Introduction
Innovations are continuously being made in bone
tissue engineering, from implanted material to bone substitutes,
and subsequently autologous bone grafting (1–4),
which avoids the issue of graft rejection; however, it has not
solved the problem of limited endogenous supply, prolonged surgical
duration and harvest complications (5–7).
Cell sources, such as bone marrow mesenchymal stem cells and
adipose-derived stem cells have generated significant interest for
their tissue engineering potential (8,9).
Perivascular stem cells (PSCs) are a homogeneous
mesenchymal stem cell (MSC) population purified by fluorescence
activated cell sorting (FACS). PSCs are cell populations that are
comprised of pericytes [cluster of differentiation
(CD)45−CD34−CD146+) (10) and adventitial cells
(CD45−CD34+CD146−), which are
selected by fluorescence-activated cell sorting (FACS) (11). The histological localization is in
blood vessels, and the two cell types have been reported to possess
characteristics of mesenchymal stem cells (MSCs) (12–14).
PSCs are the natural ancestors of MSCs, and are responsible for
homeostasis and repair in vivo. Previous studies have
indicated that purified PSCs have increased chondrogenic potential
when compared with conventional MSCs derived in culture (10,15).
Lipoaspirate-derived human PSCs have demonstrated myogenic and
angiogenic potential, and have exhibited a strong therapeutic
effect in the treatment of rotator cuff tears (16). PSC xenografting demonstrated the
osteogenic capability of this cell type in ectopic and orthotopic
bone regeneration models (17).
Additionally, a study indicated that osteoinductive growth factor
Nel-like molecule 1 induced human (h) PSC osteogenesis (18). hPSCs may present as a promising
cell source for future efforts in skeletal regenerative medicine.
hPSCs provide a stem cell-based therapeutic modality that is
readily approved by the United States Food and Drug Administration,
with potentially increased safety, purity, identity, potency and
efficacy (19).
PSCs are abundant in human white adipose tissue
(20); thus, adipose tissue is an
ideal MSC source, as it is dispensable and accessible with minimal
morbidity. Therefore, in recent years, numerous studies describing
the identification, isolation and characterization of PSCs from the
adipose tissues of humans and animals have been performed (9,11,20–22).
However, at present, to the best of our knowledge, there are no
studies on the purification and identification of PSCs in minipigs
(mps). Therefore, it is hypothesized that there is a phenotype
similar to that of the human body in mpPSCs. The findings may
benefit cosmetic dentistry and facial cosmetic surgery fields, as
the orthodontic tooth or craniofacial bone tissue repair are
closely associated with the development of bone. The aim of the
current study was to identify, isolate and characterize PSCs from
mp adipose tissue samples (n=9). Purification of mpPSCs by FACS
(which is used in human PSC purification), and investigation of
mpPSC osteogenic and adipogenic potential (using Alizarin Red S
staining and Oil Red O staining in vitro, respectively) was
performed, followed by observing the cell morphometry after cell
isolation and culture. In addition, hematoxylin and eosin (H&E)
staining was conducted to identify the fat tissue structure and
vascular distribution. Furthermore, osteogenic induction
differentiation regulating genes, osteocalcin (OCN) and collagen,
type I, α1 (COL1A1), and the adipogenesis-associated gene,
peroxisome proliferator-activated receptor-γ (PPARG) (23) were evaluated by reverse
transcription-quantitative polymerase chain reaction (RT-qPCR).
Materials and methods
PSC identification, isolation and
culture
hPSCs were provided by liposuction patients
(anonymous) following surgery at the Beijing Huangsi Cosmetic
Plastic Surgery Hospital (Beijing, China). The mp adipose tissue
samples were obtained from 9 male, health-certified Guizhou mps
(age, 10 months; mean weight, 37 kg) from the Chinese Academy of
Agricultural Sciences (Beijing, China). All protocols were approved
by the Animal Use and Care Committee of Peking University (Beijing,
China; permit no. LA2014216). Unless otherwise stated, the
subcutaneous fat from the posterior neck of the mps was used.
In order to determine the minipig stromal vascular
fraction (SVF) yield, H&E staining was performed on tissue
sections (~1×1×0.5 cm3). The mp adipose tissue samples
were snap frozen and cryosections were obtained at −30°C, then the
tissue samples were stained with H&E according to the following
protocol: Tissue samples were fixed using formalin for 24 h,
regularly dehydrated and routinely embedded. The sections were
deparaffinized and rehydrated with xylene, 100% alcohol, 95%
alcohol, 75% alcohol. The sections were stained in Harris
hematoxylin solution at room temperature for 10 min, differentiated
with 1% acid alcohol for 3–5 sec until the nucleus was colored
blue, then counterstained in eosin Y solution for 30 sec,
dehydrated with 95 then 100% alcohol, rehydrated with xylene and
mounted using neutral balsam. Images were captured using an Olympus
BX60 microscope (Olympus Corp., Tokyo, Hapan) using a ×40 objective
oil immersion lens. Subsequently, the PSCs were analyzed and
isolated from the minipig adipose tissues via flow cytometry.
Isolation and culture were performed as previously described for
hPSC isolation (14,20).
Lipoaspirate specimens were enzymatically digested
to obtain the SVF. The SVF was incubated at 37°C for 50 min with
antibodies recognizing human PSC antibodies and mp PSC antibodies.
Mp PSC antibodies were as follows: CD146-fluorescein isothiocyanate
(FITC; 1:100; cat. no. MCA2141F; Bio-Rad Laboratories, Inc.,
Hercules, CA, USA), CD45-phycoerythrin (PE; 1:100; cat. no. SM563R;
OriGene Technologies, Inc., Rockville, MD, USA) and CD34 (1:100;
cat. no. ab81289; Abcam, Cambridge, MA, USA). Human PSC antibodies
were as follows: CD146-PE (1:100; cat. no. 55305), CD45-FITC
(1:100; cat. no. 555482) and CD34-APC (1:100; cat. no. 555824) (BD,
Hercules, CA, USA). Cells were sorted using a special order
five-laser BD FACS Aria III high-speed cell sorter (BD Biosciences,
Franklin Lakes, NJ, USA). Non-viable and hematopoietic cells were
excluded based on staining for DAPI and CD45-PE, respectively.
CD34+CD146− adventitial cells and
CD146+CD34− pericytes were collected as PSCs.
The two groups of cells were not clustered clearly, but the
fluorescence minus one flow test was used as a control. Samples
with detectable pericytes and adventitial populations were
examined. Cells were cultured in Gibco Dulbecco's modified Eagle's
medium (DMEM; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
with Gibco 10% fetal bovine serum (FBS; Thermo Fisher Scientific,
Inc.) and 1% penicillin/streptomycin. The medium was changed
routinely every 3 days. Nine minipig adipose tissue samples were
examined and 6 hPSC samples were examined as an interspecies
comparison. Prior to analysis, CD34+CD146−
adventitial cells and CD146+CD34− pericytes
were purified using FACS and cultured until passages four to six in
a monolayer culture.
In vitro osteogenic differentiation
assays
For osteogenic differentiation, cells were seeded
into 24-well plates at a density of 5×104 cells/well.
Following attachment, cells were treated with osteogenic
differentiation medium (ODM) consisting of DMEM, 10% FBS, 10 mM
β-glycerophosphate and 50 µM ascorbic acid. In select studies, ODM
was supplemented with 1 µM dexamethasone (Sigma-Aldrich; Merck
KGaA, Darmstadt; cat. no. D4902-25MG). The medium was refreshed
every three days. After 14 days, osteogenic differentiation was
assessed using Alizarin Red S staining according to the
manufacturer's protocol (cat. no. 0223; ScienCell Research
Laboratories, Inc., San Diego, CA, USA). Briefly, cells were seeded
at a density of 5×104 cells/well in 24-well plates.
Cells were subsequently fixed for 15 min at 5°C in 4%
paraformaldehyde prior to staining with 2% Alizarin Red S for 10
min at room temperature. Cetylpyridinium chloride (10%;
Sigma-Aldrich; Merck KGaA) was subsequently applied for 15 min at
room temperature for quantification. Cells were examined under an
inverted fluorescence microscope (magnification, ×200) and
quantification was performed with a microplate spectrophotometer at
562 nm.
In vitro adipogenic differentiation
assays
After 14 days of differentiation, the cells were
washed gently with phosphate-buffered saline (PBS) and fixed with
4% paraformaldehyde (pH 7.4) for 10 min. The fixed liquid was
discarded. Lipid droplets were stained with 0.5% Oil Red O in 60%
isopropanol at room temperature for 15 min. The Oil Red O was
removed and the plates were rinsed with water and dried at 37°C.
Subsequently, 60% isopropanol was used to extract the dye from the
cells for 10–20 min, and the cells were washed with PBS twice, at
last immersed in PBS and observed under an inverted light
microscope (magnification, ×200).
RT-qPCR
Total RNAs were extracted from the lung tissue
samples of each group. cDNA was synthesized using a
TaqMan™ MicroRNA reverse transcription kit (cat. no.
4366596; Thermo Fisher Scientific, Inc.) and cDNA served as a
template for RT-qPCR (Applied Biosystems 7500 Real-Time PCR
Instrument; Thermo Fisher Scientific, Inc.). Reactions were
conducted in 96-well optical plates at 95°C for 10 min, followed by
40 cycles at 95°C for 15 sec and 60°C for 1 min. The quantification
cycle (Cq) data were determined using default threshold settings
(24). The Cc was defined as the
fractional cycle number at which the fluorescence passes the fixed
threshold. Reactions were run in duplicate to triplicate per RNA
isolate. The relative expression of OCN, COL1A1 and PPARG in each
group of cells were calculated by RT-qPCR and GAPDH served as an
internal standard. Primer sequences were as follows: GAPDH forward,
CAATGACCCCTTCATTGACC and reverse, GAAGATGGTGATGGCCTTTC; COL1A1
forward, GGTTTCAGTGGTTTGGATGG and reverse, TCCATTTTCACCAGGGCTAC;
OCN forward, TCACACTGCTTGCCCTACTG and reverse, CTGCACCTTTGCCAGAATC;
and PPARG forward, GCCAAGGATTCATGACAAGG and reverse,
TTGGGCTCCATAAAGTCACC.
Statistical analysis
All results were expressed as means ± standard error
and analyzed using SPSS version 19 (IBM Corp., Armonk, NY, USA).
The difference was detected by Student t-test, or one-way analysis
of variance followed by Fisher's Least Significant Difference
post-hoc test for different group data analysis and P<0.05 was
considered to indicate a statistically significant difference.
Results
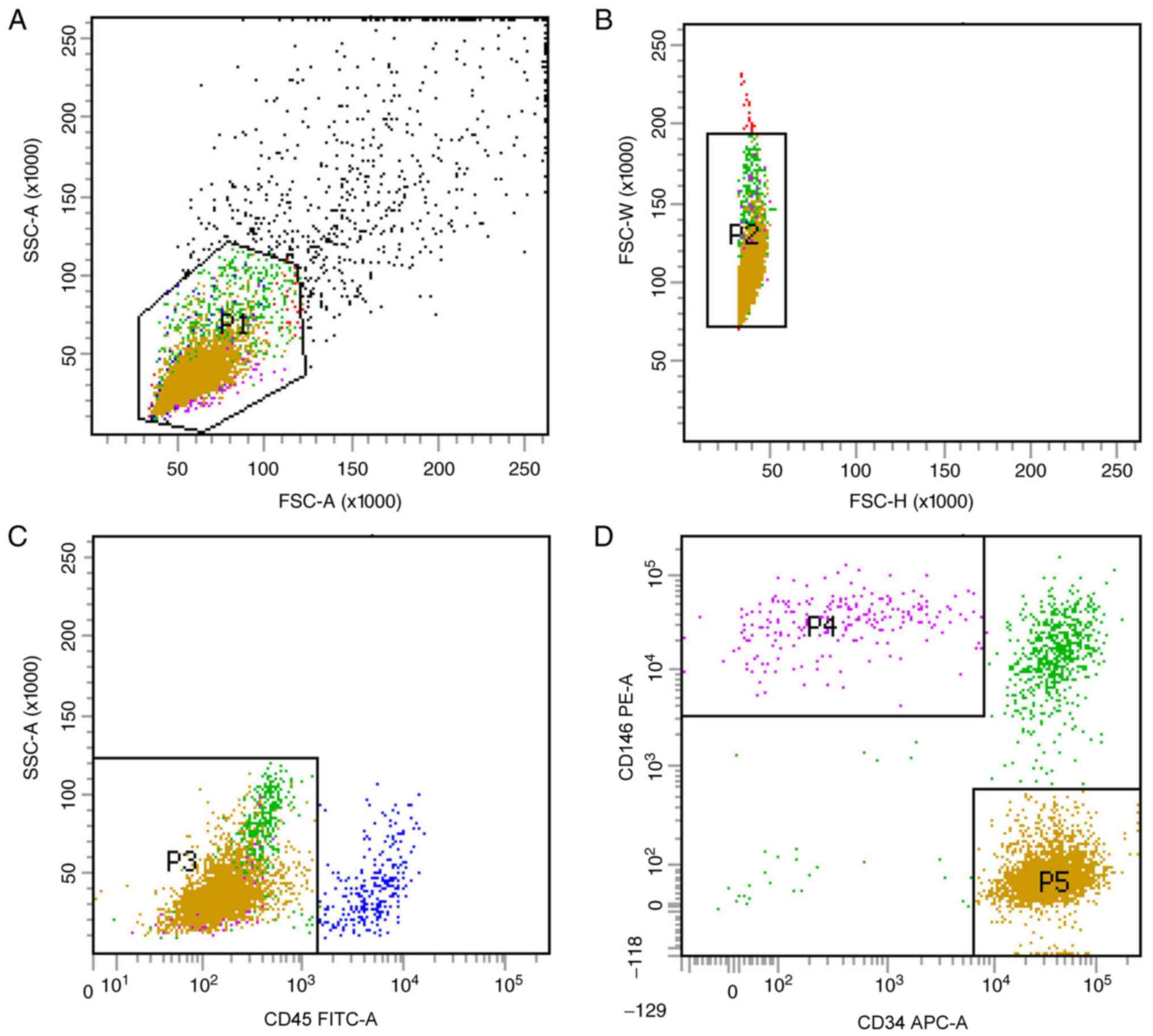
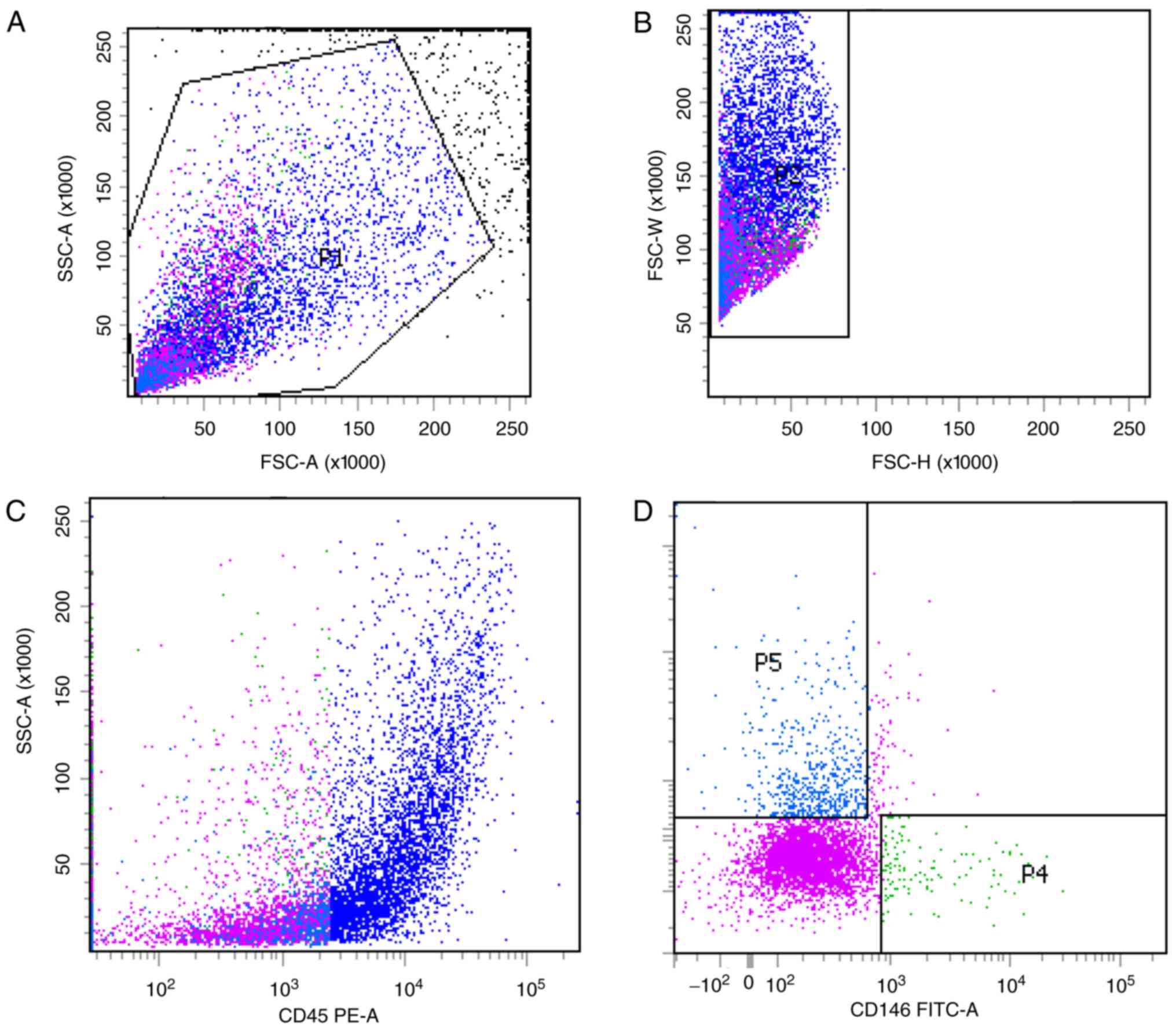
mpPSCs and hPSCs are obtained by
identical processes
Previous studies have optimized the procedure for
obtaining hPSCs (8). In the
current study, two subgroups of hPSCs, the
CD45−CD34−CD146+ phenotype of
perithelial cells (P4) and the
CD45−CD34+CD146− phenotype of
adventitial cells (P5), were isolated from human liposuction fat by
FACS (Fig. 1). Subsequently, this
protocol was applied to mp adipose tissue samples. Single cells
were gated and histograms of each antibody were compared with the
relevant isotype control (Fig. 2).
Cells were sorted into
CD45−CD34−CD146+ (P4) and
CD45−CD34+CD146− cell populations
(P5). The two groups of cells were not clustered clearly, but the
fluorescence minus one flow test was used as a loop gate and the
results were reliable.
CD45−CD34−CD146+ cell populations
and CD45−CD34+CD146− cell
populations accounted for 1.4% (P4/P1) and 7.2% (P5/P1) of SVF
cells, respectively, with a total of 8.6%, which was lower than
that previously reported in the literature (9). In the current study,
immunohistochemistry was not performed; therefore, the hPSC
phenotypes of the cells corresponding to that of the mps were used
as the supporting evidence to consider these two cell populations
as ‘PSCs’ (mpPSCs).
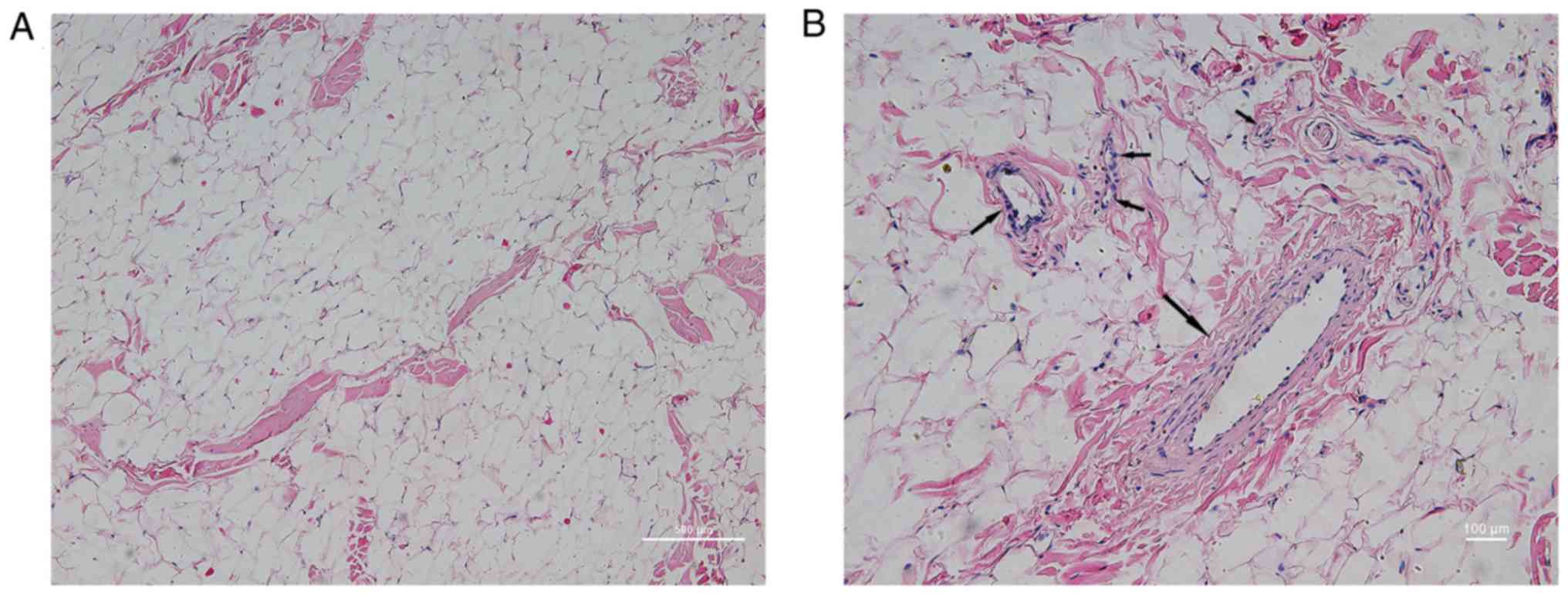
SVF are abundant in mp white adipose
tissue
In order to determine the SVF content in mp adipose
tissue, H&E staining was performed in subcutaneous adipose
tissue sampled from the posterior neck of the mps. As shown in
Fig. 3, it was observed at low
magnification that adipose tissue was predominantly composed of a
large number of vacuolar fatty cells and a small quantity of
connective tissue. Observation of connective tissue at high
magnification demonstrated abundant vascular-like structures, with
diameters from several microns to hundreds of microns. The vascular
wall exhibited three layers of arteriovenous-like structures
(endometrium, intermediate muscle layer and outer membrane) and
capillary-like monolayer structures (predominantly including the
endothelial layer and the surrounding cells). This region of the
structure was termed SVF, and according to the human PSC theory,
the fat derived mpPSCs are separate from this region.
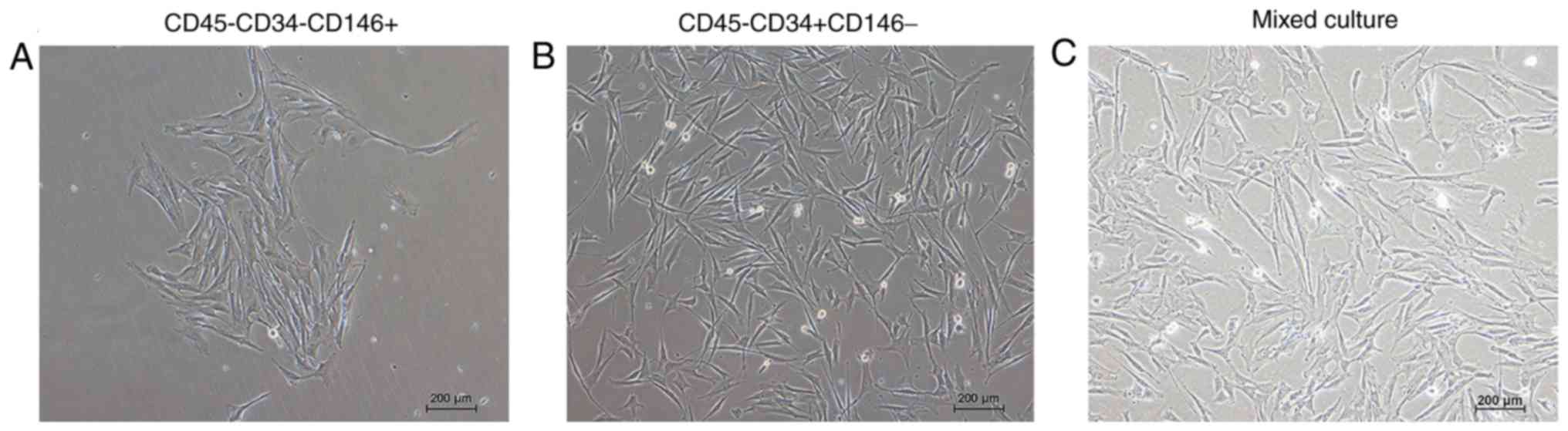
Cell morphology of the mpPSCs
The morphology of the cells adhered to the wall are
presented in Fig. 4. Following
cell sorting, the cells were separated and then mixed in culture.
The result indicated that the morphology of the two groups of cells
that had adhered to the wall was marginally different. The cell
morphology of the CD45−CD34−CD146+
group was longer and narrower, and the cell morphology of the
CD45−CD34+CD146− group was
polygonal.
mpPSCs demonstrated MSC
characteristics
mpPSCs were evaluated based on their adipogenic and
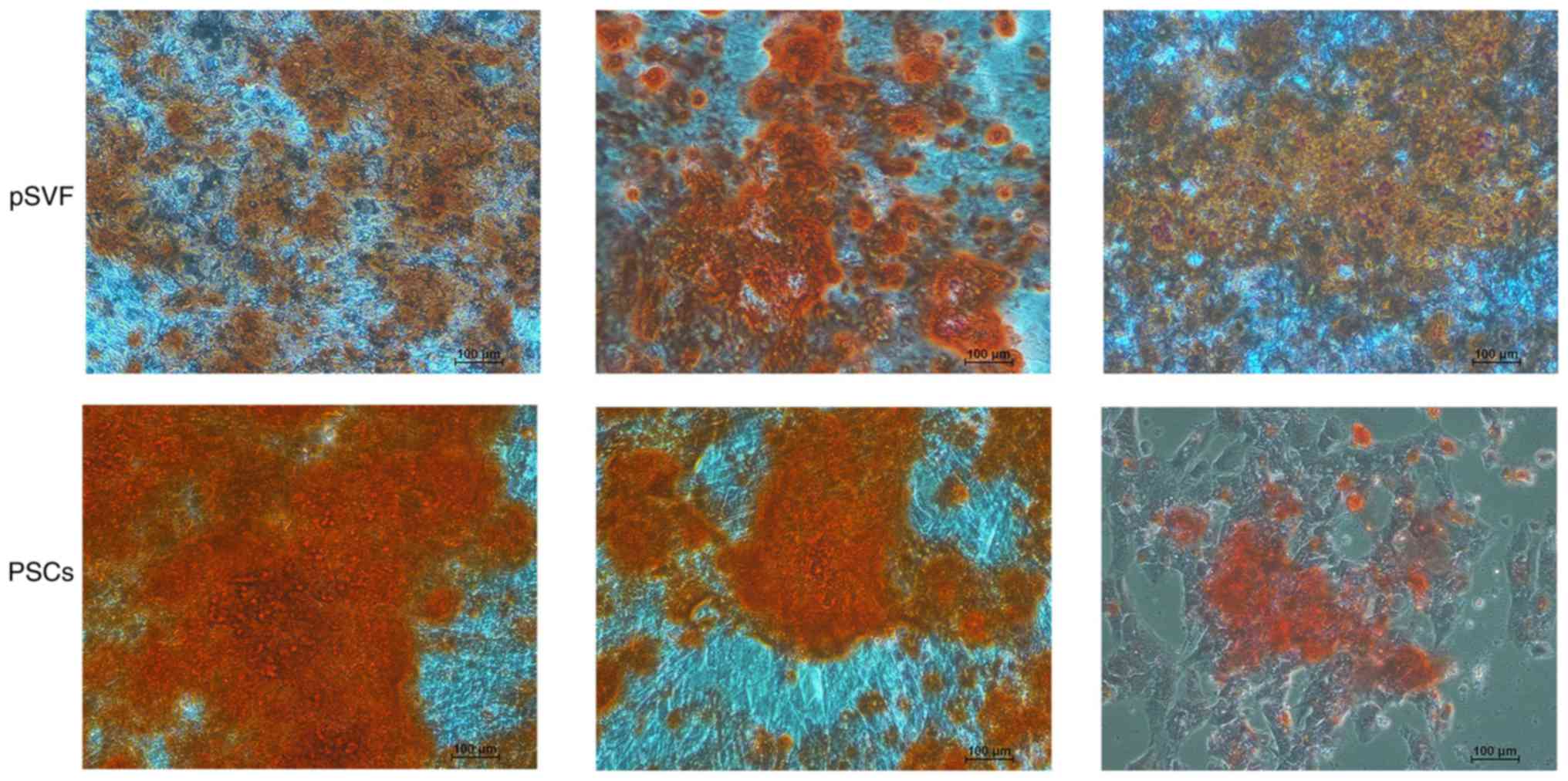
osteoplastic differentiation ability. As presented in Fig. 5, typical staining of calcium
nodules was demonstrated by Alizarin Red in the two groups. There
was a wider area of deeper staining in the group of PSCs,
indicating that mpPSCs had a stronger and more prevalent ability in
calcium nodule formation than pSVFs, which indicated that the
mpPSCs were able to undergo osteoplastic differentiation.
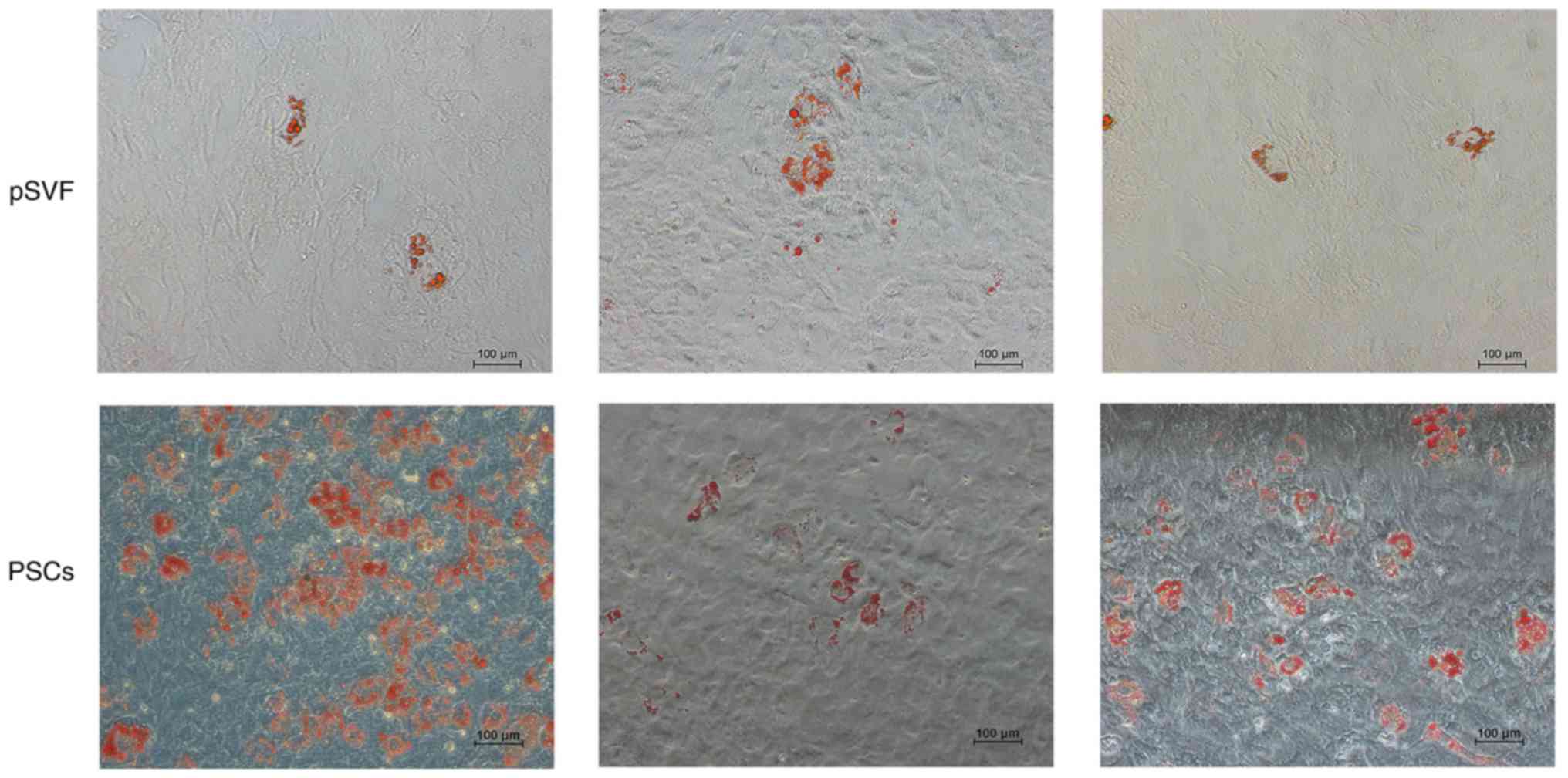
The ability of adipogenic differentiation was
detected by the results of Oil Red O staining (Fig. 6). The results demonstrated the
presence of lipid droplets, with grape-like clusters observed in in
the two groups. The lipid droplet density of the mpPSCs group was
significantly higher than that of the pSVF group, indicating that
the lipid composition ability of the mpPSCs was stronger.
Osteogenic induction differentiation
and adipogenesis-associated gene expression levels were higher in
the mpPSCs group than in the pSVF group
The results of the RT-qPCR (Fig. 7) demonstrate that compared with the
pSVF group, the mRNA expression levels of osteogenic induction
differentiation regulating genes, OCN and COL1A1, and
adipogenesis-associated gene, PPARG in the mpPSCs group were
significantly higher (P<0.05).
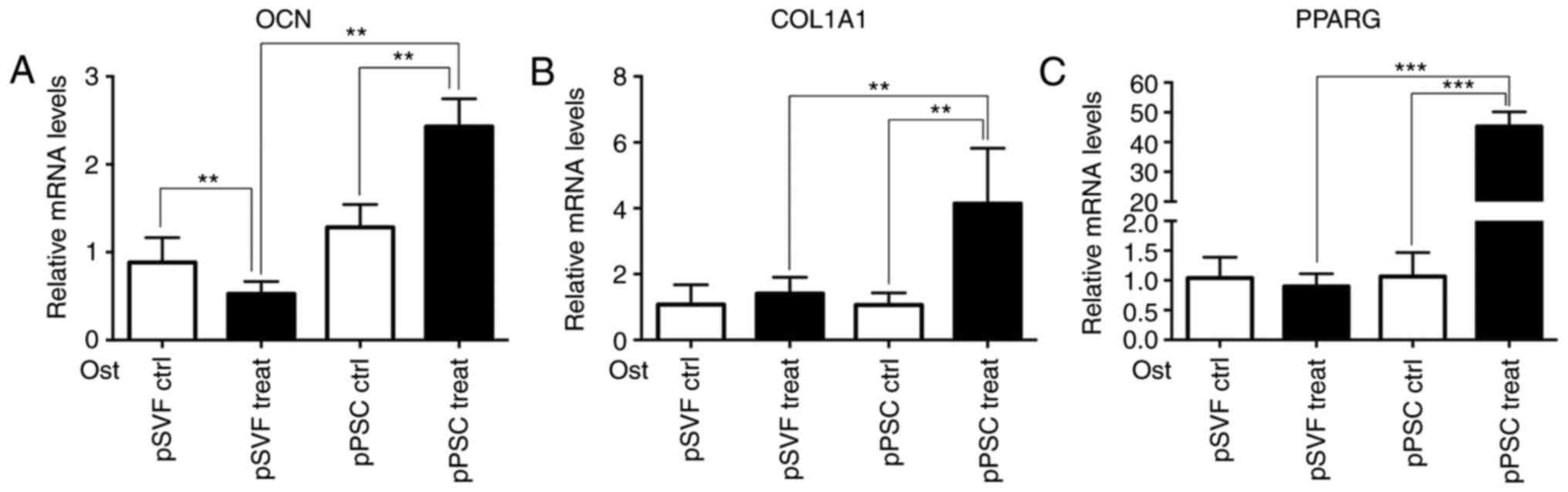 | Figure 7.Adipogenesis-associated gene
expression measured by the reverse transcription-quantitative
polymerase chain reaction. The mRNA expression levels of (A) OCN,
(B) COL1A1 and (C) PPARG in the pPSC group were significantly
higher. **P<0.01, ***P<0.001. OCN, osteocalcin; COL1A1,
collagen, type I, α1; PPARG, peroxisome proliferator-activated
receptor-γ; Ctrl, control; pPSCs, pig perivascular stem cells;
pSVF, pig stromal vascular fraction; Ost, osteogenic. |
Discussion
The current study investigated mps as a novel source
of PSCs for translational effects in bone tissue engineering. The
data revealed that mpPSCs could be isolated from SVF with an
identical isolation protocol to hPSCs. The present study further
confirmed the characteristics of mpPSCs, including the morphology
of the cells that adhered to the wall, and their adipogenic and
osteoplastic differentiation ability using Alizarin Red and Oil Red
O staining, the result demonstrated that mpPSCs exhibited the
following characteristics of stem cells: Adherent growth, clonal
formation, multipotent differentiation-osteogenesis and adipogenic.
Additionally, adipogenesis-associated gene expression was measured
using RT-qPCR, and the result demonstrated that compared with the
pSVF group, the expression levels of OCN, COL1A1 and PPARG mRNA in
the mpPSC group was significantly higher (P<0.05). The H&E
staining performed in the subcutaneous adipose tissue of the
posterior neck of the mps indicated that there was abundant SVF in
mps. These data confirm the possibility that mps provide a source
of PSCs. The possible novel source of PSCs identified during the
current study, which may supplement currently confirmed sources, as
described in different species, including humans (9,10),
canines (22), sheep (25) and mice (26). Notably, the same markers were used
to identify PSCs in these different species, using, in the majority
of instances, cross-reactive antibodies. This indicates a
highly-conserved set of perivascular markers for an ancestral
population of tissue regenerative cells.
Adipose tissue is an attractive source of PSCs, as
it is dispensable and accessible with minimal associated morbidity
(27,28), PSCs consist of pericytes and
adventitial cells; pericytes are a type of cell that typically
reside around smaller blood vessels, including capillaries, venules
and arterioles, whereas adventitial cells are located around larger
arteries and veins (10,29). PSCs have demonstrated osteogenic,
chondrogenic, adipogenic, and myogenic potential, indicating the
potential for multiple applications in skeletal regenerative
medicine without culture expansion (9,11,19,22,27,30).
mps are a type of easily cultured animal. The observation that
CD146 and CD34 have a perivascular location in mp tissues, which
coincides with that of established MSC markers is consistent with
the characteristic of pericytes and adventitial cells of PSCs
described in previous reports (22,31).
The current result further confirmed the perivascular location of
MSCs in mps, as demonstrated by H&E staining, which has already
been described in humans and other animals (22,25,30,31).
The cell can only be defined as PSCs if it satisfies
the following conditions: Located in the vascular wall and
exhibiting stem cell characteristics (22,24).
In humans, there are two types of cell that meet the
above-mentioned conditions:
CD45−CD34−CD146+ phenotype of
pericytes and CD45−CD34+CD146−
phenotype of adventitial cells. However, in the mp vascular stem
cell sorting process,
CD45−CD34−CD146+ phenotype and
CD45−CD34+CD146− phenotype cells
were prospectively isolated from SVF from minipigs, and humans had
high homology biological background and the same phenotype, and the
sorted cells demonstrated the following characteristics of stem
cells: Adherent growth, colony formation and multipotential
differentiation; however, immunofluorescence was not performed in
the present study, which could verify that
CD45−CD34−CD146+ and
CD45−CD34+CD146− are mp pericyte
and adventitial cells, respectively. Thus, in order to evaluate the
cross-reactivity of human antibodies with porcine tissue,
immunofluorescent detection of markers of pericytes (CD146),
adventitial cells (CD34), and endothelium (CD31) should be
performed on mp tissue samples in future animal experiments.
In addition, the results of flow cytometry indicated
that compared with man, the cell rate of PSC-like cells from mps
was lower, due to various potential reasons: Firstly, in the
current study, the indirect CD34 antibody marker was used, not the
mp antibody, which may have affected the identification of cell
surface molecules; additionally, the CD146-FITC antibody used in
the present study was a human monoclonal antibody, which worked in
the mp tissues, but with less antibody specificity than in human
cells; so CD34 and CD146 may not identify adventitial cells and
pericytes specifically; finally, blood vessel density in mp fat may
be lower than that of human fat. The cell sorting was not obvious
in flow cytometry, which also supported the inference of poor
antibody specificity. Therefore, it would be prudent to state that
the cells involved in the current experiment are termed mp
CD45−CD34−CD146+ and
CD45−CD34+CD146− progenitors.
In conclusion, the H&E staining results and the
characteristics of stem cells after sorting indirectly support the
existence of vascular stem cells in the fat of mps. In vitro
experiments demonstrate that the adipogenic differentiation
potential of mp vascular stem cells was significantly stronger than
that of non-selected vascular stromal cells. These results lay the
foundation for the future isolation and characterization of
CD146+ and CD34+ cell subpopulations from mp
adipose tissues.
Acknowledgements
Not applicable.
Funding
The present study was supported by the International
Science & Technology Cooperation Program of China (grant no.
2015DFB30040), the National Science Foundations of China (grant
nos. 81300897, 81571815, 81671015, and 81470717) and the Beijing
Municipal Natural Science Foundation (grant no. 7152156).
Availability of data and materials
The datasets used during the present study are
available from the corresponding author on reasonable request.
Authors' contributions
ZC and YZ conceived and designed the study. ZC, CL,
NJ, CZ, YW and HG performed the experiments. ZC and YZ wrote the
paper. All authors read and approved the manuscript.
Ethics approval and consent to
participate
All experimental protocols were approved by the
Review Board of Peking University (Beijing, China; permit no.
LA2014216).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
de Boer HH: The history of bone grafts.
Clin Orthop Relat Res. 1–298. 1988.
|
|
2
|
Kao ST and Scott DD: A review of bone
substitutes. Oral Maxillofac Surg Clin North Am. 19:513–521, vi.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Sarkar SK and Lee BT: Hard tissue
regeneration using bone substitutes: An update on innovations in
materials. Korean J Intern Med. 30:279–293. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Klenerman L: Bone grafts, derivatives and
substitutes. Br J Plas Surg. 47:5861994. View Article : Google Scholar
|
|
5
|
Giannoudis PV, Dinopoulos H and Tsiridis
E: Bone substitutes: An update. Injury. 36 Suppl 3:S20–S27. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sawin PD, Traynelis VC and Menezes AH: A
comparative analysis of fusion rates and donor-site morbidity for
autogeneic rib and iliac crest bone grafts in posterior cervical
fusions. J Neurosurg. 88:255–265. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Frodel JL Jr, Marentette LJ, Quatela VC
and Weinstein GS: Calvarial bone graft harvest. Techniques,
considerations, and morbidity. Arch Otolaryngol Head Neck Surg.
119:17–23. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Derubeis AR and Cancedda R: Bone marrow
stromal cells (BMSCs) in bone engineering: Limitations and recent
advances. Ann Biomed Eng. 32:160–165. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hindle P, Khan N, Biant L and Péault B:
The infrapatellar fat pad as a source of perivascular stem cells
with increased chondrogenic potential for regenerative medicine.
Stem Cells Transl Med. 6:77–87. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Crisan M, Yap S, Casteilla L, Chen CW,
Corselli M, Park TS, Andriolo G, Sun B, Zheng B, Zhang L, et al: A
perivascular origin for mesenchymal stem cells in multiple human
organs. Cell Stem Cell. 3:301–313. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chung CG, James AW, Asatrian G, Chang L,
Nguyen A, Le K, Bayani G, Lee R, Stoker D, Zhang X, et al: Human
perivascular stem cell-based bone graft substitute induces rat
spinal fusion. Stem Cells Transl Med. 3:1231–1241. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lee S, Zhang X, Shen J, James AW, Chung
CG, Hardy R, Li C, Girgius C, Zhang Y, Stoker D, et al: Brief
report: Human perivascular stem cells and Nel-Like protein-1
synergistically enhance spinal fusion in osteoporotic rats. Stem
Cells. 33:3158–3163. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
West CC, Hardy WR, Murray IR, James AW,
Corselli M, Pang S, Black C, Lobo SE, Sukhija K, Liang P, et al:
Prospective purification of perivascular presumptive mesenchymal
stem cells from human adipose tissue: Process optimization and cell
population metrics across a large cohort of diverse demographics.
Stem Cell Res Ther. 7:472016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Nakata M, Nakagomi T, Maeda M, Nakano-Doi
A, Momota Y and Matsuyama T: Induction of perivascular neural stem
cells and possible contribution to neurogenesis following transient
brain ischemia/reperfusion injury. Transl Stroke Res. 8:131–143.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhao H, Feng J, Seidel K, Shi S, Klein O,
Sharpe P and Chai Y: Secretion of shh by a neurovascular bundle
niche supports mesenchymal stem cell homeostasis in the adult mouse
incisor. Cell Stem Cell. 14:160–173. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Eliasberg CD, Dar A, Jensen AR, Murray IR,
Hardy WR, Kowalski TJ, Garagozlo CA, Natsuhara KM, Khan AZ, Mcbride
OJ, et al: Perivascular stem cells diminish muscle atrophy
following massive rotator cuff tears in a small animal model. J
Bone Joint Surg Am. 99:331–341. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
James AW, Zara JN, Corselli M, Chiang M,
Yuan W, Nguyen V, Askarinam A, Goyal R, Siu RK, Scott V, et al: Use
of human perivascular stem cells for bone regeneration. J Vis Exp.
e29522012.PubMed/NCBI
|
|
18
|
James AW, Zara JN, Zhang X, Askarinam A,
Goyal R, Chiang M, Yuan W, Chang L, Corselli M, Shen J, et al:
Perivascular stem cells: A prospectively purified mesenchymal stem
cell population for bone tissue engineering. Stem Cells Transl Med.
1:510–519. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
James AW, Zara JN, Corselli M, Askarinam
A, Zhou AM, Hourfar A, Nguyen A, Megerdichian S, Asatrian G, Pang
S, et al: An abundant perivascular source of stem cells for bone
tissue engineering. Stem Cells Transl Med. 1:673–684. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Mcintosh K, Zvonic S, Garrett S, Mitchell
JB, Floyd ZE, Hammill L, Kloster A, Di Halvorsen Y, Ting JP, Storms
RW, et al: The immunogenicity of human adipose-derived cells:
Temporal changes in vitro. Stem Cells. 24:1246–1253. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ruetze M and Richter W: Adipose-derived
stromal cells for osteoarticular repair: Trophic function versus
stem cell activity. Expert Rev Mol Med. 16:e92014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
James AW, Zhang X, Crisan M, Hardy WR,
Liang P, Meyers CA, Lobo S, Lagishetty V, Childers MK, Asatrian G,
et al: Isolation and characterization of canine perivascular
stem/stromal cells for bone tissue engineering. PLoS One.
12:e01773082017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Fu Y, Liu S, Cui SJ, Kou XX, Wang XD, Liu
XM, Sun Y, Wang GN, Liu Y and Zhou YH: Surface chemistry of
nanoscale mineralized collagen regulates periodontal ligament stem
cell fate. ACS Appl Mater Interfaces. 8:15958–15966. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hindle P, Baily J, Khan N, Biant LC,
Simpson AH and Péault B: Perivascular mesenchymal stem cells in
sheep: Characterisation and autologous transplantation in a model
of articular cartilage repair. Stem Cells Dev. 25:1659–1669. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kang SG, Shinojima N, Hossain A, Gumin J,
Yong RL, Colman H, Marini F, Andreeff M and Lang FF: Isolation and
perivascular localization of mesenchymal stem cells from mouse
brain. Neurosurgery. 67:711–720. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zuk PA, Zhu M, Ashjian P, De Ugarte DA,
Huang JI, Mizuno H, Alfonso ZC, Fraser JK, Benhaim P and Hedrick
MH: Human adipose tissue as a source of multipotent stem cells. Mol
Biol Cell. 13:4279–4295. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Frese L, Dijkman PE and Hoerstrup SP:
Adipose tissue-derived stem cells in regenerative medicine.
Transfus Med Hemother. 43:268–274. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Corselli M, Chen CW, Sun B, Yap S, Rubin
JP and Péault B: The tunica adventitia of human arteries and veins
as a source of mesenchymal stem cells. Stem Cells Dev.
21:1299–1308. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Askarinam A, James AW, Zara JN, Goyal R,
Corselli M, Pan A, Liang P, Chang L, Rackohn T, Stoker D, et al:
Human perivascular stem cells show enhanced osteogenesis and
vasculogenesis with Nel-like molecule I protein. Tissue Eng Part A.
19:1386–1397. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Esteves CL, Sheldrake TA, Mesquita SP,
Pesántez JJ, Menghini T, Dawson L, Péault B and Donadeu FX:
Isolation and characterization of equine native MSC populations.
Stem Cell Res Ther. 8:802017. View Article : Google Scholar : PubMed/NCBI
|