Introduction
Mesenchymal stem cells (MSCs) are derived from the
early developmental mesoderm, and they are characterized as
undifferentiated cells. They are considered to play important roles
in development, postnatal growth, repair, regeneration, and
homeostasis. These cells have attracted much interest regarding
their possible clinical applications because of their self-renewing
capacity and multi-lineage differentiation potential (1). MSCs can be used as ideal seed cells
for the repair of injury to tissue and organs. In addition, these
cells may also have immunomodulatory benefits in the treatment of
autoimmune diseases and systemic diseases (2–4).
Several investigations have shown the efficacy of MSCs in
regenerative medicine both in vitro and in vivo
(5–7). Although MSCs have been obtained from
bone marrow, umbilical cord, and various odontogenic tissues,
differences in their biological characteristics have not been fully
clarified.
MSCs obtained from human bone marrow (BMSCs) have
been intensively studied since their discovery. They have been
demonstrated to have strong self-renewal capacity and can
differentiate into a variety of tissue cells in a specific
environment. They have been used widely in animal experimental
models and clinical therapies for hematological system diseases,
cardiovascular diseases, nervous system diseases, and
osteogenesis-related disorders (8–12).
Stem cells from human exfoliated deciduous teeth
(SHED) have been isolated from naturally exfoliated deciduous teeth
with the capacity to differentiate into osteogenic and odontogenic
cells, adipocytes, and neural cells. SHEDs are derived from a very
accessible tissue resource and are capable of providing enough
cells for potential clinical application via their high
proliferation rate and expression of telomerase (13).
Umbilical cord-derived mesenchymal stem cells
(UCMSCs) collected from umbilical cord tissues of healthy full-term
babies have excellent proliferation potential and are free of
ethical issues (14). UCMSCs have
morphological, phenotypic, and multilineage differentiation
potential similar to that of BMSCs; they are able to differentiate
into functional hepatocyte-like cells in vitro, but their
immunogenicity remains low (15).
Human gingival tissue from the oral cavity can often
be easily obtained as a discarded biological sample without any
morbidity or scar formation. Gingiva-derived mesenchymal stem cells
(GMSCs) can be easily isolated from the gingival tissue, can be
expanded easily in vitro, and possess multipotent
differentiation potential and anti-inflammatory properties
(16,17). GMSCs are capable of regenerating
bone defects, and they may be potentially useful in the
reconstruction and regeneration of bone defects (18).
Besides their function in tissue reconstruction and
regeneration, the low immunogenicity and immunoregulatory potential
of MSCs are advantageous for their clinical application. It has
been reported that BMSCs do not express major histocompatibility
complex (MHC) class II molecules or costimulatory molecules
required for T-cell activation, which are responsible for
transplant rejection (19–21). UCMSCs can maintain low
immunogenicity when they differentiate into other tissue cells
(15); SHEDs show superior
immunomodulatory effects compared to BMSCs (22), and GMSCs have reproducible and
powerful immunomodulatory functions (23).
MSCs can be used for the treatment of many diseases
associated with defective tissue regeneration and immune
regulation. However, it is difficult to decide which type of stem
cell should be used in clinical treatment according to the safety
and immunogenicity of the cells. Therefore, we need to understand
the biological characteristics of MSCs from different sources and
the differences in their tumorigenicity and immunogenicity. BMSCs
from alveolar bone and GMSCs can be obtained from discarded tissues
during dental surgery, and obtaining the samples is less invasive
to the donors. SHEDs and UCMSCs are obtained from discarded medical
tissues, and are easy to obtain, non-invasive to donors, and
plentiful. In this study, we compared the cell proliferation
ability, tumorigenicity, and immunogenicity of BMSCs, SHEDs,
UCMSCs, and GMSCs. The results provide information that is helpful
for the clinical application of these cells.
Materials and methods
Isolation and culture of human
MSCs
SHEDs, GMSCs, and UCMSCs were donated by the Oral
Stem Cell Bank of Beijing, Tason Biotech Co., Ltd. (Beijing,
China), each stem cell derived from three different individuals.
Alveolar bone marrow samples were obtained from the mandibular bone
of healthy patients (from 1 male and 2 female donors, aged 30, 28,
and 20 years) after obtaining written informed consent. Sample
collection was approved by the Institutional Review Board of Peking
University School and Hospital of Stomatology (Beijing, China; no.
PKUSSIRB-201734036). BMSCs were isolated from mandibular alveolar
bone marrow. Cultures of all four types of MSCs were maintained in
α-modified Eagle's minimum essential medium (α-MEM; Gibco; Thermo
Fisher Scientific, Inc., Waltham, MA, USA) supplemented with 10%
fetal bovine serum (FBS; Biological Industries, Madison, WI, USA)
in 5% CO2 at 37°C. The cells were used in experiments
after three to five passages, and for each experiment, all MSCs had
the same passage number.
Biological characteristics of
MSCs
Cells in the logarithmic growth phase were detached
with 0.1% trypsin/0.1% EDTA to produce a single-cell suspension.
The cells were labeled with rabbit antihuman nestin (Merck KGaA,
Darmstadt, Germany), vimentin, NANOG, and CD90 antibodies, mouse
antihuman CD105 (all Abcam, Cambridge, MA, USA) and CD34 (BD
Pharmingen, San Diego, CA, USA) antibodies, and goat antihuman
cytokeratin19 (CK19; Santa Cruz Biotechnology, Santa Cruz, CA, USA)
antibody, respectively, and approximately 1×106 cells
were used for the detection of each molecule. Alexa Fluor
488-conjugated antirabbit, antimouse, and antigoat IgGs were used
as the secondary antibodies. The labeled cells were thoroughly
washed by centrifugation 3 times, resuspended in PBS solution, and
flow cytometry (Cytomics Flow Cytometer EPICS XL; Beckman Coulter,
Fullerton, CA, USA) was used to detect the fluorescence intensity
and positive rate.
The self-renewal capacity of the MSCs was evaluated
by colony-forming efficiency assays. 1×102 cells at
passage 3 were seeded in 6-well plates at 37°C with 5%
CO2 for 14 days. Then the cells were stained with 0.5%
crystal violet (Sigma-Aldrich; Merck KGaA) for 5 min at room
temperature. Stained colonies with >50 cells were counted.
Colony-forming efficiency was calculated as colony-forming unit
numbers.
MSCs were induced by osteogenic, adipogenic, and
chondrogenic differentiation kits (Biowit Technologies; Shenzhen,
China) according to the manufacturer's protocol. 2×105
cells at passage 3 were seeded in 24-well plates at 37°C with 5%
CO2. When the cells became 70–80% confluent, the medium
was replaced by differentiation induction medium for 2–3 weeks. The
cells were analyzed for osteogenesis, adipogenesis, and
chondrogenesis by Alizarin Red staining, Oil Red O staining, and
Alcian Blue staining.
Analysis of cell proliferation
capacity
MSCs were seeded into 96-well plates at a density of
3×103 cells/well and then cultured for 7 days. Cell
counting kit-8 (CCK-8; Dojindo, Kumamoto, Kyushu, Japan) solution
was added to each well of the plate and the absorbance was measured
at 450 nm every 24 h according to the manufacturer's protocol.
Tumorigenicity assay
To evaluate tumorigenicity in vitro,
anchorage-independent growth was assessed with soft agar colony
formation assay using a 6-well plate. The base agar layer was
prepared from a 0.6% soft agar solution containing α-MEM with 10%
FBS. Then, 1×104 cells were suspended in α-MEM
containing 10% FBS and 0.35% agar solution and plated onto the base
layer. HeLa cells were used as positive control. Plates were
incubated at 37°C with 5% CO2 for 21 days, after which
colony formation was observed under a microscope.
The animal study was approved by the Animal Ethics
Committee of China-Japan Friendship Hospital (Beijing, China; no.
170103). To examine tumorigenicity in vivo, 8-week-old Nu/Nu
male mice were purchased from Beijing Vital River Laboratory Animal
Technology Company [quality certificate: SCXK (Beijing) 2012-0001].
All animals were housed in the specific pathogen-free (SPF)
facility [quality certificate: SYXK (Beijing) 2010–0011] in the
Institute of Clinical Medical Sciences of China-Japan Friendship
Hospital, and rats were maintained on a 12-h light:12-h dark cycle
with free access to rodent chow and water. Mice were randomly
divided into four groups of six mice each. All four types of MSCs
were suspended in α-MEM medium at a density of 2×106
cells/100 µl, respectively. In each group, six nude mice were
anesthetized via 2% isoflurane inhalation, and 2×106
MSCs were subcutaneously inoculated into the backs of each mouse.
The grafts were observed for 6 months; twice daily for clinical
signs in the mouse and twice weekly for the presence of a tumor.
Tumor volume was calculated according to the following formula: V
(mm3)=(width2 × length)/2.
At passage 4 of all four types of MSCs, karyotypes
were analyzed respectively. 0.4 µg/ml colchicine (Gibco; Thermo
Fisher Scientific, Inc.) was added to 1×106 cells for 4
h. After centrifugation, hypotonic treatment, and then fixed. The
substrate was dripped onto a glass slide, dried at 80°C for 2 h,
then digested with 0.5% trypsin for 10 sec, stained with 10%
Giemsa, and observed and analyzed under a microscope. Chromosome
analysis was carried out by applying a scatter plot of the natural
distribution and G-bands according to the guidelines of the
International System for Chromosome Nomenclature (ISCN).
Immunogenicity assay
Mix lymphocyte proliferation
assay
Human peripheral blood mononuclear cells (PBMCs)
were isolated from healthy donors [sample collection was approved
by the Institutional Review Board of Peking University School and
Hospital of Stomatology (PKUSSIRB-201311108) and written informed
consent were obtained from all the donors] using human peripheral
blood lymphocyte separation solution (Tianjin HaoYang Biological
Manufacture Co., Ltd., Tianjin, China) and density gradient
centrifugation. Cells were cultured in RPMI-1640 medium
supplemented with 10% FBS, 2 mM glutamine (Gibco; Thermo Fisher
Scientific, Inc.), 100 U/ml penicillin, and 100 µg/ml streptomycin
(North China Pharmaceutical Limited by Share Ltd., Shijiazhuang,
China) at 37°C with 5% CO2. The PBMCs were depleted of
adherent cells overnight and further cultured in PBMC medium. The
experimental cells were divided into the following 6 groups:
Control group: 1×106/well PBMCs were cultured in a
24-well plate in normal medium as described above; positive control
group: 10 µg/ml PHA (Sigma-Aldrich; Merck KGaA) with
1×106 PBMCs; and stem cell group: Each group of the four
types of MSCs, 2×105/well, were co-cultured with
1×106 PBMCs, and the MSCs were treated with 25 µg/ml
mitomycin C (Sigma-Aldrich; Merck KGaA) at 37°C for 30 min before
seeding into wells. Each group was co-cultured for 72 h, the PBMCs
were harvested and placed in 96-well plates, and CCK-8 was used to
measure proliferation.
Detection of HLA-I, HLA-DR, CD80, and
CD86
All four types of MSCs were detected by flow
cytometry using fluorescein-isothiocyanate-conjugated or
phycoerythrin-conjugated antibodies specific for HLA-I, HLA-DR
(both Abcam), CD80, and CD86 (both R&D Systems, Minneapolis,
MN, USA) as previously described. To enhance the expression of
immune related surface molecules, MSCs were pretreated with 100
U/ml IFN-γ (interferon-γ; Peprotech, Rocky Hill, NJ, USA) for 72 h
as previously described (24), and
then the IFN-γ-treated MSCs (MSCs+IFN-γ) were detected by flow
cytometry using monoclonal antibodies specific for HLA-I, HLA-DR,
CD80, and CD86.
Statistical analysis
The analysis was conducted with SPSS 19.0 software
(IBM Corp., Armonk, NY, USA), and the data are expressed as the
mean ± standard deviation. Statistical comparisons between two
groups were performed by the t-test, and a one-way analysis of
variance followed by Tukey's test were used for comparisons among
multiple groups. P<0.05 was considered to indicate a
statistically significant difference. All experiments were repeated
at least three times as previously described (24,25).
Results
Biological characteristics of
MSCs
Cell surface marker analysis by flow cytometry
showed that all four types of MSCs positively expressed MSC
markers, including vimentin, CD90, and CD105, and they also
positively expressed nestin and NANOG, but they showed no
expression of hematopoietic stem cell marker CD34 and epithelial
cell marker CK19 (Fig. 1).
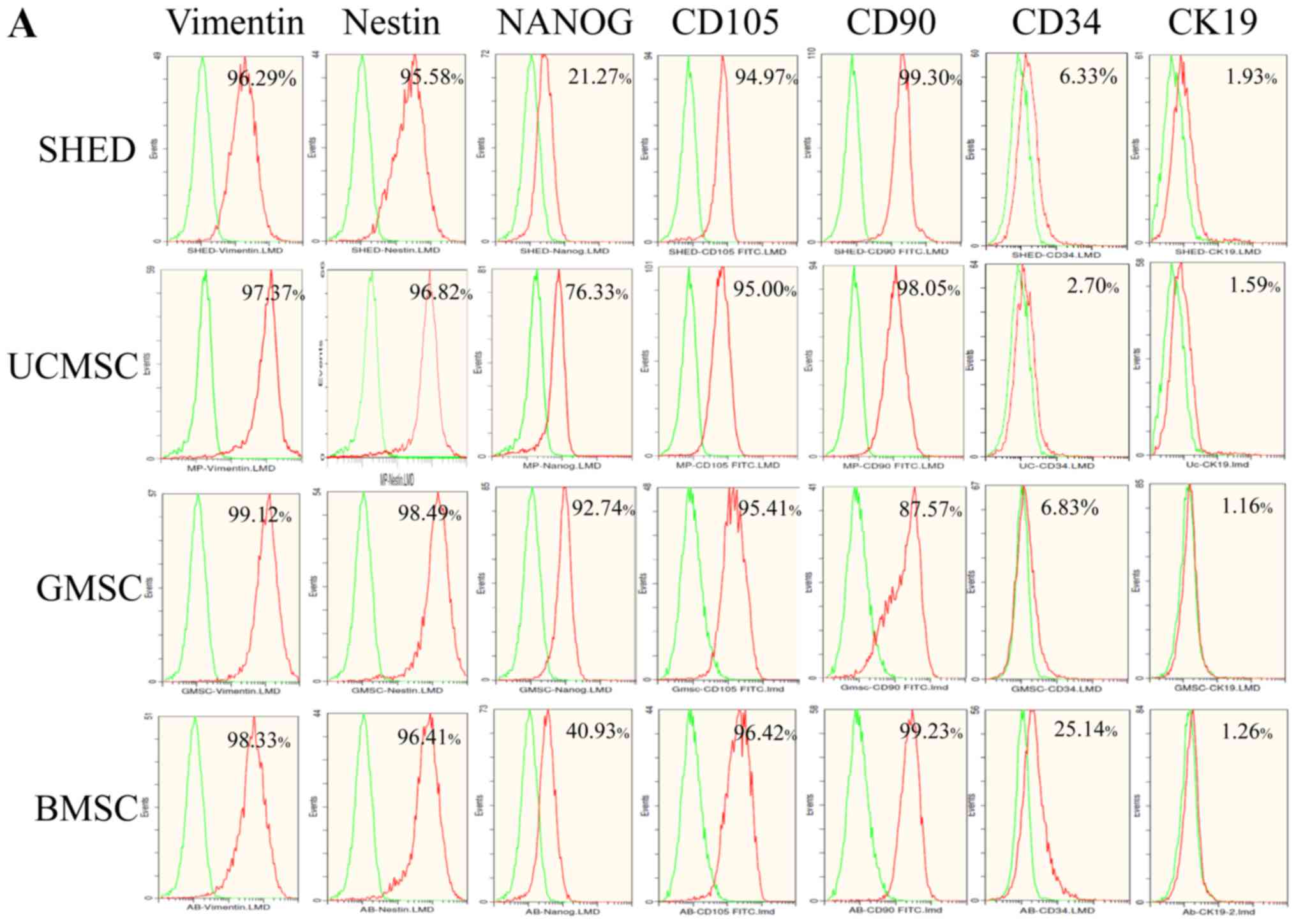 | Figure 1.(A) Surface marker expression of
MSCs. (B) All four types of MSCs were positive for vimentin,
nestin, NANOG, CD105 and CD90, and were negative for CD34 and CK19.
Results represent the mean ± standard deviation of three
independent experiments. *P<0.05, **P<0.01 and ***P<0.001,
as indicated. MSCs, mesenchymal stem cells; CD, cluster of
differentiation; SHEDs, stem cells from human exfoliated deciduous
teeth; UCMSCs, umbilical cord-derived mesenchymal stem cells;
GMSCs, gingiva-derived mesenchymal stem cells; BMSCs, bone marrow
mesenchymal stem cells; CK19, cytokeratin19; NS, not
significant. |
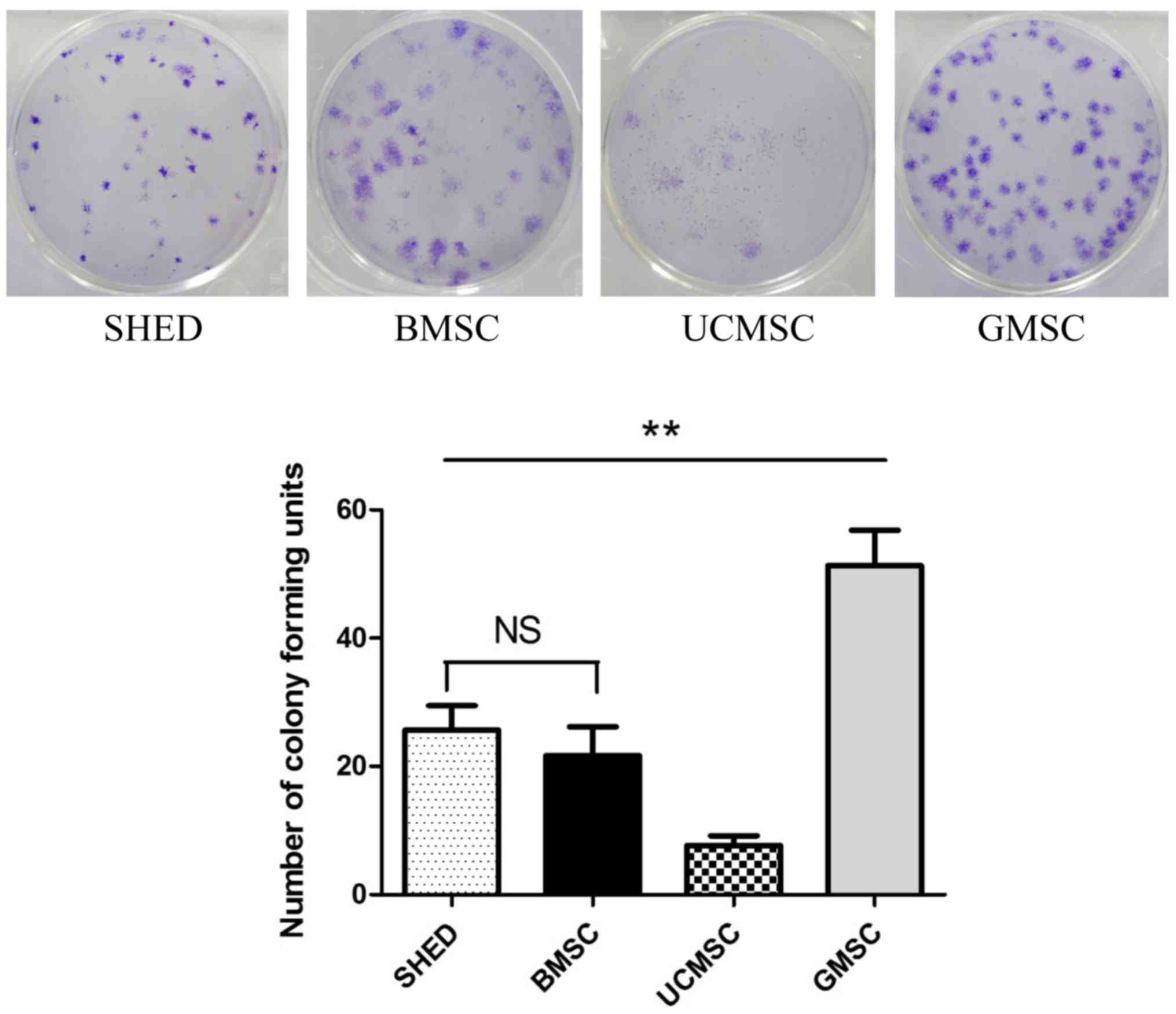
Colony-forming efficiency assays were used to
examine the self-renewal capacity of MSCs. All four MSCs at passage
3 seeded in 6-well plates at 1×102 cells/well for 14
days can form colonies. GMSCs exhibited highest self-renewal
capacity, UCMSC displayed the lowest colony formation units,
between SHED and BMSC there were no significant differences in
colony-forming efficiency (Fig.
2).
To investigate the differentiation potential of the
MSCs, cells were induced to osteogenic, adipogenic, and
chondrogenic differentiation. Osteogenic differentiation was
observed with Alizarin Red staining, and calcium deposition was
seen in all four MSCs. Adipogenic differentiation was verified by
Oil Red O staining, and the accumulation of cytoplasmic lipid
vacuoles was distinctly observed. Chondrogenic differentiation was
verified by Alcian Blue staining and was demonstrated in all tested
cells (Fig. 3). The results
indicated successful differentiation of MSCs into osteogenic,
adipogenic, and chondrogenic lineages.
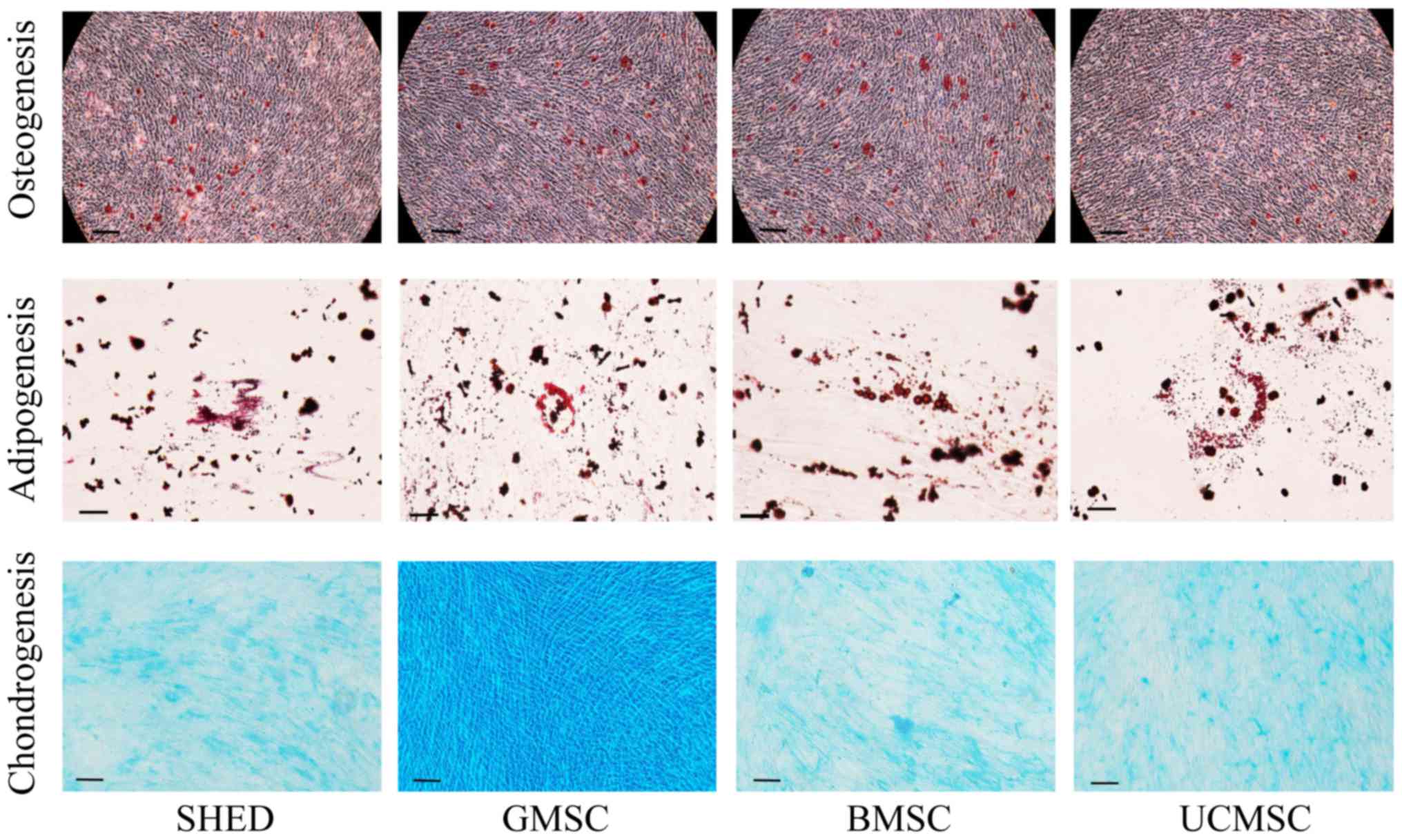 | Figure 3.Osteogenic, adipogenic and
chondrogenic differentiation of MSCs. MSCs were induced to
differentiate toward osteogenic, adipogenic and chondrogenic
lineages as verified by Alizarin Red, Oil Red O and Alcian Blue
staining, respectively (osteogenic and chondrogenic
differentiation: scale bar, 100 µm; adipogenic differentiation:
scale bar, 50 µm). MSCs, mesenchymal stem cells; SHEDs, stem cells
from human exfoliated deciduous teeth; UCMSCs, umbilical
cord-derived mesenchymal stem cells; GMSCs, gingiva-derived
mesenchymal stem cells; BMSCs, bone marrow mesenchymal stem
cells. |
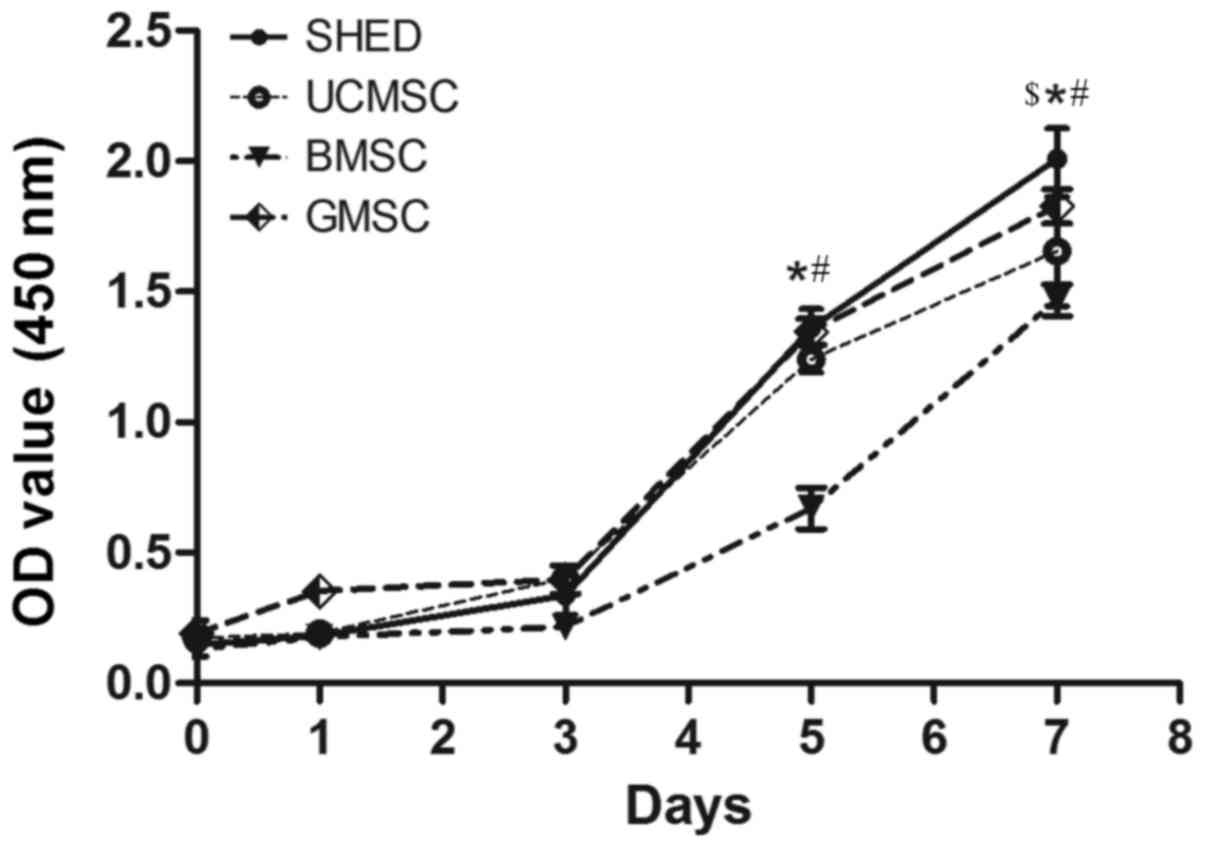
Proliferation capacity of MSCs
Cell proliferation was monitored over a period of 7
days post-seeding (Fig. 4). The
proportion of all MSCs increased markedly from the third day of
proliferation, and we found that SHED showed enhanced proliferation
capacity compared to the other MSCs. On the fifth day, except for
between the SHED and GMSC groups, and the GMSC and UCMSC groups,
there were significant differences among the other groups
(P<0.05). On the seventh day, except for between the GMSC and
UCMSC groups, there were significant differences among the other
groups (P<0.05).
Tumorigenicity of MSCs
To compare colony formation of the four types of
MSCs in vitro, we performed soft agar colony assays using
1×104 cells/well in a 6-well plate, and HeLa cells were
used as a positive control. After being cultured for 21 days, none
of the four MSC groups formed colonies, but the HeLa cell group
formed a large number of colonies (Fig. 5A). HeLa cells, but not the four
types of MSCs, showed anchorage-independent cell growth in soft
agar. MSCs showed a notably lower ability to form colonies than
HeLa cells in vitro.
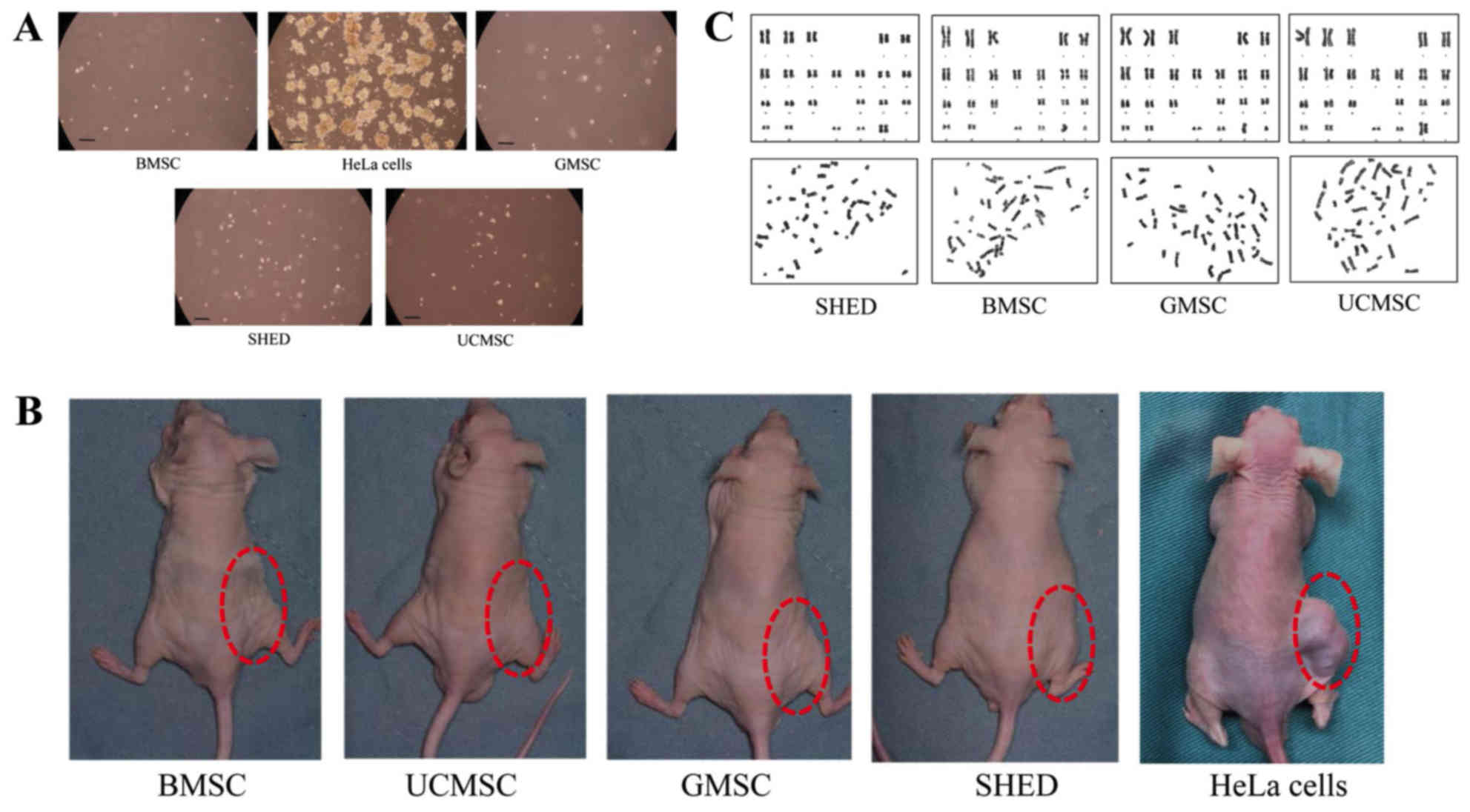 | Figure 5.Tumorigenicity of MSCs. (A) Soft agar
colony assay for MSCs and HeLa cells. Representative images of
colonies in the soft agar assay for BMSCs, GMSCs, SHEDs, UCMSCs and
HeLa cells. A large number of colonies were observed in the HeLa
group, but no colonies were identified in the MSC groups (scale
bar, 200 µm). (B) BMSCs, UCMSCs, GMSCs, SHEDs and HeLa cells were
transplanted into nude mice. Following 2 months, tumors were
identified in the injection areas of the group transplanted with
HeLa cells, but at 6 months, no tumors were observed in the other
groups transplanted with stem cells. (C) Karyotype analysis of
SHEDs, BMSCs, GMSCs and UCMSCs. The upper panel shows G-bands and
the lower panel shows natural distribution. No karyotype
abnormalities were identified for any cells. SHEDs, stem cells from
human exfoliated deciduous teeth; UCMSCs, umbilical cord-derived
mesenchymal stem cells; GMSCs, gingiva-derived mesenchymal stem
cells; BMSCs, bone marrow mesenchymal stem cells. |
Subcutaneous inoculation of MSCs into nude mice
showed that none of the four types of MSCs caused any clinical
signs during the 6-month experimental period, and there was no
tumor formation in any mice at the injection area and around that
area, but abnormal masses at injection areas were found in the HeLa
cell group (Fig. 5B). In the HeLa
cell group, 2 weeks after subcutaneous inoculation abnormal masses
at injection areas can be seen. Continuous observation for 2
months, the maximum tumor size was 1,884 mm3, no
multiple tumors were observed.
There were no significant structural chromosomal
abnormalities/aberrations in the karyotypes of any diploid cells,
indicating that the karyotypes were stable.
Immunogenicity of MSCs
To investigate whether allogeneic human MSCs can
stimulate the proliferation of PBMCs, PBMCs were co-cultured with
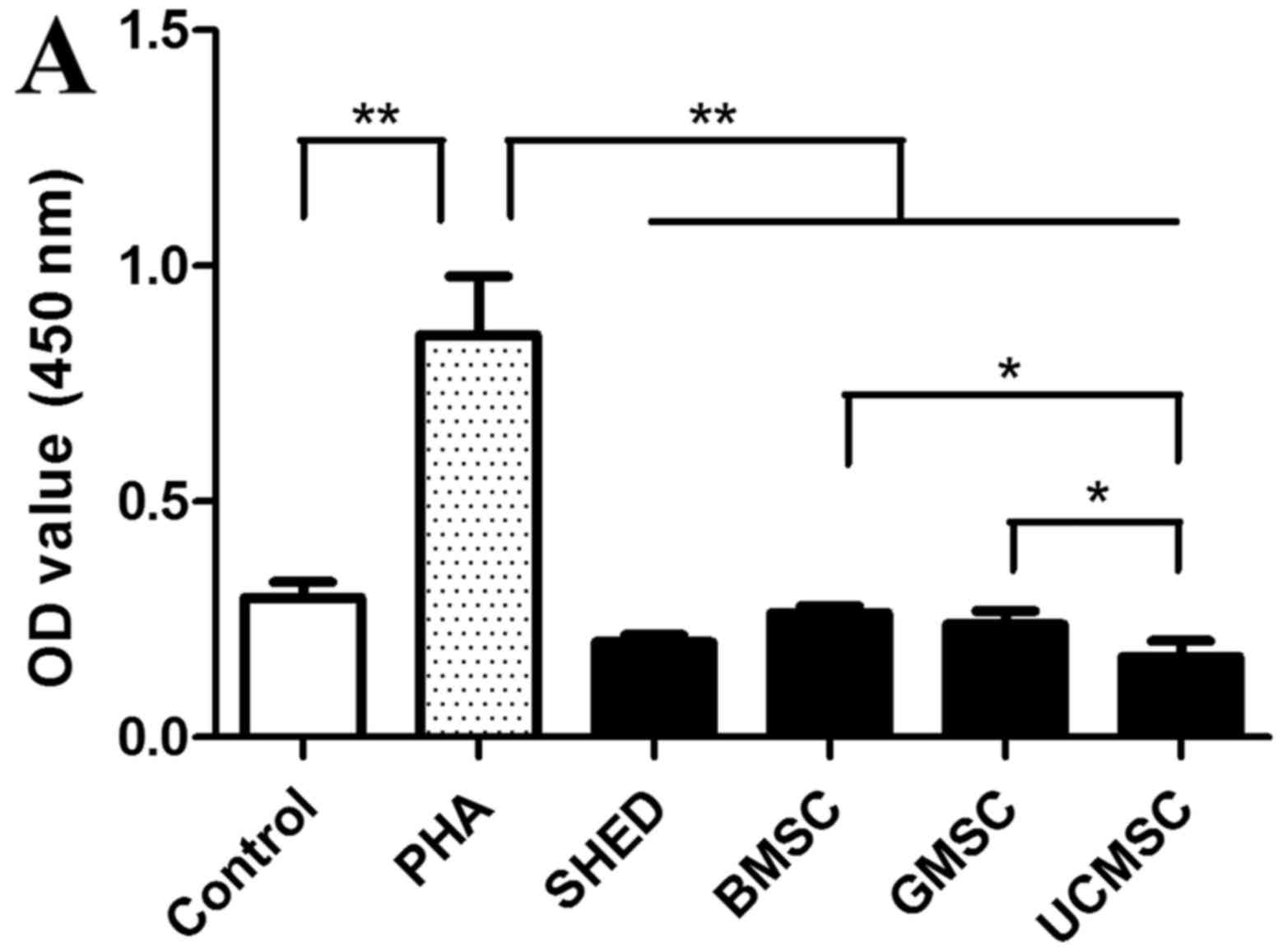
MSCs that were pretreated with mitomycin C. As shown in Fig. 6A, after a mixed co-culture for 3
days, phytohemagglutinin (PHA) induced a very strong proliferative
response in allogeneic lymphocytes (P<0.01), but none of the
allogeneic MSCs elicited any proliferative response compared to the
control group; MSCs inhibited the proliferation of lymphocytes, and
all MSC groups showed a significant difference from the control
group (P<0.01). Comparing the four types of MSCs, the BMSC and
UCMSC groups (P<0.05) and the GMSC and UCMSC groups (P<0.05)
showed a significant difference, but there was no significant
difference between the other MSC groups.
 | Figure 6.Immunogenicity characterization of
MSCs. (A) The proliferation capacity of peripheral blood
mononuclear cells following mixed co-culture for 3 days. Cell
Counting kit-8 assay revealed that the relative OD values for the
four types of MSCs were significantly lower than those in the
control and PHA groups. When comparing the four types of MSCs, the
OD value for UCMSCs was lower than that of BMSCs and GMSCs. Results
represent the mean ± standard deviation of five independent
experiments. (B and C) All MSCs exhibited high expression of HLA-I
prior to or following IFN-γ treatment. (D and E) All MSCs had a
lower expression of HLA-DR prior to IFN-γ treatment, but following
IFN-γ treatment, the expression of HLA-DR was significantly
upregulated. (F and G) The expression of CD80 was very low prior to
and following stimulation with IFN-γ. (H and I) The expression of
CD86 was very low prior to and following stimulation with IFN-γ.
Results represent the mean ± standard deviation of three
independent experiments. *P<0.05, **P<0.01 and ***P<0.001,
as indicated. NS, not significant; MSCs, mesenchymal stem cells;
OD, optical density; PHA, phytohemagglutinin; SHEDs, stem cells
from human exfoliated deciduous teeth; UCMSCs, umbilical
cord-derived mesenchymal stem cells; GMSCs, gingiva-derived
mesenchymal stem cells; BMSCs, bone marrow mesenchymal stem cells;
HLA, human leukocyte antigen; -DR, antigen D related; IFN-γ,
interferon-γ; CD, cluster of differentiation. |
The results of flow cytometry revealed that all four
types of MSCs expressed HLA-I, but they did not express HLA-DR or
the costimulatory molecules CD80 and CD86. After treatment with
IFN-γ for 72 h, the expressions of HLA-I, CD80, and CD86 showed no
obvious change in the MSCs, but the expression of HLA-DR was
significantly upregulated in BMSCs only (Fig. 6B-I).
Discussion
MSCs have been widely used in vivo and in
vitro because of their strong self-renewal, proliferation,
multilineage differentiation, and immunomodulatory abilities. MSCs
from different sources may differ in their biological
characteristics. Understanding the differences in biological
characteristics of MSCs from different sources can guide the
selection of more suitable cell sources in clinical applications
given different treatment requirements and is helpful for selecting
safer and more efficient seed cells in clinical treatment and
scientific research. In this study, we investigated the differences
in biological characteristics, including surface markers,
colony-forming efficiency, multi-potent differentiation,
proliferation capacity, tumorigenicity, and immunogenicity among
these human MSCs from different origins, and we advanced our
understanding of the advantages of each cell.
It has been found that MSCs have similar phenotypes,
and the expression of MSC markers associated with stress or aging
remains unchanged (26). All four
types of MSCs in this study positively expressed MSC surface
markers, including vimentin, CD105, and CD90, and they did not
express the hematopoietic stem cell marker CD34 or the epithelial
cell marker CK19. Nestin and NANOG are markers of embryonic stem
cells, and they play an important role in the maintenance of
pluripotency and self-renewal (27). All four types of MSCs positively
expressed nestin and NANOG, which indicates that the four types of
MSCs maintained pluripotency and have good self-renewal
ability.
Self-renewal and multi-potent differentiation are
two important properties of MSCs. All four types of MSCs in this
study had good colony-forming efficiency and could be induced to
differentiate into osteogenic, adipogenic, and chondrogenic
lineages. It has been suggested that the MSC cell type should be
selected depending on the regenerative treatment regimen (28). In addition, the proliferation
capacity of stem cells is also an important factor in stem cell
therapies. SHEDs and UCMSCs have shown a higher proliferation
capacity in comparison with BMSCs in previous studies (13,26).
Our study showed that the strongest proliferative ability was found
in SHEDs, followed by GMSCs and UCMSCs, and the lowest
proliferative ability was found in BMSCs; these results are
consistent with previous studies by other groups. SHEDs are derived
from the dental pulp tissue of deciduous teeth, an immature tissue.
The proliferative capacity of SHEDs is relatively strong since they
are in an active state and show high telomerase activity when
deciduous teeth are replaced by permanent teeth (22).
As UCMSCs are separated from the fetal umbilical
cord, a degenerative tissue, they exhibit less proliferative
capacity than SHEDs. GMSCs have a higher proliferative capacity
than BMSCs, and this may be related to their active state and fast
oral metabolism.
The evaluation of bio-safety is important for
clinical therapies utilizing MSCs, including external safety and
intrinsic safety tests. The external safety tests include pathogen
detection, mycoplasma detection, the asepsis test, and the
endotoxin test, while intrinsic safety tests include acute and
chronic toxicities, tumorigenicity, immunogenicity, and chromosome
aberrations.
In this study, stem cells at passages 3 to 5 were
used for experiments; these cells are relatively young, and their
performance is relatively stable. Cell clones were not formed in
vitro under soft agar culture in any of the four types of MSCs.
After in vivo transplantation into nude mice, the stem cells
did not form any abnormal tumor tissues for up to 6 months, and
karyotype analysis showed that no karyotypic abnormality was found
in any of the four types of MSCs, thus indicating the safety of all
four. These results are consistent with those obtained by other
groups (26,29).
MSCs have been demonstrated to have some
immunomodulatory functions in vitro, such as direct
suppression of allogeneic and mitogenic T-cell proliferation
(30). BMSCs have been implicated
as a potentially feasible treatment approach for several diseases,
such as graft-versus-host disease (31) and autoimmune diseases (32). Due to their immunomodulatory
properties, SHEDs are an accessible and feasible MSC source for
treating immune disorders like systemic lupus erythematosus
(22). The wide usefulness of MSCs
can be attributed to their low immunogenicity and immunomodulatory
functions. When MSCs were co-cultured with PBMCs under cell-cell
contact conditions, none of the allogeneic MSCs elicited a
proliferative response in PBMCs, indicating that the four types of
MSCs displayed low immunogenicity. This result is consistent with
the notion that the cell-cell contact mechanism may partly
contribute to MSC-mediated suppression of PBMC proliferation
(16,25,33).
UCMSCs are isolated from fetal tissue, and as they are more
primitive than adult stem cells, they showed higher
immunomodulatory ability, but the immunomodulatory ability of BMSCs
and GMSCs showed no significant difference in this study. This
result is not completely consistent with previous studies (16,26).
To further understand the mechanism of low
immunogenicity of the four types of MSCs, we analyzed the
expression of HLA and costimulatory molecules (CD80 and CD86). It
is well known that T-lymphocyte activation requires two signals,
the T-cell receptors with antigenic peptides presented by HLA-DR
molecules and the costimulatory molecules CD80 and CD86 (34). HLA-I is expressed on all nucleated
cell surfaces, which are mainly responsible for the presentation of
endogenous antigens, whereas HLA-DR is expressed only on the
surface of antigen-presenting cells, which are mainly responsible
for the presentation of exogenous antigens. However, recent studies
have found that BMSCs also express HLA-DR, and its expression level
is affected by individual differences and the cell microenvironment
(35). All four types of MSCs
expressed a high level of HLA-I, but HLA-DR expression was very low
under resting conditions. However, the expression of HLA-DR was
upregulated in the four types of MSCs, especially in BMSCs, after
stimulation with IFN-γ, suggesting that BMSCs exhibited the best
antigen-presenting potential. Also, the expression of costimulatory
molecules CD80 and CD86 in all four types of MSCs was low either
under resting conditions or under stimulation with IFN-γ.
In conclusion, the populations of MSCs derived from
different sources exhibited variability in their proliferative
capacity and immunomodulatory ability, although they displayed
similar phenotypes. Based on these results, in which SHEDs and
GMSCs showed a higher proliferation capacity, we conclude that
SHEDs and GMSCs are perhaps suited for tissue regeneration-related
cellular therapies. UCMSCs showed higher immunomodulatory ability,
so they are better suited for cellular therapy for some
immune-related diseases. BMSCs can more easily induce immune
reactions than the other three MSCs in the host after cell
transplantation. These data will provide helpful information for
the clinical application of MSCs.
Acknowledgements
The authors would like to thank the Oral Stem Cell
Bank of Beijing, Tason Biotech Co., Ltd. for their assistance in
stem cell collection.
Funding
The present study was supported by the National
Natural Science Fund Emergency Management Project (grant no.
81541110), the Tason Oral Medicine Development Fund (grant no.
2016.7-2019.12), and the International Scientific and Technological
Cooperation and Exchange (grant no. 2014DFA31520).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
L-HG and B-HX designed the present study. JL and
S-QX performed the experiments and analyzed the data. SY
contributed to sample collection, acquisition and data analysis.
Y-MZ contributed to animal study and data analysis. JL and Y-MZ
drafted the manuscript and revised it critically for important
intellectual content. All authors read and approved the final
manuscript, and agreed to be accountable for all aspects of the
study.
Ethics approval and consent to
participate
The present study was approved by the Institutional
Review Board of Peking University School and Hospital of
Stomatology (no. PKUSSIRB-201734036); written informed consent was
obtained from all participants. The animal studies were approved by
the Animal Ethics Committee of the China-Japan Friendship Hospital
(no. 170103).
Patient consent for publication
Written informed consent was obtained from all
participants.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
MSCs
|
mesenchymal stem cells
|
|
BMSCs
|
bone marrow mesenchymal stem cells
|
|
SHED
|
stem cells from human exfoliated
deciduous teeth
|
|
UCMSCs
|
umbilical cord-derived mesenchymal
stem cells
|
|
GMSCs
|
gingiva-derived mesenchymal stem
cells
|
|
α-MEM
|
α-modified Eagle's minimum essential
medium
|
|
FBS
|
fetal bovine serum
|
|
CK19
|
cytokeratin 19
|
|
EDTA
|
ethylenediaminetetraacetic acid
|
|
PBMCs
|
peripheral blood mononuclear cells
|
|
ELISA
|
enzyme-linked immunosorbent assay
|
|
PHA
|
phytohemagglutinin
|
|
HLA
|
human leukocyte antigen
|
|
IFN-γ
|
interferon-γ
|
References
|
1
|
Prockop DJ: Marrow stromal cels as stem
cells for nonhematopoietic tissues. Science. 276:71–74. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Pittenger MF, Mackay AM, Beck SC, Jaiswal
RK, Douglas R, Mosca JD, Moorman MA, Simonetti DW, Craig S and
Marshak DR: Multilineage potential of adult human mesenchymal stem
cells. Science. 284:143–147. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Pittenger MF and Martin BJ: Mesenchymal
stem cells and their potential as cardiac therapeutics. Circ Res.
95:9–20. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Satake K, Lou J and Lenke LG: Migration of
mesenchymal stem cells through cerebrospinal fluid into injured
spinal cord tissue. Spine (Phila Pa 1976). 29:1971–1979. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Satija NK, Singh VK, Verma YK, Gupta P,
Sharma S, Afrin F, Sharma M, Sharma P, Tripathi RP and Gurudutta
GU: Mesenchymal stem cell-based therapy: A new paradigm in
regenerative medicine. J Cell Mol Med. 13:4385–4402. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Taran R, Mamidi MK, Singh G, Dutta S,
Parhar IS, John JP, Bhonde R, Pal R and Das AK: In vitro and in
vivo neurogenic potential of mesenchymal stem cells isolated from
different sources. J Biosci. 39:157–169. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ma J, Both SK, Yang F, Cui FZ, Pan J,
Meijer GJ, Jansen JA and van den Beucken JJ: Concise review:
Cell-based strategies in bone tissue engineering and regenerative
medicine. Stem Cells Transl Med. 3:98–107. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Shang Q, Wang Z, Liu W, Shi Y, Cui L and
Cao Y: Tissue-engineered bone repair of sheep cranial defects with
autologous bone marrow stromal cells. J Craniofac Surg. 12:586–595.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ferrari G, Cusella-De Angelis G, Coletta
M, Paolucci E, Stornaiuolo A, Cossu G and Mavilio F: Muscle
regeneration by bone marrow-derived myogenic progenitors. Science.
279:1528–1530. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Shintani S, Murohara T, Ikeda H, Ueno T,
Honma T, Katoh A, Sasaki K, Shimada T, Oike Y and Imaizumi T:
Mobilization of endothelial progenitor cells in patients with acute
myocardial infarction. Circulation. 103:2776–2779. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ringdén O, Uzunel M, Rasmusson I,
Remberger M, Sundberg B, Lönnies H, Marschall HU, Dlugosz A, Szakos
A, Hassan Z, et al: Mesenchymal stem cells for treatment of
therapy-resistant graft-versus-host disease. Transplantation.
81:1390–1397. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Angelopoulou M, Novelli E, Grove JE,
Rinder HM, Civin C, Cheng L and Krause DS: Cotransplantation of
human mesenchymal stem cells enhances human myelopoiesis and
megakaryocytopoiesis in NOD/SCID mice. Exp Hematol. 31:413–420.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Miura M, Gronthos S, Zhao M, Lu B, Fisher
LW, Robey PG and Shi S: SHED: Stem cells from human exfoliated
deciduous teeth. Proc Natl Acad Sci USA. 100:5807–5812. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ren H, Sang Y, Zhang F, Liu Z, Qi N and
Chen Y: Comparative analysis of human mesenchymal stem cells from
umbilical cord, dental pulp, and menstrual blood as sources for
cell therapy. Stem Cells Int. 2016:35165742016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ren HY, Zhao QJ, Xing W, Yang SG, Lu SH,
Ren Q, Zhang L and Han ZC: Differentiation of human umbilical cord
derived mesenchymal stem cells into low immunogenic and functional
hepatocyte-like cells in vitro. Zhongguo Yi Xue Ke Xue Yuan Xue
Bao. 32:190–194. 2010.(In Chinese). PubMed/NCBI
|
|
16
|
Zhang Q, Shi S, Liu Y, Uyanne J, Shi Y,
Shi S and Le AD: Mesenchymal stem cells derived from human gingiva
are capable of immunomodulatory functions and ameliorate
inflammation-related tissue destruction in experimental colitis. J
Immunol. 183:7787–7798. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Tomar GB, Srivastava RK, Gupta N,
Barhanpurkar AP, Pote ST, Jhaveri HM, Mishra GC and Wani MR: Human
gingiva-derived mesenchymal stem cells are superior to bone
marrow-derived mesenchymal stem cells for cell therapy in
regenerative medicine. Biochem Biophys Res Commun. 393:377–383.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang F, Yu M, Yan X, Wen Y, Zeng Q, Yue W,
Yang P and Pei X: Gingiva-derived mesenchymal stem cell-mediated
therapeutic approach for bone tissue regeneration. Stem Cells Dev.
20:2093–2102. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Jacobs SA, Roobrouck VD, Verfaillie CM and
Van Gool SW: Immunological characteristics of human mesenchymal
stem cells and multipotent adult progenitor cells. Immunol Cell
Biol. 91:32–39. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Deans RJ and Moseley AB: Mesenchymal stem
cells: Biology and potential clinical uses. Exp Hematol.
28:875–884. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Inoue S, Popp FC, Koehl GE, Piso P,
Schlitt HJ, Geissler EK and Dahlke MH: Immunomodulatory effects of
mesenchymal stem cells in a rat organ transplant model.
Transplantation. 81:1589–1595. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yamaza T, Kentaro A, Chen C, Liu Y, Shi Y,
Gronthos S, Wang S and Shi S: Immunomodulatory properties of stem
cells from human exfoliated deciduous teeth. Stem Cell Res Ther.
1:52010. View
Article : Google Scholar : PubMed/NCBI
|
|
23
|
Li P, Zhao Y and Ge L: Therapeutic effects
of human gingiva-derived mesenchymal stromal cells on murine
contact hypersensitivity via prostaglandin E2-EP3 signaling. Stem
Cell Res Ther. 7:1032016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Fu X, Chen Y, Xie FN, Dong P, Liu WB, Cao
Y, Zhang WJ and Xiao R: Comparison of immunological characteristics
of mesenchymal stem cells derived from human embryonic stem cells
and bone marrow. Tissue Eng Part A. 21:616–626. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang M, Yang Y, Yang D, Luo F, Liang W,
Guo S and Xu J: The immunomodulatory activity of human umbilical
cord blood-derived mesenchymal stem cells in vitro. Immunology.
126:220–232. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
de Witte SFH, Lambert EE, Merino A, Strini
T, Douben HJCW, O'Flynn L, Elliman SJ, de Klein AJEMM, Newsome PN,
Baan CC and Hoogduijn MJ: Aging of bone marrow- and umbilical
cord-derived mesenchymal stromal cells during expansion.
Cytotherapy. 19:798–807. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Luo W, Li S, Peng B, Ye Y, Deng X and Yao
K: Embryonic stem cells markers SOX2, OCT4 and Nanog expression and
their correlations with epithelial-mesenchymal transition in
nasopharyngeal carcinoma. PLoS One. 8:e563242013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Isobe Y, Koyama N, Nakao K, Osawa K, Ikeno
M, Yamanaka S, Okubo Y, Fujimura K and Bessho K: Comparison of
human mesenchymal stem cells derived from bone marrow, synovial
fluid, adult dental pulp, and exfoliated deciduous tooth pulp. Int
J Oral Maxillofac Surg. 45:124–131. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Nakashima M, Iohara K, Murakami M,
Nakamura H, Sato Y, Ariji Y and Matsushita K: Pulp regeneration by
transplantation of dental pulp stem cells in pulpitis: A pilot
clinical study. Stem Cell Res Ther. 8:612017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Bartholomew A, Sturgeon C, Siatskas M,
Ferrer K, McIntosh K, Patil S, Hardy W, Devine S, Ucker D, Deans R,
et al: Mesenchymal stem cells suppress lymphocyte proliferation in
vitro and prolong skin graft survival in vivo. Exp Hematol.
30:42–48. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Le Blanc K, Rasmusson I, Sundberg B,
Götherström C, Hassan M, Uzunel M and Ringdén O: Treatment of
severe acute graft-versus-host disease with third party
haploidentical mesenchymal stem cells. Lancet. 363:1439–1441. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Sun L, Akiyama K, Zhang H, Yamaza T, Hou
Y, Zhao S, Xu T, Le A and Shi S: Mesenchymal stem cell
transplantation reverses multiorgan dysfunction in systemic lupus
erythematosus mice and humans. Stem Cells. 27:1421–1432. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Sheng H, Wang Y, Jin Y, Zhang Q, Zhang Y,
Wang L, Shen B, Yin S, Liu W, Cui L and Li N: A critical role of
IFNgamma in priming MSC-mediated suppression of T cell
proliferation through up-regulation of B7-H1. Cell Res. 18:846–857.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Ryan JM, Barry FP, Murphy JM and Mahon BP:
Mesenchymal stem cells avoid allogeneic rejection. J Inflamm
(Lond). 2:82005. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Bocelli-Tyndall C, Zajac P, Di Maggio N,
Trella E, Benvenuto F, Iezzi G, Scherberich A, Barbero A, Schaeren
S, Pistoia V, et al: Fibroblast growth factor 2 and
platelet-derived growth factor, but not platelet lysate, induce
proliferation-dependent, functional class II major
histocompatibility complex antigen in human mesenchymal stem cells.
Arthritis Rheum. 62:3815–3825. 2010. View Article : Google Scholar : PubMed/NCBI
|