Introduction
Dental pulp tissue contains precursor cells with
regenerative potential, which use their stem cell properties upon
pulp injury. They can proliferate and migrate to the damaged
location, differentiate into odontoblasts, and form the reparative
dentin (1,2). Following isolation from dental pulp
and culture in vitro, these cells are termed dental pulp
cells (DPCs) (3,4). DPCs are a heterogeneous population
that can differentiate into a variety of cell types, including
odontoblast-like cells, osteoblast-like cells, chondrocytes,
neural-like cells and adipocytes (1,2). The
differentiation of DPCs into functional odontoblasts is critical
for the production of a mineralized matrix during dentinogenesis
and dental pulp regeneration. Although the differentiation process
involves various mechanisms (5,6), the
potential molecular pathways underlying DPCs differentiation remain
incompletely defined.
Circular RNA (circRNA) is a group of endogenous
non-coding RNAs first detected in 1991, which was initially thought
to be a functionless byproduct of splicing errors (7). With the development of
high-throughput RNA sequencing and bioinformatics, it became clear
that circRNAs were widespread in animal cells (4,8). The
majority of circRNAs are composed of exonic sequences, which are
highly conserved across various species and frequently exhibit
tissue/developmental-stage-specific expression (9–11).
Since circRNAs are not susceptible to digestion by ribonucleases
(RNases), they possess higher stability than linear RNA (8,9).
This property confers a major advantage in utilizing circRNAs as
novel diagnostic markers. Additionally, a number of studies
demonstrated that circRNAs serve an important role in the
regulation of various cancer pathways (12–15).
Additionally, a recent study demonstrated that 19 circRNAs were
frequently upregulated and 5 circRNAs were often downregulated at
each stage of osteoclast differentiation. These circRNAs profiles
suggested that the differentially expressed circRNAs had specific
functions during osteoclastogenesis (16). However, to date, little is known
about the function of circRNAs during odontogenic differentiation
of human dental pulp cells (hDPCs). Specifically, alterations in
the expression profiles of circRNAs during odontogenesis have not
been reported. Therefore, in the present study, the role of
differentially expressed circRNAs during odontogenesis was
investigated.
Materials and methods
Cell culture and odontogenic
induction
Normal human premolars and third molars were
extracted from healthy adults (3 males and 5 females; 12–25 years
of age; recruitment date: 6th of July 2016) at the Department of
Oral and Maxillofacial Surgery at the Affiliate Stomatology
Hospital of Sun Yat-sen University (Guangzhou, China). The patients
were clearly informed and their consent was obtained. The protocols
were approved by the University Ethics Committee (Sun Yat-sen
University; ethics certificate no: ERC-2014-30). hDPCs were
isolated and cultured as previously described (17). Briefly, hDPCs from each of the 8
patients were cultuRed Separately in α-modified Eagle medium
(Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
supplemented with 15% fetal bovine serum (FBS; Gibco; Thermo Fisher
Scientific, Inc.), 10 U/ml penicillin and 10 mg/ml streptomycin
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) and were incubated
at 37°C in an atmosphere of 5% CO2.
For odontogenic differentiation, hDPCs at passage 3
were cultured for 14 days in odontogenic induction medium (10% FBS,
10 mmol/l β-glycerophosphate, 0.2 mmol/l ascorbic acid and 100
nmol/l dexamethasone in α-modified Eagle medium). Control samples
were cultured in 10% FBS medium alone.
To assess odontogenic differentiation, Alizarin Red
S staining and the alkaline phosphatase (ALP) activity test were
conducted as previously reported (18). In Alizarin Red S staining, images
were captured under a fluorescence microscope (Axio Observer Z1;
Zeiss AG, Oberkochen, Germany) and the positively stained nodules
appeared orange/red. ALP activity was measured using an ALP assay
kit (Nanjing Jiancheng Bioengineering Institute, Nanjing, China).
To normalize enzymatic activity, the concentrations of protein were
measured using a bicinchoninic protein assay kit (Wuhan Boshide
Biological Engineering Co., Ltd., Wuhan, China). Furthermore, the
expression of odontoblast-associated genes [dentin
sialophosphoprotein (DSPP) and dentin matrix acidic phosphoprotein
1 (DMP-1)] in hDPCs was monitored by reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
during odontogenic differentiation using the primers listed in
Table I. GAPDH was the internal
control. The data were analyzed using the ΔCq method
(19) and 2−ΔΔCq
represented the relative expression level of mRNAs.
 | Table I.Primer sequences of
odontoblast-associated genes used in reverse
transcription-quantitative polymerase chain reaction. |
Table I.
Primer sequences of
odontoblast-associated genes used in reverse
transcription-quantitative polymerase chain reaction.
| Gene | Primer
sequence | Product size
(bp) |
|---|
| DSPP | F:
5′-TAGCCGAGGAGATGCTTCTTATA-3′ | 180 |
|
| R:
5′-TTACCTTTGCCACTGTCTGATTT-3′ |
|
| DMP-1 | F:
5′-CAGGAGAGACAGCAAGGGTG-3′ | 178 |
|
| R:
5′-GGGGTTATCTCCCCTGGACT-3′ |
|
Preparation of RNA samples
Total RNA was extracted from hDPCs (with or without
odontogenic induction medium) using TRIzol RNA isolation reagent
(Invitrogen; Thermo Fisher Scientific, Inc.). For RNA-sequencing,
in order to enrich the circRNAs, ribosomal RNA was removed by
Mag-Bind technology and linear RNA was digested by RNases from the
samples, according to a previous study (20). Following RNA fragmentation, random
hexamer priming, cDNA synthesis [with SuperScript II Reverse
Transcriptase (Invitrogen; Thermo Fisher Scientific, Inc.) at 42°C
for 50 min followed by 70°C for 15 min for inactivation,
First-Strand Buffer, 0.1 M dithiothreitol, deoxyribonucleotides,
XhoI primer (Thermo Fisher Scientific, Inc.) and DNA
Polymerase I (Promega Corporation, Madison, WI, USA) at 16°C for
2.5 h followed by 70°C for 10 min for inactivation] and adapter
ligation, the integrity of purified products was analyzed with the
Agilent 2100 Bioanalyzer (Agilent Technologies, Inc., Santa Clara,
CA, USA) and the concentration and purity of the samples were
determined using a NanoDrop™ 2000 UV spectrophotometer (Thermo
Fisher Scientific, Inc.) Only RNA samples with absorbance 260/280
ratios between 1.9 and 2.1, and RNA integrity number >8.0 were
used for further analysis. One sample from the mixture of three
individuals was used for RNA sequencing and the remaining five
samples were utilized for RT-qPCR.
circRNA high-throughput
sequencing
Sequencing was performed using Illumina HiSeqseq™
2000 (Illumina, Inc., San Diego, CA, USA). Following raw data
filtering, the CIRI software version 1.2 (21) was applied to predict the circRNAs
from sequencing samples, that were then labeled and annotated
according to their source areas, using circBase (www.circbase.org) (22). P<0.05, or false discovery rate
(FDR) ≤0.001 and fold-change ≥2.0 was considered to indicate a
statistically significant difference.
RT-qPCR
The high-throughput sequencing demonstrated that the
amount of differentially expressed circRNAs was too large to choose
from. In view of our previous study investigating decreased
miR-135b during odontogenic differentiation of hDPCs (23), and the circRNAs' role as miRNA
sponges, a co-expression network of circRNA-miRNA was constructed
following a correlation analysis between the differentially
expressed circRNAs using Cytoscape (www.cytoscape.org) software version 3.5.1. From this,
two differentially expressed circRNAs were selected [both
upregulated: Chr1:172520652 |172548407 (ID: hsa_circ_0015260) and
chr9:130206308|130207528 (hsa_circ_0006984)] to validate the
results obtained with circRNA high-throughput sequencing.
Validation was performed via RT-qPCR assay with SYBR-Green (Qiagen,
Inc., Valencia, CA, USA) on a StepOnePlus™ Real-Time PCR system
(Applied Biosystems; Thermo Fisher Scientific, Inc.). RT was
performed using M-MLV reverse transcriptase (Promega Corporation),
according to the manufacturer's instructions. The PCR cycling
conditions were as follows: 95°C for 2 min, followed by 40 cycles
of 95°C for 15 sec and 60°C for 30 sec. Primers were synthesized by
Guangzhou Forevergen Biosciences Co., Ltd. (Guangdong, China) and
listed in Table II. The RT-qPCR
results were presented as the fold-change of each circRNA in
induced cells relative to control cells, as calculated by the
2−ΔΔCq method. These data were produced from three
biological replicates consisting of five different cell samples
each. The circRNA expression levels were normalized using GAPDH as
the reference gene.
| Table II.Primer sequences of circRNAs used in
reverse transcription-quantitative polymerase chain reaction. |
Table II.
Primer sequences of circRNAs used in
reverse transcription-quantitative polymerase chain reaction.
| Genes | Sequence | Size (bp) |
|---|
|
hsa_circ_0015260 | circ F:
5′-CGCCAGGAACTATTTGATGAGG-3′ | 177 |
|
| circ R:
5′-GTGCGCAGTTGTCATCTTGA-3′ |
|
|
| linear F:
5′-GTCCACCTGACAGCCTTGTT-3′ | 130 |
|
| linear R:
5′-GATTTTGGATCACCACCGCC-3′ |
|
|
hsa_circ_0006984 | circ F:
5′-AGCTCTGCCTTCGTTAGACATC-3′ | 133 |
|
| circ R:
5′-CTTGCTCTGGTGGTGATCCAG-3′ |
|
|
| linear F:
5′-CCCATGTGTAGAGATGCCCC-3′ | 94 |
|
| linear -F:
5′-CCCATGTGTAGAGATGCCCC-3′ |
|
miRNA prediction and functional
analysis
miRanda (issue 2010; www.microrna.org/microrna/) and TargetScan (issue
2011; www.targetscan.org/vert_50/) software were used to
predict the binding sites for human miRNA within these
differentially expressed circRNAs. To further investigate the
biological function and associated pathways of the predicted target
genes, the DAVID Database (issue 2016; david-d.ncifcrf.gov/) was used for Gene Ontology and
the Kyoto Encyclopedia of Genes and Genomes (KEGG) database (issue
2016; www.genome.jp/kegg/) was used for
pathway analysis of these differentially expressed genes.
Statistical analysis
The experiments were repeated three times and the
data are presented as the mean ± standard deviation. SPSS software
version 13.0 (SPSS, Inc., Chicago, IL, USA) was used to analyze the
results. Comparisons between two groups were performed using paiRed
Student's t-test and one-way analysis of variance followed by the
least significant difference multiple-comparison test was used for
multiple group comparisons. P<0.05, or false discovery rate
(FDR) ≤0.001 and fold-change ≥2.0 was considered to indicate a
statistically significant difference.
Results
Odontogenic differentiation of
hDPCs
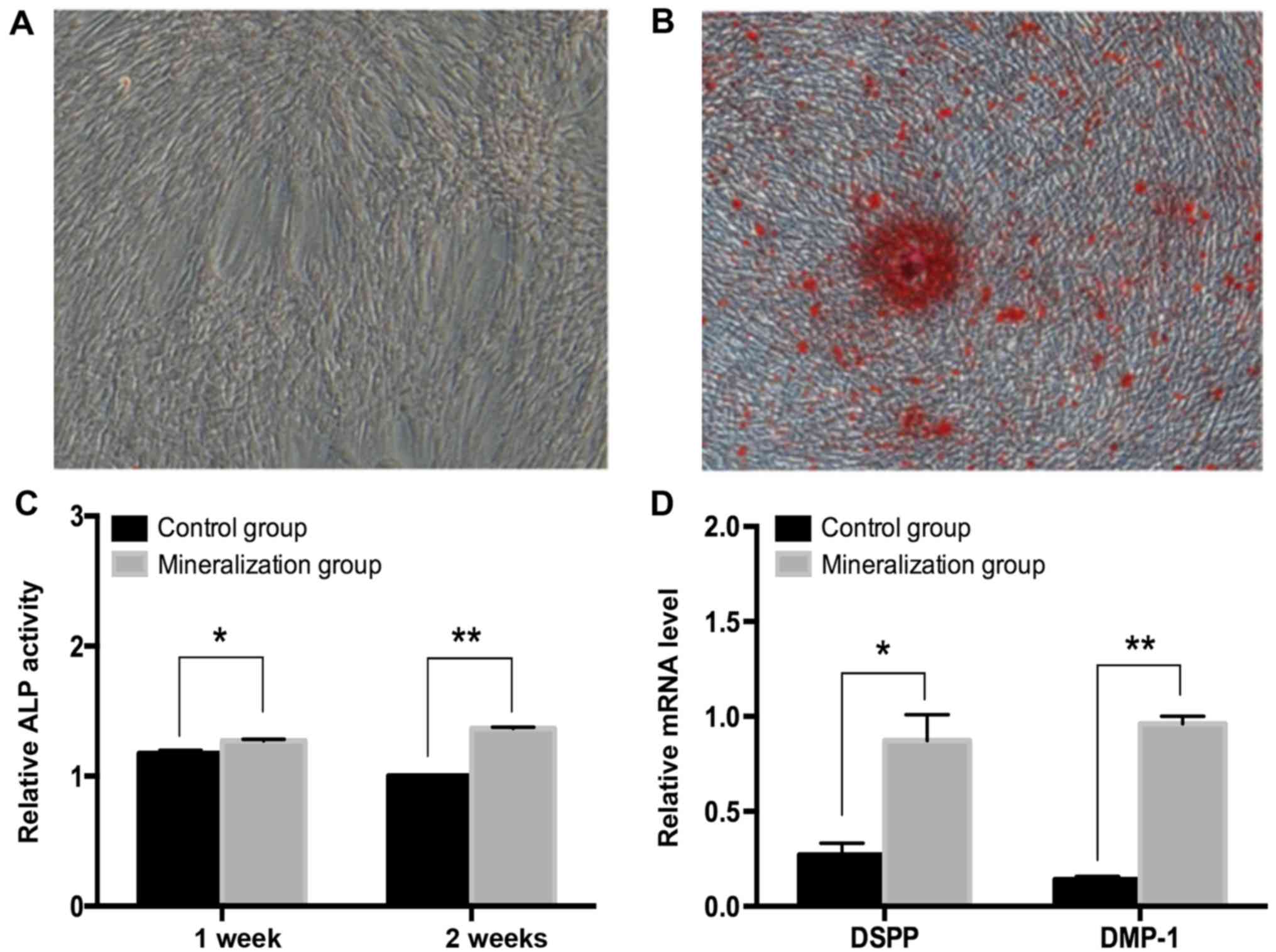
DPCs were incubated in odontogenic induction medium
for 14 days (Fig. 1A). Alizarin
Red S staining positively demonstrated mineralized matrix
deposition in the induced group, which was evident after 14 days
(Fig. 1B). Additionally, the ALP
activity was measured at 7 and 14 days and demonstrated to be
significantly increased (P<0.5; Fig. 1C). The expression of odontoblast
marker genes including DSPP and DMP-1 significantly increased
following day 14 in the induced group (P<0.05; Fig. 1D). These results were confirmed by
RT-qPCR assays.
Expression profiles of circRNAs during
the odontogenic differentiation of hDPCs
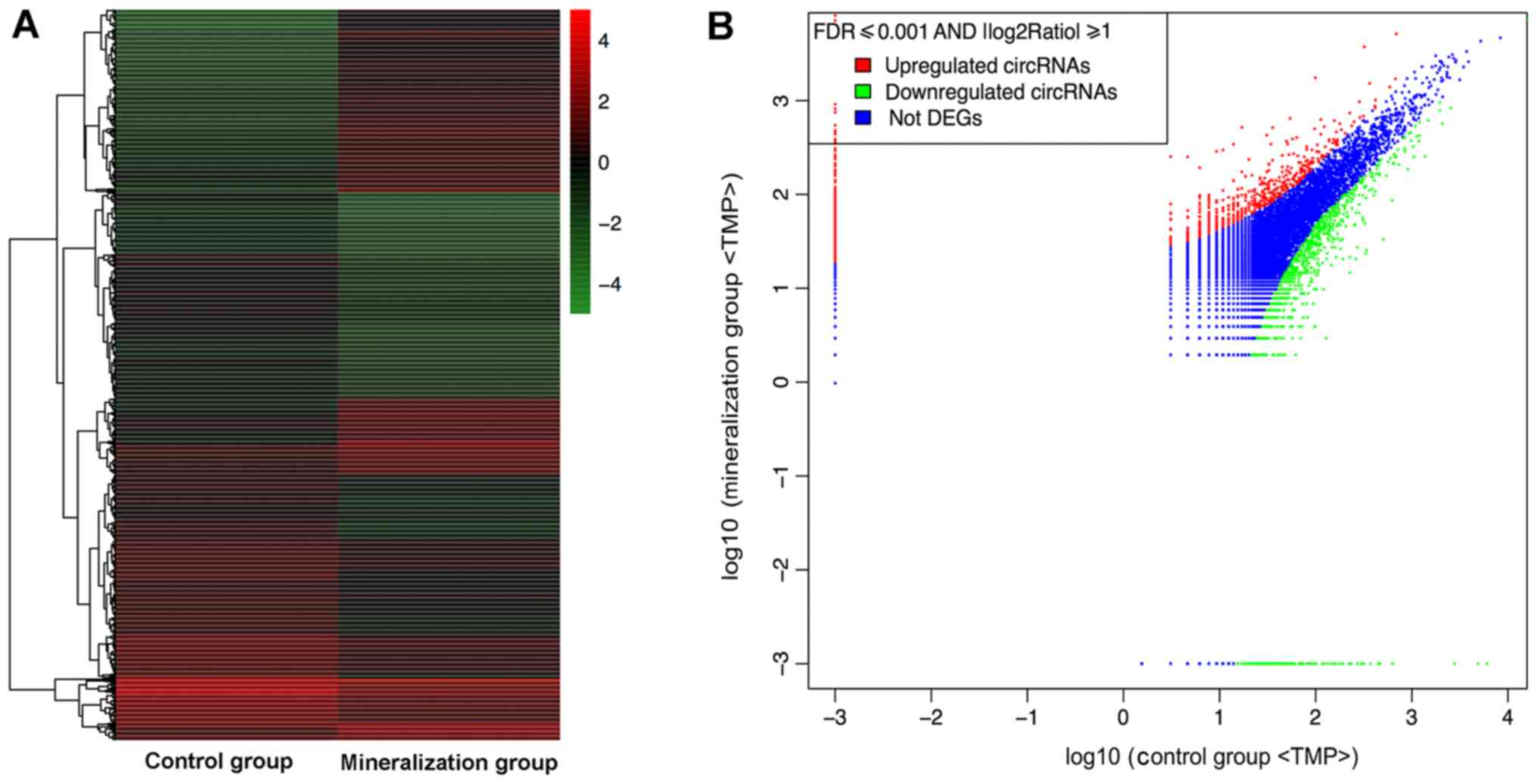
The RNA sequencing results revealed that during
odontogenic induction 1,314 circRNAs were upregulated and 1,780
were downregulated (FDR ≤0.001 and fold-change ≥2.0; Fig. 2).
Validation of circRNA expression
levels by RT-qPCR
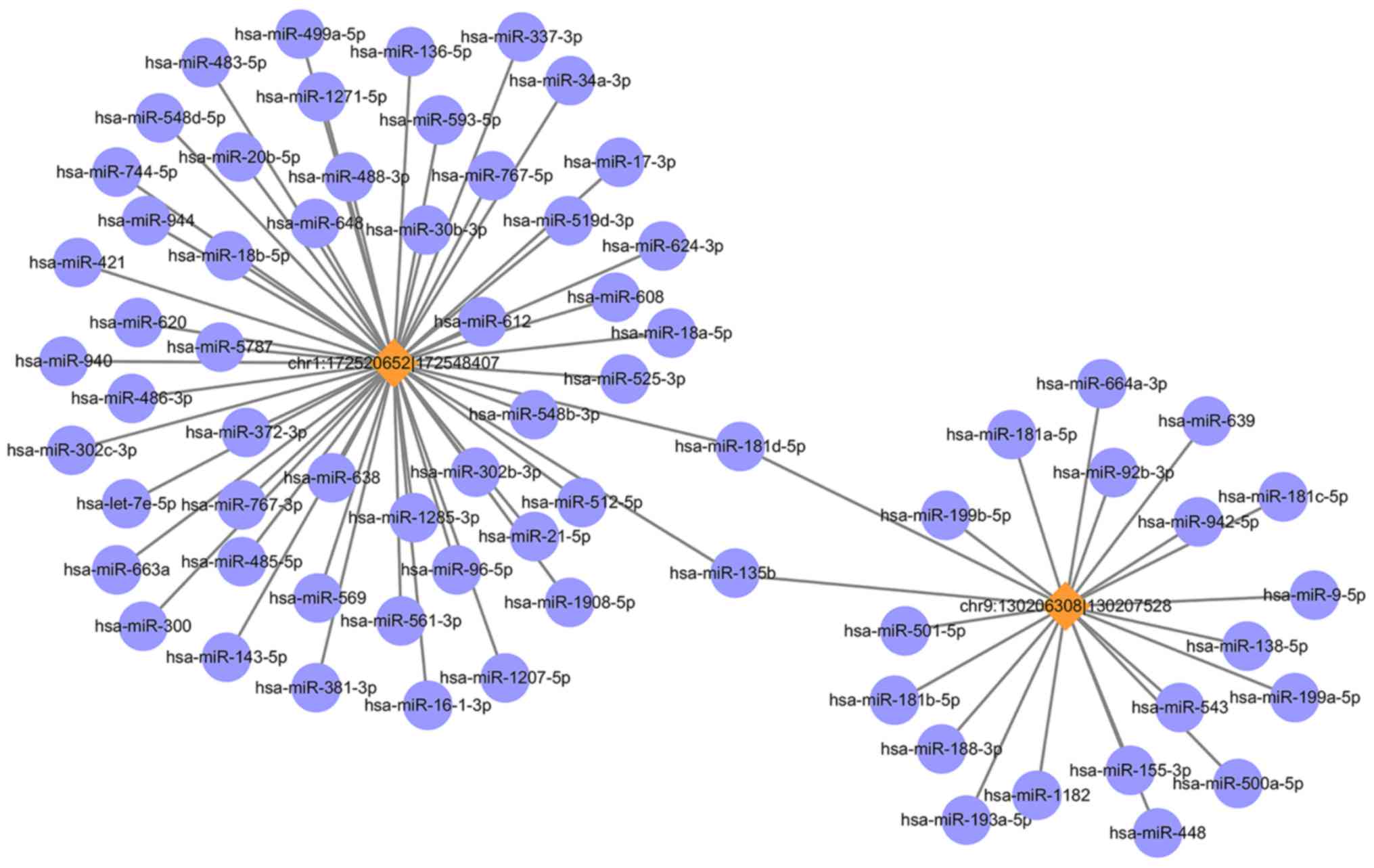
The circRNA-miRNA co-expression network was
constructed containing 2 circRNAs and 71 miRNAs (Fig. 3). In the network, the circles
represent miRNA and the diamonds represent circRNA. The yellow
color represents upregulation. The size of diamond represents the
fold-change of circRNAs with the larger size indicating a higher
fold-change. In the network, the two circRNA i.e., chr1: 172520652
|172548407 (ID: hsa_circ_0015260) and chr9: 130206308|130207528
(hsa_circ_0006984) have been predicted to contain binding sites for
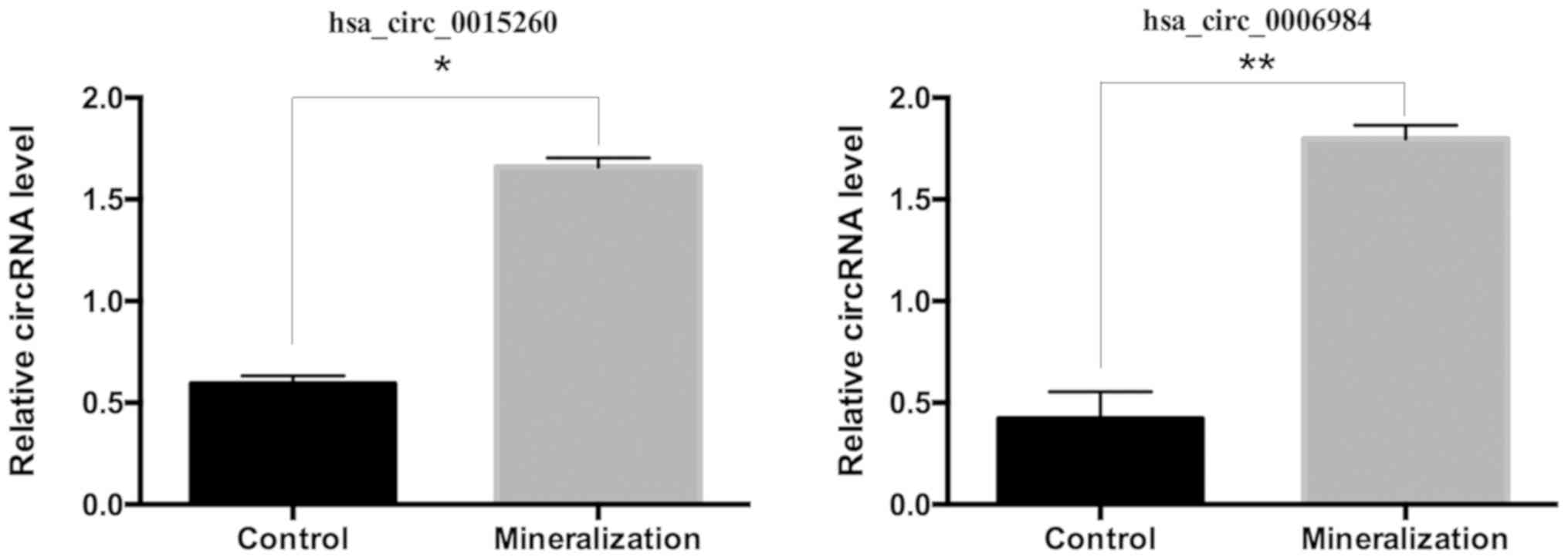
miR-135b. To validate the circRNA results, the RT-qPCR was
performed for the above two circRNAs. These results were consistent
with the RNA sequencing results, as presented in Fig. 4.
miRNA prediction and function
analysis
A total of 2,588 miRNAs were identified as
differentially expressed circRNAs. Through GO analysis, it was
demonstrated that a great quantity of the differentially expressed
circRNA parental genes were associated with important biological
processes, including binding and catalytic activity, molecular
transducer activity, biological regulation, cellular components
organization or biogenesis, cellular process, immune system
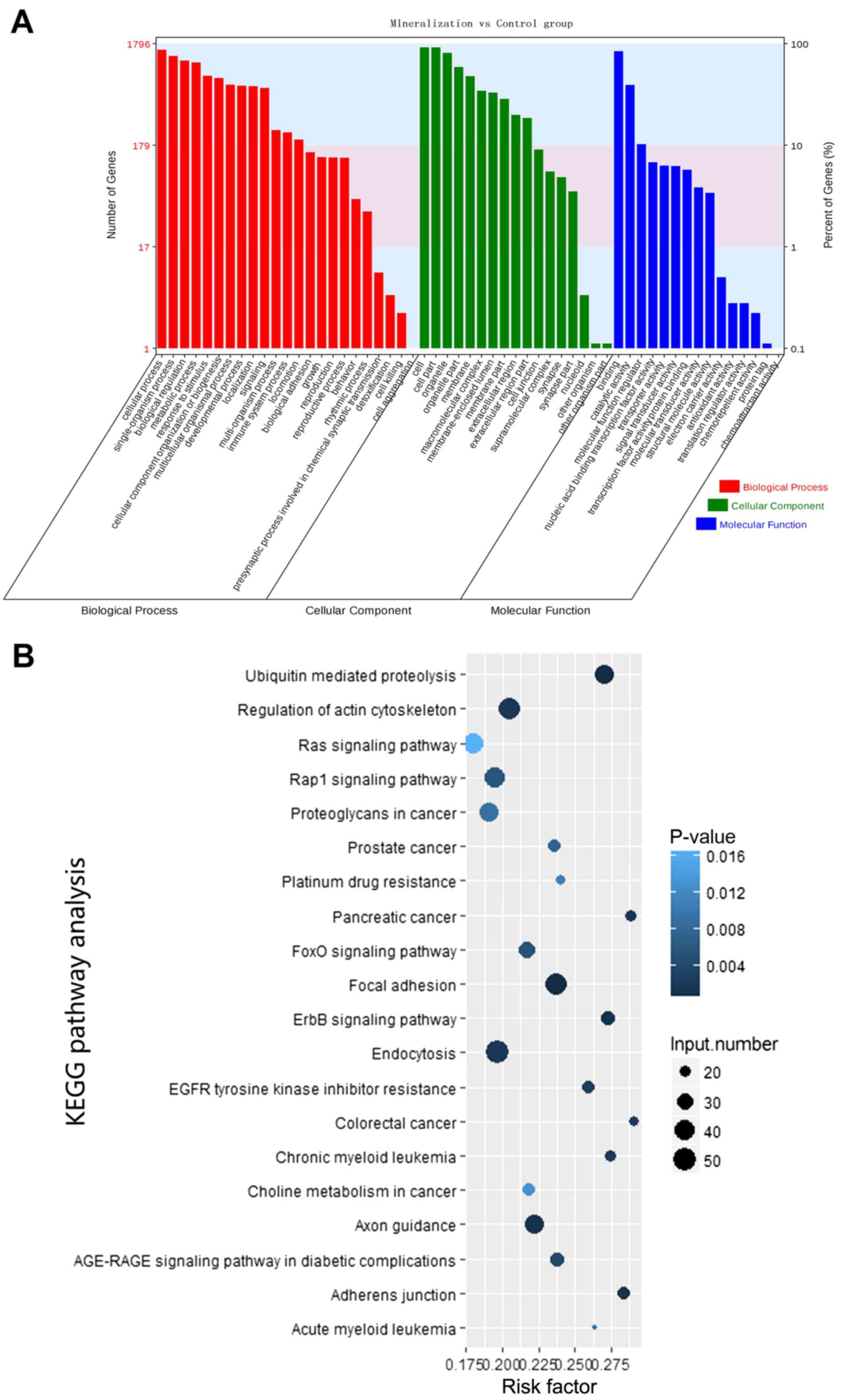
process, metabolic processes and response to stimulus (Fig. 5A). Additionally, KEGG pathway
analysis identified several enriched pathways, including adherens
junction, regulation of actin cytoskeleton, forkhead box protein O1
signaling pathway and mitogen activated protein kinase (MAPK)
signaling pathway, associated with odontogenic differentiation of
hDPCs (Table III). Certain
identified KEGG pathways were consistent with the biological
processes identified by the GO analysis. The top 20 enriched
signaling pathways are shown in Fig.
5B; however, in the present study there were a total of 37
significantly enriched signaling pathways, which included the MAPK
signaling pathway. Therefore, the results from the functional
analysis of the identified circRNAs target genes, using different
bioinformatics approaches, suggest that circRNAs are essential
regulators of the odontogenic differentiation of hDPCs through
their role in key signaling pathways.
| Table III.Enriched KEGG pathways involving
target genes of the differentially expressed circular RNAs during
the odontogenic differentiation of human dental pulp cells. |
Table III.
Enriched KEGG pathways involving
target genes of the differentially expressed circular RNAs during
the odontogenic differentiation of human dental pulp cells.
| Term | KEGG ID | N | P-value |
|---|
| Adherence
junction | hsa04520 | 21 | 0.00115035 |
| Regulation of actin
cytoskeleton | hsa04810 | 44 | 0.00186187 |
| FoxO signaling
pathway | hsa04068 | 29 | 0.00529183 |
| MAPK signaling
pathway | hsa04010 | 38 | 0.04811483 |
Discussion
In previous years, increasing evidence has indicated
that circular RNAs (circRNAs) participate in the regulation of gene
expression and can be used as ideal diagnostic biomarkers for
certain diseases (24–26). A recent study focusing on
osteogenesis identified circ19142 and circ5846 as being associated
with osteoblast differentiation (27). It also reported that BMP2 may
induce osteogenic differentiation through the
circ19142/circ5846-targeted miRNA-mRNA axis. In an investigation of
the circRNA landscape of periodontal ligament stem cells (PDLSCs)
during osteogenesis, a temporal dynamic alteration in the
expression was observed during the osteoblast differentiation and
biomineralization of PDLSCs. This suggested that circRNAs served a
role in osteoblast differentiation and bone formation (28). However, the role of circRNAs in
odontogenic differentiation has never been reported. In the present
study, RNA-Seq was used to screen and compare the 3,094
differentially expressed circRNAs during odontogenesis ofhuman
dental pulp cells (hDPCs). In this analysis, 1,314 circRNAs were
upregulated, while 1,780 circRNAs were downregulated in the
induction group as compared with the control group. Two circRNAs
(hsa_circ_0015260 and hsa_circ_0006984) were selected for RT-qPCR,
which produced results that were consistent with those obtained via
the circRNA high-throughput sequencing. Subsequently, whether the
differentially expressed circRNAs had a regulatory function during
odontogenic differentiation was investigated.
Recently, circRNAs have been proposed to act as
miRNA sponges to regulate gene expression and consequently regulate
linear RNA transcription and protein synthesis (29,30).
The most well-known circRNA is CDR1 antisense RNA, which contains
70 common binding sites and functions as a sponge for miR-7.11
(24). The expression levels of
circRNAs are higher by comparison to their linear isomers and
circRNAs are enriched with miRNA binding sites (31). The results suggested that specific
circRNAs may function as competing endogenous RNAs to promote PDLSC
osteogenic differentiation and periodontal regeneration (32). In addition, circRNAs can chelate
miRNAs more efficiently. Therefore, the roles of circRNAs in
osteoblast differentiation may be associated with their
miRNA-mediated effects. A previous study has demonstrated that
miR-135b was decreased prominently during odontogenic
differentiation of hDPCs (23).
Through target prediction and functional analysis, it was also
hypothesized that the downregulation of miR-135b increased the
expression of APC, myocyte enhancer factor 2C, and collagen type V
α1 chain, subsequently mediating the MAPK and the Wnt signaling
pathways and therefore promoting hDPC odontogenic differentiation
(23). In the present study, the
two RT-qPCR validated circRNAs (hsa_circ_0015260 and
hsa_circ_0006984) increased. In addition, both circRNAs were
enriched with miRNA binding sites for miR-135b, which was
consistent with the hypothesis of the circRNAs acting as miRNA
sponges. Based on the previous studies (23,29,32),
it is reasonable to hypothesize that the upregulation of the
circRNAs hsa_circ_0015260 and hsa_circ_0006984 may competitively
release the inhibition of miR-135b on its target genes, and
consequently mediate the associated signaling pathways, which
ultimately facilitate hDPC odontogenic differentiation. The
specific interaction between the two validated circRNAs and
miR-135b requires further investigation.
From the bioinformatics analyses, the GO analysis
revealed several differentially expressed circRNA parental genes,
which were associated with many important biological processes. The
results suggested that the regulation of the affected genes during
cellular processes was vital during odontogenic differentiation.
Furthermore, the KEGG pathway analysis indicated that the majority
of the identified genes were enriched in several signaling
pathways. Certain pathways, including focal adhesion, regulation of
the actin cytoskeleton and MAPK signaling were associated with
odontogenic differentiation. The results of the present study, in
accordance with previous studies (7–10),
infer that circRNA has a potential role as a regulator of various
biological processes. Among the enriched signaling pathways
identified in this study, MAPK signaling is widely associated with
the differentiation of dental pulp cells and the process of tooth
development (33,34). Woo et al (35) reported that 17-estradiol induced
odontoblastic differentiation via activation of the c-Src/MAPK
signaling pathway in hDPCs. Insulin-like growth factor 1 has been
revealed to promote the proliferation and osteogenic/odontogenic
differentiation of human dental pulp stem cells through MAPK
signaling pathways (36).
Additionally, the magnetic nanofiber scaffold could induce the
stimulation of odontogenesis and pro-angiogenesis of hDPCs through
the Wnt/MAPK/nuclear factor-κB signaling pathway (37). Furthermore, a previous study
observed that the elasticity of the hDPCs cytoskeleton increased
over time and the focal adhesion was enhanced by the odontogenic
medium (38).
In conclusion, in the present study, high-throughput
sequencing was used to screen for differentially expressed circRNAs
during the odontogenic differentiation of hDPCs. In addition, the
potential functions of these circRNAs using GO and KEGG analyses
were examined. The obtained results suggest that the enrichment of
target genes involved in the MAPK signaling pathway is of
particular importance. This study adds to the current understanding
of the regulatory mechanisms underlying the differentiation of
hDPCs.
Acknowledgements
Not applicable.
Funding
This study was supported by the Natural Science
Foundation of Guangdong Province (grant no. 2017A030313713).
Availability of data and materials
The datasets generated and/or analyzed during the
current study are not publicly available due to confidentiality of
another study in our group, but are available from the
corresponding author on reasonable request.
Authors' contributions
HJ conceived the study. CL designed and performed
the experiments, analyzed the data, and wrote the manuscript.
Ethics approval and consent to
participate
The patients were clearly informed and their consent
was obtained. The protocols were approved by the University Ethics
Committee (ethic certificate no: ERC-2014-30).
Patient consent for publication
The patients were clearly informed and their consent
was obtained.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Wang J, Ma H, Jin X, Hu J, Liu X, Ni L and
Ma PX: The effect of scaffold architecture on odontogenic
differentiation of human dental pulp stem cells. Biomaterials.
32:7822–7830. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lambrichts I, Driesen RB, Dillen Y,
Gervois P, Ratajczak J, Vangansewinkel T, Wolfs E, Bronckaers A and
Hilkens P: Dental pulp stem cells: Their potential in reinnervation
and angiogenesis by using scaffolds. J Endod. 43 (Suppl 9):S12–S16.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wang J, Wei X, Ling J, Huang Y, Gong Q and
Huo Y: Identification and characterization of side population cells
from adult human dental pulp after ischemic culture. J Endod.
38:1489–1497. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hilkens P, Gervois P, Fanton Y,
Vanormelingen J, Martens W, Struys T, Politis C, Lambrichts I and
Bronckaers A: Effect of isolation methodology on stem cell
properties and multilineage differentiation potential of human
dental pulp stem cells. Cell Tissue Res. 353:65–78. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Qin W, Yang F, Deng R, Li D, Song Z, Tian
Y, Wang R, Ling J and Lin Z: Smad 1/5 is involved in bone
morphogenetic protein-2-induced odontoblastic differentiation in
human dental pulp cells. J Endod. 38:66–71. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Geng YW, Zhang Z, Liu MY and Hu WP:
Differentiation of human dental pulp stem cells into neuronal by
resveratrol. Cell Biol Int. 41:1391–1398. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Nigro JM, Cho KR, Fearon ER, Kern SE,
Ruppert JM, Oliner JD, Kinzler KW and Vogelstein B: Scrambled
exons. Cell. 64:607–613. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Memczak S, Jens M, Elefsinioti A, Torti F,
Krueger J, Rybak A, Maier L, Mackowiak SD, Gregersen LH, Munschauer
M, et al: Circular RNAs are a large class of animal RNAs with
regulatory potency. Nature. 495:333–338. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Salzman J, Chen RE, Olsen MN, Wang PL and
Brown PO: Cell-type specific features of circular RNA expression.
PLoS Genet. 9:e10037772013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Barrett SP, Wang PL and Salzman J:
Circular RNA biogenesis can proceed through an exon-containing
lariat precursor. Elife. 4:e075402015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Qu S, Yang X, Li X, Wang J, Gao Y, Shang
R, Sun W, Dou K and Li H: Circular RNA: A new star of noncoding
RNAs. Cancer Lett. 365:141–148. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Qi X, Zhang DH, Wu N, Xiao JH, Wang X and
Ma W: ceRNA in cancer: Possible functions and clinical
implications. J Med Genet. 52:710–718. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Qu S, Song W, Yang X, Wang J, Zhang R,
Zhang Z, Zhang H and Li H: Microarray expression profile of
circular RNAs in human pancreatic ductal adenocarcinoma. Genom
Data. 5:385–387. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Huang YS, Jie N, Zou KJ and Weng Y:
Expression profile of circular RNAs in human gastric cancer
tissues. Mol Med Rep. 16:2469–2476. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Liang HF, Zhang XZ, Liu BG, Jia GT and Li
WL: Circular RNA circ-ABCB10 promotes breast cancer proliferation
and progression through sponging miR-1271. Am J Cancer Res.
7:1566–1576. 2017.PubMed/NCBI
|
|
16
|
Dou C, Cao Z, Yang B, Ding N, Hou T, Luo
F, Kang F, Li J, Yang X, Jiang H, et al: Changing expression
profiles of lncRNAs, mRNAs, circRNAs and miRNAs during
osteoclastogenesis. Sci Rep. 6:214992016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wei X, Ling J, Wu L, Liu L and Xiao Y:
Expression of mineralization markers in dental pulp cells. J Endod.
33:703–708. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yu J, He H, Tang C, Zhang G, Li Y, Wang R,
Shi J and Jin Y: Differentiation potential of STRO-1+
dental pulp stem cells changes during cell passaging. BMC Cell
Biol. 11:322010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression using real-time quantitative PCR and the
2−ΔΔCT method. Methods. 25:402–408. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yin QF, Chen LL and Yang L: Fractionation
of non-polyadenylated and ribosomal-free RNAs from mammalian cells.
Methods Mol Biol. 1206:69–80. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Gao Y, Wang J and Zhao F: CIRI: An
efficient and unbiased algorithm for de novo circular RNA
identification. Genome Biol. 16:42015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Glažar P, Papavasileiou P and Rajewsky N:
circBase: A database for circular RNAs. RNA. 20:1666–1670. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Gong Q, Wang R, Jiang H, Lin Z and Ling J:
Alteration of microRNA expression of human dental pulp cells during
odontogenic differentiation. J Endod. 38:1348–1354. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hansen TB, Jensen TI, Clausen BH, Bramsen
JB, Finsen B, Damgaard CK and Kjems J: Natural RNA circles function
as efficient microRNA sponges. Nature. 495:384–388. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lukiw WJ: Circular RNA (circRNA) in
Alzheimer's disease (AD). Front Genet. 4:3072013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Li Z, Huang C, Bao C, Chen L, Lin M, Wang
X, Zhong G, Yu B, Hu W, Dai L, et al: Exon-intron circular RNAs
regulate transcription in the nucleus. Nat Struct Mol Biol.
22:256–264. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Qian DY, Yan GB, Bai B, Chen Y, Zhang SJ,
Yao YC and Xia H: Differential circRNA expression profiles during
the BMP2-induced osteogenic differentiation of MC3T3-E1 cells.
Biomed Pharmacother. 90:492–499. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zheng Y, Li X, Huang Y, Jia L and Li W:
The circular RNA landscape of periodontal ligament stem cells
during osteogenesis. J Periodontol. 88:906–914. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Salmena L, Poliseno L, Tay Y, Kats L and
Pandolfi PP: A ceRNA hypothesis: The rosetta stone of a hidden RNA
language? Cell. 146:353–358. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Chen I, Chen CY and Chuang TJ: Biogenesis,
identification, and function of exonic circular RNAs. Wiley
Interdiscip Rev RNA. 6:563–579. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Shang X, Li G, Liu H, Li T, Liu J, Zhao Q
and Wang C: Comprehensive circular RNA profiling reveals that
hsa_circ_0005075, a new circular RNA biomarker, is involved in
hepatocellular cacinoma development. Medicine. 95:e38112016.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Gu X, Li M, Jin Y, Liu D and Wei F:
Identification and integrated analysis of differentially expressed
lncRNAs and circRNAs reveal the potential ceRNA networks during
PDLSC osteogenic differentiation. BMC Genet. 18:1002017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Miyata H, Genma T, Ohshima M, Yamaguchi Y,
Hayashi M, Takeichi O, Ogiso B and Otsuka K: Mitogen-activated
protein kinase/extracellular signal-regulated protein kinase
activation of cultured human dental pulp cells by low-power
gallium-aluminium-arsenic laser irradiation. Int Endod J.
39:238–244. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhao JW, Gao ZL, Mei H, Li YL and Wang Y:
Differentiation of human mesenchymal stem cells: The potential
mechanism for estrogen-induced preferential osteoblast versus
adipocyte differentiation. Am J Med Sci. 341:460–468. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Woo SM, Seong KJ, Oh SJ, Park HJ, Kim SH,
Kim WJ and Jung JY: 17β-estradiol induces odontoblastic
differentiation via activation of the c-Src/MAPK pathway in human
dental pulp cells. Biochem Cell Biol. 93:587–595. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Lv T, Wu Y, Mu C, Liu G, Yan M, Xu X, Wu
H, Du J, Yu J and Mu J: Insulin-like growth factor 1 promotes the
proliferation and committed differentiation of human dental pulp
stem cells through MAPK pathways. Arch Oral Biol. 72:116–123. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Yun HM, Kang SK, Singh RK, Lee JH, Lee HH,
Park KR, Yi JK, Lee DW, Kim HW and Kim EC: Magnetic nanofiber
scaffold-induced stimulation of odontogenesis and pro-angiogenesis
of human dental pulp cells through Wnt/MAPK/NF-κB pathways. Dent
Mater. 32:1301–1311. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Jones TD, Naimipour H, Sun S, Cho M and
Alapati SB: Mechanical changes in human dental pulp stem cells
during early odontogenic differentiation. J Endod. 41:50–55. 2015.
View Article : Google Scholar : PubMed/NCBI
|