases stress resistance in Caenorhabditis elegans.
Free Radic Res. 44:813–820. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Xue H, Ji Y, Wei S, Yu Y, Yan X, Liu S,
Zhang M, Yao F, Lan X and Chen L: HGSD attenuates neuronal
apoptosis through enhancing neuronal autophagy in the brain of
diabetic mice: The role of AMP-activated protein kinase. Life Sci.
153:23–34. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Longa EZ, Weinstein PR, Carlson S and
Cummins R: Reversible middle cerebral artery occlusion without
craniectomy in rats. Stroke. 20:84–91. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yenari MA, Xu L, Tang XN, Qiao Y and
Giffard RG: Microglia potentiate damage to blood-brain barrier
constituents: Improvement by minocycline in vivo and in vitro.
Stroke. 37:1087–1093. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wang N, Zhang Y, Wu L, Wang Y, Cao Y, He
L, Li X and Zhao J: Puerarin protected the brain from cerebral
ischemia injury via astrocyte apoptosis inhibition.
Neuropharmacology. 79:282–289. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang D, Yuan X, Liu T, Liu L, Hu Y, Wang Z
and Zheng Q: Neuroprotective activity of lavender oil on transient
focal cerebral ischemia in mice. Molecules. 17:9803–9817. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kwon SH, Hong SI, Kim JA, Jung YH, Kim SY,
Kim HC, Lee SY and Jang CG: The neuroprotective effects of Lonicera
japonica THUNB. against hydrogen peroxide-induced apoptosis via
phosphorylation of MAPKs and PI3K/Akt in SH-SY5Y cells. Food Chem
Toxicol. 49:1011–1019. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yu SM and Kim SJ: Thymoquinone-induced
reactive oxygen species causes apoptosis of chondrocytes via
PI3K/Akt and p38kinase pathway. Exp Biol Med (Maywood).
238:811–820. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kim DW, Jeong HJ, Kang HW, Shin MJ, Sohn
EJ, Kim MJ, Ahn EH, An JJ, Jang SH, Yoo KY, et al: Transduced human
PEP-1-catalase fusion protein attenuates ischemic neuronal damage.
Free Radic Biol Med. 47:941–952. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Pan R, Rong Z, She Y, Cao Y, Chang LW and
Lee WH: Sodium pyruvate reduced hypoxic-ischemic injury to neonatal
rat brain. Pediatr Res. 72:479–489. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang F, Wang S, Signore AP and Chen J:
Neuroprotective effects of leptin against ischemic injury induced
by oxygen-glucose deprivation and transient cerebral ischemia.
Stroke. 38:2329–2336. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chan PH: Role of oxidants in ischemic
brain damage. Stroke. 27:1124–1129. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Keller JN, Kindy MS, Holtsberg FW, St
Clair DK, Yen HC, Germeyer A, Steiner SM, Bruce-Keller AJ, Hutchins
JB and Mattson MP: Mitochondrial manganese superoxide dismutase
prevents neural apoptosis and reduces ischemic brain injury:
Suppression of peroxynitrite production, lipid peroxidation, and
mitochondrial dysfunction. J Neurosci. 18:687–697. 1998.PubMed/NCBI
|
|
23
|
Ryter SW, Kim HP, Hoetzel A, Park JW,
Nakahira K, Wang X and Choi AM: Mechanisms of cell death
inoxidative stress. Antioxid Redox Signal. 9:49–89. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Namura S, Zhu J, Fink K, Endres M,
Srinivasan A, Tomaselli KJ, Yuan J and Moskowitz MA: Activation and
cleavage of caspase-3 in apoptosis induced by experimental cerebral
ischemia. J NeuroSci. 18:3659–3668. 1998.PubMed/NCBI
|
|
25
|
Green DR and Reed JC: Mitochondria and
apoptosis. Science. 281:1309–1312. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Cardone MH, Roy N, Stennicke HR, Salvesen
GS, Franke TF, Stanbridge E, Frisch S and Reed JC: Regulation of
cell death protease caspase-9 by phosphorylation. Science.
282:1318–1321. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Xing Y, Zhang X, Zhao K, Cui L, Wang L,
Dong L, Li Y, Liu Z, Wang C, Zhang X, et al: Beneficial effects of
sulindac in focal cerebral ischemia: A positive role in
Wnt/β-catenin pathway. Brain Res. 1482:71–80. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yoshida H, Kong YY, Yoshida R, Elia AJ,
Hakem A, Hakem R, Penninger JM and Mak TW: Apaf1 is required for
mitochondrial pathways of apoptosis and brain development. Cell.
94:739–750. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
González-Sarrías A, Núñez-Sánchez MÁ,
Tomás-Barberán FA and Espín JC: Neuroprotective effects of
bioavailable polyphenol-derived metabolites against oxidative
stress-induced cytotoxicity in human neuroblastoma SH-SY5Y Cells. J
Agric Food Chem. 65:752–758. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Li F, Omori N, Jin G, Wang SJ, Sato K,
Nagano I, Shoji M and Abe K: Cooperative expression of survival
p-ERK and p-Akt signals in rat brain neurons after transient MCAO.
Brain Res. 962:21–26. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Friguls B, Justicia C, Pallàs M and Planas
AM: Focal cerebischemia causes two temporal waves of Akt
activation. Neuroreport. 12:3381–3384. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Sun B, Feng M, Tian X, Lu X, Zhang Y, Ke
X, Huang S, Cao J and Ding X: DL-3-n-Butylphthalide protects rat
bone marrow stem cells against hydrogen peroxide-induced cell death
through antioxidation and activation of PI3K-Akt pathway. Neurosci
Lett. 516:247–252. 2012. View Article : Google Scholar : PubMed/NCBI
|