Introduction
The major function of the intestinal epithelial
barrier is to regulate the transport of molecules through
transcellular and paracellular pathways between the hostile
environment of the intestinal epithelium and the sub-epithelial
tissue (1). The intestinal
epithelial barrier has a selective permeable barrier that limits
the permeation of luminal noxious molecules, including toxins,
pathogens and antigens, while regulating the absorption of
nutrients and water. This barrier is established by intercellular
tight junction (TJ) structures (2). TJ assembly involves at least four
types of transmembrane proteins: Tricellulin, claudins, occludin
and junctional adhesion molecules. Claudins are structural proteins
present in TJs and are involved in the regulation of paracellular
movement by forming a barrier across the epithelial monolayer
(3). Altered functions of claudins
are associated with different forms of cancer, inflammatory bowel
diseases and diarrhea (4).
Claudin-1, a major structural and functional TJ protein is
responsible for modulating the permeability of the epidermis
(5).
Myosin light chain kinase (MLCK) is a
calcium-dependent enzyme present in the cytoskeleton and the
cytoplasm (6). MLCK may promote
phosphorylation of the myosin light chain, which regulates the
cytoskeleton (6). The MLCK pathway
mediates intestinal epithelial barrier permeability by regulating
TJs (7). A previous study revealed
that downregulation of MLCK may attenuate damage to TJs (8). The results of the aforementioned
studies suggest that MLCK has an important role in TJ damage under
endotoxemic conditions.
Aquaporins (AQPs), including AQP0-AQP12, are a
family of small integral membrane proteins that transport water
across the cytoplasmic membrane. AQP9 is a subtype of the AQP
protein family that is present in mammalian cells (9), and murine AQP9 protein expression has
been demonstrated in liver, skin, epididymal, epidermal and
neuronal cells (10,11). It has been well established that
AQP9 functions as an aquaglyceroporin protein in the liver,
facilitates hepatic uptake of glycerol (12–14),
and attenuates glaucomatous damage (15). AQP9 was first reported to be
present in the duodenum, jejunum, ileum and colon (16) and a more recent study demonstrated
that AQP9 has important roles in the digestive system (17). Therefore, AQP9 may have an
important role in energy metabolism by functioning as channels in
metabolic pathways.
The present study aimed to determine the association
between AQP9 expression in the intestinal mucosa and conditions in
which TJs are disrupted. This was investigated by comparing the
expression levels of claudin-1 and MLCK; revealing the pathological
features of ileum tissue in the normal control (NC) group, and
acute liver injury groups at 2, 48, 72 h time intervals; and
determining the association between the expression levels of AQP,
Claudin-1 and MLCK.
Materials and methods
Materials
A total of 20 male Sprague Dawley rats (age, 3
months; body weight, 180–240 g), were purchased from the Department
of Animal Science and Technology, Nanchang University (Nanchang,
China). D-galactosamine was purchased from Shanghai Oripharm Co.
Ltd. (Shanghai, China). Antibodies against claudin 1 (cat. no.
4933s; Cell Signaling Technology, Inc., Danvers, MA, USA), MLCK
(cat. no. ab76092), AQP9 (cat. no. ab15129) and β-actin (cat. no.
ab8228; all Abcam, Cambridge, UK) were used. An immunohistochemical
staining kit was purchased from Nanjing KeyGen Biotech Co., Ltd.
(Nanjing, China).
Establishment of an acute liver injury animal model
and preparation of tissue specimens. All animal experiments were
approved by the Animal Ethics Association of the First Affiliated
Hospital of Nanchang University. Rats were housed in a specific
pathogen-free environment at 23±2°C with 45–65% humidity and under
a 12 h light/dark cycle. The animals had free access to food and
water. The rats were randomly divided into two groups following one
week of adaptive feeding: An acute liver injury group (15 rats) and
a NC group (5 rats). Animal models of acute liver injury were
established by one intraperitoneal injection of D-galactosamine
(D-GalN; 1,200 mg/kg). A total of five animals were sacrificed at
24, 48 and 72 h time intervals post-injection. Rats in the NC group
were subjected to intraperitoneal injection of normal saline (12
ml/kg) and then sacrificed at the 72 h time interval. Following
this, serum, liver and ileum tissue specimens from all rats were
collected for subsequent investigation.
Serum biochemical examination
Whole blood samples were centrifuged at a speed of
1,500 × g for 15 min at 4°C to separate the serum. Following this,
levels of alanine aminotransferase (ALT), aspartate
aminotransferase (AST), total bilirubin (Tbil) and albumin (ALB)
were detected in serum samples using an automatic biochemical
analyzer (Beckman Coulter Chemistry Analyzer AU5800 Series; Beckman
Coulter, Inc., Brea, CA, USA).
Pathological examination of liver and
ileum tissues
Liver and ileal tissues (~0.5 g) were collected,
washed with PBS, fixed in 10% neutral formaldehyde solution for 24
h at 4°C and then embedded in paraffin sections. The pathological
alterations in liver and ileum tissue were observed under a light
microscope (BX41; Olympus, Corporation, Tokyo, Japan), following
hematoxylin and eosin staining for 5 min at room temperature. At
least five fields of view were captured and analyzed.
Western blot analysis
The expression levels of MLCK, claudin-1 and AQP9 in
terminal ileum tissues were detected via western blotting. The same
weight (~30 mg) of terminal ileum tissue samples was obtained from
rats belonging to each group and total protein was isolated using
the illustra triplePrep kit (cat. no. 28-9425-44; GE Healthcare
Life Sciences, Little Chalfont, UK). The bicinchoninic acid assay
method (BCA Protein Assay kit; cat. no. P0011; Beyotime Institute
of Biotechnology, Haimen, China) was used for the determination of
protein concentration in the supernatant. Equal amounts of protein
(20 µg) were loaded into lanes of 6 and 12% SDS-PAGE. Once the
proteins had migrated through the gel, the current was terminated
and gels were transferred to polyvinylidene difluoride (PVDF)
membranes in transfer buffer at 18 V for 60 min. PVDF membranes
were then blocked with 5% non-fat dry milk in Tris-buffered saline
containing 0.1% Tween-20 at room temperature for 1 h. Membranes
were incubated overnight at 4°C with primary antibodies against the
following proteins: MLCK (1:250; cat. no. ab76092), AQP9 (1:250;
cat. no. ab15129; both Abcam), claudin-1 (1:250; cat. no. 4933s;
Cell Signaling Technology, Inc.) and β-actin (1:5,000; cat. no.
ab8228; Abcam). Following this, membranes were incubated with
horseradish peroxidase-conjugated goat anti-rabbit secondary
antibodies (1:5,000; cat. no. ab6721; Abcam) for 2 h at room
temperature. Membranes were subsequently washed three times with
Tris-buffered saline containing 0.1% Tween-20 and proteins were
then visualized using Pierce™ ECL Western Blotting Substrate
(Pierce; Thermo Fisher Scientific, Inc., Waltham, MA, USA),
according to the manufacturer's protocol. Each sample was
normalized against β-actin expression. The protein bands were
analyzed for densitometry using Quantity One software version 1.4.6
(Bio-Rad Laboratories, Inc., Hercules, CA, USA).
Immunohistochemistry (IHC)
IHC assays were performed to determine protein
expression levels of claudin-1 and MLCK in terminal ileum tissues
obtained from each group. Tissue sections (4 µm) were fixed with
10% neutral-buffered formalin for 24 h at 4°C and then embedded in
paraffin. Paraffin sections were incubated for 2 h at 67°C and then
subjected to dewaxing, boiling and washing with PBS. Following
this, sections were incubated with rabbit anti-mouse monoclonal
primary antibodies against MLCK (1:250; cat. no. ab76092; Abcam)
for 60 min at room temperature, followed by incubation with
anti-rabbit IgG horseradish peroxidase-linked antibody (1:200; cat.
no. 7074; Cell Signaling Technology, Inc.) at 37°C for 15 min.
Immunohistochemical staining was visualized by incubating sections
with 3,3′-diaminobenzidine chromogen for 3 min at room temperature.
A 50i Nikon Fluorescence Microscope (Nikon Corporation, Tokyo,
Japan) and Adobe Photoshop CS4 software version 11.0 (Adobe
Systems, Inc., San Jose, CA, USA) were used to capture and analyze
the images. Finally, samples were counterstained with 1 mg/ml
hematoxylin for 3 min at room temperature, dehydrated in alcohol
and then mounted. Two investigators blinded to the clinical data,
semi-quantitatively scored the slides by evaluating the staining
intensity and percentage of stained cells in selected areas. At
least five fields of view were captured at magnification ×200. The
staining intensity was scored as 0 (no signal), 1 (weak), 2
(moderate), or 3 (high). The percentage of cells stained was scored
as 1 (1–25%), 2 (26–50%), 3 (51–75%), or 4 (76–100%). A final score
of 0–12 was obtained by multiplying the intensity and percentage
scores (18).
ELISA
Expression levels of secreted tumor necrosis factor
(TNF)-α, interleukin (IL)-6 and IL-10 in the supernatant were
investigated using rat TNF-α (cat. no. ab100785), IL-6 (cat. no.
ab100772) and IL-10 (cat. no. ab100765; all Abcam) ELISA kits
according to the manufacturer's protocol. Absorbance values were
determined using a Synergy HT Microplate Reader (BioTek
Instruments, Inc., Winooski, VT, USA) at 450 nm, and IL-10
concentrations were calculated according to a standard curve and
dilution ratio. All assays were performed in triplicate.
Statistical analysis
Statistical analysis was performed using SPSS
software (version 19.0; IBM Corps., Armonk, NY, USA). Experiments
were repeated six times and all data are expressed as the mean ±
standard error of the mean. For statistical comparison of normal
distributed data, a one-way analysis of variance followed by
Tukey's post hoc test was used. P<0.05 was considered to
indicate a statistically significant difference.
Results
Levels of serum biochemical markers in
the NC and acute liver injury model groups
The results demonstrated that ALT and AST levels
detected in rats in the acute liver injury groups increased in a
time-dependent manner until the 48 h time interval; then were
levels of ALT and AST were decreased at the 72 h time interval. The
levels of Tbil were revealed to increase in a time-dependent
manner, and the results also demonstrated that Alb levels were
significantly reduced at the 72 h time interval (Table I). These results suggested that
rats were suffering from acute liver damage (19). Compared with the NC group,
differences regarding levels of ALT, AST and Tbil in acute liver
injury groups were statistically significant compared with the
(P<0.05; Table I).
 | Table I.Comparative analysis of serum
biochemical index levels in acute liver injury model rats at three
time intervals and NC rats. |
Table I.
Comparative analysis of serum
biochemical index levels in acute liver injury model rats at three
time intervals and NC rats.
| Groups | Rats (n) | ALT (U/l) | AST (U/l) | Tbil (µmol/l) | Alb (g/l) |
|---|
| NC | 5 | 26.67±5.03 | 113.33±7.02 | 0.53±0.21 | 23.33±1.79 |
| M-24 h | 5 |
1,633.53±449.11a |
3,472.67±314.42a |
22.15±6.02a | 23.83±2.78 |
| M-48 h | 5 |
3,886.64±255.62a |
5,013.50±749.38a |
60.47±11.74a | 21.50±1.89 |
| M-72 h | 5 |
935.57±173.05a |
2,230.75±297.13a |
114.45±24.46a |
20.88±2.38a |
Pathological features of liver tissues
obtained from rats in each group
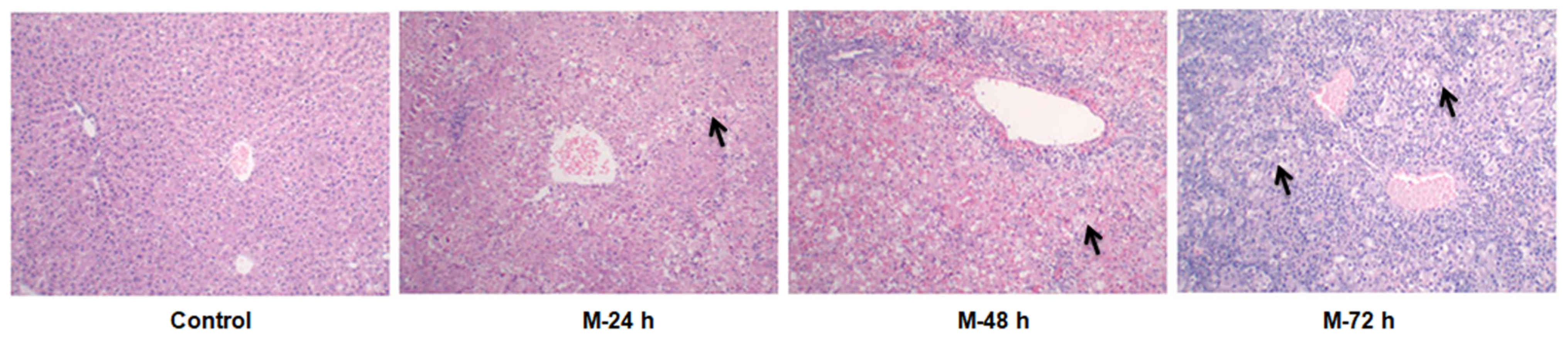
The liver tissues obtained from rats in the NC group
exhibited intact hepatic lobules, with the lobular central vein
presented at the center and the hepatic cord radially arranged, in
the absence of marked levels of inflammation, necrosis and
degeneration (Fig. 1). The normal
structure of liver tissues obtained from rats with acute liver
injury at the 72 h time interval disappeared, with inflammatory
cell infiltration, liver sinus congestion and hemorrhaging, and the
liver cells displayed signs of necrosis (Fig. 1).
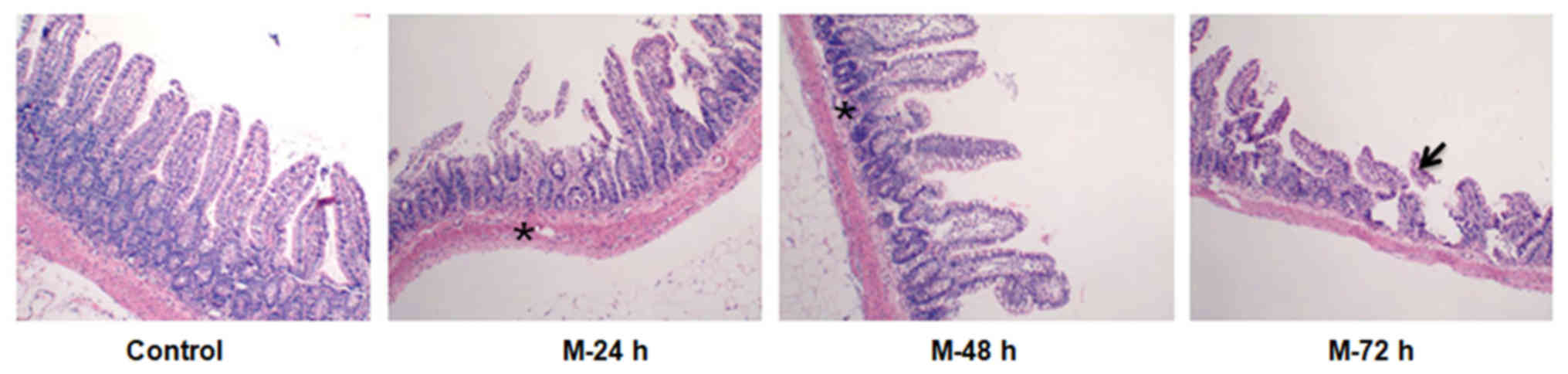
Pathological features of terminal
ileum tissues in NC rats and acute liver injury rats
Complete mucosal structure was demonstrated in
terminal ileum tissues obtained from NC rats, in the absence of
inflammation, necrosis and shedding (Fig. 2). However, terminal ileum tissues
obtained from rats belonging to the acute liver injury groups
exhibited visibly increased levels of neutrophil infiltration in a
time-dependent manner, enhanced levels of interstitial edema in
mucosa and submucosa villi, and, at the 72 h time interval, part of
the tip of the villi was eroded in the terminal ileum mucosa model
24, 48 and 72 h (Fig. 2).
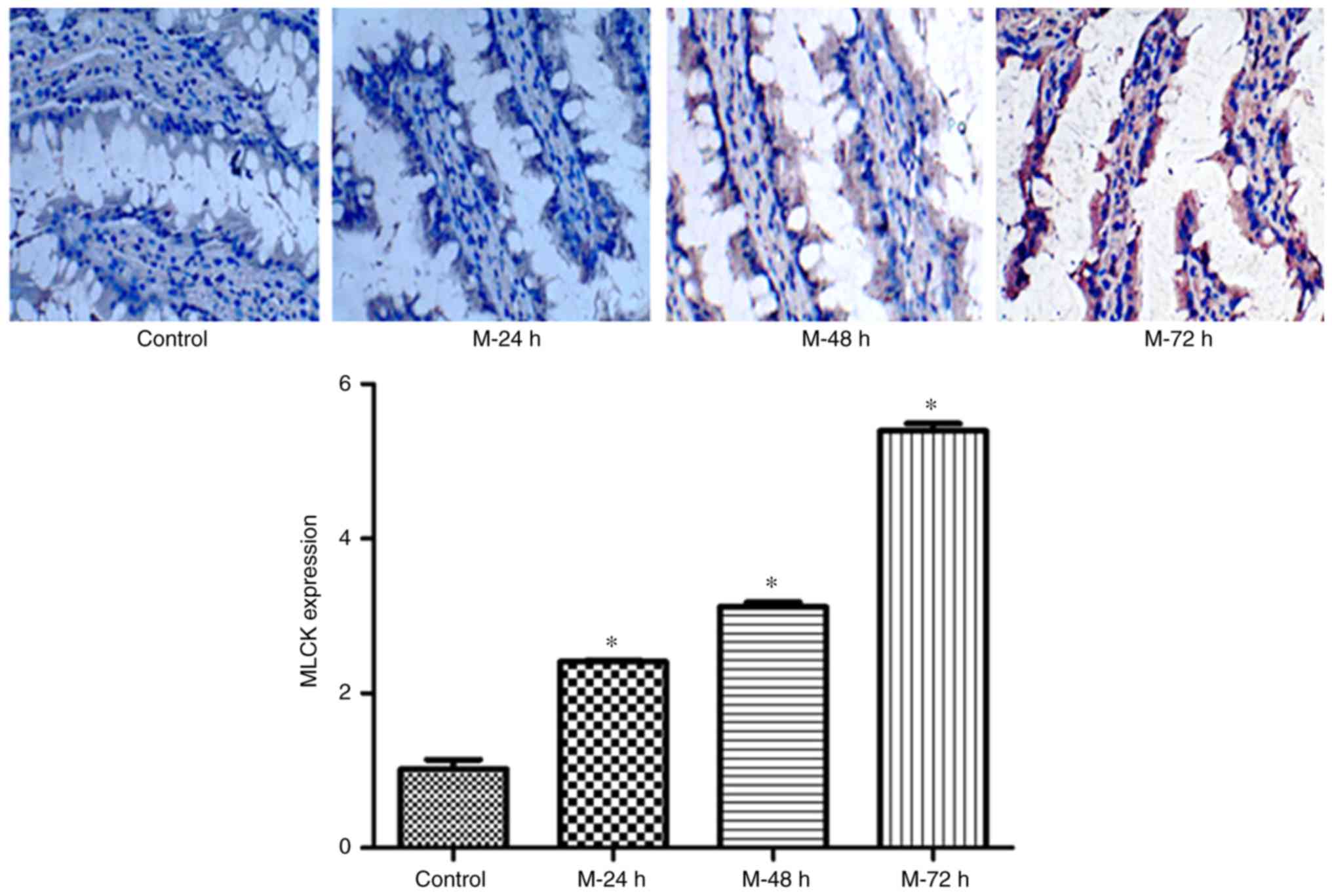
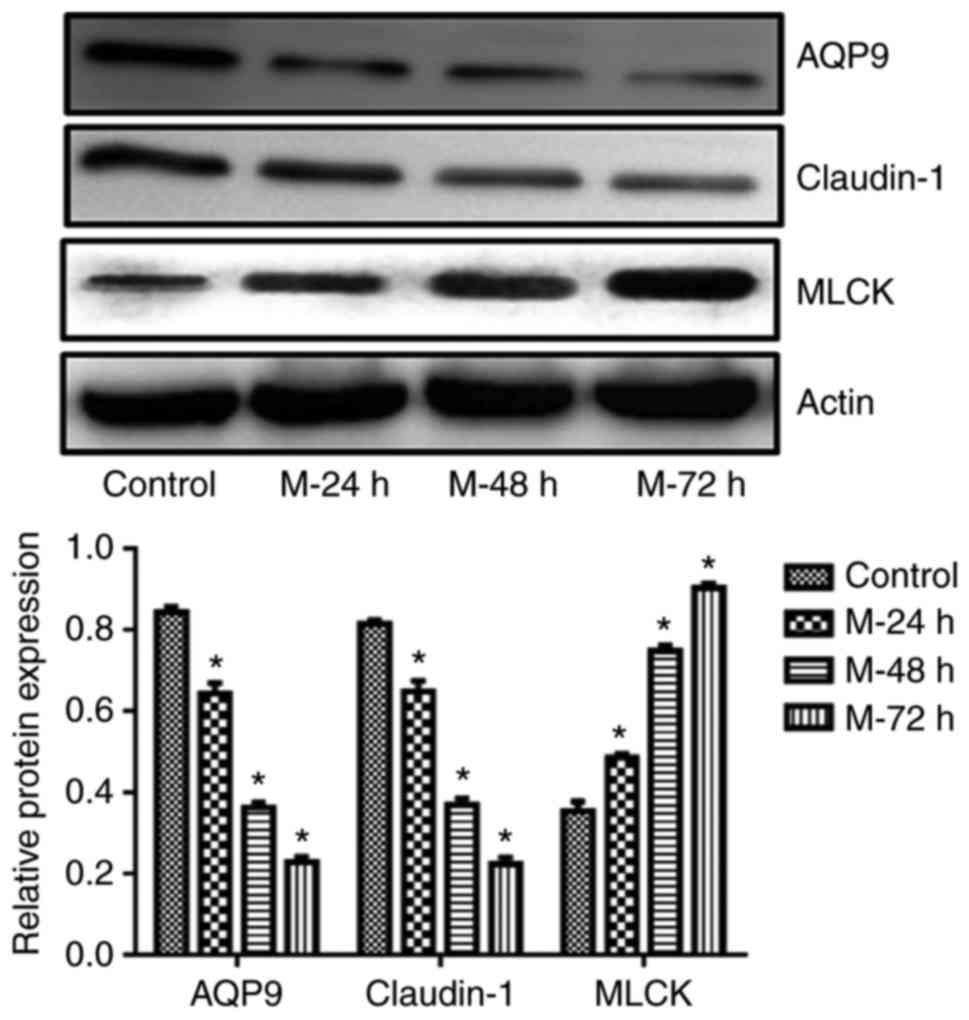
MLCK, AQP9 and claudin-1 expression
levels in terminal ileum tissues obtained from each group as
determined by IHC and western blotting
IHC staining was used to determine MLCK protein
expression levels, and the results demonstrated that in the
sections of ileum obtained from NC rats, slight positive brown
staining revealing MLCK protein expression in the cytoplasm was
observed; however, positive staining in the mucosa of terminal
ileum tissues obtained from acute liver injury rats at 24, 48 and
72 h time intervals was revealed to be significantly increased in a
time-dependent manner (P<0.05; Fig.
3). The greatest protein expression levels of MLCK were
demonstrated in the model 72 h group (Fig. 3). The results of western blotting
revealed that MLCK protein expression levels were significantly
increased compared with the NC group in a time-dependent manner
(Fig. 4).
Western blotting was also used to investigate AQP9
and claudin-1 protein expression levels in terminal ileum tissues
obtained from all rats. The results revealed that both AQP9 and
claudin-1 expression levels were significantly decreased in the
terminal ileum of model groups compared with the NC group in a
time-dependent manner (P<0.05; Fig.
4).
Concentrations of serum cytokines in
NC rats and acute liver injury model rats
Levels of serum TNF-α, IL-6 and IL-10 in each group
were determined using ELISA. The results demonstrated that levels
of serum TNF-α, IL-6 and IL-10 were significantly increased in the
24 h model group compared with the NC group (P<0.05; Table II). The serum levels of TNF-α,
IL-6 and IL-10 were further increased in the 48 h model group.
However, in the 72 h model group, levels of TNF-a and IL-6 were
suppressed compared with the 48 h group; whereas levels of IL-10
continued to increase in a time-dependent manner (Table II).
| Table II.Concentrations of TNF-α, IL-6 and
IL-10 in acute liver injury model rats at three time intervals and
normal control rats. |
Table II.
Concentrations of TNF-α, IL-6 and
IL-10 in acute liver injury model rats at three time intervals and
normal control rats.
|
| TNF-α (pg/ml) | IL-6 (pg/ml) | IL-10 (pg/ml) |
|---|
| NC | 8.20±3.12 | 5.12±1.12 | 7.12±2.15 |
| M-24 h |
29.20±3.54a |
21.60±3.59a |
18.69±4.56a |
| M-48 h |
53.40±5.21a |
46.52±3.26a |
25.78±5.89a |
| M-72 h |
30.60±3.58a |
36.89±4.69a |
34.65±5.68a |
Discussion
Liver diseases have previously been reported to be
associated with gastrointestinal function (20). When liver function is severely
impaired, it is frequently accompanied by endotoxemia; meanwhile in
intestinal barrier function disorders, a large number of intestinal
bacteria translocate from the intestinal lumen causing infection,
and increasing the production of endotoxins (21). In addition, the intestinal canal
represents a bacterial repository, and the largest gram-negative
bacterial endotoxin pool (22,23).
Therefore, it is very important to maintain intestinal barrier
function, limit bacterial translocation and reduce endotoxemia in
the treatment of liver disease (23,24).
Intestinal mucosal barrier function is essential for
the working of the intestine (25). The normal intestinal mucosal
barrier is composed of mechanical, chemical, immune and biological
barriers, of which the intestinal mechanical barrier is the most
important, and is predominantly composed of intestinal epithelial
cells and TJs between the epithelial cells. The mechanical barrier,
which physically separates the lumen and the internal milieu,
facilitates vectorial transport of nutrients ions, and other
substances (26). In addition,
there is communication between epithelial cells and the immune
system (27). Therefore, the
mucosal barrier exhibits physical, biochemical and immune features
(25). However, numerous
inflammatory diseases may induce intestinal mucosal barrier
dysfunction, including fulminant hepatitis (28), chronic obstructive pulmonary
disease, uremia, severe acute pancreatitis and peritoneal air
exposure (29–31). A number of traditional Chinese
medicines have been demonstrated to improve intestinal barrier
function via inhibition of the nuclear factor-κB-mediated signaling
pathway or antioxidative action, inhibiting the production of
inflammatory cytokines and upregulation of zona occludens protein 1
mRNA and protein expression levels (29,32–34).
The involvement of inflammatory cells and inflammatory factors, as
well as the immune response, results in damage to the intestinal
epithelial barrier caused by liver injury, and our model displays
these pathophysiological changes.
The most important element of the intestinal barrier
is TJs, which are composed of both cytosolic and integral membrane
proteins. TJs represent the main component of the intestinal
mucosal mechanical barrier structure and have an important role in
maintaining the integrity and permeability of the intestinal mucosa
(1,2). Claudin-1 is an important structural
protein within TJs (5,35). In liver injury, the permeability of
TJs between intestinal epithelia is increased and the distribution
of the TJ proteins are markedly altered on the intestinal mucosal
surface, as well as in membrane microdomains (36).
TNF-α is an important regulator of intestinal
inflammation that induces an increase in intestinal permeability
via extracellular signal-regulated kinase 1/2-dependent activation
of ETS transcription factor (Elk-1), which subsequently activates
the MLCK gene in the nucleus by binding to the promoter region of
MLCK (37,38). However, restoration of impaired
mucosal barrier function may be achieved via downregulation of MLCK
expression (7,8). The results of the present study
demonstrated that levels of serum TNF-α in acute liver injury rats
increased in a time-dependent manner. All the aforementioned
results suggested that the model of acute liver injury-induced
impaired mucosal barrier was successfully established.
Water homeostasis is important for neural signal
transduction in the nervous system and the function of the
intestinal epithelial barrier (4).
AQPs were first observed in the nervous system (39). Following this, AQPs were revealed
to be widely distributed in the liver, duodenum, jejunum, ileum and
colon. AQP9 is a transmembrane protein composed of a single
polypeptide transmembrane peptide 6 (40). Both rat and human AQP9 are
abundantly expressed in the liver, whereas in contrast to rat AQP9
(41), human AQP9 is also
expressed in peripheral leukocytes and in tissues that accumulate
leukocytes, including the lung, spleen and bone marrow (10,11–14).
The promoter region of AQP9 contains putative tonicity and
glucocorticoid-responsive elements, suggesting that AQP9 may be
regulated by osmolality and catabolism (42), involving osmoregulation of small
molecules (43). This mechanism is
important to the function of intestinal epithelial barrier
(44).
To the best of our knowledge, the present study is
the first to investigate the expression and function of AQP9 in the
ileum mucosa, and the results demonstrated that expression levels
of AQP9 in damaged ileal mucosa were decreased compared with the NC
group. Furthermore, the results revealed that claudin-1 expression
levels in damaged ileum mucosa were also decreased compared with
the NC group; however, MLCK expression levels were increased. These
results suggested that AQP9 may have various associations with
ileal mucosal damage.
However, the results of the present study have
predominantly focused on the association between AQP9 expression
levels and the expression of two important makers of the intestinal
epithelium; however, the specific molecular mechanism has not been
determined. In addition, the present study only investigated the
ileum mucosa; the situation of the rest of the small intestine
remains unclear.
The results of the present study revealed that AQP9
in the intestinal epithelium has an important role in the
development of acute liver injury-induced intestinal epithelium
damage.
Acknowledgements
Not applicable.
Funding
The present study was financially supported by the
Jiangxi Provincial Health Department (grant nos. 20155100 and
20167202).
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
TX and NC conceived and designed the study. TX, SG,
JW, JX, LY and XW performed the experiments. TX and NC wrote the
manuscript. All authors read and approved the manuscript.
Ethics approval and consent to
participate
All animal experiments were approved by the Animal
Ethics Association of the First Affiliated Hospital of Nanchang
University.
Patient consent for publication
Not applicable.
Competing interests
The authors state that they have no competing
interests.
References
|
1
|
Suzuki T: Regulation of intestinal
epithelial permeability by tight junctions. Cell Mol Life Sci.
70:631–659. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Dokladny K, Zuhl MN and Moseley PL:
Intestinal epithelial barrier function and tight junction proteins
with heat and exercise. J Appl Physiol (1985). 120:692–701. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Muto S: Physiological roles of claudins in
kidney tubule paracellular transport. Am J Physiol Renal Physiol.
312:F9–F24. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Khan N and Asif AR: Transcriptional
regulators of claudins in epithelial tight junctions. Mediators
Inflamm. 2015:2198432015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Gan H, Wang G, Hao Q, Wang QJ and Tang H:
Protein kinase D promotes airway epithelial barrier dysfunction and
permeability through down-regulation of claudin-1. J Biol Chem.
288:37343–37354. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ohta A, Yang SS, Rai T, Chiga M, Sasaki S
and Uchida S: Overexpression of human WNK1 increases paracellular
chloride permeability and phosphorylation of claudin-4 in MDCKII
cells. Biochem Biophys Res Commun. 349:804–808. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Du J, Chen Y, Shi Y, Liu T, Cao Y, Tang Y,
Ge X, Nie H, Zheng C and Li YC: 1,25-Dihydroxyvitamin D protects
intestinal epithelial barrier by regulating the myosin light chain
kinase signaling pathway. Inflamm Bowel Dis. 21:2495–2506. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Xiong Y, Wang J, Chu H, Chen D and Guo H:
Salvianolic acid B restored impaired barrier function via
downregulation of MLCK by microRNA-1 in rat colitis model. Front
Pharmacol. 7:1342016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Takata K, Matsuzaki T and Tajika Y:
Aquaporins: Water channel proteins of the cell membrane. Prog
Histochem Cytochem. 39:1–83. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Carbrey JM, Gorelick-Feldman DA, Kozono D,
Praetorius J, Nielsen S and Agre P: Aquaglyceroporin AQP9: Solute
permeation and metabolic control of expression in liver. Proc Natl
Acad Sci U S A. 100:2945–2950. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lindskog C, Asplund A, Catrina A, Nielsen
S and Rutzler M: A systematic characterization of aquaporin-9
expression in human normal and pathological tissues. J Histochem
Cytochem. 64:287–300. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Rojek AM, Skowronski MT, Fuchtbauer EM,
Füchtbauer AC, Fenton RA, Agre P, Frøkiaer J and Nielsen S:
Defective glycerol metabolism in aquaporin 9 (AQP9) knockout mice.
Proc Natl Acad Sci U S A. 104:3609–3614. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Spegel P, Chawade A, Nielsen S, Kjellbom P
and Rutzler M: Deletion of glycerol channel aquaporin-9 (Aqp9)
impairs long-term blood glucose control in C57BL/6 leptin
receptor-deficient (db/db) obese mice. Physiol Rep. 3:e125382015.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Okada S, Misaka T, Matsumoto I, Watanabe H
and Abe K: Aquaporin-9 is expressed in a mucus-secreting goblet
cell subset in the small intestine. FEBS Lett. 540:157–162. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Matsuzaki T, Tajika Y, Ablimit A, Aoki T,
Hagiwara H and Takata K: Aquaporins in the digestive system. Med
Electron Microsc. 37:71–80. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu W, Hou S, Yan J, Zhang H, Ji Y and Wu
X: Quantification of proteins using enhanced etching of Ag coated
Au nanorods by the Cu(2+)/bicinchoninic acid pair with improved
sensitivity. Nanoscale. 8:780–784. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Rudler M, Mouri S, Charlotte F, Lebray P,
Capocci R, Benosman H, Poynard T and Thabut D: Prognosis of treated
severe alcoholic hepatitis in patients with gastrointestinal
bleeding. J Hepatol. 62:816–821. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Konno R, Yamakawa H, Utsunomiya H, Ito K,
Sato S and Yajima A: Expression of survivin and Bcl-2 in the normal
human endometrium. Mol Hum Reprod. 6:529–534. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang C, Li Y, Zhang XY, Liu L, Tong HZ,
Han TL, Li WD, Jin XL, Yin NB, Song T, et al: Therapeutic potential
of human minor salivary gland epithelial progenitor cells in liver
regeneration. Sci Rep. 7:127072017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Elamin E, Masclee A, Troost F, Pieters HJ,
Keszthelyi D, Aleksa K, Dekker J and Jonkers D: Ethanol impairs
intestinal barrier function in humans through mitogen activated
protein kinase signaling: A combined in vivo and in vitro approach.
PloS One. 9:e1074212014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Armand-Lefevre L, Angebault C, Barbier F,
Hamelet E, Defrance G, Ruppé E, Bronchard R, Lepeule R, Lucet JC,
El Mniai A, et al: Emergence of imipenem-resistant gram-negative
bacilli in intestinal flora of intensive care patients. Antimicrob
Agents Chemother. 57:1488–1495. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Resino E, San-Juan R and Aguado JM:
Selective intestinal decontamination for the prevention of early
bacterial infections after liver transplantation. World J
Gastroenterol. 22:5950–5957. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Baranova IN, Souza AC, Bocharov AV,
Vishnyakova TG, Hu X, Vaisman BL, Amar MJ, Chen Z, Kost Y, Remaley
AT, et al: Human SR-BI and SR-BII potentiate
lipopolysaccharide-induced inflammation and acute liver and kidney
injury in mice. J Immunol. 196:3135–3147. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
de Medina Sanchez F, Romero-Calvo I,
Mascaraque C and Martinez-Augustin O: Intestinal inflammation and
mucosal barrier function. Inflamm Bowel Dis. 20:2394–2404. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Pelaseyed T, Bergstrom JH, Gustafsson JK,
Ermund A, Birchenough GM, Schütte A, van der Post S, Svensson F,
Rodríguez-Piñeiro AM, Nyström EE, et al: The mucus and mucins of
the goblet cells and enterocytes provide the first defense line of
the gastrointestinal tract and interact with the immune system.
Immunol Rev. 260:8–20. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Turner JR: Intestinal mucosal barrier
function in health and disease. Nat Rev Immunol. 9:799–809. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wang X, Yan S, Xu D, Li J, Xie Y, Hou J,
Jiang R, Zhang C and Sun B: Aggravated liver injury but attenuated
inflammation in PTPRO-deficient mice following LPS/D-GaIN induced
fulminant hepatitis. Cell Physiol Biochem. 37:214–224. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yu C, Tan S, Zhou C, Zhu C, Kang X, Liu S,
Zhao S, Fan S, Yu Z, Peng A and Wang Z: Berberine reduces
uremia-associated intestinal mucosal barrier damage. Biol Pharm
Bull. 39:1787–1792. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li HC, Fan XJ, Chen YF, Tu JM, Pan LY,
Chen T, Yin PH, Peng W and Feng DX: Early prediction of intestinal
mucosal barrier function impairment by elevated serum procalcitonin
in rats with severe acute pancreatitis. Pancreatology. 16:211–217.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Tan SJ, Yu C, Yu Z, Lin ZL, Wu GH, Yu WK,
Li JS and Li N: High-fat enteral nutrition reduces intestinal
mucosal barrier damage after peritoneal air exposure. J Surg Res.
202:77–86. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wen JB, Zhu FQ, Chen WG, Jiang LP, Chen J,
Hu ZP, Huang YJ, Zhou ZW, Wang GL, Lin H and Zhou SF: Oxymatrine
improves intestinal epithelial barrier function involving
NF-κB-mediated signaling pathway in CCl4-induced cirrhotic rats.
PLoS One. 9:e1060822014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chen J, Zhang R, Wang J, Yu P, Liu Q, Zeng
D, Song H and Kuang Z: Protective effects of baicalin on
LPS-induced injury in intestinal epithelial cells and intercellular
tight junctions. Can J Physiol Pharmacol. 93:233–237. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zong X, Hu W, Song D, Li Z, Du H, Lu Z and
Wang Y: Porcine lactoferrin-derived peptide LFP-20 protects
intestinal barrier by maintaining tight junction complex and
modulating inflammatory response. Biochem Pharmacol. 104:74–82.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Saeedi BJ, Kao DJ, Kitzenberg DA,
Dobrinskikh E, Schwisow KD, Masterson JC, Kendrick AA, Kelly CJ,
Bayless AJ, Kominsky DJ, et al: HIF-dependent regulation of
claudin-1 is central to intestinal epithelial tight junction
integrity. Mol Biol Cell. 26:2252–2262. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Al-Sadi R, Guo S, Ye D, Rawat M and Ma TY:
TNF-α modulation of intestinal tight junction permeability is
mediated by NIK/IKK-α axis activation of the canonical NF-κB
pathway. Am J Pathol. 186:1151–1165. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Liu R, Li X, Huang Z, Zhao D, Ganesh BS,
Lai G, Pandak WM, Hylemon PB, Bajaj JS, Sanyal AJ and Zhou H: C/EBP
homologous protein-induced loss of intestinal epithelial stemness
contributes to bile duct ligation-induced cholestatic liver injury
in mice. Hepatology. 67:1441–1457. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Al-Sadi R, Guo S, Ye D and Ma TY: TNF-α
modulation of intestinal epithelial tight junction barrier is
regulated by ERK1/2 activation of Elk-1. Am J Pathol.
183:1871–1884. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Guérin CF, Regli L and Badaut J: Roles of
aquaporins in the brain. Med Sci (Paris). 21:747–752. 2005.(In
French). View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Dibas A, Yang MH, Bobich J and Yorio T:
Stress-induced changes in neuronal Aquaporin-9 (AQP9) in a retinal
ganglion cell-line. Pharmacol Res. 55:378–384. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Chen XF, Li CF, Lu L and Mei ZC:
Expression and clinical significance of aquaglyceroporins in human
hepatocellular carcinoma. Mol Med Rep. 13:5283–5289. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Liu Z, Carbrey JM, Agre P and Rosen BP:
Arsenic trioxide uptake by human and rat aquaglyceroporins. Biochem
Biophys Res Commun. 316:1178–1185. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Tsukaguehi H, Weremowiez S, Morton CC and
Hediger MA: Funetional and molecular characterization of the human
neutral solute channel aquaporin-9. Am J Physiol. 277:F685–F696.
1999.PubMed/NCBI
|
|
43
|
Yang M, Gao F, Liu H, Yu WH, Zhuo F, Qiu
GP, Ran JH and Sun SQ: Hyperosmotic induction of aquaporin
expression in rat astrocytes through a different MAPK pathway. J
Cell Biochem. 114:111–119. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Lei Q, Qiang F, Chao D, Di W, Guoqian Z,
Bo Y and Lina Y: Amelioration of hypoxia and LPS-induced intestinal
epithelial barrier dysfunction by emodin through the suppression of
the NF-kappaB and HIF-1alpha signaling pathways. Int J Mol Med.
34:1629–1639. 2014. View Article : Google Scholar : PubMed/NCBI
|