Introduction
There is growing concern regarding the safety of
anesthetics on the developing brain. Previous studies demonstrated
that early post-natal anesthesia has neurotoxic effects in rodents
and other animals (1,2). Anesthesia of children aged <3
years old in surgery increases the risk of learning and memory
impairment (3). The use of
ketamine is well established in pediatric anesthesia and it is used
for induction and maintenance of anesthesia (4,5). An
increasing number of studies demonstrated that ketamine causes
anesthetic neurotoxicity. Previous studies in rodent and primate
models have confirmed that ketamine anesthesia during critical
periods of brain development may result in functional deficits
(6,7).
Dendritic spines are the anatomical locus of the
majority of excitatory synapses in the central nervous system
(8). Dendritic spines arise as
small protrusions from the dendritic shaft of various types of
neuron and receive inputs from excitatory axons (9). Alterations in the morphology and
number of dendritic spines are associated with alterations in
synaptic efficacy, learning and memory, and other cognitive
processes (10,11). Previous studies demonstrated that
ketamine causes learning and behavioral impairment, primarily due
to a reduction in synaptic structural plasticity (12,13).
The present study investigated the underlying mechanism by which
ketamine affects synaptogenesis in early developing hippocampal
neurons of rats.
Transforming protein RhoA (RhoA) guanosine
triphosphate hydrolases (GTPases) and their downstream signals are
critical in the association of neuronal activity with various forms
of neuronal plasticity (14,15).
Volatile anesthetics lead to neuronal apoptosis and
length-shortening of dendritic protrusions by activating RhoA in
developing neurons (16,17). In the present study, it was
hypothesized that ketamine may alter the number and length of
dendritic spines in early developing hippocampal neurons via
activation of the RhoA/Rho-associated kinase (ROCK) signaling
pathway.
Materials and methods
Ethics statement
The Institutional Animal Care and Use Committee of
Hebei Medical University (Shijiazhuang, China) specifically
approved the present study. All of the animals were handled
according to this approved protocol.
Animals
A total of eight 24 h postnatal Sprague-Dawley rats,
weighing 4.5~6.5 g, were provided by the Laboratory Animal Center
of Hebei Medical University (Hebei, China). Animals were kept in a
40–60% humidity and temperature-controlled (22–23°C) room under a
12-h light/dark cycle (light on at 7:00 AM); standard rat chow and
water were available ad libitum. Housing was provided with
appropriate tactile, olfactory, visual, and auditory stimuli.
Primary neuron cultures
The rats were quickly decapitated and their brains
were aseptically dissected and immersed in ice-cold dissecting
solution, consisting of Hanks' Balanced Salt Solution. Bilateral
hippocampal tissues were exposed and removed from the 24-h
post-natal Sprague-Dawley rats and subsequently disinfected with
75% alcohol. Subsequent to the removal of blood vessels and
meninges, the hippocampal tissues were cut into pieces that were as
small as possible. These pieces were centrifuged for 100 × g and 5
min at room temperature. Subsequent to discarding the supernatant,
Accutase® (Thermo Fisher Scientific, Inc., Waltham, MA,
USA) was added to the hippocampal tissues at a volume of 3:1. The
samples were placed in an incubator for 10–20 min for digestion.
The tissues were vortexed numerous times during the digestion and
Dulbecco's modified Eagle's medium (DMEM) with 10% fetal bovine
serum (FBS; Gibco; Thermo Fisher Scientific, Inc.) was added to
terminate the digestion process. The tissues were thoroughly
dispersed by pipetting and subsequently filtered through a 200-mesh
copper filter. The filtered cells were collected and centrifuged at
100 × g for 5 min at room temperature. Subsequently, the
supernatant was discarded, DMEM with 10% FBS was added, and the
cells were dispersed in order to obtain a cell suspension. Cells
(1×106 cells/well) were plated in 6-well plates for 24
h. The culture medium was replaced with neurobasal medium
(Invitrogen; Thermo Fisher Scientific, Inc.) containing 1% N2 and
2% B27. Half of the culture medium was changed every two days.
Cells were cultured at 37°C in a 5% CO2 incubator.
Immunofluorescence procedures and
purity identification
Cells were fixed with 4% formaldehyde for at room
temperature for 30 min. The cells were then incubated with 5% goat
serum (Novus Biologicals, LLC, Littleton, CO, USA) at room
temperature for 10 min and immunostained with
microtubule-associated protein 2 (MAP2; cat. no. 554002, 1:50; BD
Biosciences, Franklin Lakes, NJ, USA) antibody at 4°C overnight,
followed by incubation with goat anti-mouse immunoglobulin G
secondary antibody (cat. no. NBP2-44080, 1:50, Novus Biologicals,
Colorado, USA) at room temperature for 45 min. The nuclei were
stained with DAPI (Enzo Life Sciences, Inc., Farmingdale, NY, USA)
at room temperature for 30 min. Images of each slide were captured
using a Zeiss Axiovert Upright Fluorescence microscope
(magnification, ×40 and ×200; Zeiss AG, Oberkochen, Germany) and
fluorescent intensity was quantified using Image Pro Plus version
6.0 (Media Cybernetics, Inc., Rockville, MD, USA). The purity of
neurons (%) was calculated as follows: (Number of MAP2 positive
cells/number of DAPI positive cells) ×100.
Cell treatment with reagents
After culturing for 5 days, cells were exposed to
control (PBS), ketamine (100, 300 and 500 µM), 10 µM Y27632
(ScienCell Research Laboratories, Inc., San Diego, CA, USA) or
ketamine and Y27632 at 5 days in vitro (DIV). After
treatment for 6 h, cells were washed with PBS and subsequently
incubated for 5 min at 37°C with Vybrant™ CM-Dil
Cell-Labeling solution (cat. no. V22888; Thermo Fisher Scientific,
Inc.). Cells were fixed with 4% formaldehyde at room temperature
for 30 min. Following rinsing, slides were mounted for imaging.
Fluorescence images were captured using confocal microscopy
(magnification, ×400 and ×1,000; Nikon Corporation, Tokyo, Japan).
Vybrant™ CM-Dil Cell-Labeling solution-stained fluorescence images
appeared in red or, following transformation using Image-J version
2.0 software (National Institutes of Health, Bethesda, MD, USA), in
black and white.
Analysis of spine density
All dendrite and spine tracing was performed by an
investigator, who was blind to the experimental treatment group.
Fluorescence images were captured using confocal microscopy
(magnification, ×400 and ×1,000; Nikon Corporation). Analysis of
spine plasticity was performed using ImageJ software, as previously
described (17). Dendrite segments
located distal to the first branch point from the cell soma and
including the most distally-located dendritic segments (2–5 per
neuron) were randomly selected for analysis. For each examined
neuron, one 100-µm long segment from a second- or third-order
dendrite (protruding from its parent apical dendrite) was selected
for spine density quantification (18). For spine length quantification,
length was determined as the distance between the base and top of
the spine head.
Western blot analysis
Cells were harvested 6 h after treatment with
reagents and lysed in a buffer containing 50 mM Tris-HCl (pH 7.5),
150 mM NaCl, 2 mM EDTA, 1% Triton, 1 mM PMSF and Protease Inhibitor
Cocktail (1:50; Beyotime Institute of Biotechnology, Shanghai,
China) for 20 min on ice. Lysates were separated by centrifugation
at 15,000 × g at 4°C for 20 min. Supernatants were collected and
the protein concentrations were determined by a Bradford assay.
Proteins (150 µg) were subjected to electrophoresis on 12% SDS-PAGE
and were subsequently transferred to polyvinylidene difluoride
membranes. Membranes were blocked in Tris-buffered saline (pH 7.4)
with 5% non-fat dry milk at room temperature for 1 h, and probed
with primary antibodies at 4°C overnight. The following antibodies
were used: RhoA (cat. no. ab187027; 1:500), ROCK (cat. no. ab71598;
1:500) and β-actin (cat. no. ab8226; 1:500; Abcam, Cambridge, UK).
Then, the cells were washed with TBST three times for 10 min each
and then incubated with peroxidase-labeled affinity purified
antibody to Rabbit IgG (cat. no. ZDR-5306; OriGene Technologies,
Inc., Beijing, China) or peroxidase-labeled affinity purified
antibody to Mouse IgG (cat. no. ZDR-5307; OriGene Technologies,
Inc., Beijing, China) at room temperature for 2 h. Blots were
detected using an enhanced chemiluminescence plus system (MiniChemi
610 Plus; 304002L; Beijing Sage Creation Science Co., Ltd.,
Beijing, China). Quantification (sageCapture3.0, Service For Life
Science) of the western blots was conducted using sageCapture
version 3.0 (Beijing Sage Creation Science Co., Ltd.,).
Statistical analysis
All experiments were repeated three times.
Statistical analysis was performed using GraphPad Prism version 5.0
software (GraphPad Software, Inc., La Jolla, CA, USA). All data are
presented as the mean ± standard error of the mean. The density and
mean length of dendritic spines were analyzed with one-way analysis
of variance. Post hoc individual means comparisons were performed
using the LSD-t-test. P<0.05 was considered to indicate a
statistically significant difference.
Results
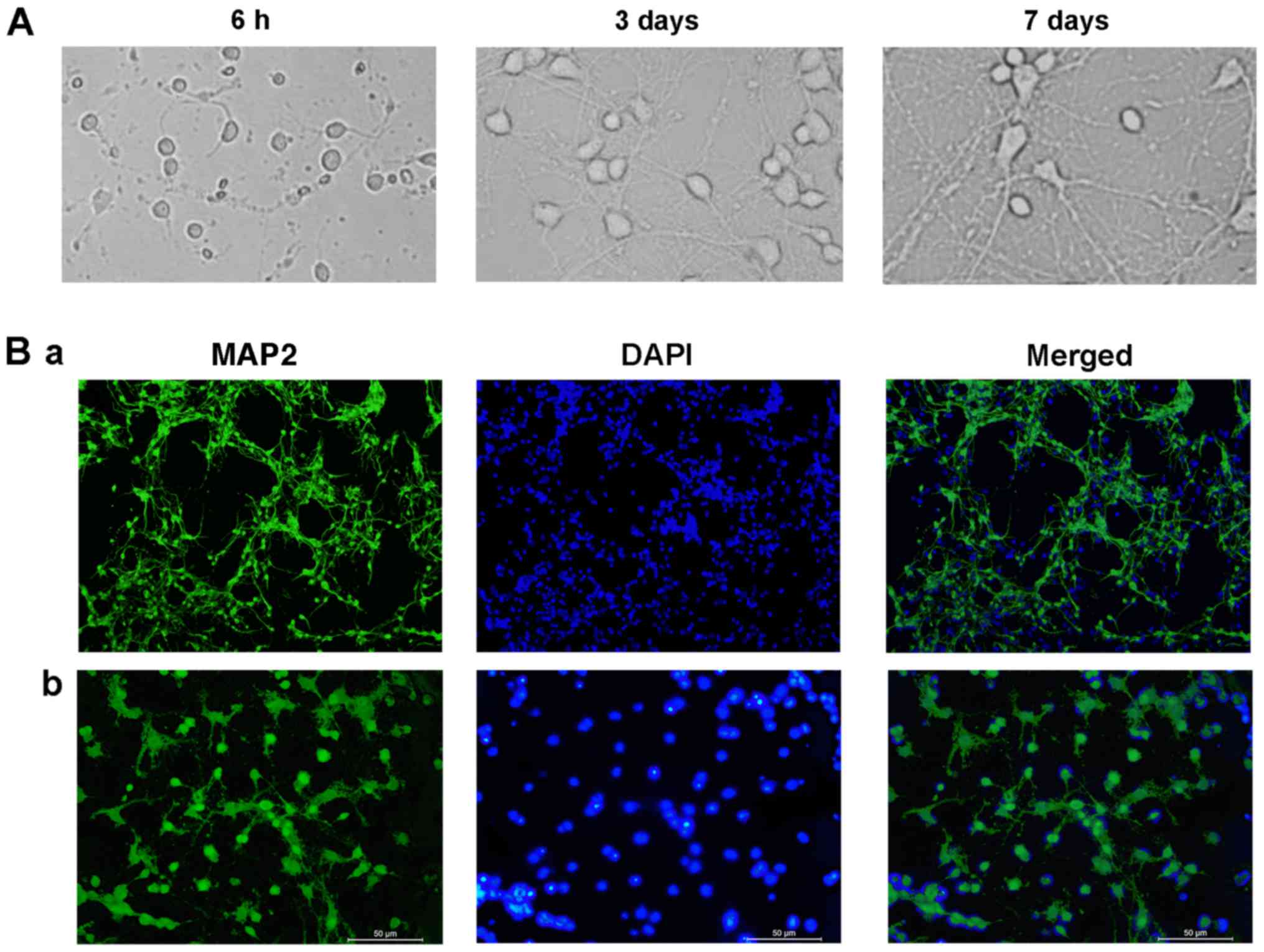
Morphology and purity of hippocampal
neurons
Cells cultured in vitro adhered to the
surface and demonstrated a spindle shape. At 3 DIV, the neurons
were typically spindle-shaped and developed into a sparse network
that primarily formed an in vitro neural network. After 7
DIV, the network and effective synaptic connections formed by axons
and dendrites were enhanced (Fig.
1A). The purity of hippocampal neurons was subsequently
determined. The microtubule-associated protein MAP2 preferentially
localizes to dendrites; however, not to axons, and is a useful
marker for dendrites in cell culture (19). In total, 91% of the cells were
identified as neurons, demonstrated by MAP2 staining (Fig. 1B).
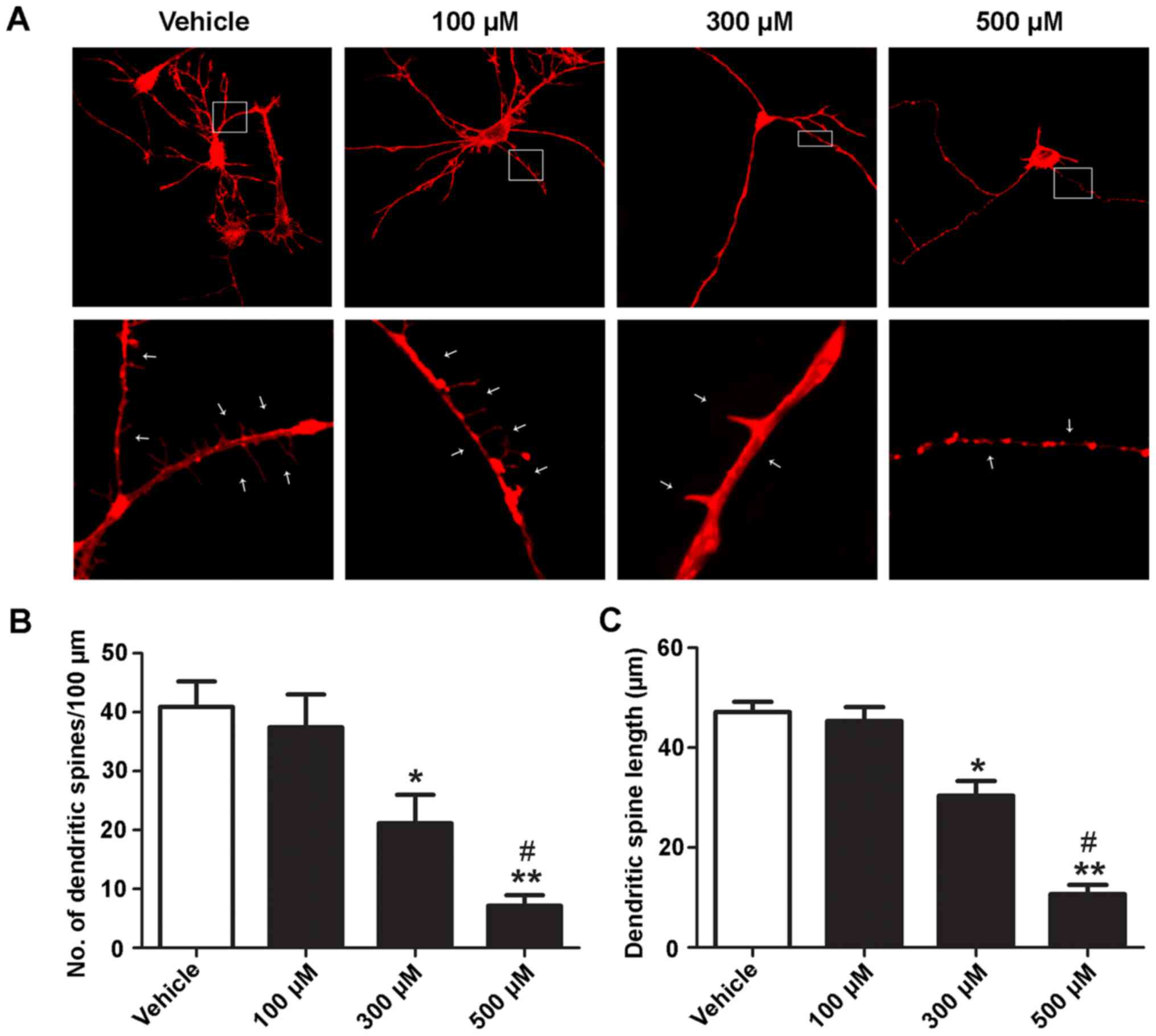
Ketamine decreases the density and
length of dendritic spines in a dose-dependent manner
The effects of ketamine on the length and density of
dendritic spines were assessed. At 5 DIV, hippocampal neurons were
treated with different concentrations (100, 300 and 500 µM) of
ketamine for 6 h. Representative images of dendritic spines in
vehicle (unexposed) or ketamine-exposed neurons are presented in
Fig. 2A. Ketamine caused a
dose-dependent decrease in the number and length of dendritic
spines (Fig. 2B and C). There was
no significant difference between the 100 µM ketamine-exposure
group (number, 36.47±8.23; length, 45.87±5.01 µm) and control cells
(number, 40.88±7.61; length, 47.69±4.28 µm). Concentrations of 300
and 500 µM ketamine significantly decreased the density and length
of dendritic spines (number, 20.31±5.87, length, 30.71±5.52 µm; and
number, 7.23±2.01; length, 10.68±3.53 µm, respectively; P<0.05)
compared with the control. Furthermore, 500 µM ketamine
significantly decreased the number and length of dendritic spines
compared with 300 µM ketamine (P<0.05). These results suggested
that ketamine decreases the density and length of dendritic spines
in a dose-dependent manner.
ROCK inhibitor attenuates
ketamine-induced dendritic spine loss and length-shortening
During early development, activation of the
RhoA-ROCK pathway is involved in immature dendritic protrusions on
hippocampal neurons (20).
Therefore, the role of the RhoA-ROCK pathway in ketamine-induced
dendritic spine loss and length-shortening was investigated in the
present study. Neurons treated with ketamine for 5 DIV were
incubated in the presence of 10 µM Y27632, a specific inhibitor of
ROCK (21). Ketamine upregulated
RhoA and ROCK protein expression levels, whereas, Y27632 attenuated
the expression of ROCK (Fig. 3A).
The number and length of dendritic spines differed among the four
groups. Neurons treated with ketamine had fewer (15.57±6.23) and
shorter length (20.01±3.87 µm) dendritic spines compared with the
control neurons (number, 40.65±5.78; length, 50.78±6.38 µm).
Treatment with Y27632 in the presence of ketamine significantly
prevented dendritic spine loss and length-shortening (number,
28.67±5.83; length, 34.89±4.87 µm). Compared with the control
group, the neurons treated with Y27632 demonstrated no significant
difference in number or length of dendritic spines (Fig. 3B-D).
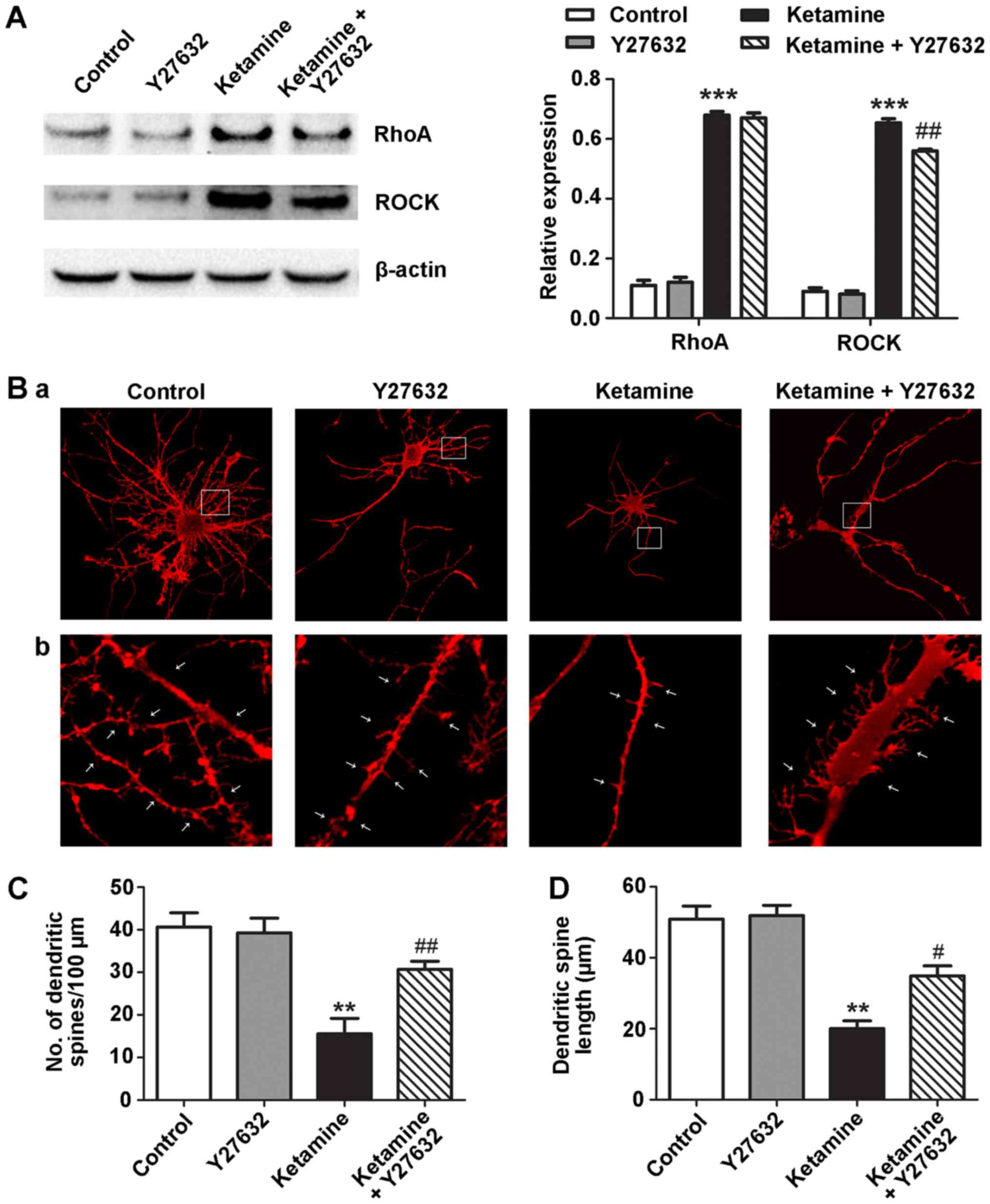 | Figure 3.ROCK Y27632 prevents ketamine-induced
dendritic spine loss and shortening in 5 DIV hippocampal neurons.
The 5 DIV rat hippocampal neurons were exposed to Y27632 (10 µM),
ketamine (300 µM) or ketamine (300 µM) and Y27632 (10 µM) for 6 h.
(A) Ketamine (300 µM) induced upregulation of RhoA and ROCK
compared with the control group. ROCK protein expression levels
were attenuated by co-incubating ketamine-exposed neurons with
Y27632. (B) Representative dendritic arbor development and spine
images of neurons treated with control, Y27632, ketamine or
ketamine plus Y27632. White arrows indicate dendritic spines. (C)
Number of dendritic spines per 100 µm. (D) Dendritic spine length.
n=10. Magnification, ×400 and ×1,000. **P<0.01, ***P<0.001
vs. control; #P<0.05, ##P<0.01 vs.
ketamine. RhoA, transforming protein RhoA; ROCK, Rho-associated
kinase; DIV, days in vitro. |
Discussion
The results of the present study suggested that
early exposure of rat hippocampal neurons to ketamine substantially
decreases the density and length of dendritic spines. The selective
ROCK inhibitor Y27632 partly attenuates the effects induced by
ketamine, suggesting that the RhoA/ROCK signaling pathway is
involved in ketamine-induced neurotoxicity in the developing
brain.
A previous study demonstrated that in rats the blood
concentration of ketamine was ~20 µM and the brain plasma ratio of
ketamine was 6.5:1 (22), thus,
100–300 µM ketamine is clinically relevant (blood concentration of
ketamine was ~20 µM and the brain plasma ratio of ketamine was
6.5:1, (20×6.5=130 µM). A high concentration of ketamine has been
widely used in a large number of in vitro experiments to
investigate its mechanisms of neurotoxicity (23–25).
In the present study, concentrations of ketamine ranging between
100 and 500 µM were selected.
Previous studies demonstrated that the impact of
ketamine on dendritic spines is dependent on developmental age.
Exposure to ketamine decreases the density of synapses or spines in
the hippocampus of neonatal rodents at 5–13 days post-natal
(12). Treatment of mouse
hippocampal neurons with ketamine for 5 h at 15 and 30 days
post-natal caused an age-dependent significant increase in
dendritic spine density (26).
These previous studies identified a mechanism for the modulation of
synaptogenesis via modulation of the excitation/inhibition balance
by ketamine. In the present study, rats were sacrificed 24 h
post-natal. At 5 DIV, neurons treated with ketamine for 6 h
exhibited a decrease in dendritic spine density and mean length in
a dose-dependent manner, which is consistent with the results of a
previous study (12). This
suggested that ketamine impairs the morphology of dendritic spines
during early post-natal development.
The small GTPase Rho is critical in regulating actin
remodeling in dendritic spines (27). RhoA regulates maturation of
adhesions and the postsynaptic density of dendritic spines
(28). Activation of RhoA/ROCK
signaling leads to dendritic spine shrinkage and synapse loss
(29). Conversely, blocking
activation of ROCK induces novel, long spines and protrusive
motility (30). In the present
study, ketamine induced upregulation of RhoA and ROCK protein
expression levels. Furthermore, Y27632, an inhibitor of ROCK,
attenuated dendritic spine loss and length-shortening, suggesting
that the mechanism of ketamine anesthetic neurotoxicity is partly
via activation of the RhoA/ROCK signaling pathway. A previous study
demonstrated that the anesthetic agent isoflurane destabilizes
spine actin, which results in alterations to dendritic spine
morphology and number (31).
Zimering et al (17)
additionally observed that sevoflurane causes actin
depolymerization, leading to reversible length-shortening of
dendritic protrusions through a mechanism involving activation of
RhoA/ROCK signaling. Ketamine is commonly used as an intravenous
general anesthetic for pediatric patients. These results are
consistent with the possibility that activation of the RhoA/ROCK
signaling pathway is one of the mechanisms of the neurotoxicity of
ketamine when used as an anesthetic.
The ROCK inhibitor Y27632 has been demonstrated to
strongly stimulate neurite outgrowth (32). A previous study identified that
treatment of 13 DIV hippocampal neurons with 100 µM Y27632 for 3
days increased the density and length of dendritic spines of
cultured neurons (33). Swanger
et al (21) demonstrated
that exposure of 16–17 DIV neurons to 10 µM Y27632 altered the
morphology of dendritic spines. In the present study, an increase
in number and length of dendritic spines was not observed following
treatment of 5 DIV neurons with 10 µM Y27632 for 6 h, which may be
due to the relatively fewer developing neurons or the low
concentration of Y27632. Y27632 in the presence of ketamine
attenuated the dendritic spine loss and length-shortening induced
by ketamine. Therefore, it was hypothesized that in addition to the
RhoA/ROCK signaling pathway, there are other molecular mechanisms
involved in the synaptic structural plasticity induced by
ketamine.
Previous studies demonstrated that there are a
number of possible mechanisms by which ketamine induces
neurotoxicity in the developing brain, including suppression of
phosphorylated (p)-protein kinase Cγ and p-extracellular
signal-regulated kinase (34),
upregulation of neuronal microRNA 124 (35), inhibition of the frequency and
amplitude of calcium oscillations (36), decrease in glycogen synthase
kinase-3β phosphorylation (37),
and activation of the phosphoinositide 3-kinase/protein kinase B
signaling pathways. However, there are fewer studies focused on the
effect of ketamine on hippocampal dendritic spines. The present
study has identified another mechanism whereby ketamine affects the
plasticity of hippocampal dendritic spines and may contribute to
neurotoxicity in the developing brain.
The present study has a number of limitations.
Exposure to anesthetics transiently affects the dynamics of
dendritic spines, and dendritic protrusion loss and shrinkage are
reversible (17,31,38).
The present study was based on single time-point observations; it
is not known whether ketamine-induced effects on the morphology and
number of dendritic spines may be attenuated upon ketamine
elimination. Repeated exposure to ketamine during early development
impairs motor learning and learning-dependent plasticity of
dendritic spines later in life (13). The in vitro model used in
the present study only demonstrated that ketamine decreases the
number and length of dendritic spines. Further experiments on
animals are required to investigate the possible effects of
ketamine on behavior.
In summary, the present study demonstrated that
exposure to ketamine during the period of rapid synaptogenesis
causes loss and length-shortening of dendritic spines in cultured
rat hippocampal neurons. Inhibition of ROCK by Y27632 attenuates
the effects of ketamine on dendritic spines. The present novel data
suggested that ketamine affects the formation of synapses partly
through the RhoA/ROCK signaling pathway. Therefore, further studies
on the RhoA/ROCK signaling pathways in ketamine-induced
neurotoxicity in children are required.
Acknowledgements
The authors would like to thank Ms. Hanying Xing of
the Hebei General Hospital (Shijiazhuang, China) for her technical
assistance.
Funding
The present study was supported by the Natural
Science Foundation of Hebei Province (grant no. H2014206454).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
LH and RK conceived and designed the study. Primary
neuron cultures and immunofluorescence procedures were performed by
ZH, RZ and YW. Spine densities were analyzed by XL and LB. XD and
ZH performed the western blot analysis. SJ participated in the
analysis and processing of experiments and data, and wrote the
paper. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
Animal experiments were performed according to the
regulations of laboratory animal management by the Ministry of
Science and Technology of the People's Republic of China (1988) no.
134, which conforms to the internationally recognized National
Institutes of Health (Bethesda, MD, USA) guidance for care and use
of laboratory animals. The Institutional Animal Care and Use
Committee of Hebei Medical University specifically approved the
present study.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Young C, Jevtovic-Todorovic V, Qin YQ,
Tenkova T, Wang H, Labruyere J and Olney JW: Potential of ketamine
and midazolam, individually or in combination, to induce apoptotic
neurodegeneration in the infant mouse brain. Br J Pharmacol.
146:189–197. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lee BH, Chan JT, Hazarika O, Vutskits L
and Sall JW: Early exposure to volatile anesthetics impairs
long-term associative learning and recognition memory. PLoS One.
9:e1053402014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
DiMaggio C, Sun LS and Li G: Early
childhood exposure to anesthesia and risk of developmental and
behavioral disorders in a sibling birth cohort. Anesth Analg.
113:1143–1151. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Green SM, Roback MG, Kennedy RM and Krauss
B: Clinical practice guideline for emergency department ketamine
dissociative sedation: 2011 update. Ann Emerg Med. 57:449–461.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Mistry RB and Nahata MC: Ketamine for
conscious sedation in pediatric emergency care. Pharmacotherapy.
25:1104–1111. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Paule MG, Li M, Allen RR, Liu F, Zou X,
Hotchkiss C, Hanig JP, Patterson TA, Slikker W Jr and Wang C:
Ketamine anesthesia during the first week of life can cause
long-lasting cognitive deficits in rhesus monkeys. Neurotoxicol
Teratol. 33:220–230. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Scallet AC, Schmued LC, Slikker W Jr,
Grunberg N, Faustino PJ, Davis H, Lester D, Pine PS, Sistare F and
Hanig JP: Developmental neurotoxicity of ketamine: Morphometric
confirmation, exposure parameters, and multiple fluorescent
labeling of apoptotic neurons. Toxicol Sci. 81:364–370. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Matus A: Actin-based plasticity in
dendritic spines. Science. 290:754–758. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Rochefort NL and Konnerth A: Dendritic
spines: From structure to in vivo function. EMBO Rep. 13:699–708.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Blanpied TA and Ehlers MD: Microanatomy of
dendritic spines: Emerging principles of synaptic pathology in
psychiatric and neurological disease. Biol Psychiatry.
55:1121–1127. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
McCann RF and Ross DA: A Fragile balance:
Dendritic Spines, Learning, and Memory. Biol Psychiatry.
82:e11–e13. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Tan H, Ren RR, Xiong ZQ and Wang YW:
Effects of ketamine and midazolam on morphology of dendritic spines
in hippocampal CA1 region of neonatal mice. Chin Med J (Engl).
122:455–459. 2009.PubMed/NCBI
|
|
13
|
Huang L and Yang G: Repeated exposure to
ketamine-xylazine during early development impairs motor
learning-dependent dendritic spine plasticity in adulthood.
Anesthesiology. 122:821–831. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Olson MF: Contraction reaction: Mechanical
regulation of Rho GTPase. Trends Cell Biol. 14:111–114. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hedrick NG and Yasuda R: Regulation of Rho
GTPase proteins during spine structural plasticity for the control
of local dendritic plasticity. Curr Opin Neurobiol. 45:193–201.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lemkuil BP, Head BP, Pearn ML, Patel HH,
Drummond JC and Patel PM: Isoflurane neurotoxicity is mediated by
p75NTR-RhoA activation and actin depolymerization. Anesthesiology.
114:49–57. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zimering JH, Dong Y, Fang F, Huang L,
Zhang Y and Xie Z: Anesthetic sevoflurane causes Rho-dependent
filopodial shortening in mouse neurons. PLoS One. 11:e01596372016.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Nagy D, Kocsis K, Fuzik J, Marosi M, Kis
Z, Teichberg VI, Toldi J and Farkas T: Kainate postconditioning
restores LTP in ischemic hippocampal CA1: Onset-dependent second
pathophysiological stress. Neuropharmacology. 61:1026–1032. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Caceres A, Banker G, Steward O, Binder L
and Payne M: MAP2 is localized to the dendrites of hippocampal
neurons which develop in culture. Brain Res. 315:314–318. 1984.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Castaneda P, Munoz M, Garcia-Rojo G, Ulloa
JL, Bravo JA, Márquez R, García-Pérez MA, Arancibia D, Araneda K,
Rojas PS, et al: Association of N-cadherin levels and downstream
effectors of Rho GTPases with dendritic spine loss induced by
chronic stress in rat hippocampal neurons. J Neurosci Res.
93:1476–1491. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Swanger SA, Mattheyses AL, Gentry EG and
Herskowitz JH: ROCK1 and ROCK2 inhibition alters dendritic spine
morphology in hippocampal neurons. Cell Logist. 5:e11332662016.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Cohen ML, Chan SL, Way WL and Trevor AJ:
Distribution in the brain and metabolism of ketamine in the rat
after intravenous administration. Anesthesiology. 39:370–376. 1973.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Giardina SF, Cheung NS, Reid MT and Beart
PM: Kainate-induced apoptosis in cultured murine cerebellar granule
cells elevates expression of the cell cycle gene cyclin D1. J
Neurochem. 71:1325–1328. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Soriano SG, Liu Q, Li J, Liu JR, Han XH,
Kanter JL, Bajic D and Ibla JC: Ketamine activates cell cycle
signaling and apoptosis in the neonatal rat brain. Anesthesiology.
112:1155–1163. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zheng X, Zhou J and Xia Y: The role of
TNF-α in regulating ketamine-induced hippocampal neurotoxicity.
Arch Med Sci. 11:1296–1302. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
De Roo M, Klauser P, Briner A, Nikonenko
I, Mendez P, Dayer A, Kiss JZ, Muller D and Vutskits L: Anesthetics
rapidly promote synaptogenesis during a critical period of brain
development. PLoS One. 4:e70432009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Elia LP, Yamamoto M, Zang K and Reichardt
LF: p120 catenin regulates dendritic spine and synapse development
through Rho-family GTPases and cadherins. Neuron. 51:43–56. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Newell-Litwa KA, Badoual M, Asmussen H,
Patel H, Whitmore L and Horwitz AR: ROCK1 and 2 differentially
regulate actomyosin organization to drive cell and synaptic
polarity. J Cell Biol. 210:225–242. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Pozueta J, Lefort R, Ribe EM, Troy CM,
Arancio O and Shelanski M: Caspase-2 is required for dendritic
spine and behavioural alterations in J20 APP transgenic mice. Nat
Commun. 4:19392013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Tashiro A and Yuste R: Regulation of
dendritic spine motility and stability by Rac1 and Rho kinase:
Evidence for two forms of spine motility. Mol Cell Neurosci.
26:429–440. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Platholi J, Herold KF, Hemmings HC Jr and
Halpain S: Isoflurane reversibly destabilizes hippocampal dendritic
spines by an actin-dependent mechanism. PLoS One. 9:e1029782014.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Darenfed H, Dayanandan B, Zhang T, Hsieh
SH, Fournier AE and Mandato CA: Molecular characterization of the
effects of Y-27632. Cell Motil Cytoskeleton. 64:97–109. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kang MG, Guo Y and Huganir RL: AMPA
receptor and GEF-H1/Lfc complex regulates dendritic spine
development through RhoA signaling cascade. Proc Natl Acad Sci USA.
106:3549–3554. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Huang L, Liu Y, Jin W, Ji X and Dong Z:
Ketamine potentiates hippocampal neurodegeneration and persistent
learning and memory impairment through the PKCgamma-ERK signaling
pathway in the developing brain. Brain Res. 1476:164–171. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Xu H, Zhang J, Zhou W, Feng Y, Teng S and
Song X: The role of miR-124 in modulating hippocampal neurotoxicity
induced by ketamine anesthesia. Int J Neurosci. 125:213–220. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Huang L, Liu Y, Zhang P, Kang R, Liu Y, Li
X, Bo L and Dong Z: In vitro dose-dependent inhibition of the
intracellular spontaneous calcium oscillations in developing
hippocampal neurons by ketamine. PLoS One. 8:e598042013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Liu JR, Baek C, Han XH, Shoureshi P and
Soriano SG: Role of glycogen synthase kinase-3β in ketamine-induced
developmental neuroapoptosis in rats. Br J Anaesth. 110 Suppl
1:i3–i9. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Yang G, Chang PC, Bekker A, Blanck TJ and
Gan WB: Transient effects of anesthetics on dendritic spines and
filopodia in the living mouse cortex. Anesthesiology. 115:718–726.
2011. View Article : Google Scholar : PubMed/NCBI
|