Introduction
Paraquat (1,1′-dimethyl-4,4′-bipyridinium
dichloride; PQ) is a widely used herbicide in agricultural
production, as it possesses fast-acting and nonselective properties
(1); however, the mortality rate
following PQ poisoning is >90%, due to the lack of a specific
antidote or effective therapy (2).
It is known that the redox response is one of the initial factors
involved in the toxic effects of PQ (3). Potent redox responses to PQ induce
the production of a large quantity of reactive oxygen species
(ROS), subsequently resulting in multiple organ injury via lipid
peroxidation (4,5). The lungs are usually one of the most
damaged organs; accumulation of PQ in the lungs induces pulmonary
edema, hemorrhage and alveolar epithelial cell destruction,
ultimately leading to acute lung injury (ALI), one of the most
frequent causes of PQ poisoning-associated mortality (6,7).
Numerous therapeutic strategies have been developed to target the
pathological mechanisms of PQ poisoning; however, their clinical
efficacies have not been satisfactory (8–10).
Thioredoxin (Trx) interacting protein (TXNIP) has
been reported to interact with and inhibit Trx, which can modulate
cellular ROS by reducing disulfides into thiol groups (11); however, under conditions of
oxidative stress, the elevation of ROS levels induces the
dissociation of TXINP from Trx, enabling Trx to contribute to ROS
regulation (12). Of note, certain
studies have demonstrated that TXNIP is involved in the activation
of NLR pyrin domain containing 3 (NLRP3) (13). NLRP3, the best characterized
inflammasome, consists of NLRP and the adaptor protein,
apoptosis-associated speck-like protein containing caspase-1
activator domain (14). When the
inflammasome is activated, caspase-1 is cleaved to generate the
active form of caspase-1, which in turn cleaves interleukin (IL)-1β
and IL-18 into their active forms; these proinflammatory cytokines
are subsequently involved in inflammation (15). Activation of the NLRP3 inflammasome
has been demonstrated to serve an important role in PQ-induced lung
damage (16). Therefore, NLRP3 may
be a promising therapeutic target to treat PQ-induced ALI.
Curcumin (C21H20O6)
is a natural polyphenolic compound, which can be extracted from the
powdered rhizome of the plant Curcuma longa L. Increasing
evidence has indicated that curcumin possesses potent antioxidative
and anti-inflammatory properties, with minimal side effects in
humans (17–19). Notably, the inhibitory effects of
curcumin on NLRP3-mediated inflammatory pathways have been
reported; for example, Gong et al (20) demonstrated that curcumin
effectively improves dextran sulfate sodium (DSS)-induced colitis
in mice by inhibiting activation of the NLRP3 inflammasome; this
study also reported that inhibiting the NLRP3 inflammasome in
vivo with a specific NLRP3 inhibitor notably abrogates the
additional inhibitory effects of curcumin on DSS-induced
inflammatory bowel disease. Similarly, in fructose-induced hepatic
inflammation, curcumin markedly inhibits NLRP3 inflammasome
activation by upregulating microRNA-200a in mice, subsequently
alleviating hepatic inflammation (21). Therefore, curcumin may be a
promising drug for the treatment of NLRP3 inflammasome-associated
inflammatory disease. Due to the central role of NLRP3 activation
in PQ-induced ALI, the present study aimed to investigate whether
curcumin could regulate activation of the NLRP3 inflammasome to
improve PQ-induced lung damage.
Materials and methods
Cell culture
Normal lung fibroblasts [WI-38VA13; American Type
Culture Collection (ATCC®) CCL-75.1™] were obtained from
the ATCC. Cells were cultured in Minimum Essential Medium (MEM;
ATCC® 30-2003™) and 10% fetal bovine serum (FBS;
ATCC® 30-2020™; both ATCC) in a humidified atmosphere
consisting of 5% CO2 at 37°C.
MTT assay
The effects of PQ or curcumin on the viability of
WI-38VA13 cells were determined using an MTT assay (cat. no. M2128;
Sigma-Aldrich; Merck KGaA) as previously described (22). Briefly, a single-cell suspension
was incubated in 96-well plates at a cell density of
4×103 cells/well and supplemented with MEM containing
10% FBS. To determine the appropriate experimental concentrations
of curcumin and PQ, cells were treated with a series of
concentrations of curcumin (0, 100, 300 and 600 µmol/l; cat. no.
08511; Sigma-Aldrich; Merck KGaA) or PQ (0, 5, 10 and 20 µmol/l;
cat. no. 36541; Sigma-Aldrich; Merck KGaA) for 0, 24 or 48 h prior
to cell viability detection. Subsequently, 10 µl MTT solution (5
mg/ml) was added to each well, and the plates were incubated at
37°C for a further 4 h. After removing the supernatant, DMSO (cat.
no. D2650; Sigma-Aldrich; Merck KGaA) was added to dissolve the
formazan crystals. The absorbance was detected at 450 nm using a
microplate reader (Shanghai Cany Precision Instrument Co.,
Ltd.).
Effects of curcumin on PQ-induced
WI-38VA13 cell injury
WI-38VA13 cells in the logarithmic growth phase were
seeded in culture flasks, and were adjusted to a concentration of
1–2×104 cells/ml in MEM (500 µl). The following day, the
medium was replaced with MEM containing 2% FBS. The cells were
divided into four groups: Group 1 (Control) was treated with normal
medium (MEM containing 2% FBS) for 96 h; Group 2 (PQ) was exposed
to PQ (10 µmol/l) for 48 h and then transferred to complete medium
(MEM containing 10% FBS) for a further 48-h incubation; Group 3 (PQ
+ Cur) was exposed to PQ (10 µmol/l) for 48 h, followed by
treatment with 300 µmol/l curcumin for 48 h; and Group 4 (Cur) was
cultured with normal medium for 48 h and then treated with curcumin
(300 µmol/l) for 48 h. All groups were incubated in a humidified
atmosphere consisting of 5% CO2 at 37°C. The cells were
collected at three time points (0, 24 and 48 h) following
completion of the 96 h treatment of cells for further MTT assays.
Other subsequent experiments were conducted ≥48 h following
completion of the 96-h treatment of cells.
ROS detection assay
WI-38VA13 cells were seeded in 96-well plates
(4×103 cells/well) following various treatments.
Following incubation for 48 h, cells were trypsinized and
harvested. A Reactive Oxygen Species Assay kit (Beyotime Institute
of Biotechnology, cat. no. S0033) was used to determine the ROS
levels. Briefly, cells were cultured with complete medium
containing dichlorodihydrofluorescein diacetate at a final
concentration of 10 µmol/l at 37°C for 20 min. The cells were then
harvested and washed with PBS at least three times. ROS levels were
detected using a FACSCalibur flow cytometer (BD Biosciences), using
BD CellQuest™ Pro Software version 5.1 (BD Biosciences).
Cell apoptosis analysis
Cells from each group were transferred to 96-well
plates (4×103 cells/well). Cells were incubated for 48
h, trypsinized and harvested by centrifugation at 3,000 × g for 5
min at 4°C. The apoptosis rates were determined by Annexin-V/PI
(propidine iodide) double-stain assay (BD Biosciences). The cells
were resuspended in binding buffer containing 5 µl Annexin V-FITC.
Cells were stained in the dark for 15 min at room temperature.
Subsequently, 5 µl PI was added 5 min before the assay, and the
apoptotic rates of the treated WI-38VA13 cells were determined
using a FACS Calibur flow cytometer (BD Biosciences), using BD
CellQuest™ Pro Software version 5.1 (BD Biosciences).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from treated WI-38VA13 cells
using TRIzol® reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) and was then purified using RNase-free DNase
(Takara Biotechnology Co., Ltd.). A PrimeScript™ First Strand cDNA
Synthesis kit (Takara Biotechnology Co., Ltd.) was used to
synthesize cDNA as previously described (23) under the following conditions: 65°C
for 5 min, 30°C for 6 min and 50°C for 50 min. Sequences of the
specific primers are presented in Table I. An ABI 7500 Fast Real-Time PCR
system (Applied Biosystems; Thermo Fisher Scientific, Inc.) was
used to perform qPCR reactions using a reaction mixture containing
1 µl forward and reverse primers (10 µmol/l), 10 µl SYBR
fluorescent dye (Applied Biosystems; Thermo Fisher Scientific,
Inc.), 2 µl cDNA and RNase-free dH2O. The thermocycling
parameters were as follows: Initial denaturation at 94°C for 3 min,
followed by repeated amplification for 40 cycles at 94°C for 30
sec, 56°C for 30 sec and 72°C for 2 min, prior to a final step at
72°C for 10 min. GAPDH served as an internal control. The relative
mRNA expression levels were calculated using the 2−ΔΔCq
method (24).
 | Table I.Primers for reverse
transcription-quantitative polymerase chain reaction. |
Table I.
Primers for reverse
transcription-quantitative polymerase chain reaction.
| Gene name | Primer
sequences |
|---|
| Bax | Forward:
5′-GATGCGTCCACCAAGAAG-3′ |
|
| Reverse:
5′-AGTTGAAGTTGCCGTCAG-3′ |
| Bcl-2 | Forward:
5′-GTTCCCTTTCCTTCCATCC-3′ |
|
| Reverse:
5′-TAGCCAGTCCAGAGGTGAG-3′ |
| GAPDH | Forward:
5′-ATGGCACCGTCAAGGCTGAG-3′ |
|
| Reverse:
5′-TGTCAGGTACGGTAGTGACG-3′ |
| TXNIP | Forward:
5′-GCTCAATCATGGTGATGTTCAAG-3′ |
|
| Reverse:
5′-CTTCACACACTTCCACTGTCAC-3′ |
| NLRP3 | Forward:
5′-GGAGAGACCTTTATGAGAAAGCAA-3′ |
|
| Reverse:
5′-GCTGTCTTCCTGGCATATCACA-3′ |
| Notch1 | Forward:
5′-CACCCATGACCACTACCCAGTT-3′ |
|
| Reverse:
5′-CCTCGGACCAATCAGAGATGTT-3′ |
Enzyme-linked immunosorbent assay
(ELISA)
The treated WI-38VA13 cells were collected by
centrifugation at 300 × g for 7 min at 4°C. After washing with PBS,
the cells were lysed using cell extraction buffer (cat. no.
FNN0011; Thermo Fisher Scientific, Inc.) with PMSF (1 mM) and
protease inhibitor cocktail (cat. no. P-2714; Sigma-Aldrich; Merck
KGaA). Subsequently, the mixture was incubated at 4°C for 30 min.
After centrifuging the cells at 13,000 × g for 10 min at 4°C, the
supernatant was removed. IL-1β and IL-18 levels were determined
using IL-1β and IL-18 assay kits (cat. nos. H002 and H015; Nanjing
Jiancheng Bioengineering Institute Co., Ltd.), according to the
manufacturer's protocols. The primary IL-1β and IL-18 primary
antibodies were added to the wells. The prepared standard liquid
and diluted supernatant (1:4) were added to ELISA plates. The
horseradish peroxidase (HRP)-labeled secondary antibodies were
added to each well, and then the plate was incubated in a
humidified box at 37°C for 60 min. After the solution was
discarded, substrate solution was added to each well, and the
optical density was measured using a spectrophotometer (cat. no.
722S; Shanghai Precision & Scientific Instrument Co.,
Ltd.).
Western blotting
Total protein was extracted from WI-38VA13 cells
using RIPA solution (Sigma-Aldrich; Merck KGaA), and then
quantified via a bicinchoninic acid protein assay (Takara
Biotechnology Co., Ltd.). Equal quantities of total protein (30 µg)
were separated by 12% SDS-PAGE and then transferred to PVDF
membranes (EMD Millipore). After blocking in 5% non-fat milk in
TBS-Tween for 1 h at room temperature, membranes were incubated at
4°C overnight with the following primary antibodies (all from Cell
Signaling Technology, Inc.): Anti-Bcl-2 (rabbit; 28 kDa; 1:1,000;
cat. no. 2872); anti-Bax (rabbit; 20 kDa; 1:1,000; cat. no. 2774);
anti-cleaved caspase-1 (rabbit; 20 kDa; 1:1,000; cat. no. 4199);
anti-caspase-1 (rabbit; 48 kDa; 1:1,000; cat. no. 3866); anti-NLRP3
(rabbit; 110 kDa; 1:1,000; cat. no. 15101), anti-TXINP (rabbit; 55
kDa; 1:1,000; cat. no. 14715); anti-Notch1 (rabbit; 120 kDa;
1:1,000; cat. no. 4380), anti-phosphorylated (p)-extracellular
signal-regulated kinase 1/2 (p-ERK1/2; rabbit; 42 and 44 kDa;
1:1,000; cat. no. 9101); anti-ERK1/2 (rabbit; 42 and 44 kDa;
1:1,000; cat. no. 9102); and anti-GAPDH (rabbit; 37 kDa; 1:1,000;
cat. no. 5174). Subsequently, membranes were incubated with
HRP-conjugated goat anti-rabbit immunoglobulin G secondary
antibodies (1:2,000; cat. no. 7074; Cell Signaling Technology,
Inc.) for 2 h at room temperature. Proteins were detected using
Pierce™ ECL western blotting substrate (cat. no. 32106; Thermo
Fisher Scientific, Inc.). Optical band density was semi-quantified
using Bio-Rad ChemiDoc™ XRS+ System with Image Lab™ Software
version 4.1 (Bio-Rad Laboratories, Inc.). GAPDH was used as an
internal control.
Statistical analysis
Statistically significant differences among three or
more groups were analyzed by one-way analysis of variance followed
by Tukey's post hoc test, using SPSS version 20 (IBM Corp.). Each
experiment was repeated three times. All data were presented as the
mean ± standard error of the mean. P<0.05 was considered to
indicate a statistically significant difference.
Results
Curcumin mitigates PQ-induced
apoptosis of WI-38VA13 cells
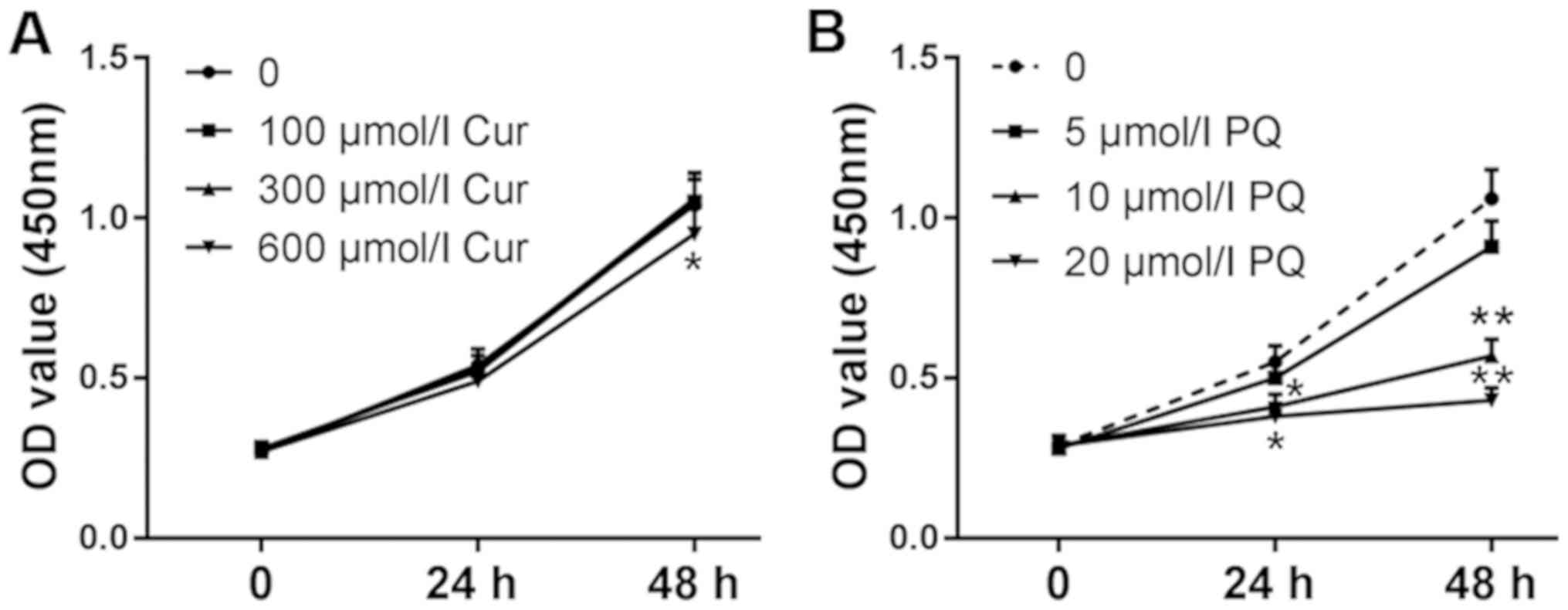
The viability of WI-38VA13 cells was detected at 24
and 48 h. As shown in Fig. 1A,
cell viability was unchanged following treatment with 300 µmol/l
curcumin; however, a decrease in viability was observed in response
to treatment with 600 µmol/l curcumin for 48 h compared with in the
control group. In addition, the effects of 10 µmol/l PQ on cell
viability were more noticeable compared with 5 µmol/l PQ (Fig. 1B). Therefore, 10 µmol/l PQ was
selected to induce lung cell injury and the positive effects of 300
µmol/l curcumin were assessed on the toxicity of PQ. As shown in
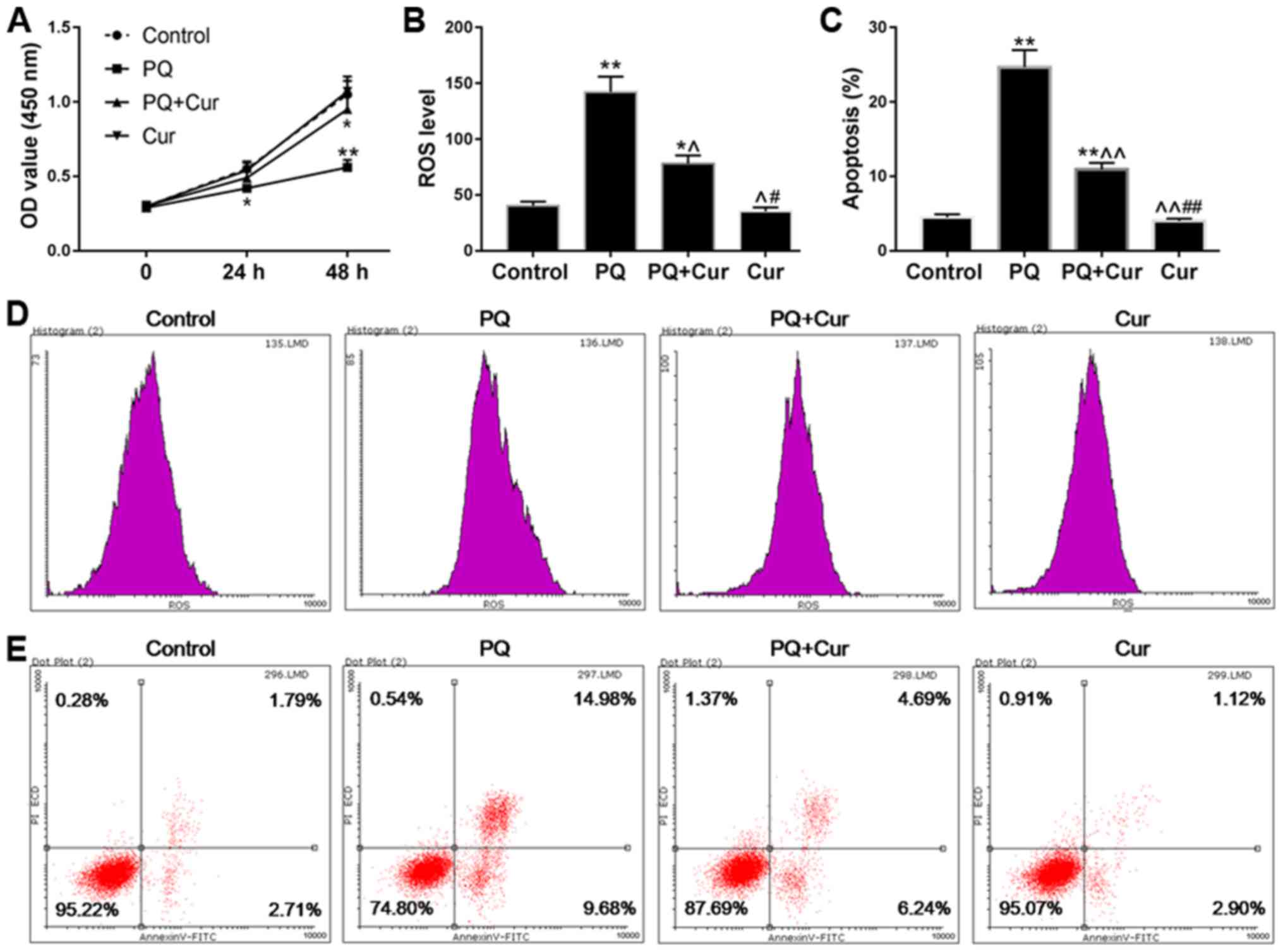
Fig. 2A, the viability of cells
treated with PQ was significantly reduced compared with in the
control group, whereas the cell viability of the PQ + Cur group was
higher than that of the PQ group. In addition, ROS levels and
apoptosis were measured by flow cytometry. As predicted, treatment
with PQ promoted the production of ROS and significantly increased
apoptosis (Fig. 2B and C). As
shown in Fig. 2B-E, PQ treatment
induced ROS generation and increased the percentage of apoptotic
cells from 4.50 to 24.68% (P<0.01) compared with in the control
group. Conversely, the apoptosis of cells treated with curcumin
alone was no different from that in the control group. Furthermore,
compared with in the PQ-treated group, the ROS levels were
significantly reduced in the PQ + Cur group (P<0.05), and the
percentage of apoptosis fell to 10.92%. These results indicated
that curcumin may be able to greatly reduce PQ-induced ROS
production and apoptosis; however, curcumin could not completely
protect lung cells against PQ-induced damage.
Curcumin suppresses
apoptosis-associated protein expression in PQ-damaged WI-38VA13
cells
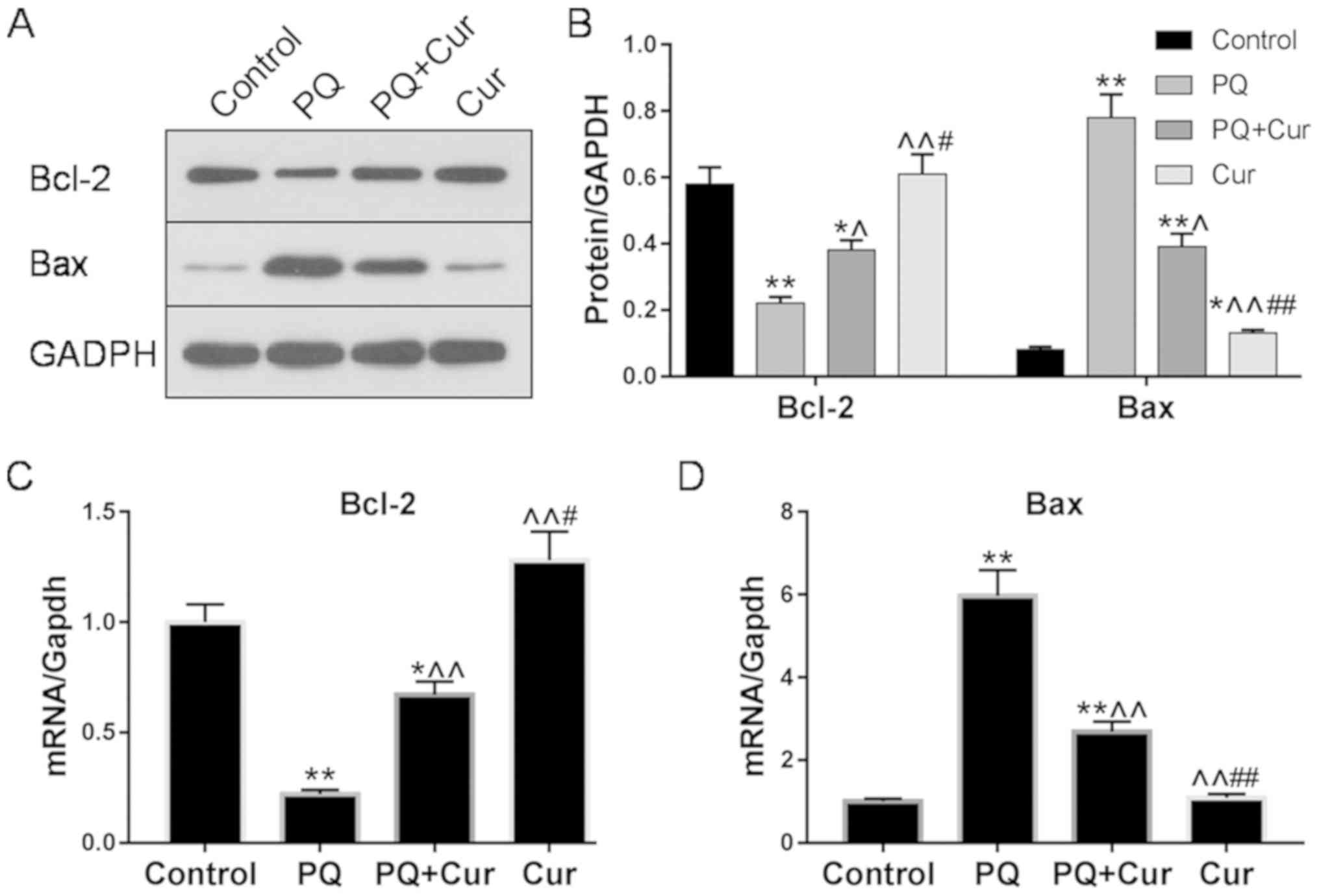
The expression levels of the apoptosis-associated
proteins Bcl-2 and Bax in WI-38VA13 cells were determined via
western blotting and RT-qPCR. Significantly upregulated Bax mRNA
and protein levels were detected following treatment of cells with
10 µmol/l PQ for 48 h compared with the control; however, treatment
with PQ + Cur significantly decreased the expression levels of Bax
(Fig. 3A-C). Bcl-2, an
anti-apoptotic protein, was significantly downregulated by PQ,
whereas curcumin inhibited these effects and significantly
increased the expression of Bcl-2 (Fig. 3A-C). Conversely, compared with the
effects of curcumin on the protein expression levels of Bax and
Bcl-2 in PQ-damaged cells (PQ + Cur group), WI-38VA13 cells treated
with curcumin alone (Cur group) exhibited a more upregulated Bcl-2
levels and downregulated Bax levels.
Curcumin inhibits the activated
TXNIP/NLRP3 axis in cultured PQ-treated WI-38VA13 cells
An increasing number of studies have reported that
inflammation serves an important role in lung injury (25,26).
To investigate whether curcumin reduced lung inflammation induced
by PQ, the expression levels of TXNIP, NLRP3, cleaved caspase-1 and
caspase-1 were evaluated by western blotting and RT-qPCR. As
presented in Fig. 4A-D,
significant upregulation of TXNIP, NLRP3 and cleaved caspase-1 was
observed in the PQ-treated group compared with the control;
however, there was no significant change observed in caspase-1
expression. Additionally, following treatment of lung cells with
curcumin, the mRNA and protein expression levels of TXNIP and NLRP3
were significantly decreased compared with PQ treatment alone,
whereas the effects on cleaved caspase-1 levels were only slightly
attenuated.
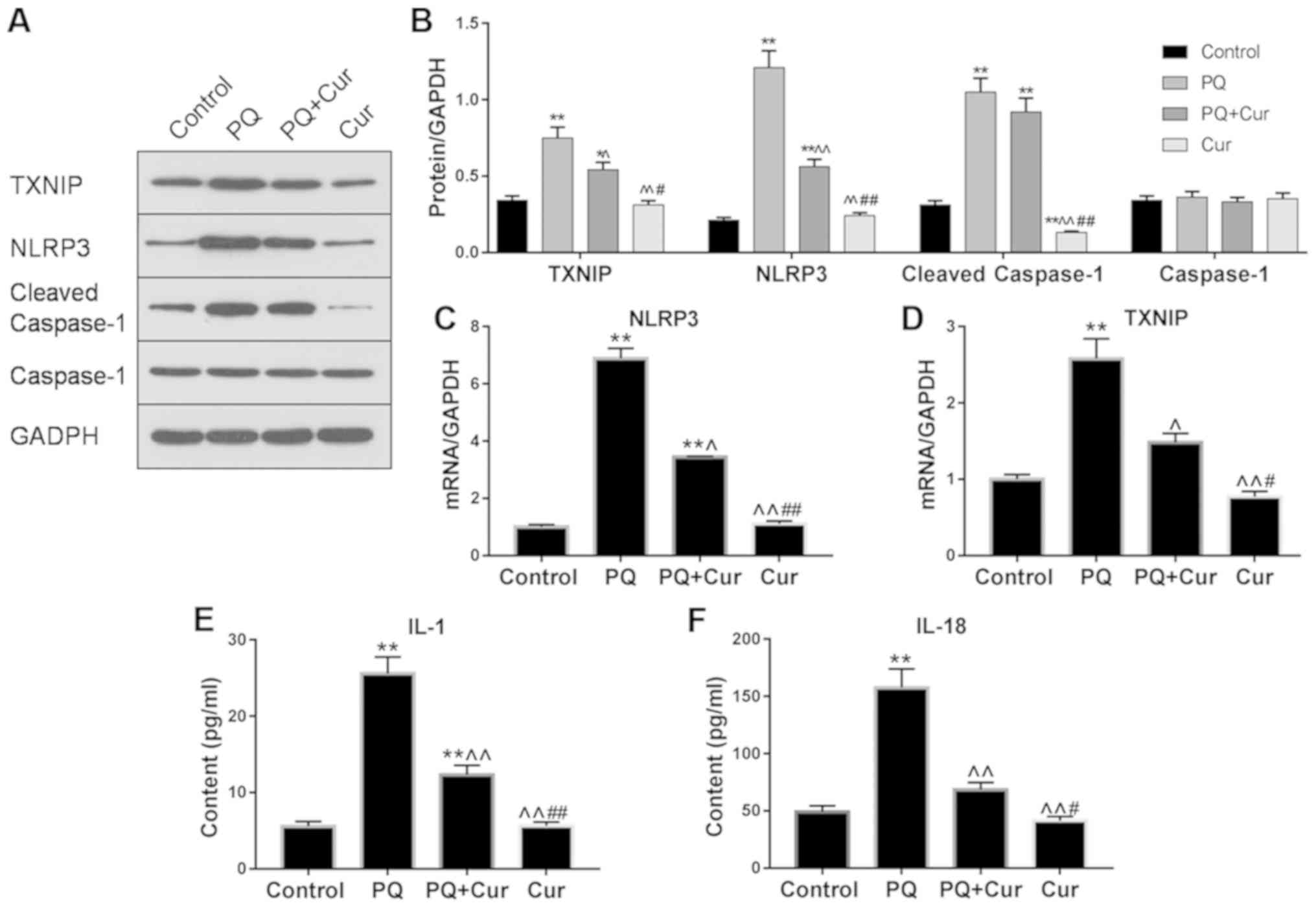 | Figure 4.Curcumin inhibits the TXNIP/NLRP3
axis and proinflammatory cytokine release in PQ-treated WI-38VA13
cells. PQ (10 µmol/l) and curcumin (300 µmol/l) were used to study
the mechanisms underlying the effects of curcumin on PQ-induced
lung cell injury. (A and B) Protein levels of TXNIP, NLRP3, cleaved
caspase-1 and caspase-1 as determined via western blotting. mRNA
levels of (C) NLRP3 and (D) TXNIP were detected via reverse
transcription-quantitative polymerase chain reaction. GAPDH was
used as an internal control. Levels of (E) IL-1β and (F) IL-18 were
quantified by enzyme-linked immunosorbent assay. Data are presented
as the mean ± standard error of the mean (n=3). *P<0.05,
**P<0.01 vs. control; ^P<0.05,
^^P<0.01 vs. PQ; #P<0.05,
##P<0.01 vs. PQ + Cur. Cur, curcumin; IL,
interleukin; NLRP3, NLR pyrin domain containing 3; PQ, paraquat;
TXNIP, thioredoxin interacting protein. |
To further determine whether curcumin affected
PQ-induced activation of proinflammatory cytokines, the levels of
IL-1β and IL-18 in cell extracts were determined by ELISA. It was
revealed that PQ treatment alone significantly increased IL-1β and
IL-18 levels compared with the control; however, treatment with
curcumin significantly suppressed the elevated levels of these
proinflammatory cytokines (Fig.
4E). Collectively, these results demonstrated that curcumin
suppressed PQ-induced NLRP3 activation and downstream inflammatory
pathways.
Involvement of signaling pathways in
the effects of curcumin on PQ-induced WI-38VA13 cell injury
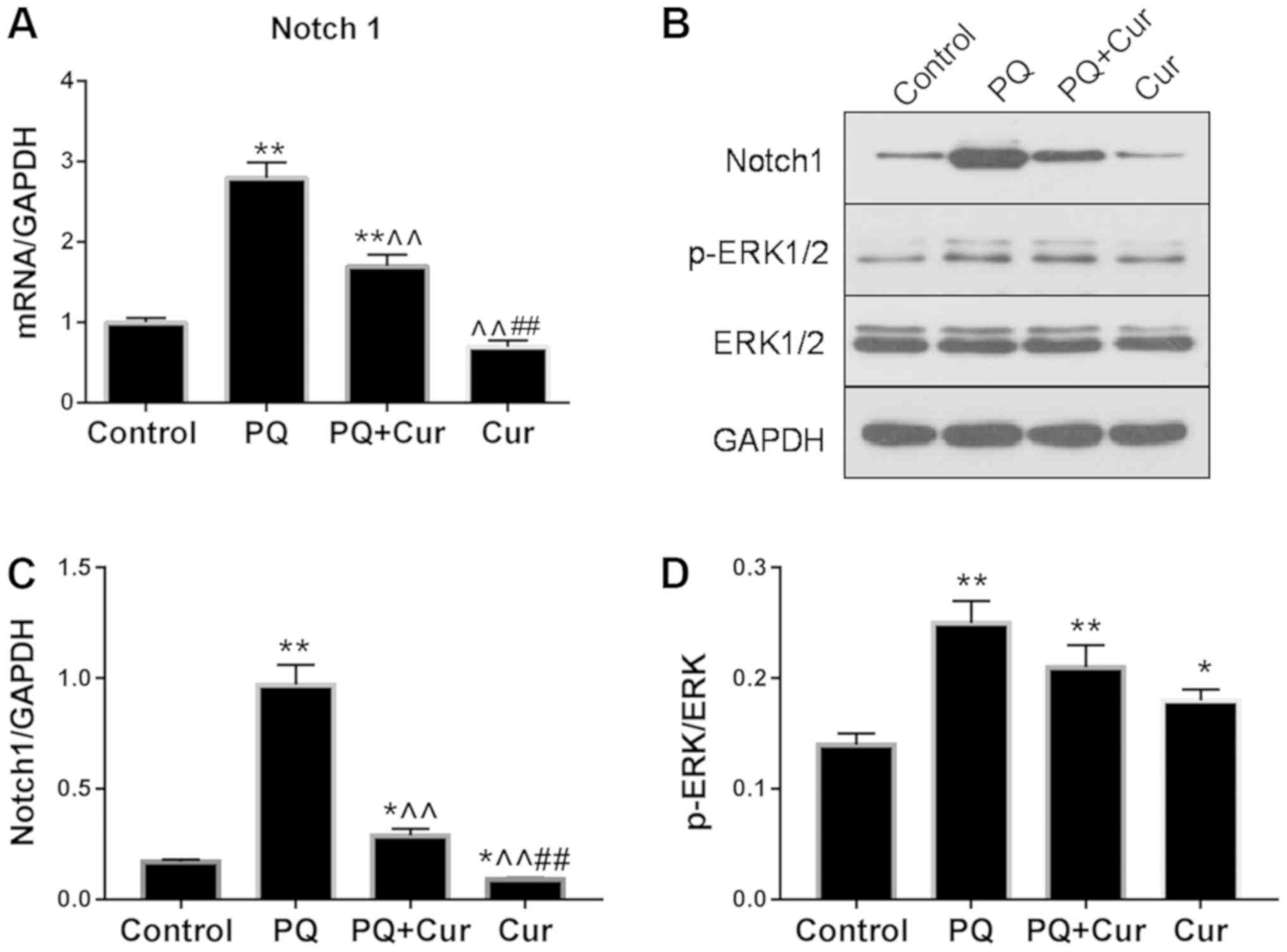
As presented in Fig.
5A-C, PQ treatment significantly upregulated the mRNA and
protein expression levels of Notch1, whereas they were
significantly reduced by curcumin. Additionally, the levels of
p-ERK1/2 and ERK1/2 were assessed by western blotting (Fig. 5B). The ratio of p-ERK1/2 to total
ERK1/2 for each group is presented in Fig. 5D. Significant increases in Notch1
and p-ERK1/2/ERK1/2 levels were observed in the PQ group compared
with the control (Fig. 5C and D).
Curcumin treatment significantly attenuated the PQ-induced
upregulation of Notch1; however, no significant difference in
p-ERK1/2/ERK1/2 was observed compared with PQ treatment alone. Of
note, significantly increased phosphorylation of ERK1/2 was also
observed following treatment with curcumin alone compared with the
control.
Discussion
The toxic effects of PQ are primarily mediated via
oxidative stress and inflammatory responses (27) In 2014, Tyagi et al (28) reported that intranasal curcumin
directly targets the lungs of mice, and significantly improves
PQ-induced lung damage by inhibiting the production of
proinflammatory cytokines and ROS. In the present study, it was
revealed that treatment with curcumin significantly attenuated
PQ-induced elevated ROS levels and apoptosis in WI-38VA13 cells.
Additionally, the expression of proteins in the
TXNIP/NLRP3-mediated proinflammatory pathway was determined, and it
was demonstrated that curcumin also significantly decreased
activation of the NLRP3 inflammasome. Furthermore, downregulation
of Notch1 was observed, indicating that the Notch signaling pathway
may serve a potential role in the mechanisms underlying the effects
of curcumin on PQ-induced ALI. Collectively, the present findings
suggested that curcumin may aid in relieving oxidative stress and
inflammation induced by PQ, and that these effects may be mediated
by decreasing ROS levels and inhibiting the activation of
NLRP3.
Previous studies have reported that PQ poisoning
leads to the depletion of cellular nicotinamide adenine
dinucleotide phosphate by generating large quantities of ROS, which
potently promotes oxidative stress and inflammatory responses, and
that the increasing lipid peroxidation may alter the function of
cell membrane receptors, inducing immune cells to produce ILs and
other inflammatory cytokines (28–30).
The Trx/TXNIP complex has been revealed to be an important
regulator of cellular redox status; overproduction of ROS induces
the separation of Trx and TXNIP, after which Trx promotes the
biosynthesis of manganese superoxide dismutase to inhibit ROS
production (31,32), whereas TXNIP directly interacts
with NLRP3 and promotes activation of the NLRP3 inflammasome
(13). In 2016, Kim et al
(3) demonstrated that ROS exhibits
the ability to induce upregulated TXNIP expression. In the present
study, it was demonstrated that the expression of TXNIP was
significantly increased following PQ treatment. In addition to
affecting the Trx/TXNIP complex, ROS have also been proposed to
directly activate NLRP3 (33,34).
The NLRP3 inflammasome serves an important role in inflammation and
innate immunity, and has been reported to be involved in the
pathogenesis of various types of inflammatory diseases (35–37),
including PQ-induced ALI (16).
Considering the association between TXNIP and NLRP3, it was
hypothesized that the mechanisms underlying PQ-induced inflammatory
responses in the lung may be mediated by the promoting effects of
ROS on TXNIP expression and subsequent NLRP3 activation. In the
present study, treatment with curcumin significantly reduced ROS
generation, which may suppress ROS-mediated activation of the
TXNIP/NLRP3 inflammatory pathway, and downstream caspase-1
cleavage, and IL-1β and IL-18 secretion. Furthermore, it was
observed that the apoptosis of WI-38VA13 cells was significantly
increased by treatment with PQ. Previous studies revealed that
ROS-associated apoptosis is mainly associated with the
apoptosis-signal kinase 1/Trx/TXNIP and TXNIP/NLRP3 inflammasome
signaling pathways (38,39). According to the results of flow
cytometry, and the expression of the anti-apoptotic protein Bcl-2
and proapoptotic protein Bax, the present results suggested that
curcumin inhibited PQ-induced apoptosis and inflammation in
WI-38VA13 cells, potentially by reducing ROS generation.
The protein levels of Notch1, p-ERK1/2 and ERK1/2
were also investigated by western blotting. Previous studies have
indicated that activation of Notch signaling promotes tissue
inflammation, neoplasia and metastasis (40,41).
In the present study, significant upregulation of Notch1 was
observed in PQ-treated WI-38VA13 cells. In 2014, Singla et
al (42) confirmed that
increased Notch1 levels promote proinflammatory cytokine release by
activating macrophage differentiation, whereas inhibiting Notch1
expression or blocking the Notch signaling pathway decreases
proinflammatory cytokine production and secretion. The present
findings suggested that curcumin downregulated Notch1 signaling,
potentially subsequently attenuating the synthesis and release of
proinflammatory cytokines; however, Jiang et al (43) reported that Notch signaling induces
inhibitory effects on NLRP3 activation by suppressing the high
mobility group protein B1/Toll-like receptor 4/NF-κB pathway in
acetaminophen-induced liver injury, in contrast with the present
study. It was also observed that PQ induced upregulation of
p-ERK1/2; however, curcumin treatment did not significantly affect
PQ-induced phosphorylation, indicating that ERK1/2 signaling may be
involved in the mechanisms underlying PQ-induced ALI but not those
contributing towards the protective effects of curcumin.
Collectively, Notch signaling may serve a role in the effects of
curcumin on PQ-induced ALI; however, the underlying molecular
mechanisms require further investigation.
The present study possessed certain limitations.
Additional inflammatory cytokines should be measured to more fully
characterize the activation of TXNIP/NLRP3-mediated inflammatory
pathways. In addition, the mechanisms underlying the potential
involvement of the Notch pathway in the effects of curcumin on
PQ-induced ALI are yet to be investigated.
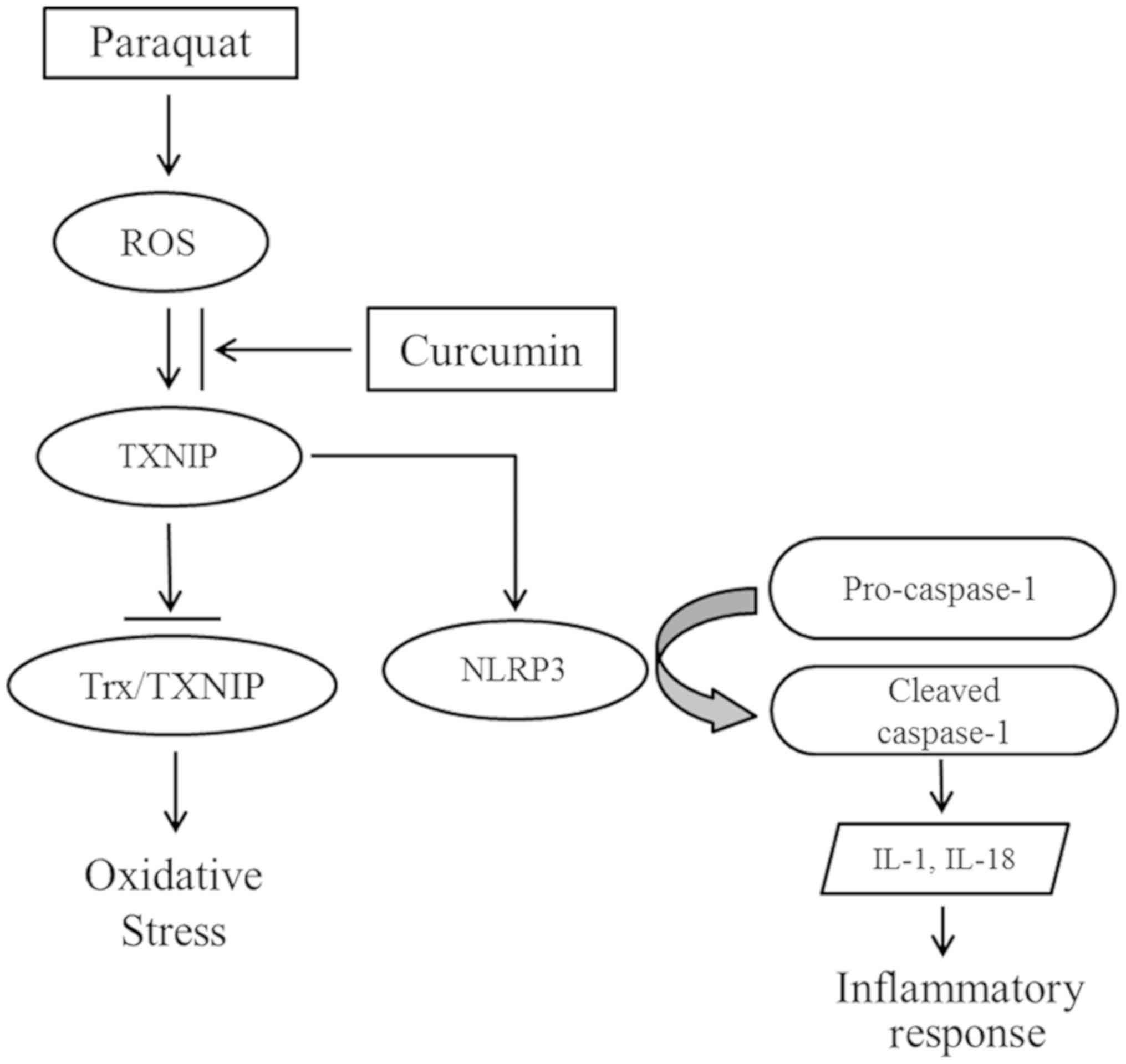
In conclusion, the present study suggested that PQ
poisoning induced the overproduction of ROS, which may promote
TXNIP expression. TXNIP not only inhibits the antioxidative ability
of Trx, but also contributes to NLRP3 activation, and the
maturation and secretion of proinflammatory cytokines (13). Following treatment with curcumin,
inflammation and apoptosis were significantly decreased, and
protein expression analysis suggested that the beneficial effects
of curcumin may be mediated by suppressing the TXNIP/NLRP3
inflammatory axis (Fig. 6). In
addition, it was observed that Notch1 signaling served a potential
role in the mechanisms via which curcumin attenuated PQ poisoning.
The present findings provide further understanding of the molecular
mechanisms underlying the effects of curcumin on PQ-induced lung
injury, and indicated that TXNIP/NLRP3 may be a novel therapeutic
target for the treatment of PQ-induced ALI.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant nos. 81401583 and
81701894), Social Development Projects of Jiangsu Province (grant
no. BE2017720), Jiangsu Provincial Medical Youth Talent (grant nos.
QNRC2016908 and QNRC2016909), the Major Projects Foundation of
General Logistics Department of PLA (grant no. CNJ14L002), the
Peking Union Farsighted Emergency Project (grant no. RE2016002) and
the Natural Science Foundation of Jinling Hospital (grant nos.
2015027 and 2016032).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XC and SN conceived and designed the study. YR
studied the effect of curcumin on ROS and apoptosis induced by
paraquat. ZY studied the effect of curcumin on apoptosis related
factors induced by paraquat. ZS studied the effect of curcumin on
Notch pathway activated by paraquat. WZ studied the effect of
curcumin on proinflammatory cytokines induced by paraquat. SN was a
major contributor in writing the manuscript. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Baltazar T, Dinis-Oliveira RJ, Duarte JA,
de Lourdes Bastos M and Carvalho F: Paraquat research: Do recent
advances in limiting its toxicity make its use safer? Br J
Pharmacol. 168:44–45. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zhang B, Lang Y and Yi O: Literature
analysis of paraquat poisoning in China (1991–2008). Chin J Crit
Care Med. 30:139–141. 2010.(In Chinese).
|
|
3
|
Kim E, Leverage WT, Liu Y, Panzella L,
Alfieri ML, Napolitano A, Bentley WE and Payne GF: Paraquat-melanin
redox-cycling: Evidence from electrochemical reverse engineering.
ACS Chem Neurosci. 7:1057–1067. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Dinis-Oliveira RJ, Duarte JA,
Sanchez-Navarro A, Remião F, Bastos ML and Carvalho F: Paraquat
poisonings: Mechanisms of lung toxicity, clinical features, and
treatment. Crit Rev Toxicol. 38:13–71. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Tomita M, Okuyama T, Katsuyama H, Miura Y,
Nishimura Y, Hidaka K, Otsuki T and Ishikawa T: Mouse model of
paraquat-poisoned lungs and its gene expression profile.
Toxicology. 231:200–209. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Liu J, Xiong Y and Jiang M: Ratio of
injured lung volume fraction in prognosis evaluation of acute PQ
poisoning. Biomed Res Int. 2018:e45015362018.
|
|
7
|
Yang W, Liu W, Yu W, Fei D, Meng X, Yang
S, Meng S and Zhao M: Angptl2 deficiency attenuates paraquat
(PQ)-induced lung injury in mice by altering inflammation,
oxidative stress and fibrosis through NF-κB. Biochem Biophys Res
Commun. 503:94–101. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hu S, Qiao C, Yuan Z, Li M, Ye J, Ma H,
Wang J, Xin S and Zhang J: Therapy with high-dose long-term
antioxidant free radicals for severe paraquat poisoning: A pilot
study. Exp Ther Med. 16:5149–5155. 2018.PubMed/NCBI
|
|
9
|
Buendia JA, Chavarriaga GJR and Zuluaga
AF: Burden of paraquat poisoning in the department of Antioquia,
Colombia. BMC Pharmacol Toxicol. 20:112019. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Nasr Isfahani S, Farajzadegan Z,
Sabzghabaee AM, Rahimi A, Samasamshariat S and Eizadi-Mood N: Does
hemoperfusion in combination with other treatments reduce the
mortality of patients with paraquat poisoning more than
hemoperfusion alone: A systematic review with meta-analysis. J Res
Med Sci. 24:22019. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Devi TS, Lee I, Hüttemann M, Kumar A,
Nantwi KD and Singh LP: TXNIP links innate host defense mechanisms
to oxidative stress and inflammation in retinal Muller glia under
chronic hyperglycemia: Implications for diabetic retinopathy. Exp
Diabetes Res. 2012:4382382012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yoshihara E, Masaki S, Matsuo Y, Chen Z,
Tian H and Yodoi J: Thioredoxin/Txnip: Redoxisome, as a redox
switch for the pathogenesis of diseases. Front Immunol. 4:5142014.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhou R, Tardivel A, Thorens B, Choi I and
Tschopp J: Thioredoxin-interacting protein links oxidative stress
to inflammasome activation. Nat Immunol. 11:136–140. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lamkanfi M and Dixit VM: Inflammasomes and
their roles in health and disease. Annu Rev Cell Dev Biol.
28:137–161. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Schroder K and Tschopp J: The
inflammasomes. Cell. 140:821–832. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu Z, Zhao H, Liu W, Li T, Wang Y and
Zhao M: NLRP3 inflammasome activation is essential for
paraquat-induced acute lung injury. Inflammation. 38:433–444. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Narayanan A, Kehn-Hall K, Senina S,
Lundberg L, Van Duyne R, Guendel I, Das R, Baer A, Bethel L, Turell
M, et al: Curcumin inhibits rift valley fever virus replication in
human cells. J Biol Chem. 287:33198–33214. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Rahmani AH, Al Zohairy MA, Aly SM and Khan
MA: Curcumin: A potential candidate in prevention of cancer via
modulation of molecular pathways. Biomed Res Int. 2014:7616082014.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Shehzad A, Lee J and Lee YS: Curcumin in
various cancers. Biofactors. 39:56–68. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gong Z, Zhao S, Zhou J, Yan J, Wang L, Du
X, Li H, Chen Y, Cai W and Wu J: Curcumin alleviates DSS-induced
colitis via inhibiting NLRP3 inflammsome activation and IL-1β
production. Mol Immunol. 104:11–19. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ding XQ, Wu WY, Jiao RQ, Gu TT, Xu Q, Pan
Y and Kong LD: Curcumin and allopurinol ameliorate fructose-induced
hepatic inflammation in rats via miR-200a-mediated TXNIP/NLRP3
inflammasome inhibition. Pharmacol Res. 137:64–75. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Han G, Wei Z, Cui H, Zhang W, Wei X, Lu Z
and Bai X: NUSAP1 gene silencing inhibits cell proliferation,
migration and invasion through inhibiting DNMT1 gene expression in
human colorectal cancer. Exp Cell Res. 367:216–221. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Huang Y, An L, Hui KM, Ren Q and Wang W:
An LDLa domain-containing C-type lectin is involved in the innate
immunity of Eriocheir sinensis. Dev Comp Immunol. 42:333–344. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Goodman RB, Pugin J, Lee JS and Matthay
MA: Cytokine-mediated inflammation in acute lung injury. Cytokine
Growth Factor Rev. 14:523–535. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wen SH, Wu HJ, Lin L, Chong L, Zhu LL,
Zhang WX, Zhang HL and Li CC: Adjunctive dexamethasone therapy
improves lung injury by inhibiting inflammation and reducing RIP3
expression during Staphylococcus aureus pneumonia in mice. Int
Immunopharmacol. 23:709–718. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chen Y, Nie YC, Luo YL, Lin F, Zheng YF,
Cheng GH, Wu H, Zhang KJ, Su WW, Shen JG and Li PB: Protective
effects of naringin against paraquat-induced acute lung injury and
pulmonary fibrosis in mice. Food Chem Toxicol. 58:133–140. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Tyagi N, Kumari A, Dash D and Singh R:
Protective effects of intranasal curcumin on paraquot induced acute
lung injury (ALI) in mice. Environ Toxicol Pharmacol. 38:913–921.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li GP, Yang H, Zong SB, Liu Q, Li L, Xu
ZL, Zhou J, Wang ZZ and Xiao W: Diterpene ginkgolides meglumine
injection protects against paraquat-induced lung injury and
pulmonary fibrosis in rats. Biomed Pharmacother. 99:746–754. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Cheresh P, Kim SJ, Tulasiram S and Kamp
DW: Oxidative stress and pulmonary fibrosis. Biochim Biophys Acta.
1832:1028–1040. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Das KC, Lewis-Molock Y and White CW:
Elevation of manganese superoxide dismutase gene expression by
thioredoxin. Am J Respir Cell Mol Biol. 17:713–726. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Hwang J, Suh HW, Jeon YH, Hwang E, Nguyen
LT, Yeom J, Lee SG, Lee C, Kim KJ, Kang BS, et al: The structural
basis for the negative regulation of thioredoxin by
thioredoxin-interacting protein. Nat Commun. 5:29582014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yu X, Lan P, Hou X, Han Q, Lu N, Li T,
Jiao C, Zhang J, Zhang C and Tian Z: HBV inhibits LPS-induced NLRP3
inflammasome activation and IL-1β production via suppressing the
NF-κB pathway and ROS production. J Hepatol. 66:693–702. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Dai J, Zhang X, Li L, Chen H and Chai Y:
Autophagy inhibition contributes to ROS-producing NLRP3-dependent
inflammasome activation and cytokine secretion in high
glucose-induced macrophages. Cell Physiol Biochem. 43:247–256.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Heneka MT, Carson MJ, El Khoury J,
Landreth GE, Brosseron F, Feinstein DL, Jacobs AH, Wyss-Coray T,
Vitorica J, Ransohoff RM, et al: Neuroinflammation in Alzheimer's
disease. Lancet Neurol. 14:388–405. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Masters SL, Dunne A, Subramanian SL, Hull
RL, Tannahill GM, Sharp FA, Becker C, Franchi L, Yoshihara E, Chen
Z, et al: Activation of the NLRP3 inflammasome by islet amyloid
polypeptide provides a mechanism for enhanced IL-1β in type 2
diabetes. Nat Immunol. 11:897–904. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wang Z, Hu W, Lu C, Ma Z, Jiang S, Gu C,
Acuña-Castroviejo D and Yang Y: Targeting NLRP3 (Nucleotide-binding
domain, leucine-rich-containing family, pyrin domain-containing-3)
inflammasome in cardiovascular disorders. Arterioscler Thromb Vasc
Biol. 38:2765–2779. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zhang M, Harashima N, Moritani T, Huang W
and Harada M: The roles of ROS and caspases in TRAIL-induced
apoptosis and necroptosis in human pancreatic cancer cells. PLoS
One. 10:e01273862015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Friedemann T, Otto B, Klätschke K,
Schumacher U, Tao Y, Leung AK, Efferth T and Schroder S: Coptis
chinensis franch. exhibits neuroprotective properties against
oxidative stress in human neuroblastoma cells. J Ethnopharmacol.
155:607–615. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Piggott K, Deng J, Warrington K, Younge B,
Kubo JT, Desai M, Goronzy JJ and Weyand CM: Blocking the NOTCH
pathway inhibits vascular inflammation in large-vessel vasculitis.
Circulation. 123:309–318. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Poulsen LC, Edelmann RJ, Krüger S,
Diéguezhurtado R, Shah A, Stavnoraas TE, Renzi A, Szymanska M, Wang
J, Ehling M, et al: Inhibition of endothelial NOTCH1 signaling
attenuates inflammation by reducing cytokine-mediated histone
acetylation at inflammatory enhancers. Arterioscler Thromb Vasc
Biol. 38:854–869. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Singla RD, Wang J and Singla DK:
Regulation of Notch 1 signaling in THP-1 cells enhances M2
macrophage differentiation. Am J Physiol Heart Circ Physiol.
307:H1634–H1642. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Jiang L, Ke M, Yue S, Xiao W, Yan Y, Deng
X, Ying QL, Li J and Ke B: Blockade of Notch signaling promotes
acetaminophen-induced liver injury. Immunol Res. 65:739–749. 2017.
View Article : Google Scholar : PubMed/NCBI
|