Introduction
Liver cancer is one of the leading causes of
cancer-associated mortality in worldwide (1). Patients suffering from this disease
are often diagnosed with late-stage cancer and severely impaired
liver function. Due to the poor response to hepatic resection,
patients may only be able to be treated with chemotherapy (2). At present, the primary
chemotherapeutic drugs for liver cancer are shikonin,
5-fluorouracil (5-FU) and cisplatin, but these drugs have severe
side effects and are expensive (3,4).
Therefore, novel therapeutic agents with improved efficiency and
lower toxicity are urgently required for liver cancer.
Apoptosis is an active form of chemotherapy-induced
cell death characterized by orderly cell death, and is mediated by
mitochondrial dysfunction. The caspase cascade serves an important
role in this process (5). The
B-cell lymphoma 2 (Bcl-2) family consists of proteins that either
promote or inhibit apoptosis. Inhibiting anti-apoptotic protein
Bcl-2 and promoting pro-apoptotic protein Bcl-2-associated X
protein (Bax) expression can be targeted for cancer chemoprevention
(6). Caspase-3 is one of the most
important molecules in the regulation of apoptosis and cell
survival, and exists as an inactive enzyme that undergoes
proteolytic cleavage at conserved aspartic residues, to produce
large and small subunits that dimerize to form the active enzyme
(7).
The mitogen-activated protein kinase (MAPK) pathway
is activated by upstream genomic events and/or activation of
multiple signaling events where information coalesces at this
important nodal pathway point (8).
The p38 MAPK pathway is an important regulator of a number of
cellular responses. It is well established that enhanced p38
activity correlates with a poor clinical prognosis in certain tumor
types (9–11). The c-Jun N-terminal kinase (JNK) is
a master protein kinase that regulates a number of physiological
processes including inflammatory responses, morphogenesis, and cell
proliferation, differentiation, survival and death (12,13).
Extracellular signal-regulated kinase (ERK) phosphorylation results
in the activation of multiple substrates that are responsible for
stimulating cell proliferation (14). In addition, signal transducer and
activator of transcription 3 (STAT3) is persistently activated in
several types of cancer, and also regulates numerous cardinal
features of cancer, including cancer cell growth, apoptosis,
metastasis and tumor angiogenesis (15,16).
In addition, the protein expression level of STAT3 in HCC tissues
was identified to be increased compared with that in normal liver
and adjacent tissues (17).
Reactive oxygen species (ROS) within cells including
hydrogen peroxide, superoxide anions and hydroxyl radicals serve as
second messengers in the regulation of a number of important
cellular events including transcription factor activation, gene
expression and cellular proliferation, differentiation and
senescence (18,19). In addition to endogenous sources of
ROS, ROS levels may also increase due to chemical stimulation,
ultraviolet radiation and thermal exposure (20). ROS have also been implicated in the
metabolic reprogramming of cancer cells, serving important roles in
tumor initiation, progression and metastasis (21). Excessive production of ROS leads to
disruption of the homeostasis of the intracellular redox status,
which may directly induce oxidative damage in lipids, proteins and
nucleic acids, thereby killing cancer cells by disturbing their
metabolism and signal transduction (22). In addition, based on the different
redox status of normal and cancer cells, a promising therapeutic
strategy based on drugs that increase ROS generation and induce
apoptosis in cancer cells has arisen cancer therapy (23).
As naphthalene organic derivatives,
1,4-naphthoquinone compounds have been extensively investigated for
their potential biological benefits, including their
anti-inflammatory and anti-bacterial activities (24). In addition, the 1,4-naphthoquinone
pharmacophore exhibits anti-cancer activity and has been the focus
of previous studies (25,26). Among the 1,4-naphthoquinone
compounds, plumbagin, mitomycin and shikonin have been used in the
development of potent anti-cancer drugs (27). However, these compounds exhibit
high levels of cytotoxicity and significant side effects, making
their application as anti-cancer drugs in the clinical setting
problematic (28). Therefore, our
previous study used 1,4-naphthoquinone as a common compound to
synthesize novel naphthoquinone derivatives, which indicated
significantly improved cytotoxicity and increased anti-tumor
activity in several types of cancer cells (29,30).
In an attempt to develop compounds with decreased
side effects and optimized antitumor effects, two novel types of
1,4-naphthoquinone derivatives were synthesized, namely
2,3-dihydro-2,3-epoxy-2-propylsulfonyl-5,8-dimethoxy-1,4-naphthoquinone
(EPDMNQ) and
2,3-dihydro-2,3-epoxy-2-nonylsulfonyl-5,8-dimethoxy-1,4-naphthoquinone
(ENDMNQ). Then, their effects on anti-proliferation, apoptosis
induction and ROS generation in liver cancer cells were detected.
The molecular mechanisms of apoptosis induced by EPDMNQ and ENDMNQ
were also explored in Hep3B cells.
Materials and methods
Synthesis of the 1,4-naphthoquinone
derivatives EPDMNQ and ENDMNQ
AlCl3 (142 g, 1.06 mol) and NaCl (28.3 g,
0.48 mol) were melted at 150–155°C, and a mixture of
1,4-dimethoxybenzene (0.12 mol) and maleic anhydride (0.24 mol) was
added to the melted mixture. The temperature was sustained at
170–175°C for 1–2 min and the dark red melt was allowed to cool.
Next, the distilled water (1,400 ml) and 98% HCl (100 ml) were
added to the melt mixture and continuously mixed for 12 h to
generate naphthazarin. The mixture of naphthazarin (19.0 g),
tetrahydrofuran (C4H8O; 200 ml),
Na2S2O4 (10.6 g), distilled water
(200 ml) and tetrabutylammonium bromide
(C16H36BrN, 2.0 g) were mixed for 1 h. Then,
a solution of C2H6O4S (25 ml),
NaOH (40 ml) and Na2S2O4 (10.6 g)
were added to the mixture stirred for 22 h at room temperature. The
1,4,5,8-tetramethoxynaphthalene was then recovered by filtration
and recrystallized from petroleum ether (boiling point 90–120°C).
The residue was recrystallized from MeOH to give the title compound
5,8-dimethoxy-1,4-naphthoquinone (DMNQ) The 1-Mercaptopropane (1.65
mmol) and 1-Nonanethiol (1.65 mmol) was added to a solution of DMNQ
(1.38 mmol) and MeOH (30 ml) respectively. The mixture was then
stirred at room temperature for 4 h. A solution of sodium
dichromate
(Na2Cr2O7·2H2O, 0.76
mmol) and 98% H2SO4 (0.23 mmol) was then
added and stirred for 2 min. The reaction was conducted at room
temperature with m-chloroperoxybenzoic acid to produce the final
products, 1,4-naphthoquinone derivatives EPDMNQ and ENDMNQ. The
acidic solution was extracted with dichloromethane
(CHCl2; 60 ml). The organic layer was washed with brine
(Fig. 1A and B).
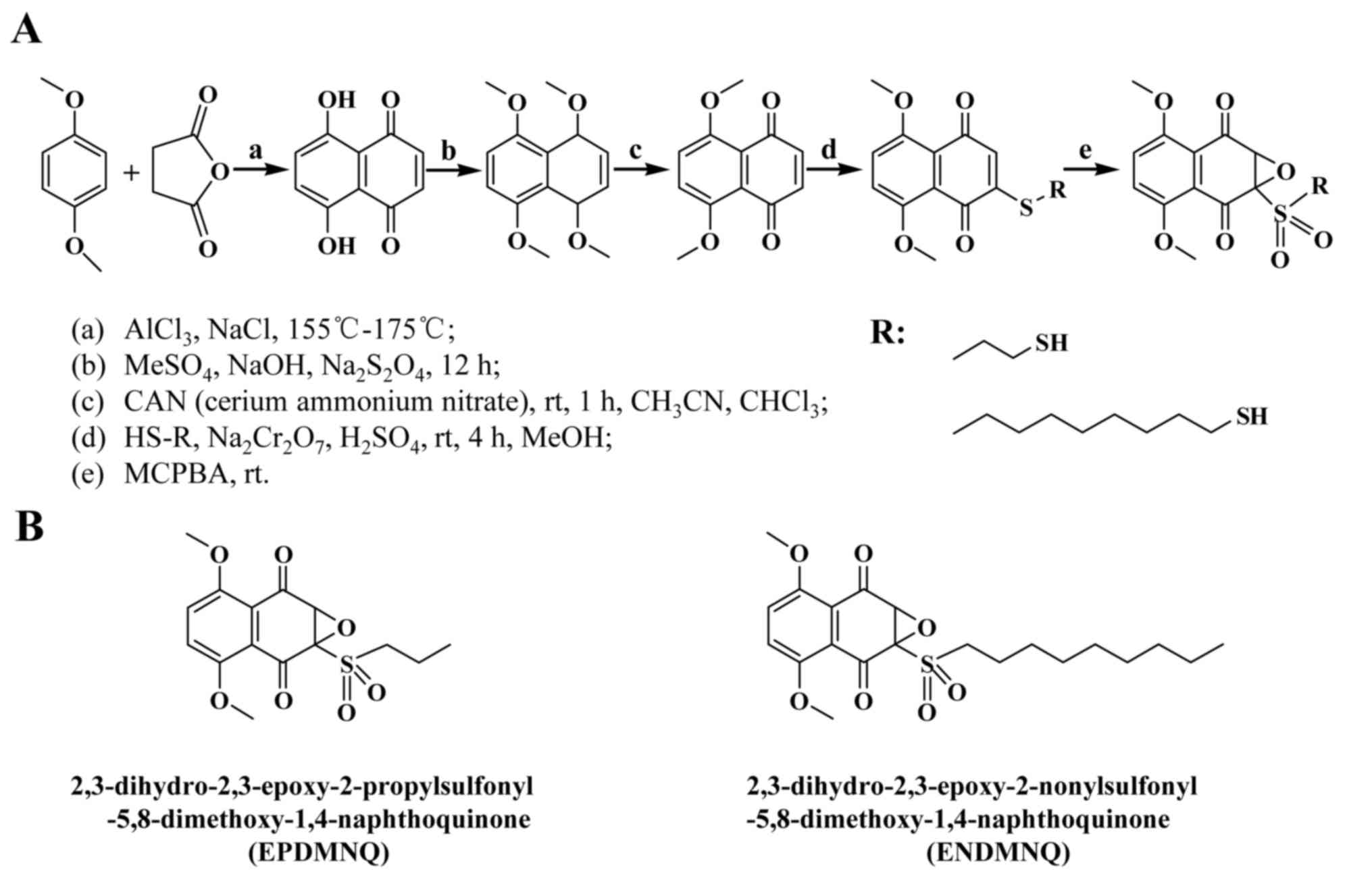 | Figure 1.Synthesis of 1,4-naphthoquinone
derivatives EPDMNQ and ENDMNQ. (A) Process of synthetic EPDMNQ and
ENDMNQ generation. (B) Structural formulas of EPDMNQ and ENDMNQ.
EPDMNQ,
2,3-dihydro-2,3-epoxy-2-propylsulfonyl-5,8-dimethoxy-1,4-naphthoquinone;
ENDMNQ,
2,3-dihydro-2,3-epoxy-2-nonylsulfonyl-5,8-dimethoxy-1,4-naphthoquinone. |
Nuclear magnetic resonance (NMR) spectra were
recorded on JNM-AL 600 (600 MHz) and JNM-AL 150 (150 MHz)
spectrometers. Chemical shifts (d) were measured as ppm downfield
from tetramethylsilane as the internal standard. Mass spectra were
collected with the AB SCIEX API 2000 LC/MS/MS System (Applied
Biosystems; Thermo Fisher Scientific, Inc., Waltham, MA, USA) and
LCMS-IT-TOF (Shimadzu (China) Co., Ltd., Beijing, China).
Cell lines and cell culture
Human hepatocellular carcinoma cells Hep3B and Huh7,
and human hepatoblastoma HepG2 cells were obtained from the
American Type Culture Collection (Manassas, VA, USA), and normal
liver L-02, normal lung IMR-90 and normal stomach GES-1 cell lines
were obtained from (Saiqi Biological Engineering Co., Ltd.,
Shanghai, China). Cells were cultured in Dulbecco's modified
Eagle's medium (DMEM; Gibco; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) supplemented with 10% fetal bovine serum (Gibco;
Thermo Fisher Scientific, Inc.). The cultures were maintained at
37°C in a humidified atmosphere of 5% CO2.
MTT assay
Human hepatocellular carcinoma cells Hep3B and Huh7,
human hepatoblastoma HepG2, normal liver L-02, normal lung IMR-90
and normal stomach GES-1 cells were seeded in 96-well plates at a
density of 1×104 cells/well. Following overnight
incubation at 37°C, cells were treated with different
concentrations (1, 3, 10, 30 or 100 µmol/l) of 5-FU, EPDMNQ and
ENDMNQ for 24 h at 37°C. Following treatment, 15 µl MTT (5 mg/ml)
was added to each well and incubated for 2 h. The absorbance values
of the solution were measured at 490 nm with a microplate
illuminometer (BioTek Instruments Inc., Winooski, VT, USA).
Annexin V-fluorescein isothiocyanate
(FITC)/propidium iodide (PI) double staining
Hep3B cells were seeded in 6-well plates at a
density of 1×106 cells/well. After 24 h incubation at
37°C, cells were treated with 4 µmol/l 5-FU, EPDMNQ and ENDMNQ for
different time points (0, 3, 6, 12 or 24 h). Cells were washed with
PBS, and stained with Annexin V-FITC (10 µl) and PI (5 µl) in the
dark for 15 min. Then, cells were observed using the Leica
fluorescence microscope DM 2500 (Leica Microsystems GmbH, Wetzlar,
Germany) at magnification, ×400.
Flow cytometry analysis
Hep3B cells were seeded in 6-well plates at a
density of 1×106 cells/well. After 24 h incubation at
37°C, cells were treated with 4 µmol/l 5-FU, EPDMNQ and ENDMNQ for
different time points (0, 3, 6, 12 or 24 h), and pretreated with
N-acetyl cysteine (NAC, 5 mmol/l, Sigma-Aldrich; Merck KGaA) for 30
min and incubated with 4 µmol/l EPDMNQ and ENDMNQ for 24 h,
respectively. Cells were stained with Annexin V-FITC (10 µl) and PI
(5 µl) (Beyotime Institute of Biotechnology, Shanghai, China) in
the dark for 15 min. The frequency of apoptotic cells in the
treatment groups was determined by flow cytometry (Beckman Coulter,
Inc., Brea, CA, USA). Harvested cells were treated with
2′,7-dichlorodihydrofluorescein diacetate (10 mmol/l) (DCFH-DA;
Merck, Shanghai, China) at 37°C for 30 min to allow ROS
measurement. ROS contents were determined by flow cytometry.
Harvested cells were treated with JC-1 (10 µg/ml) (Beyotime
Institute of Biotechnology) at room temperature for 5 min to detect
mitochondrial membrane potential (ΔΨm) depolarization. States of
ΔΨm depolarization were determined by flow cytometry and CytExpert
software (version 1.2; Beckman Coulter, Inc., Brea, CA, USA) was
used to analyze the data.
Western blot analysis
Harvested Hep3B cells were lysed in lysis buffer (50
mmol/l Tris (pH 7.4), 150 mmol/l NaCl, 1% Triton X-100, 1% sodium
deoxycholate, 0.1% SDS, 20 mg/ml AEBSF, 0.5 mg/ml pepstatin, 0.5
mg/ml leupeptin and 2 mg/ml aprotinin; Beyotime Institute of
Biotechnology), The protein concentrations were determined using
Bradford reagent (Bio-Rad Laboratories, Inc., Hercules, CA, USA),
following which protein lysates (30 µg) were resolved on 8–12%
SDS-PAGE and electrotransferred onto nitrocellulose membranes (EMD
Millipore, Billerica, MA, USA). The membranes were blocked for 2 h
at room temperature in fresh 5% non-fat milk in 10 mM Tris-HCl
containing 150 mM NaCl (TBS; pH 7.5) and TBS+0.2% Tween-20 (TBST),
followed by incubation with specific primary antibodies (all
obtained from Santa Cruz Biotechnology, Inc., Dallas, TX, USA)
against mouse monoclonal α-tubulin (1:2,500; cat. no. sc-8035),
Bcl-2 (1:1,500; cat. no. sc-7382), Bax (1:1,500; cat. no. sc-493),
cleaved (cle)-poly (adenosine 5-diphosphate-ribose) polymerase
(cle-PARP; 1:1,500; cat. no. sc-8007), cle-caspase-3 (1:1,500; cat.
no. sc-373730), phosphorylated (p)-p38 (Tyr182, 1:1,500;
cat. no. sc-7973), p-JNK (Tyr183 and Tyr185,
1:1,500; cat. no. sc-6254), JNK (1:1,500; cat. no. sc-7345), p-ERK
(Tyr204, 1:1,500; cat. no. sc-8059), p-STAT3
(Tyr705, 1:1,500; cat. no. sc-8059) and STAT3 (1:1,500;
cat. no. sc-8019). Rabbit polyclonal antibodies included p38α/β
(1:1,500; cat. no. sc-7972) and ERK2 (1:1,500; cat. no. sc-154),
overnight at 4°C. Following five washes with TBST, the membranes
were incubated with peroxidase-conjugated AffiniPure goat
anti-mouse IgG (1:5,000; cat. no. ZB-2305; OriGene Technologies,
Inc., Beijing, China) and goat anti-rabbit IgG (1:5,000; cat. no.
ZB-2305; OriGene Technologies, Inc.) secondary antibodies for 1 h
at room temperature, and immunoreactive protein bands were detected
with an Amersham imager (AI600; GE Healthcare, Chicago, IL, USA).
The blots were analyzed using Image J version 1.46r (National
Institutes of Health, Bethesda, MD, USA) and protein levels were
normalized to the matching densitometry value of α-tubulin as the
internal control. The change of the expression levels of p-p38,
p-JNK, p-ERK and p-STAT3 was based on the expression levels of p38,
JNK, ERK and STAT3.
Statistical analysis
Data are presented as the mean ± standard deviation
of three independent experiments. The samples of each group were
compared by analysis of variance, and multiple comparisons between
groups were performed using one-way analysis of variance followed
by Tukey's post hoc tests using SPSS version 18.0 statistical
software (SPSS, Inc., Chicago, IL, USA). P<0.05 was considered
to indicate a statistically significant difference.
Results
Synthesis of the 1,4-naphthoquinone
derivatives EPDMNQ and ENDMNQ
To improve activity and decrease side effects, the
chemical synthesis of the 1,4-naphthoquinone derivatives EPDMNQ and
ENDMNQ was modified (Fig. 1A and
B). By performing NMR at a wavelength of 400 MHz, the H and C
spectra were analyzed in deuterated chloroform solvent and the
following structures were identified.
EPDMNQ: 1H-NMR (CDCl3, 600
MHz): δ 7.14 [singlet (s), 2H], 4.5 (s, 1H), 3.99 (s, 6H), 3.60
[multiplet (m), J=6.5 Hz, 2H], 1.9 (m, 2H), 1.1 (m, 3H).
13C NMR (CDCl3, 150 MHz) δ187.2 (C-1), 184.6
(C-4), 153.4 (C-5), 152.8 (C-8), 152.7 (C-2), 130.3 (C-3), 128.3
(C-7), 120.7 (C-6), 120.3 (C-10), 120.1 (C-9), 56.8
(OCH3), 56.4 (OCH3), 56.9 (C-1′), 49.8
(C-2′), 14.9 (C-3′); ion trap-time of flight mass spectrometer
(IT-TOF/MS): m/z 364.41 (M+Na)+.
ENDMNQ: 1H-NMR (CDCl3, 600
MHz): δ 7.14 (s, 2H), 4.5 (s, 1H), 3.99 (s, 6H), 3.60 (m,
J=6.4 Hz, 2H), 1.9 (m, 14H), 1.1 (m, 3H); 13C NMR
(CDCl3, 150 MHz). δ 191.1 (C-1), 189.3 (C-4), 154.8
(C-5), 153.4 (C-8), 152.8 (C-2), 129.9 (C-3), 128.3 (C-7), 120.7
(C-6), 120.6 (C-10), 119.1 (C-9), 56.8 (OCH3), 56.5
(OCH3), 56.9 (C-1′), 45.3 (C-2′), 29.7 (C-3′), 25.9
(C-4′), 20.5 (C-5′), 15.2 (C-6′), 14.7 (C-6′), 14.7 (C-7′), 14.1
(C-8′), 13.1 (C-9′); IT-TOF/MS: m/z 447.56 (M+Na)+.
EPDMNQ and ENDMNQ treatment
selectively kills liver cancer cells but not normal cells
To determine whether EPDMNQ and ENDMNQ had cytotoxic
effects in liver cancer cells, cell viabilities were determined by
the MTT assay. As demonstrated in Fig.
2A, EPDMNQ and ENDMNQ inhibited Hep3B, HepG2 and Huh7 cell
proliferation in a dose-dependent manner. The cytotoxic effect of
EPDMNQ and ENDMNQ on liver cancer cells was significantly greater
compared with 5-FU. The effects of EPDMNQ half maximal inhibitory
concentration (IC50) values of Hep3B, HepG2 and Huh7
cells were 3.89±1.22, 5.26±1.64 and 7.68±1.54 µmol/l, respectively.
The effects of ENDMNQ IC50 values of Hep3B, HepG2 and
Huh7 cells were 4.89±2.09, 6.90±2.21 and 16.54±1.24 µmol/l,
respectively. As indicated in Fig.
2B, EPDMNQ and ENDMNQ exhibited lower cytotoxicity compared
with 5-FU treatment in normal liver L-02, normal lung IMR-90 and
normal stomach GES-1 cell lines. As Hep3B cells exhibited the
lowest IC50 values, and were most sensitive to EPDMNQ
and ENDMNQ of the 3 cancer cell lines, Hep3B cells were used for
the subsequent studies
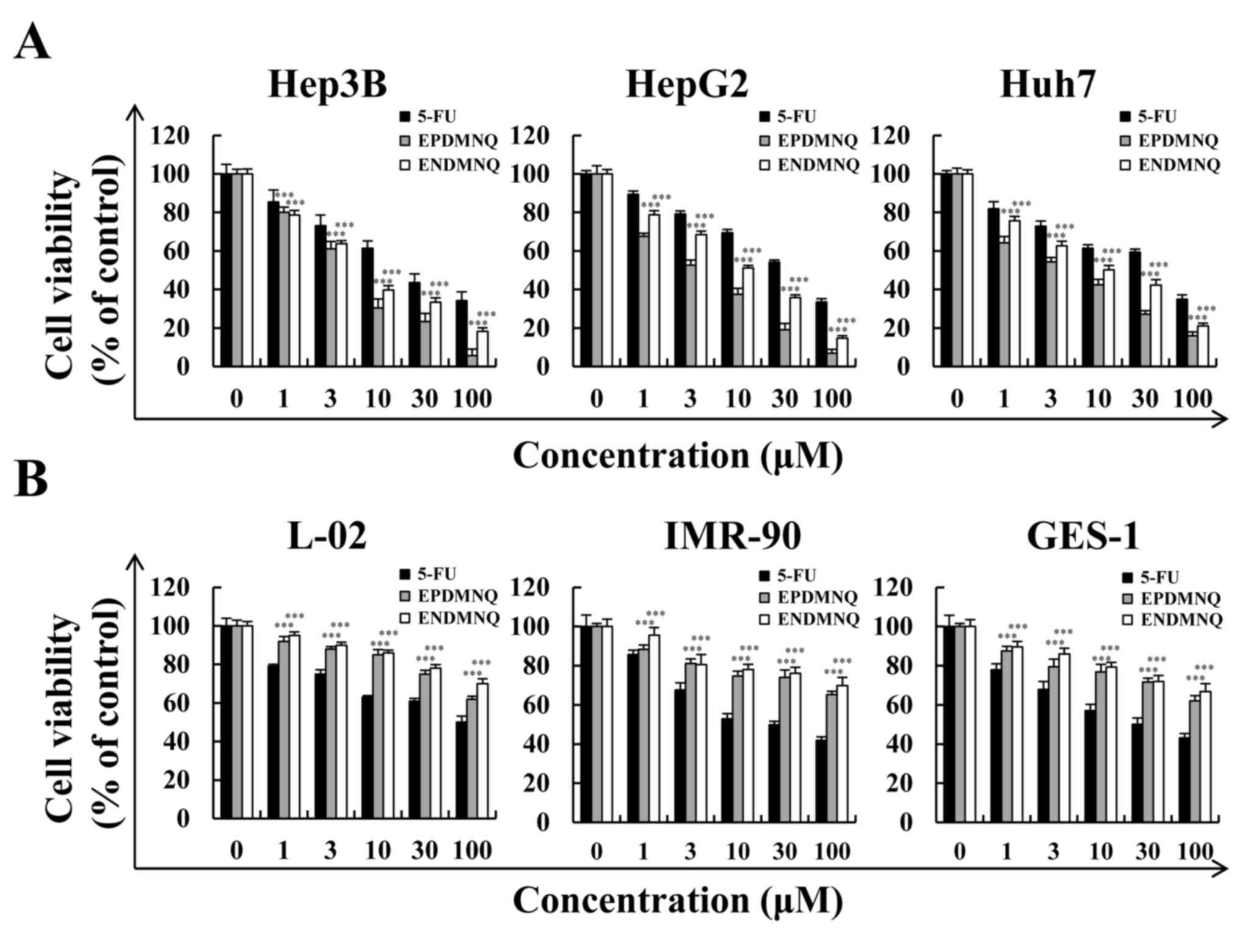 | Figure 2.Effects of EPDMNQ and ENDMNQ on the
viabilities of liver cancer and normal cells. (A) Hep3B, HepG2 and
Huh7 cells were treated with different concentrations (1, 3, 10, 30
or 100 µmol/l) of 5-FU, EPDMNQ or ENDMNQ for 24 h. Cell viability
was determined by MTT assay. (B) Normal liver L-02, normal lung
IMR-90 and normal stomach GES-1 cell line viabilities. Data are
expressed as the percentage of viable cells. ***P<0.001 vs. 5-FU
group. EPDMNQ,
2,3-dihydro-2,3-epoxy-2-propylsulfonyl-5,8-dimethoxy-1,4-naphthoquinone;
ENDMNQ,
2,3-dihydro-2,3-epoxy-2-nonylsulfonyl-5,8-dimethoxy-1,4-naphthoquinone;
5-FU, 5-fluorouracil. |
EPDMNQ and ENDMNQ induce apoptosis in
Hep3B cells
To determine whether the anti-proliferative effects
of EPDMNQ and ENDMNQ were due to effects on cell apoptosis, cell
populations were detected by fluorescence microscopy. As
demonstrated in Fig. 3A and B, the
fluorescence intensities of Annexin V-FITC and PI were increased in
a time-dependent manner. Early and late apoptotic cells were
detected by flow cytometry, and identified that the percentage of
apoptotic cells after 24 h of EPDMNQ and ENDMNQ treatment were
59.03 and 47.39%, respectively (Fig.
3C and D). EPDMNQ and ENDMNQ treatment groups exhibited a
significant increase (P<0.001) cell apoptosis compared with the
5-FU groups. These results suggested that the anti-cancer effects
of EPDMNQ and ENDMNQ were also associated with the induction of
cell apoptosis.
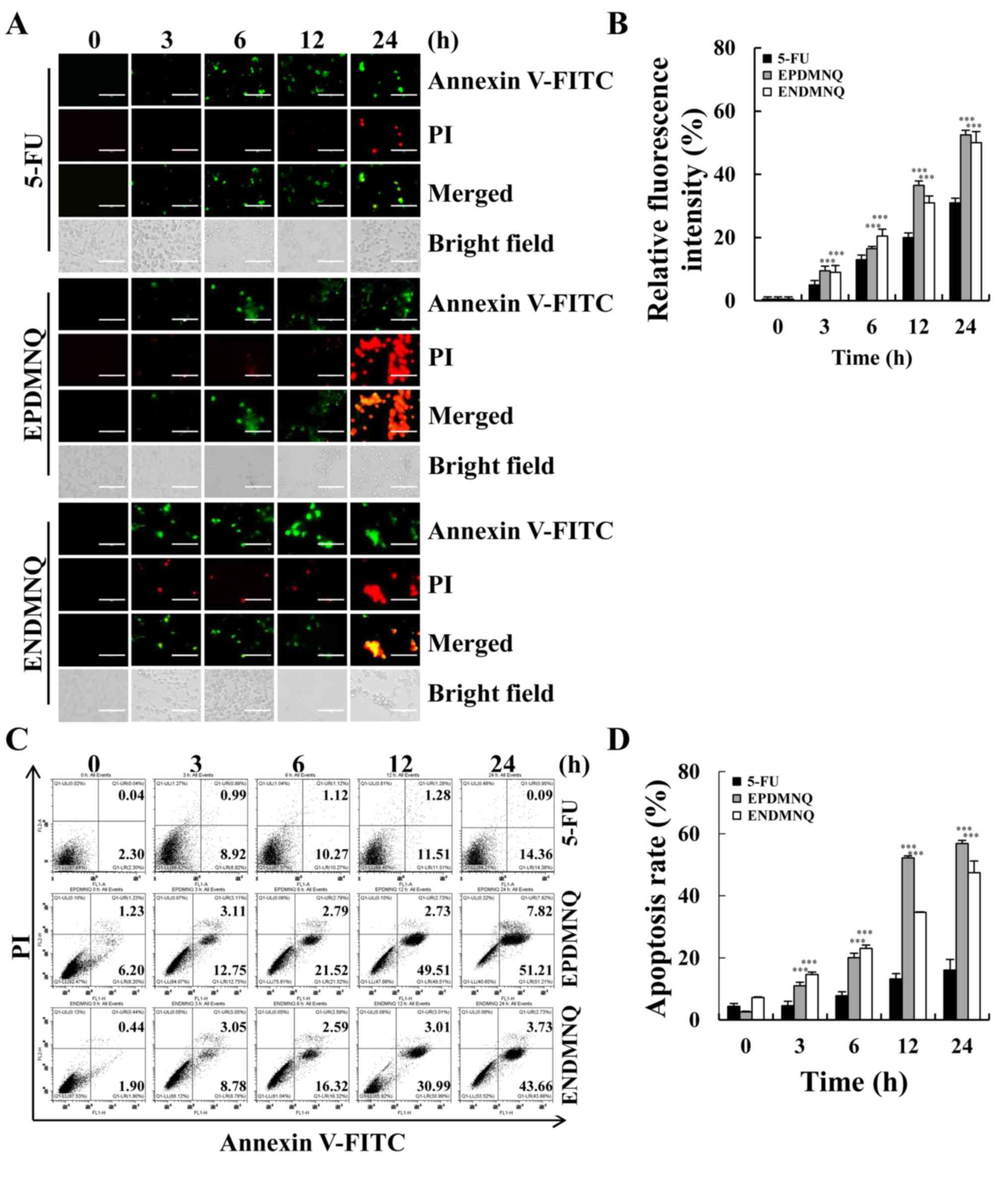 | Figure 3.EPDMNQ and ENDMNQ induce apoptosis in
hepatocellular carcinoma cells. (A) Hep3B cells were treated with 4
µmol/l 5-FU, EPDMNQ or ENDMNQ for different time intervals (3, 6,
12 or 24 h) and stained with Annexin V-FITC/PI. Images represent
fluorescence microscopic images (original magnifications, ×400).
(B) Quantification of fluorescence intensities from A. (C)
Apoptosis distribution was determined by flow cytometry following
treatment with 4 µmol/l 5-FU, EPDMNQ or ENDMNQ for different time
intervals (3, 6, 12 or 24 h). (D) Quantification of flow cytometry
from C. Data are expressed as the percentage of viable cells.
***P<0.001 vs. 5-FU group. EPDMNQ,
2,3-dihydro-2,3-epoxy-2-propylsulfonyl-5,8-dimethoxy-1,4-naphthoquinone;
ENDMNQ,
2,3-dihydro-2,3-epoxy-2-nonylsulfonyl-5,8-dimethoxy-1,4-naphthoquinone;
5-FU, 5-fluorouracil; FITC, fluorescein isothiocyanate; PI,
propidium iodide. |
EPDMNQ and ENDMNQ induce apoptosis by
modulating the mitochondrial, MAPK and STAT3 signaling pathways in
Hep3B cells
To determine the molecular mechanisms of apoptosis
induced by EPDMNQ and ENDMNQ, the expression levels of proteins
were assessed by western blot analysis. As indicated in Fig. 4A and B, the expression levels of
Bax, cle-PARP and cle-caspase-3 were increased and that of Bcl-2
was decreased in a time-dependent manner, with maximum changes
occurring at 24 h. As demonstrated in Fig. 4C and D, EPDMNQ and ENDMNQ
significantly increased the phosphorylation levels of p38 and JNK,
whereas the phosphorylation levels of ERK and STAT3 were decreased
in a time-dependent manner. These data demonstrated that EPDMNQ and
ENDMNQ induced cell apoptosis through the mitochondrial, MAPK, and
STAT3 signaling pathways.
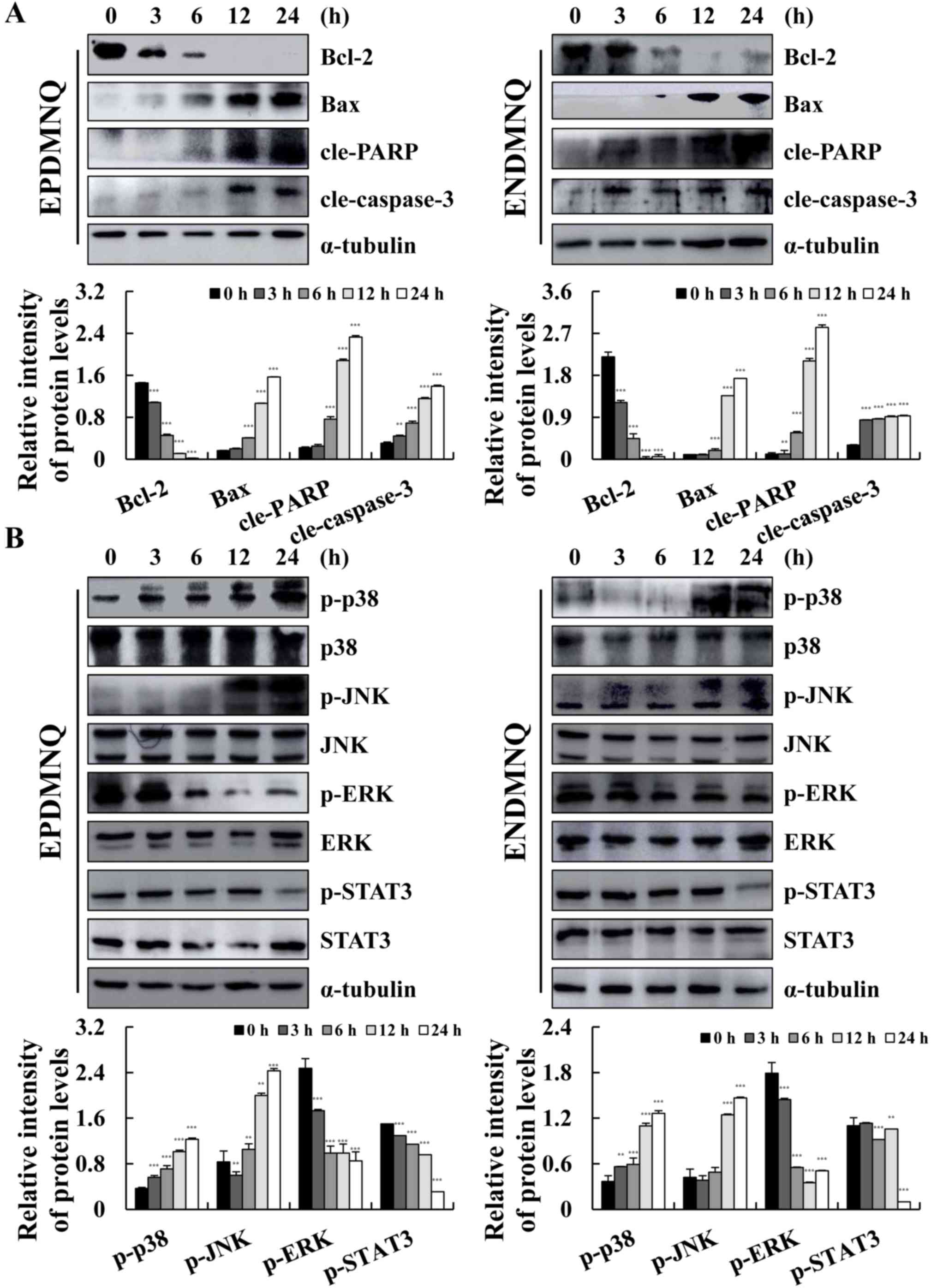 | Figure 4.EPDMNQ and ENDMNQ regulate
mitochondrial, MAPK and STAT3 signaling pathways-associated
proteins in Hep3B cells. (A) Hep3B cells were treated with EPDMNQ
or ENDMNQ for different time intervals (3, 6, 12 or 24 h). The
expression levels of mitochondrial pathway-associated proteins were
analyzed by western blot analysis. (B) The expression levels of
MAPK and STAT3 pathway-associated proteins were analyzed by western
blot analysis. α-tubulin was used as the internal control. Band
intensity was quantified using the Image J software. **P<0.01
and ***P<0.001 vs. untreated control group. EPDMNQ,
2,3-dihydro-2,3-epoxy-2-propylsulfonyl-5,8-dimethoxy-1,4-naphthoquinone;
ENDMNQ,
2,3-dihydro-2,3-epoxy-2-nonylsulfonyl-5,8-dimethoxy-1,4-naphthoquinone;
Bcl-2, B-cell lymphoma 2; Bax, Bcl-2-associated X protein; cle,
cleaved; PARP, poly (adenosine 5-diphosphate-ribose) polymerase; p,
phosphorylated; p38, p38 mitogen-activated protein kinase; JNK,
c-Jun N-terminal kinase; ERK, extracellular signal regulated
kinase; STAT3, signal transducer and activator of transcription
3. |
Accumulation of ROS induced by EPDMNQ
and ENDMNQ participates in the apoptosis of Hep3B cells
To determine whether EPDMNQ and ENDMNQ-induced
apoptosis was associated with intracellular ROS generation in Hep3B
cells, the association between intracellular ROS levels and
apoptosis was evaluated by flow cytometry. As presented in Fig. 5A and B, Hep3B cells treated with
EPDMNQ and ENDMNQ exhibited increased levels of ROS in a
time-dependent manner. ΔΨm depolarization was measured by JC-1, as
the processing time was extended from 0 to 24 h. The green
fluorescence intensity was continuously enhanced, indicating that
apoptosis gradually increased in Hep3B cells in a time-dependent
manner. Treatment with the ROS inhibitor N-acetyl-L-cysteine
(NAC) reversed the degree of apoptosis induced by EPDMNQ and ENDMNQ
(Fig. 5E and F). It was also
identified that the treatment of Hep3B cells with NAC reversed the
expression levels of proteins regulated by EPDMNQ and ENDMNQ in the
mitochondrial, MAPK, and STAT3 signaling pathways, namely JNK, ERK,
STAT3, Bcl-2, Bax, cle-PARP and cle-caspase-3 (Fig. 6A and B). These data demonstrated
that ROS generation was the key regulator of EPDMNQ and
ENDMNQ-induced apoptosis.
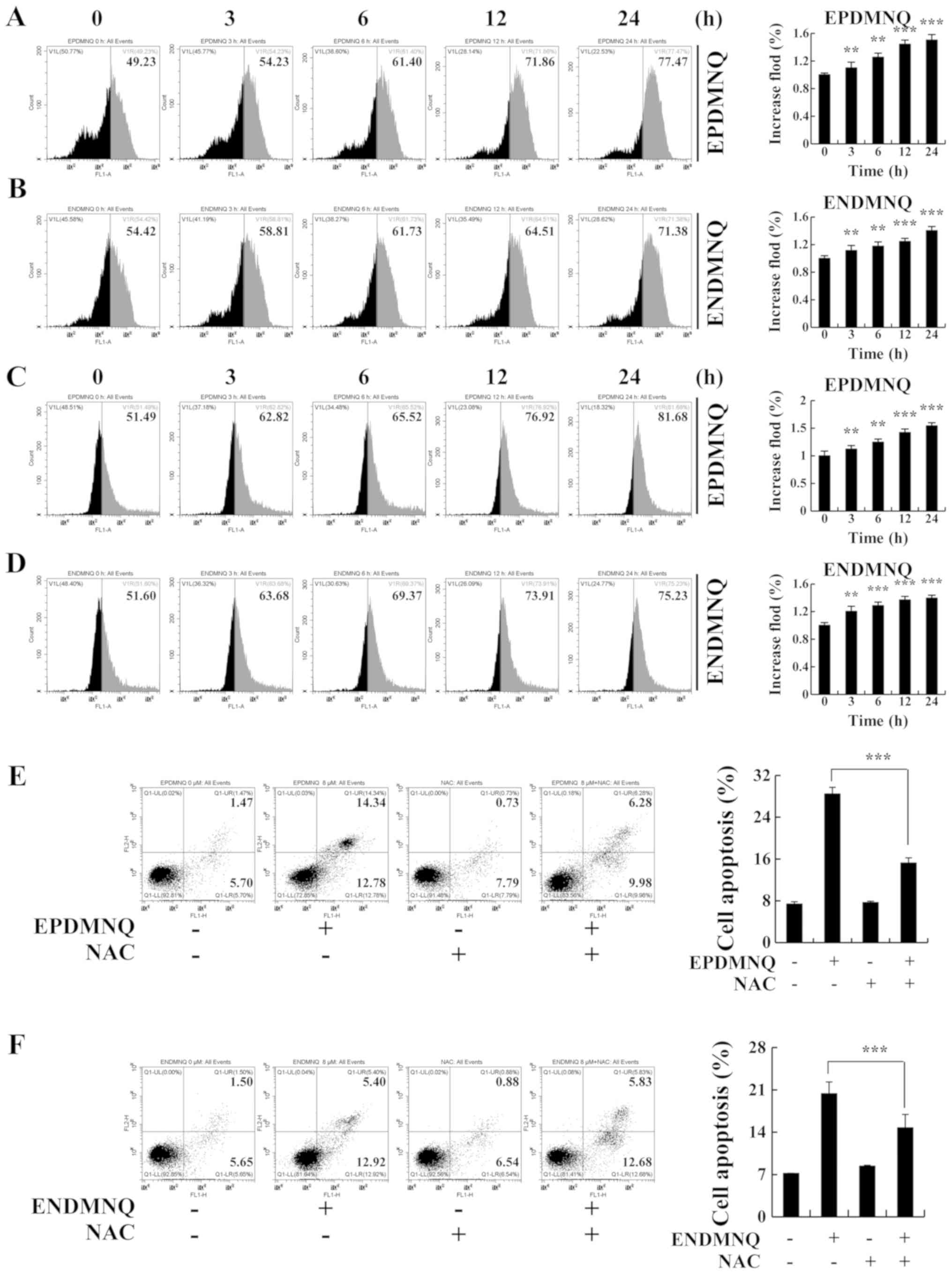 | Figure 5.EPDMNQ and ENDMNQ-induced apoptosis
is restored by ROS inhibition in Hep3B cells. (A and B)
Intracellular ROS generation induced by EPDMNQ or ENDMNQ was
measured in Hep3B cells following staining with
2′,7′-dichlorodihydrofluorescein diacetate and detected by flow
cytometry. (C and D) Hep3B cells were treated with EPDMNQ or ENDMNQ
for 24 h, and mitochondrial membrane potential depolarization value
was detected by flow cytometry following JC-1 staining. (E) Hep3B
cells were treated with NAC for 30 min and then incubated with
EPDMNQ for 24 h. Cell apoptosis was determined by flow cytometry.
(F) Apoptotic cell numbers were detected following ENDMNQ treatment
by flow cytometry. Data are expressed as the apoptotic cell
numbers. **P<0.01 and ***P<0.001 vs. untreated control group.
EPDMNQ,
2,3-dihydro-2,3-epoxy-2-propylsulfonyl-5,8-dimethoxy-1,4-naphthoquinone;
ENDMNQ,
2,3-dihydro-2,3-epoxy-2-nonylsulfonyl-5,8-dimethoxy-1,4-naphthoquinone;
ROS, reactive oxygen species; NAC, N-acetyl-L-cysteine. |
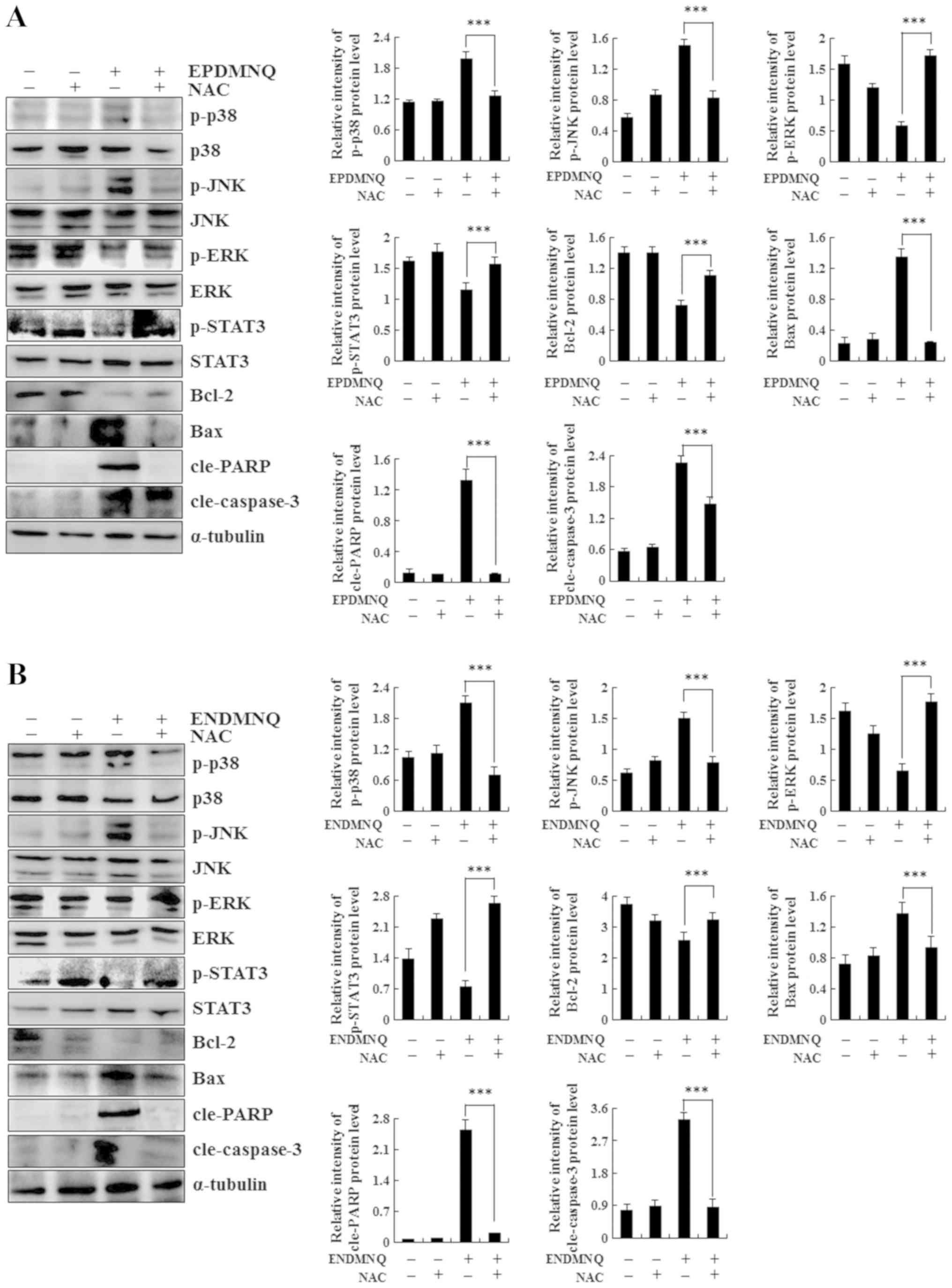 | Figure 6.Roles of mitochondrial,
mitogen-activated kinase and STAT3 signaling pathways in the
apoptosis of EPDMNQ- and ENDMNQ-treated Hep3B cells. Detection of
p-ERK, p-JNK, p-p38, p-STAT3, Bax, Bcl-2, cle-PARP and
cle-caspase-3 protein expression in (A) EPDMNQ- and (B)
ENDMNQ-treated Hep3B cells by western blot analysis. ***P<0.001
vs. untreated control groupl. EPDMNQ,
2,3-dihydro-2,3-epoxy-2-propylsulfonyl-5,8-dimethoxy-1,4-naphthoquinone;
ENDMNQ,
2,3-dihydro-2,3-epoxy-2-nonylsulfonyl-5,8-dimethoxy-1,4-naphthoquinone;
Bcl-2, B-cell lymphoma 2; Bax, Bcl-2-associated X protein; cle,
cleaved; PARP, poly (adenosine 5-diphosphate-ribose) polymerase; p,
phosphorylated; p38, p38 mitogen-activated protein kinase; JNK,
c-Jun N-terminal kinase; ERK, extracellular signal regulated
kinase; STAT3, signal transducer and activator of transcription
3. |
Discussion
The compound 1,4-naphthoquinone is a type of organic
molecule derived from naphthalene; naphthoquinone derivatives have
a number of pharmacological effects including anti-viral,
anti-bacterial and anti-inflammatory activities (31). At present, studies on
naphthoquinone derivatives have primarily focused on
5,8-dihydroxy-1,4-naphthoquinone and substitutions at the C2 and C6
positions of 5-dimethoxy-1,4-naphthoquinone, but rarely on
1,4-naphthoquinone (32,33). Although 1,4-naphthoquinone may
inhibit the growth of a variety of cancer cells, the majority of
1,4-naphthoquinone derivatives exhibit high cytotoxicity and side
effects, so they are not suitable for clinical treatment (34). Therefore, a number of attempts have
been made to identify novel therapeutic compounds that are
efficacious but have fewer side effects.
In the present study, 2 types of novel
1,4-naphthoquinone derivatives (ENDMNQ and EPDMNQ) were synthesized
and their anti-proliferation effects were screened in liver cancer
Hep3B, HepG2 and Huh7 cell lines. The liver cancer cells commonly
used in drug screening experiments include HepG2, Hep3B, Huh7,
SMMC7721, Bel-7402, MHCC97 and PLC/PRF/5 (35–38).
HepG2 cells are human hepatoblastoma that were isolated and
established from a primary hepatic blastoma of a 15-year-old
Caucasian boy in Argentina in 1979 (39). Hep3B cells are human hepatoma cell
line isolated from a liver cancer tissue of an 8-year-old African
male (40). Huh7 cells are human
hepatoma cell line obtained from a 57-year-old Japanese male liver
cancer tissue specimen in 1982 (41). These 3 cell lines have a high
degree of differentiation; the metabolism remains stable and will
not change due to the increased number of passages and in drug
screening studies. Therefore, the present study used Hep3B, HepG2
and Huh7 cells. Compared with 5-FU treatment, the cytotoxic effects
of EPDMNQ and ENDMNQ were more potent in Hep3B cells compared with
HepG2 and Huh7 cells. Certain doses (1, 3, 10, 30 or 100 µmol/l) of
EPDMNQ and ENDMNQ effectively induced liver cancer cell apoptosis
but exhibited less harmful effects in normal liver L-02, normal
lung IMR-90 and normal stomach GES-1 cell lines. It was also
identified that the IC50 of EPDMNQ was decreased
compared with IC50 of ENDMNQ. As the substituent chains
of EPDMNQ and ENDMNQ are different in length, the cytotoxic effects
of the 2 compounds on liver cancer cells are significantly
different. The cytotoxic effect of EPDMNQ with longer carbon chains
on liver cancer cells is more marked compared with that of
ENDMNQ.
Apoptosis is a common way of removing aged cells
from the body that serves an essential role in organism development
and tissue homeostasis. The majority of anti-cancer therapies
trigger apoptosis induction and regulate signaling pathways to
eliminate cancer cells (42). As a
canonical pathway that allows cells to undergo a highly regulated
form of cell death in response to pro-apoptotic stimuli, apoptosis
is triggered by multiple signaling pathways (43). The mitochondrion serves an
important role in the integration and transmission of signals to
induce apoptotic cell death and is regulated by pro-apoptotic Bax
and anti-apoptotic Bcl-2 proteins. Caspase-3 is a critical
executioner of apoptosis and acts by cleaving several essential
cellular proteins, including PARP (44). The results of the present study
indicated that EPDMNQ and ENDMNQ treatment induced a significant
change in cell morphology. Annexin V-FITC/PI double staining and
flow cytometry demonstrated that Hep3B cells treated with EPDMNQ
and ENDMNQ induced apoptosis in a dose-dependent manner.
Concomitantly, it was identified that EPDMNQ treatment exhibited a
more marked induction of apoptosis compared with ENDMNQ.
Furthermore, Hep3B cell apoptosis was regulated by increased
protein levels of Bax, cle-PARP and cle-caspase-3 and decreased
levels of Bcl-2. We hypothesized that the EPDMNQ and ENDMNQ
treatment of Hep3B cells induced apoptosis via a
mitochondrial-dependent pathway. MAPK is a critical signaling
pathway that serves a role in cancer cell survival, dissemination
and resistance to drug therapy (45–47).
STAT3 regulates a number of cardinal features of cancer, including
cancer cell growth and resistance to apoptosis, and has been
validated as a drug target for cancer therapy (48). Following treatment with EPDMNQ and
ENDMNQ, the phosphorylation levels of p38 and JNK increased and
those of ERK and STAT3 decreased in a time-dependent manner. These
data suggest that the anti-apoptotic effects of EPDMNQ and ENDMNQ
observed in the present study may be associated with MAPK and
STAT3.
It has been well established that ROS serve
important biological roles in cell homeostasis, but several studies
have also suggested that high intracellular ROS levels are usually
associated with the apoptosis of cancer cells (49–51).
However, ROS accumulation contributes to abnormal changes in
mitochondrial membrane permeability. As a positive feedback loop
between intracellular ROS levels and mitochondria, a large amount
of ROS is released into the cytoplasm during mitochondria-dependent
apoptosis (52). In addition, ROS
accumulation contributes to MAPK activation and inactivates other
potential inhibitory factors to promote tumor cell apoptosis
(53). In the present study,
EPDMNQ- and ENDMNQ-induced apoptosis was accompanied by ROS
accumulation, which was reversed by pretreatment with the ROS
inhibitor NAC. In the present study, EPDMNQ- and ENDMNQ-induced
apoptosis was accompanied by ROS accumulation and mitochondrial
membrane potential depolarization, which was reversed by
pretreatment with the ROS inhibitor NAC in a time-dependent manner.
The expression levels of the pro-apoptotic protein Bax, cle-PARP
and cle-caspase-3 and the anti-apoptotic protein Bcl-2 were also
examined by western blot analysis. EPDMNQ and ENDMNQ treatment
significantly increased the levels of Bax, cle-PARP and
cle-caspase-3 and decreased Bcl-2 protein levels; these effects
were reversed upon NAC pretreatment. In addition, EPDMNQ and ENDMNQ
significantly promoted the phosphorylation of JNK and p38 but
inhibited that of ERK; these effects were also reversed with NAC
pretreatment. These results demonstrated that EPDMNQ and ENDMNQ
induced ROS-mediated apoptosis via the MAPK and STAT3 signaling
pathways. The effects of EPDMNQ and ENDMNQ demonstrated in
vivo should be evaluated in future studies. EPDMNQ and
ENDMNQ-induced apoptosis are in addition to the changes in
intracellular ROS levels already mentioned in the present study,
including many other causes, including cell autophagy, endoplasmic
reticulum stress and DNA damage. The experimental results of the
present study do not fully explain the causes of apoptosis caused
by EPDMNQ and ENDMNQ. Therefore, the other potential mechanisms of
apoptosis, with the exception of ROS, induced by EPDMNQ and ENDMNQ
will be examined in future studies.
Taken together, the results of the present study
demonstrated that 2 novel types of 1,4-naphthoquinone derivatives,
EPDMNQ and ENDMNQ, induced apoptosis through ROS modulation of the
MAPK and STAT3 signaling pathways in Hep3B cells. Therefore, EPDMNQ
and ENDMNQ may be potential chemotherapeutic agents for the
treatment of HCC.
Acknowledgements
Not applicable.
Funding
The present study was funded by the Multigrain
Production and Processing Characteristic Discipline Construction
Project, the Postdoctoral Scientific Research Foundation of
Heilongjiang Province of China (grant no. LBH-Q13132) and the
Scientific Research Foundation of Heilongjiang Provincial Education
Department of China (grant no. 1252HQ007).
Availability of data and materials
The datasets used or analysed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
CHJ and DJZ conceived and designed the study. YW,
YHL, XJP, GNS, LQM, YZ, JRW, JQL, HW, WTX, YL, YZ, YHH; MHJ and YQZ
performed the experiments. TZ, SNW and HNS analyzed the data. YW
and YHL wrote the manuscript. All authors read and approved the
final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Zhang ZQ, Meng H, Wang N, Liang LN, Liu
LN, Lu SM and Luan Y: Serum microRNA 143 and microRNA 215 as
potential biomarkers for the diagnosis of chronic hepatitis and
hepatocellular carcinoma. Diagn Pathol. 9:1352014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Sellam F, Harir N, Khaled MB, Mrabent NM,
Salah R, Benchouk A and Diaf M: Delayed diagnosis of pancreatic
cancer reported as more common in a population of North African
young adults. J Gastrointest Oncol. 6:505–510. 2015.PubMed/NCBI
|
|
3
|
Bruix J, Reig M and Sherman M:
Evidence-based diagnosis, staging, and treatment of patients with
hepatocellular carcinoma. Gastroenterology. 150:835–853. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Li Q, Zhu LZ, Yang RJ and Zhu X: Cytotoxic
activity of anticancer drugs on hepatocellular carcinoma cells in
hypoxic-hyponutritional culture. Int Surg. 99:745–752. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Loureiro R, Magalhães-Novais S, Mesquita
KA, Baldeiras I, Sousa IS, Tavares LC, Barbosa IA, Oliveira PJ and
Vega-Naredo I: Melatonin antiproliferative effects require active
mitochondrial function in embryonal carcinoma cells. Oncotarget.
6:17081–17096. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wang X, Yan Y, Yang L, Li M and Zhong X:
Effect of quercetin on the expression of Bcl-2/Bax apoptotic
proteins in endometrial cells of
lipopolysaccharide-induced-abortion mice. J Tradit Chin Med.
36:737–742. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yang SY, Miah A, Sales KM, Fuller B,
Seifalian AM and Winslet M: Inhibition of the p38 MAPK pathway
sensitises human colon cancer cells to 5-fluorouracil treatment.
Int J Oncol. 38:1695–1702. 2011.PubMed/NCBI
|
|
8
|
Bao H, Guo CG, Qiu PC, Zhang XL, Dong Q
and Wang YK: Long non-coding RNA Igf2as controls hepatocellular
carcinoma progression through the ERK/MAPK signaling pathway. Oncol
Lett. 14:2831–2837. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lu X, Li C, Wang YK, Jiang K and Gai XD:
Sorbitol induces apoptosis of human colorectal cancer cells via p38
MAPK signal transduction. Oncol Lett. 7:1992–1996. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Giachello CN, Fiumara F, Giacomini C,
Corradi A, Milanese C, Ghirardi M, Benfenati F and Montarolo PG:
MAPK/Erk-dependent phosphorylation of synapsin mediates formation
of functional synapses and short-term homosynaptic plasticity. J
Cell Sci. 123:881–893. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Pereira L, Igea A, Canovas B, Dolado I and
Nebreda AR: Inhibition of p38 MAPK sensitizes tumour cells to
cisplatin-induced apoptosis mediated by reactive oxygen species and
JNK. EMBO Mol Med. 5:1759–1774. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Qu J, Zhao M, Teng Y, Zhang Y, Hou K,
Jiang Y, Yang X, Shang H, Qu X and Liu Y: Interferon-α sensitizes
human gastric cancer cells to TRAIL-induced apoptosis via
activation of the c-CBL-dependent MAPK/ERK pathway. Cancer Biol
Ther. 12:494–502. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Atay O and Skotheim JM: Spatial and
temporal signal processing and decision making by MAPK pathways. J
Cell Biol. 216:317–330. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Gong J, Lv L and Huo J: Roles of F-box
proteins in human digestive system tumors (Review). Int J Oncol.
45:2199–2207. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Tasaki S, Horiguchi A, Asano T, Ito K,
Asano T and Asakura H: Docosahexaenoic acid inhibits the
phosphorylation of STAT3 and the growth and invasion of renal
cancer cells. Exp Ther Med. 14:1146–1152. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yuan J, Zhang F and Niu R: Multiple
regulation pathways and pivotal biological functions of STAT3 in
cancer. Sci Rep. 5:176632015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bharadwaj U, Eckols TK, Kolosov M,
Kasembeli MM, Adam A, Torres D, Zhang X, Dobrolecki LE, Wei W,
Lewis MT, et al: Drug-repositioning screening identified
piperlongumine as a direct STAT3 inhibitor with potent activity
against breast cancer. Oncogene. 34:1341–1353. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kron KJ, Murison A, Zhou S, Huang V,
Yamaguchi TN, Shiah YJ, Fraser M, van der Kwast T, Boutros PC,
Bristow RG, et al: TMPRSS2-ERG fusion co-opts master transcription
factors and activates NOTCH signaling in primary prostate cancer.
Nat Genet. 49:1336–1345. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lai ZQ, Ip SP, Liao HJ, Lu Z, Xie JH, Su
ZR, Chen YL, Xian YF, Leung PS and Lin ZX: Brucein D, a naturally
occurring tetracyclic triterpene quassinoid, induces apoptosis in
pancreatic cancer through ROS-associated PI3K/Akt signaling
pathway. Front Pharmacol. 8:9362017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sengupta D, Mazumdar ZH, Mukherjee A,
Sharma D, Halder AK, Basu S and Jha T: Benzamide porphyrins with
directly conjugated and distal pyridyl or pyridinium groups
substituted to the porphyrin macrocycles: Study of the
photosensitising abilities as inducers of apoptosis in cancer cells
under photodynamic conditions. J Photochem Photobiol B.
178:228–236. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sun J, Song B, Ye Z and Yuan J:
Mitochondria targetable time-gated luminescence probe for singlet
oxygen based on a β-diketonate-europium complex. Inorg Chem.
54:11660–11668. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Gonzalez AA, Zamora L, Reyes-Martinez C,
Salinas-Parra N, Roldan N, Cuevas CA, Figueroa S, Gonzalez-Vergara
A and Prieto MC: (Pro)renin receptor activation increases
profibrotic markers and fibroblast-like phenotype through
MAPK-dependent ROS formation in mouse renal collecting duct cells.
Clin Exp Pharmacol Physiol. 44:1134–1144. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Liu XM, Peyton KJ and Durante W: Ammonia
promotes endothelial cell survival via the heme
oxygenase-1-mediated release of carbon monoxide. Free Radic Biol
Med. 102:37–46. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Guerriero E, Sorice A, Capone F,
Napolitano V, Colonna G, Storti G, Castello G and Costantini S:
Vitamin C effect on mitoxantrone-induced cytotoxicity in human
breast cancer cell lines. PLoS One. 9:e1152872014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Coelho-Cerqueira E1, Netz PA, do Canto VP,
Pinto AC and Follmer C: Beyond topoisomerase inhibition: Antitumor
1,4-naphthoquinones as potential inhibitors of human monoamine
oxidase. Chem Biol Drug Des. 83:401–410. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Graciani FS and Ximenes VF:
2-Bromo-1,4-naphthoquinone: A potentially improved substitute of
menadione in Apatone™ therapy. Braz J Med Biol Res. 45:701–710.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ju Woo H, Jun DY, Lee JY, Park HS, Woo MH,
Park SJ, Kim SC, Yang CH and Kim YH: Anti-inflammatory action of
2-carbomethoxy-2,3-epoxy-3-prenyl-1,4-naphthoquinone (CMEP-NQ)
suppresses both the MyD88-dependent and TRIF-dependent pathways of
TLR4 signaling in LPS-stimulated RAW264.7 cells. J Ethnopharmacol.
205:103–115. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
P R KR, Fernandez A, Laila SP, B A, C S S
and V S V: Synthesis, spectral characterization, crystal structure,
cytotoxicity and apoptosis-Inducing activity of two derivatives of
2-hydroxy-1,4-naphthaquinone. Photodiagnosis Photodyn Ther.
17:250–259. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kawiak A and Lojkowska E: Ramentaceone, a
naphthoquinone derived from drosera sp., induces apoptosis by
suppressing PI3K/Akt signaling in breast cancer cells. PLoS One.
11:e01477182016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Liu C, Shen GN, Luo YH, Piao XJ, Jiang XY,
Meng LQ, Wang Y, Zhang Y, Wang JR, Wang H, et al: Novel
1,4-naphthoquinone derivatives induce apoptosis via ROS-mediated
p38/MAPK, Akt and STAT3 signaling in human hepatoma Hep3B cells.
Int J Biochem Cell Biol. 96:9–19. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
P1rachayasittikul V, Pivngaew R,
Worachartcheewan A, Nantasenamat C, Prachayasittikul S, Ruchirawat
S and Prachayasittikul V: Synthesis, anticancer activity and QSAR
study of 1,4-naphthoquinone derivatives. Eur J Med Chem.
84:247–263. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lee JJ, Zhang WY, Yi H, Kim Y, Kim IS,
Shen GN, Song GY and Myung CS: Anti-proliferative actions of
2-decylamino-5,8-dimethoxy-1,4-naphthoquinone in vascular smooth
muscle cells. Biochem Biophys Res Commun. 411:213–218. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lara LS, Moreira CS, Calvet CM, Lechuga
GC, Souza RS, Bourguignon SC, Ferreira VF, Rocha D and Pereira MCS:
Efficacy of 2-hydroxy-3-phenylsulfanylmethyl-[1,4]-naphthoquinone
derivatives against different Trypanosoma cruzi discrete type
units: Identification of a promising hit compound. Eur J Med Chem.
144:572–581. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kishore N, Binneman B, Mahapatra A, van de
Venter M, du Plessis-Stoman D, Boukes G, Houghton P, Marion Meyer
JJ and Lall N: Cytotoxicity of synthesized 1,4-naphthoquinone
analogues on selected human cancer cell lines. Bioorg Med Chem.
22:5013–5019. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Bahman AA, Abaza MSI, Khoushiash SI and
Al-Attiyah RJ: Sequence-dependent effect of sorafenib in
combination with natural phenolic compounds on hepatic cancer cells
and the possible mechanism of action. Int J Mol Med. 42:1695–1715.
2018.PubMed/NCBI
|
|
36
|
You ML, Chen YJ, Chong QY, Wu MM, Pandey
V, Chen RM, Liu L, Ma L, Wu ZS, Zhu T and Lobie PE: Trefoil factor
3 mediation of oncogenicity and chemoresistance in hepatocellular
carcinoma is AKT-BCL-2 dependent. Oncotarget. 8:39323–39344. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhang Y, Li H, Chang H, Du L, Hai J, Geng
X and Yan X: MTP18 overexpression contributes to tumor growth and
metastasis and associates with poor survival in hepatocellular
carcinoma. Cell Death Dis. 9:9562018. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Guamán-Ortiz LM, Orellana MI and
Ratovitski EA: Natural compounds as modulators of non-apoptotic
cell death in cancer cells. Curr Genomics. 18:132–155. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Aden DP, Fogel A, Plotkin S, Damjanov I
and Knowles BB: Controlled synthesis of HBsAg in a differentiated
human liver carcinoma-derived cell line. Nature. 282:615–616. 1979.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Morris KM, Aden DP, Knowles BB and Colten
HR: Complement biosynthesis by the human hepatoma-derived cell line
HepG2. J Clin Invest. 70:906–913. 1982. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Nakabayashi H, Taketa K, Miyano K, Yamane
T and Sato J: Growth of human hepatoma cells lines with
differentiated functions in chemically defined medium. Cancer Res.
42:3858–3863. 1982.PubMed/NCBI
|
|
42
|
Mohammad RM, Muqbil I, Lowe L, Yedjou C,
Hsu HY, Lin LT, Siegelin MD, Fimognari C, Kumar NB, Dou QP, et al:
Broad targeting of resistance to apoptosis in cancer. Semin Cancer
Biol. 35 Suppl:S78–S103. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Panganiban RA, Snow AL and Day RM:
Mechanisms of radiation toxicity in transformed and non-transformed
cells. Int J Mol Sci. 14:15931–15958. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Kodama T, Hikita H, Kawaguchi T, Saito Y,
Tanaka S, Shigekawa M, Shimizu S, Li W, Miyagi T, Kanto T, et al:
The Bcl-2 homology domain 3 (BH3)-only proteins Bim and bid are
functionally active and restrained by anti-apoptotic Bcl-2 family
proteins in healthy liver. J Biol Chem. 288:30009–30018. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Bubici C and Papa S: JNK signalling in
cancer: In need of new, smarter therapeutic targets. Br J
Pharmacol. 171:24–37. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Calvo N, Carriere P, Martin MJ and Gentili
C: RSK activation via ERK modulates human colon cancer cells
response to PTHrP. J Mol Endocrinol. 59:13–27. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Kosako H and Kou M: Global Identification
of ERK substrates by phosphoproteomics based on IMAC and 2D-DIGE.
Methods Mol Biol 1487. 137–149. 2017. View Article : Google Scholar
|
|
48
|
Yao X, Liu H, Zhang X, Zhang L, Li X, Wang
C and Sun S: Cell surface GRP78 accelerated breast cancer cell
proliferation and migration by activating STAT3. PLoS One.
10:e01256342015. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Ishaq M, Kumar S, Varinli H, Han ZJ, Rider
AE, Evans MD, Murphy AB and Ostrikov K: Atmospheric gas
plasma-induced ROS production activates TNF-ASK1 pathway for the
induction of melanoma cancer cell apoptosis. Mol Bio Cell.
25:1523–1531. 2014. View Article : Google Scholar
|
|
50
|
Hempel N and Trebak M: Crosstalk between
Calcium and reactive oxygen species signaling in cancer. Cell
Calcium. 63:70–96. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Hseu YC, Tsai TJ, Korivi M, Liu JY, Chen
HJ, Lin CM, Shen YC and Yang HL: Antitumor properties of Coenzyme
Q0 against human ovarian carcinoma cells via induction
of ROS-mediated apoptosis and cytoprotective autophagy. Sci Rep.
7:80622017. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Kim MO, Moon DO, Jung JM, Lee WS, Choi YH
and Kim GY: Agaricus blazei extract induces apoptosis through
ROS-dependent JNK activation involving the mitochondrial pathway
and suppression of constitutive NF-κB in THP-1 cells. Evid Based
Complement Alternat Med 2011. 8381722011.
|
|
53
|
Sakon S, Xue X, Takekawa M, Sasazuki T,
Okazaki T, Kojima Y, Piao JH, Yagita H, Okumura K, Doi T, et al:
NF-kappaB inhibits TNF-induced accumulation of ROS that mediate
prolonged MAPK activation and necrotic cell death. EMBO J.
22:3898–3909. 2003. View Article : Google Scholar : PubMed/NCBI
|














