Introduction
Postoperative cognitive dysfunction (POCD) is a
central nervous system (CNS) complication that can occur at any age
but is higher among elderly patients who are exposed to major
surgery under general anesthesia (1). With the continuous aging of the
population, the prevalence and consequences of POCD are increasing,
and interventions that prevent POCD represent an unmet medical
need. Across a variety of studies, POCD has been consistently
associated with prolonged hospitalization, reduced quality of life,
and increased morbidity (2).
Factors that can increase the risk of POCD include increasing age,
severity of the surgery, duration of anesthesia, occurrence of
complications, pre-existing cognitive impairment, hemorrhagic
shock, and education level (3,4).
Hyperlipidemia is among the many risk factors associated with the
development of POCD (3,4). The development and maintenance of
cerebral function relies on circulating lipids. Abnormalities in
gene expression, function, and regulation induce changes in lipid
metabolism that predispose patients to hyperlipidemia, which has
been shown to be associated with the development and progression of
Alzheimer's disease (5,6). The introduction of hyperlipidemia
into POCD-related research better reflects the clinical scenario
where increased lipids correlate with the incidence of POCD.
The blood-brain barrier (BBB) provides a protective
barrier between plasma and neuronal cells. While the selective
permeability of the BBB has important biological significance for
maintaining the normal physiological state of the CNS, it also
provides a barrier preventing passage of drugs into the CNS
(7). The BBB has been shown to be
disturbed in a variety of diseases, and animal models of these
conditions have been shown to display cognitive impairments leading
to Alzheimer's disease (8). The
permeability of the BBB can also increase in response to
inflammation, leading to damage to neuronal function.
Major facilitator superfamily domain-containing
protein 2 (Mfsd2a) is exclusively expressed in endothelial cells of
the BBB and is regulated by pericytes to facilitate BBB integrity
(9). Mfsd2a-deficient mice
exhibit neurological abnormalities, including ataxia (10). Tumor necrosis factor (TNF)-α is an
important mediator in the initiation and amplification of the
inflammatory cascade. Activation of immune cells in the peripheral
circulation in response to inflammation can result in learning and
memory dysfunction in animals (11). Interleukin (IL)-1β is also involved
in the inflammatory response, and upregulated expression of IL-1β
in the brain can lead to cognitive impairment (12).
In recent years, the protective effects of various
anesthetics have been observed and studied. Dexmedetomidine (Dex),
a selective agonist of the α2-adrenergic receptor, is widely used
in intensive care units due to its sedative and analgesic effects.
A large body of recent work supports its favorable effects for
improving outcomes and long-term brain function in the critically
ill. The source of these benefits may lie in the neuroprotective
properties that are seen in experimental models and in the clinical
setting, in which it can attenuate delirium, preserve sleep
architecture, preserve ventilatory drive and decrease sympathetic
tone and inflammatory response (13–15).
In a rat model of focal cerebral ischemia, Dex decreased the total
ischemic volume by 40% in the cortex compared to control treatment
with NaCl, and a study examining whether DEX can ameliorate
long-term cognitive dysfunction indicated that pre-treatment with
DEX may improve long-term cognitive function and protect against
neuronal degeneration (16).
Indeed, Dex treatment was found to reduce cerebral ischemia-induced
oxidative stress, apoptosis, and intracellular Ca2+
signaling by inhibiting transient receptor potential cation channel
subfamily M member 2 (TRPM2) and transient receptor potential
cation channel subfamily V member 1 (TRPV1) in the rat hippocampus
and dorsal root ganglion (17).
From the research observations described above, we
hypothesized that the incidence of POCD will be higher in rats with
hyperlipidemia than in controls and that Dex treatment may protect
against cognitive impairment through actions on Mfsd2a.
Materials and methods
Animals
Healthy male Sprague-Dawley rats (6–7 months old,
weighing 200–250 g) were purchased from the Dalian Medical
University Laboratory Animal Center and individually housed in
24×24×36 cm cages in a room with a temperature of 20±2°C and
humidity of 50±10%. Rats had access to sufficient food and water
and were kept on a 12:12 light/dark cycle (lights on at 9:00 a.m.).
All rats were allowed to adapt to their new environment for 7 days
prior to experiments. Any rats with poor eyesight or swimming
ability, as judged by the inability to swim to the platform shown
to them immediately prior to their placement in the water, would
have been excluded, but none of the rats obtained for this
experiment met these criteria. All animal procedures were approved
by the Animal Experiment Ethics Committee of Dalian Medical
University, China. All studies involving animals are carried out in
accordance with the ARRIVE guidelines for reporting experiments
involving animals.
Hyperlipidemia model
High-fat emulsion production
The composition of the high-fat emulsion was chosen
based on previous research (18).
A 200-ml beaker was put into a 60°C water bath, and 30 ml distilled
water, 20 ml propylene glycol, and 2 g sodium deoxycholate were
added and fully dissolved. In a separate beaker, 25 g fat, 10 g
cholesterol, 1 g propylthiouracil, and 25 ml Tween-80 were added,
and the beaker was heated to 100°C. The contents of the two beakers
were combined by stirring the contents of the first beaker into
that of the second beaker. The mixture was stored at 4°C and melted
in a water bath at 37°C prior to use.
Generation of the animal model
Two initial groups of rats were developed: a normal
diet-fed group and high-fat-treated group (Fig. 1). Every day between 8:00 and 9:00
am, rats in the high-fat-treated group (n=30) were fed the high-fat
emulsion preparation by gavage at a dosage of 10 ml/kg body weight.
The rats were fed this diet for 14 days to induce hyperlipidemia.
Rats in the normal diet-fed group (n=30) were administered the same
doses of saline.
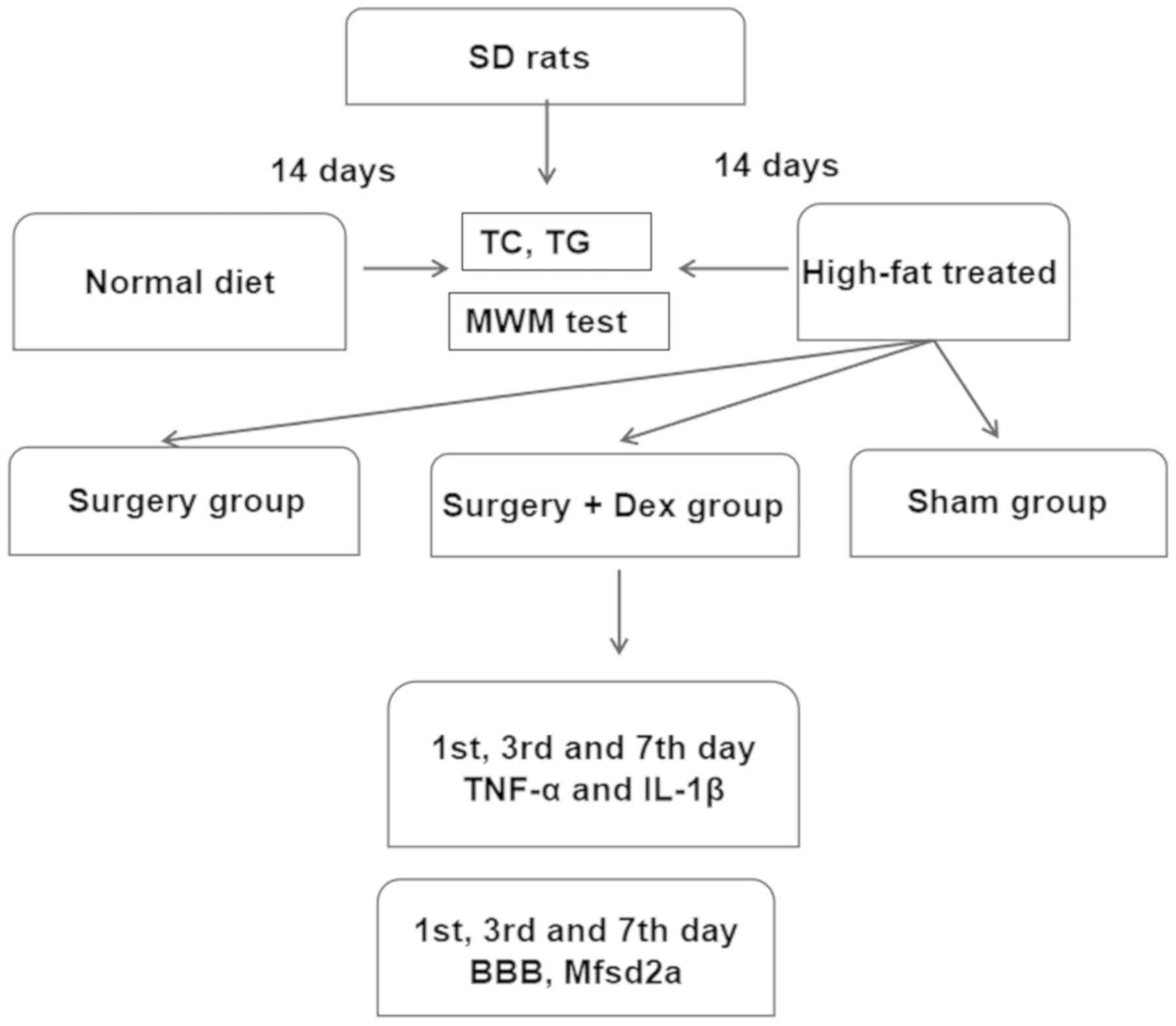 | Figure 1.Experimental flow chart. There were
two stages to our study. i) Establishment of the hyperlipidemia
model. The two groups were fed a normal diet and a high-fat
emulsion diet. These groups were established by 14 days of feeding.
ii) Establishment of the surgical groups. The sham group was fed
the high-fat diet and underwent the sham surgery procedure; the
surgery group was fed the high-fat diet, received anesthesia, and
underwent exploratory laparotomy surgery; and the surgery+Dex group
was fed the high-fat diet, received anesthesia, received preloading
treatment with 3 µg/kg Dex prior to exploratory laparotomy surgery,
underwent exploratory laparotomy surgery, and then received
subsequent infusion of Dex at 3 µg/kg/h via an osmotic pump for 2
h. BBB, blood-brain barrier; Mfsd2a, major facilitator superfamily
domain-containing protein 2; TNF-α, tumor necrosis factor-α; IL-1β,
interleukin-1β; TG, triglyceride; TC, total cholesterol; MWM,
Morris water maze. |
Following the last gavage treatment, rats were
fasted for 8 h. Then 2–3 ml blood was collected in heparin
anticoagulation tubes from the orbital venous plexus, and plasma
was collected via centrifugation (1,000 × g for 10 min).
Cholesterol was measured using a total cholesterol (TC)
enzyme-linked immunosorbent assay (ELISA) kit (American TSZ Co.).
Triglyceride (TG) levels were measured using a glycerol three-lipid
kit (F001-2 GPO-PAP; Nanjing Institute of Biological Engineering).
All assays were performed according to the manufacturer's
instructions. After successful establishment of the hyperlipidemia
model, navigation training was performed for 6 days. During the
training period, the high-fat-treated group rats continued to
receive gavage at the dose of 10 mg/kg to maintain elevated
cholesterol levels in the plasma. Hyperlipidemia rats were selected
to carry out further experiments.
Anesthesia and surgery
The high-fat-treated rats were randomly divided into
three groups (n=10 in each group; Fig.
1): A surgery group, in which rats were preloaded with 3 ml/kg
saline, subsequently infused with saline at 3 ml/kg/h with an
osmotic pump for 2 h, and then received anesthesia and underwent
exploratory laparotomy surgery; a surgery + Dex group, in which
rats received anesthesia, were preloaded with 3 µg/kg Dex prior to
exploratory laparotomy, and subsequently received Dex infusion at 3
µg/kg/h via an osmotic pump for 2 h; and a sham surgery group, in
which rats received anesthesia, were preloaded with 3 ml/kg saline
and subsequently infused with saline at 3 ml/kg/h with an osmotic
pump for 2 h. The selected DEX concentration (3 µg/kg) was
determined to provide the most significant protective effect in a
preliminary experiment. For the POCD model, brief anesthesia (1.5%
sevoflurane for 5 min) was administered in an anesthetic chamber.
Rats were endotracheal intubated and mechanically ventilated with
1–2% sevoflurane in 100% O2. For the exploratory
laparotomy, the abdominal region was shaved and sterilized, and a
3-cm vertical incision was made approximately 0.5 cm below the
lower right rib. The gastrointestinal tract was exteriorized, and
the upper mesenteric artery clamped for 30 min, leading to
restriction of blood flow in the mesenteric vascular bed. The rats
in the sham surgery group were treated in the same way without
laparotomy. All rats were kept under sevoflurane anesthesia for 2 h
and given analgesia (0.25% bupivacaine, 0.03 mg/kg subcutaneously)
at the end of surgery. Prior to abdominal surgery, the femoral vein
of the rats was released from clamping to allow anesthetic drug or
saline infusion. In order to alleviate incision pain for the
animals, local anesthetics were used during sealing of the
incision. In the event that any animal experienced death, severe
infection, incision dehiscence, and weakness, they were excluded
from the experiment. No experimental animals lost more than 15–20%
of their body weight or were unable to eat independently for more
than 24 h.
Morris water maze (MWM)
MWM testing was used to assess spatial learning,
spatial memory, and cognitive abilities. The water maze consisted
of a circular pool (diameter of 150 cm, height of 50 cm, black
interior wall) that was filled with water kept at a temperature of
26±1°C. The maze was divided into four quadrants with equidistantly
placed cameras in each quadrant that were connected to a
computerized image analysis system. These cameras were used to
record the swimming course of the rats. A transparent cylinder
platform (diameter of 10 cm, height of 27 cm) was placed in the
center of a quadrant, 10 cm underwater. All rats were trained three
times a day for 6 consecutive days. The training involved the
hidden platform discovery experiment and the space exploration
experiment (19), with hidden
platform discovery training occurring first, followed by the space
exploration training on the same day. After 6 days of training,
both assessments were made on days 1, 3, and 7 after surgery.
For the hidden platform discovery experiment, rats
were trained to find the hidden platform. Daily training started at
10:00 am and consisted of three sessions that lasted for 1 h and
were repeated three times a day. Each rat was sequentially placed
in each quadrant and allowed to find the platform. The order of the
quadrants was randomly chosen. The experiment was concluded when
the rat found the platform and remained on it for 10 sec. At the
completion of the experiment, the rats were wiped dry with towels
and returned to their cages. The average time to find the platforms
was recorded as the space learning ability.
Space exploration experiments were conducted by
removing the platform in order to record the swimming course of the
rats over a 35-sec duration. At 10:00 am, a probe trial was
performed to assess spatial memory. The platform was removed from
the maze. Each rat was randomly placed in one of the quadrants and
allowed to explore the maze for 60 sec. Behavior was analyzed with
the computerized image analyzing system. Moving distance and time
spent were measured as indices of spatial memory. Our water maze
detection device has a lifting platform, thus before each test, the
platform was raised to the water level, allowing the animals to see
the platform. When the animals were placed in the swimming pool, if
they swam directly to the platform without any difficulty, this
showed that their swimming ability and vision were normal and they
could be used for the experiment.
TNF-α and IL-1β measurement
Serum levels of TNF-α and IL-1β were measured using
ELISA kits (Wuhan USCN Business Co., Ltd.). For the measurements,
2–3 ml blood was collected from the orbital venous plexus on days
1, 3 and 7 after surgery at the end of the light phase and
coagulated for 30 min. Serum was collected via centrifugation
(1,000 × g for 10 min). ELISAs were performed according to the
manufacturer's instructions. Briefly, microtiter plates (96-well
flat-bottom) were coated for 24 h with the samples diluted at a
ratio of 1:2 in diluent to a final volume of 100 µl. The samples
were analyzed in duplicate. The plates were washed three times with
diluent, and monoclonal anti-TNF-α (cat. no. SEA133Ra) or
anti-IL-1β (cat. no. SEA563Ra) antibodies from Wuhan USCN Business
Co., Ltd., diluted 1:1,000 in diluent were added to each well and
incubated for 3 h at room temperature. After washing, a
peroxidase-conjugated anti-rabbit secondary antibody (diluted
1:1,000) was added to each well and incubated at room temperature
for 1 h. After addition of streptavidin-enzyme, substrate and stop
solution, the concentrations of TNF-α and IL-1β were determined
based on the absorbance at 450 nm in a spectrophotometer. The
standard curve demonstrated a direct relationship between optical
density (OD) and test concentrations. Total protein was measured by
Lowry's method using bovine serum albumin (BSA) as a standard.
Measurement of BBB permeability
BBB permeability was evaluated by Evans blue
(Solarbio) extravasation measurements. Rats were anesthetized with
2% sevoflurane and injected with 2% Evans blue dye intravenously (4
ml/kg) through the femoral vein. After 1 h, rats were perfused with
200 ml normal saline through the left ventricle and sacrificed for
hippocampus tissue collection. Hippocampal samples were homogenized
in 3 ml cold 7.5% trichloroacetic acid (Solarbio) and centrifuged
at 10,000 × g for 10 min. The absorbance of each hippocampus tissue
extraction was measured at 632 nm using a spectrophotometer
(Synergy H1 Hybrid Reader; BioTek Instruments, Inc.). Evans blue
dye content was expressed as µg/mg of hippocampal tissue and
evaluated against positive control standard curves.
Quantitative real-time PCR (qPCR)
analysis for Mfsd2a
Total RNA was extracted from brain tissue using the
RNAiso Plus Purification Kit (9108; Takara Biotechnology Co.,
Ltd.). RNA was evaluated by A260 (OD) measurement and integrity was
checked by 2% agarose gel electrophoresis. RNA was reverse
transcribed into cDNA using PrimeScript™ RT Reagent Kit for gDNA
Eraser Reverse Transcript Reagent Kit (RR047A; Takara Biotechnology
Co., Ltd.). qPCR was performed in a LightCycler machine (Roche)
with commercial SYBR-Green reaction reagent (RR820A; Takara
Biotechnology Co., Ltd.). GAPDH was used as an internal control.
The conditions for PCR consisted of 40 cycles of 95°C for 5 sec and
60°C for 20 sec followed by extension at 72°C for 10 min. The
primer sequences used for gene amplification (Invitrogen; Thermo
Fisher Scientific, Inc.) were as follows: MFSD2A-forward,
5′-CCACATTCACCATCCCTATCT-3′ and reverse,
5′-TTCTTATTCTGTCGCCGCTTC-3′, GAPDH-forward, 5′-ATGCCGCCTGGAAACC-3′
and reverse, 5′-GCATCAAAGTGGAAGAATGG-3′. The amplified products
were 199 bp in length; gene expression was quantified via the
2−∆∆Cq method (20).
Statistical analysis
All data were analyzed with SPSS version 20 (SPSS,
Inc., Chicago, IL, USA). Statistical data are expressed as mean ±
standard error of the mean (SEM). MWM results were assessed with
repeated measures analysis of variance (ANOVA) with Bonferroni
post-test. All other data were analyzed with one-way ANOVA with
Bonferroni post-test. Effects were regarded as significantly
different at P<0.05.
Results
High-fat diet feeding induces
hyperlipidemia
After 14 days of feeding on the normal diet, the
serum TC concentration was 1.65±0.20 mmol/l and the serum TG
concentration was 0.38±0.10 mmol/l. Comparatively, the rats fed the
high-fat diet had a serum TC concentration of 3.33±0.59 mmol/l and
serum TG concentration of 0.63±0.18 mmol/l. Thus, the serum TC and
TG concentrations were elevated in the high-fat-treated rats
compared with those in the rats fed a normal diet (Table I; both P<0.05), indicating the
development of hyperlipidemia. Hyperlipidemia was maintained by
continual high-fat emulsion lavage.
 | Table I.Serum total cholesterol (TC) and
triglyceride (TG) levels after high-fat treatment. |
Table I.
Serum total cholesterol (TC) and
triglyceride (TG) levels after high-fat treatment.
|
|
| Day 14 | Day 20 |
|---|
|
|
|
|
|
|---|
| Group | Dose | TC (mmol/l) | TG (mmol/l) | TC (mmol/l) | TG(mmol/l) |
|---|
| Saline | 10 ml/kg | 1.65±0.20 | 0.38±0.10 | 1.53±0.20 | 0.30±0.10 |
| High-fat
treated | 10 ml/kg |
3.33±0.59b |
0.63±0.18a |
4.28±0.59b |
0.72±0.18a |
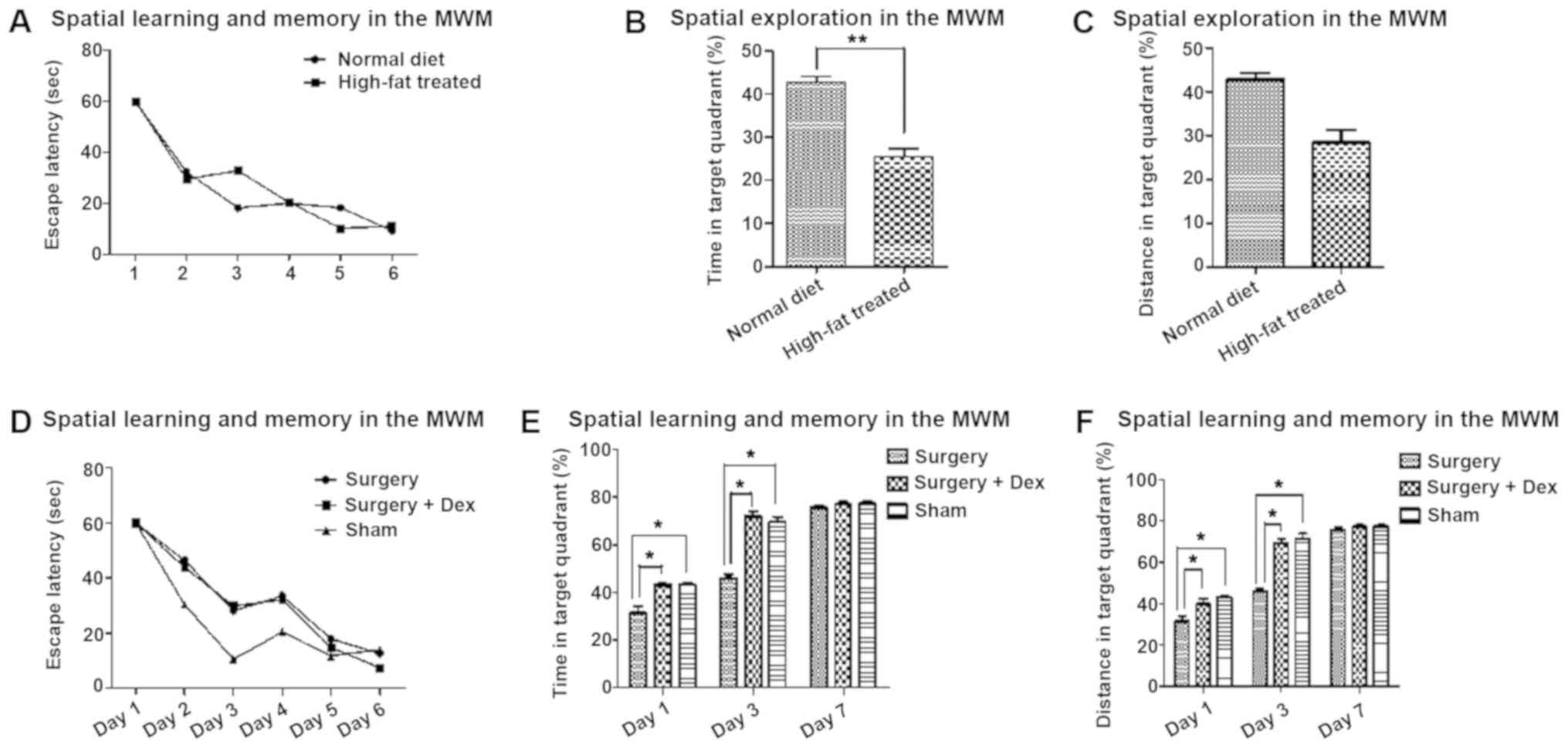
Cognitive function is reduced by both
high-fat diet and surgery, and Dex treatment prevents POCD
After training on the escape latency test, the rats
were able to learn and remember the location of the platform
(Fig. 2A and D). In the space
exploration test, the time and distance ratio of the high-fat
treated rats were lower for targeting the quadrant than those of
the rats fed a normal diet (Fig. 2B
and C, P<0.01), indicating hyperlipidemia-induced cognitive
impairment.
After surgery, the time and distance ratio of the
sham and surgery+Dex rats were both higher than those of the
surgery group on days 1 and 3 (Fig. 2E
and F, P<0.05), with no significant difference between them.
The differences between the sham and surgery groups indicate that
surgical stimulation of POCD occurred in the early post-operative
period, and the differences between the surgery+Dex and surgery
groups suggest that Dex prevented the occurrence of POCD. On day 7,
there were no differences among the groups.
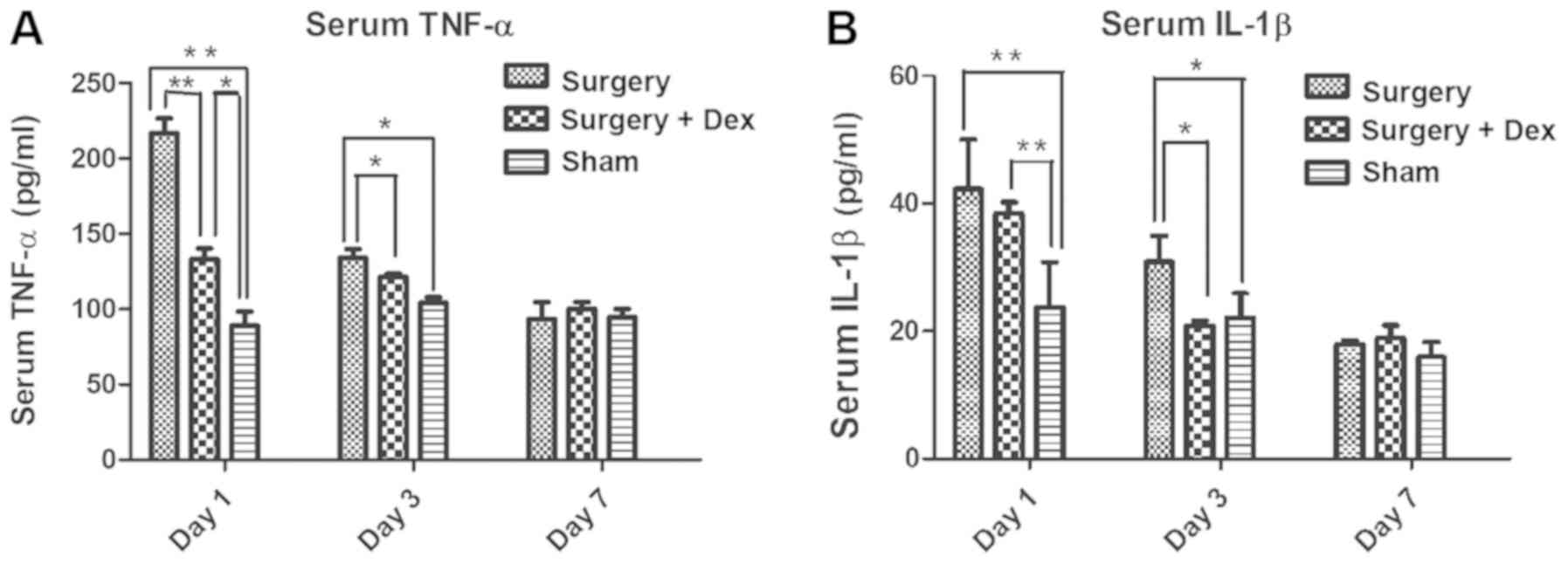
The surgery-induced inflammatory
response is alleviated by Dex treatment
The serum TNF-α concentrations in both the sham and
surgery+Dex groups were lower than that in the surgery group on day
1 (Fig. 3A, both P<0.01), and
the TNF-α concentration was higher in the surgery+Dex group than
that noted in the sham group (P<0.01). On day 3, the TNF-α
concentrations in the sham and surgery+Dex groups were still lower
than that in the surgery group (Fig.
3A, both P<0.05), but no difference was observed in the
TNF-α concentrations in the sham and surgery+Dex groups. These
results revealed that anesthesia and exploratory laparotomy induced
an inflammatory response and that Dex treatment alleviated this
inflammatory response in the early post-operative period. On day 7
post-operatively, no significant differences in serum TNF-α
concentration were observed among the groups, indicating that the
inflammation induced by surgery had returned to baseline
levels.
The serum IL-1β concentrations in the surgery and
surgery+Dex groups were both higher than that in the sham group on
day 1 (Fig. 3B, P<0.01). On day
3 however, while the IL-1β concentration in the surgery group was
still higher than that in the sham group (P<0.05), the IL-1β
concentration in the surgery+Dex group had decreased to less than
that in the surgery group (P<0.05), implying a reduction in
inflammation with Dex treatment.
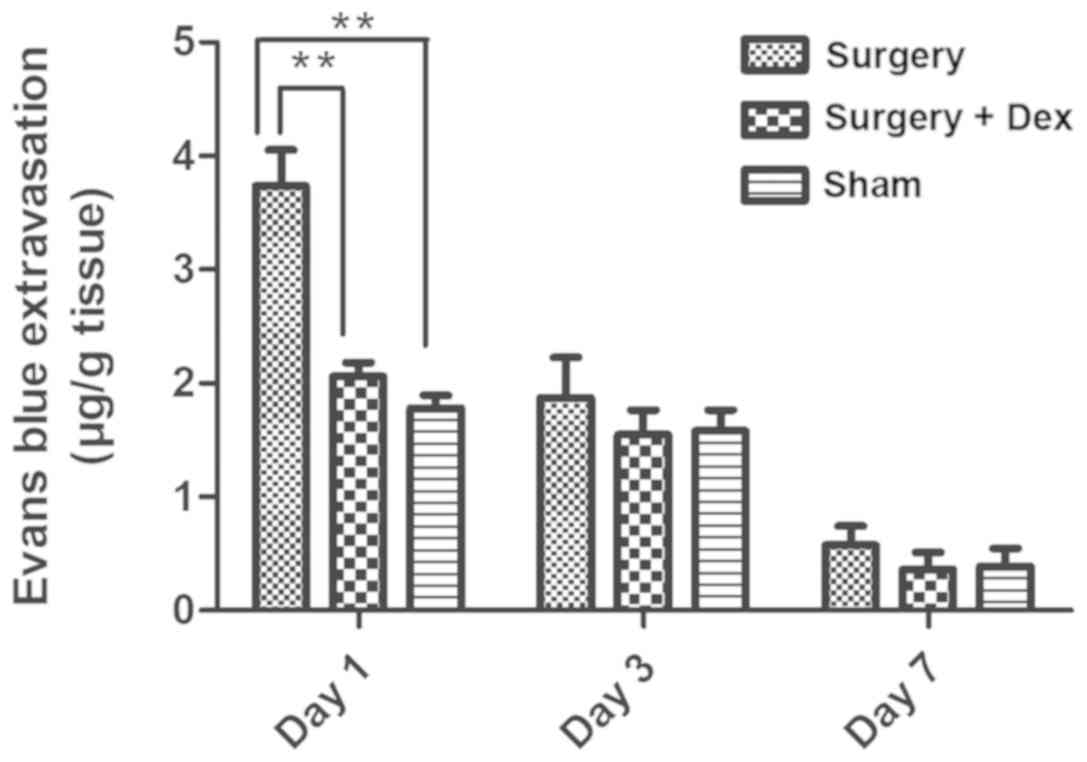
BBB permeability
Quantification of the observed Evans Blue staining
in each group revealed significantly more staining in the surgery
group than in the sham group on day 1 (Fig. 4, P<0.01), indicating that
surgery induced BBB dysfunction. In addition, Evans Blue staining
was significantly reduced in the surgery+Dex compared with the
surgery group (P<0.01) and not statistically different from that
in the sham group on day 1 (P>0.05), indicating that Dex
treatment attenuated the increase in BBB permeability caused by
surgical trauma. No differences were observed among the groups on
days 3 and 7.
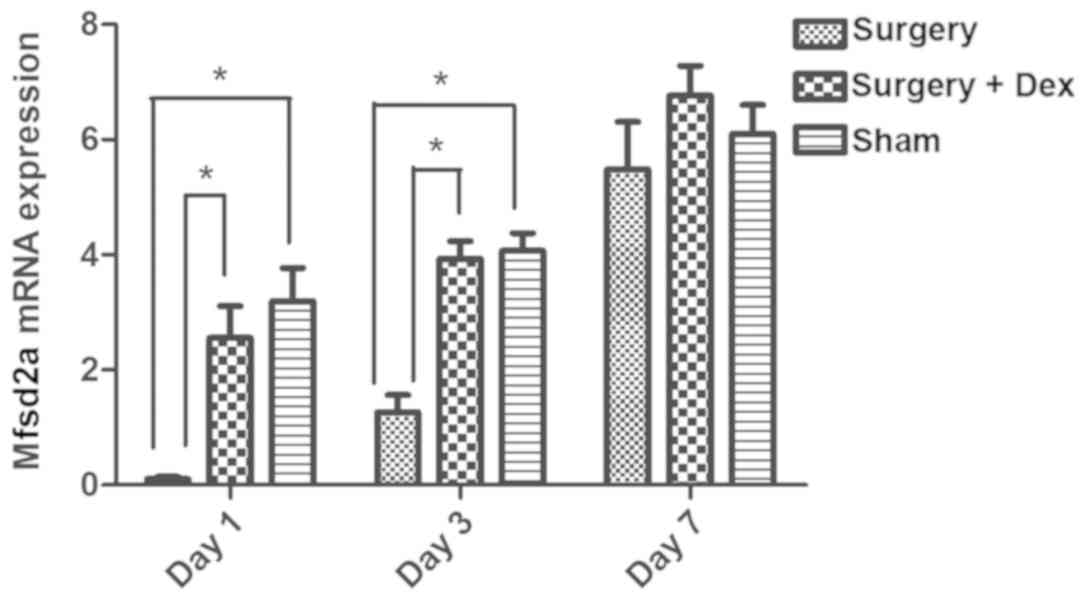
Downregulation of Mfsd2a mRNA
expression in the early postoperative period is prevented by Dex
treatment
The relative expression of Mfsd2a mRNA in the
surgery group was significantly lower than that in the sham group
on days 1 and 3 (Fig. 5, both
P<0.05), indicating that Mfsd2a mRNA expression was
downregulated in the early post-operative period. However,
Mfsd2a expression did not differ between the sham and
surgery+Dex groups on days 1 and 3, with significantly higher
Mfsd2a expression observed in the surgery+Dex group compared
with the surgery group at these time points (P<0.05). These
results indicate that the downregulation of Mfsd2a following
surgery was prevented by Dex treatment. By day 7, no differences in
relative expression of Mfsd2a mRNA were observed among the
three groups.
Discussion
The aim of the present study was to evaluate the
effect of dexmedetomidine (Dex) on the incidence of postoperative
cognitive dysfunction (POCD) in an animal model of
hyperlipidemia-induced blood-brain barrier (BBB) dysfunction,
through its effects on Mfsd2a gene expression. The major
findings of the present study were that: i) Anesthesia and surgery
increased BBB permeability and, correspondingly, induced cognitive
dysfunction, and ii) treatment with Dex attenuated these effects.
Combined, our results revealed an inflammation-based mechanism for
the development of POCD.
Surgery and anesthesia have been shown to induce
tissue damage and activate the peripheral innate immune system,
resulting in the release of inflammatory mediators (21). Following surgery, elderly patients
often develop POCD, which can both lengthen the recovery process
and accelerate future progression to Alzheimer's disease.
Anesthetics, particularly volatile anesthetics, have been strongly
associated with the development of POCD (22–24).
With respect to pre-clinical studies, both cell culture and animal
studies have suggested that anesthetics may stimulate
neuroapoptosis, caspase activation, β-amyloid protein (Aβ)
accumulation and oligomerization, and, ultimately,
neurodegeneration and deficits in neurocognition. While the primary
risk factors for POCD are increased age, surgery, and use of
anesthesia, other risk factors include hyperlipidemia, diabetes
mellitus, obesity, vascular factors, and depression, all of which
play a role in the pathogenesis of POCD (25). Among these risk factors, the
effects of hyperlipidemia have been largely understudied.
Hyperlipidemia is a systemic disorder of lipid
metabolism resulting in elevated levels of fats in the blood,
including total cholesterol (TC), triglycerides (TG), free fatty
acids, high-density lipoprotein (HDL), very low-density lipoprotein
(VLDL), and low-density lipoprotein (LDL), which may eventually
result in atherosclerosis. Atherosclerosis is a known trigger for
immune responses that drives neuroinflammation and
neurodegeneration. Evaluation of the effects of anesthetics on
cognitive dysfunction in hyperlipidemia animal models have not, to
date, been reported. In our experiments, rats fed a high-fat
emulsion for 14 days developed elevated serum TC and TG levels,
which led to learning and memory disturbances according to results
obtained using the Morris water maze. The Morris water maze is a
particularly useful tool for judging the spatial learning and
memory ability of mice (26). It
can provide more experimental parameters for systematically and
comprehensively investigating the spatial cognitive processing of
experimental animals and objectively reflects their cognitive
level. The learning and memory impairment and sensory and motor
impairment of experimental animals were separated to reduce their
interference in the detection of learning and memory processing.
Additionally, our laboratory experience with using the Morris water
maze to detect behavioral changes in animals is superior to that
for other experimental methods.
The BBB limits the entry of plasma components, red
blood cells, and leukocytes into the brain (27). The BBB is often disrupted after
ischemic injury, inflammation, or in a variety of vascular
disorders that can generate neurotoxic products, thereby
compromising synaptic and neuronal functions. Furthermore, an
intact BBB is a major obstacle for drug delivery to the central
nervous system (CNS). A limited understanding of the molecular
mechanisms that control BBB formation has hindered our ability to
manipulate the BBB in disease and drug therapy. Therefore,
understanding how the BBB is impaired following surgery could
provide insight and aid in improving therapies targeting the CNS
(28). Major facilitator
superfamily domain-containing protein 2 (Mfsd2a) is a key regulator
of BBB function that may act by suppressing transcytosis in
CNS-specific endothelial cells. Mfsd2a, therefore, may play a
protective role in maintaining BBB function (29). In the present study, Mfsd2a
expression was reduced following surgery, an effect that was
inhibited by Dex treatment. While this effect on Mfsd2a
expression provides insight into the mechanism of the therapeutic
effects of Dex, future studies of additional genes will be
valuable. In a rat model of lipopolysaccharide (LPS)-induced
neuroinflammation, additional genes showed altered expression after
Dex treatment (30). These include
the IL1-β and TNF-α genes as well as miR132 expression and
deacetylation of histone H3. Dex decreased the LPS-induced
reductions in IL1-β and TNF-α gene expression levels and reversed
the Aβ-induced deacetylation of histone H3, thereby, increasing
BDNF production. Studies of the effects of their altered expression
following Dex treatment are warranted. A previous study showed that
treatment with Dex reduced inflammation after trauma, increased the
expression of tight junction proteins, and diminished secondary BBB
damage and apoptosis, and these protective effects were likely
mediated via reduced activation of the nuclear factor (NF)-κB and
NLRP3 inflammasome pathways (31).
Future research into the various effects of Dex treatment will
provide a more comprehensive understanding of how the drug can be
used clinically.
The neuroprotective properties of Dex are largely
attributed to its agonistic actions on α2-adrenoreceptors, which
belong to the G-protein coupled family of transmembrane receptors
and are present on both pre- and post-synaptic autonomic ganglia in
the central and peripheral nervous systems (15). Binding of agonists, whether
endogenous (norepinephrine) or exogenous (dexmedetomidine), results
in G-protein coupling with the inhibition of both adenylyl cyclase
and phospholipase C activity and additional downstream effects
caused by such inhibition. Neuroinflammation may be the principal
cause for cognitive dysfunction after general anesthesia or surgery
in rodents. The results of the present study demonstrated that Dex
improved learning and memory function in the early postoperative
period in a rat model of hyperlipidemia.
Surgical trauma and anesthesia activate the immune
response of both the peripheral and CNS. Surgical trauma will
inevitably affect the blood pressure and respiration of animals,
leading to changes in the synthesis and release of inflammatory
factors. Therefore, in the pre-experimental stage, we performed one
femoral artery puncture manometry and another femoral vein
administration and blood collection in rats. We found no
significant effect of anesthesia and operation on blood pressure
and heart rate among the groups in the same experiment. Therefore,
femoral artery puncture was not performed in order to reduce the
incidence of infection after operation in the water maze
experiment.
An increase in inflammatory cytokines in the serum
may be an important risk factor for cognitive decline after surgery
(32). We did not include a group
treated with Dex after sham surgery, as surgical trauma is the
inducing factor for POCD in rats. A surgery group, in which rats
received anesthesia and surgery served as a positive control group.
The impairment of surgical trauma on cognitive function of rats in
the sham-operation group was slightly less than that in operation
group, which was regarded as a negative control group. The Dex
group was used as the experimental group to observe the protective
effect of Dex on cognitive function. Thus, Dex treatment without
surgery would not provide useful information for the purpose of our
study.
Both TNF-α and IL-1β have been implicated in the
development of cognitive dysfunction (33). Pro-inflammatory cytokines can
inhibit long-term potentiation and the expression of brain-derived
neurotrophic factor in the hippocampus. Moreover, TNF-α and IL-1β
are known to contribute to neuroinflammation in many CNS disorders,
as well as the development of inflammatory responses within the
brain. Dex administration has been shown to decrease the production
of inflammatory cytokines in severely septic patients (15). In the present study, Dex treatment
attenuated the postoperative increase in serum TNF-α but not IL-1β
on days 1 and 3 after surgery. A limitation of the present study is
that we measured TNF-α and IL-1β levels in serum only, and the
effects of DEX on TNF-α and IL-1β concentrations in the brain
should be determined in future studies.
In conclusion, our results revealed that
hyperlipidemia in rats increased postoperative cognitive impairment
following surgery, and these effects could be attenuated by Dex
treatment. Neuronal dysfunction was ameliorated by inhibiting the
inflammatory response and increasing Mfsd2a gene expression
to preserve the integrity of the BBB. Our findings provide
rationale for examining Dex as a preemptive therapy for preventing
POCD in elderly patients.
Acknowledgements
The authors would like to thank Professor Shen Lv of
the Research Center of the Second Affiliated Hospital of Dalian
Medical University, and Miss Yu-Hua Gao of the Department of
Anesthesiology, Henan Province Hospital of Traditional Chinese
Medicine, for their technical support.
Funding
This research was supported by funding from the
Science and Technology Department of Liaoning Province of China
(grant no. 2013023009). The authors declare that they have no
financial relationship with the organization that sponsored the
research, and the funding body was not involved in study design,
data collection, analysis and writing of the study.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XPZ and DBL conceived and designed the experiments.
XPZ and YRL performed the experiments. YRL analyzed the data. XPZ
wrote the manuscript. MC conducted data analysis and edited the
manuscript. HTY was involved in performing the animal behavioral
tests. GW served an important role in the establishment of the
hyperlipidemia animal model. MH determined the dose of Dex used in
the experiments. DBL provided guidance during the whole process and
assisted writing the article, and ensured the integrity of
experiments and approved the final version. All authors edited and
approved the submission and agree to be accountable for all aspects
of the research in ensuring that the accuracy or integrity of any
part of the work are appropriately investigated and resolved. DBL
is the corresponding author.
Ethics approval and consent to
participate
All animal procedures were approved by the Animal
Experiment Ethics Committee of Dalian Medical University, China.
All studies involving animals are reported in accordance with the
ARRIVE guidelines for reporting experiments involving animals.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
BBB
|
blood-brain barrier
|
|
POCD
|
postoperative cognitive
dysfunction
|
|
CNS
|
central nervous system
|
|
Mfsd2a
|
Major facilitator superfamily
domain-containing protein 2
|
|
TNF-α
|
tumor necrosis factor-α
|
|
IL-1β
|
interleukin-1β
|
|
TRPM2
|
transient receptor potential cation
channel, subfamily M, member 2
|
|
TRPV1
|
transient receptor potential cation
channel, subfamily V, member 1
|
|
TGs
|
triglycerides
|
|
TC
|
total cholesterol
|
|
MWM
|
Morris water maze
|
|
OD
|
optical density
|
|
qPCR
|
quantitative real-time PCR
|
|
Aβ
|
β-amyloid protein
|
|
HDL
|
high-density lipoprotein
|
|
VLDL
|
very low-density lipoprotein
|
|
LDL
|
low-density lipoprotein
|
|
BDNF
|
brain-derived neurotrophic factor
|
|
LTP
|
long-term potentiation
|
References
|
1
|
Pappa M, Theodosiadis N, Tsounis A and
Sarafis P: Pathogenesis and treatment of post-operative cognitive
dysfunction. Electron Physician. 9:3768–3775. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bilotta F, Qeva E and Matot I: Anesthesia
and cognitive disorders: A systematic review of the clinical
evidence. Expert Rev Neurother. 16:1311–1320. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Price CC, Garvan CW and Monk TG: Type and
severity of cognitive decline in older adults after noncardiac
surgery. Anesthesiology. 108:8–17. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lin Y, Chen J and Wang Z: Meta-analysis of
factors which influence delirium following cardiac surgery. J Card
Surg. 27:481–492. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Loffler T, Flunkert S, Temmel M and
Hutter-Paier B: Decreased plasma Abeta in hyperlipidemic APPSL
transgenic mice is associated with BBB dysfunction. Front Neurosci.
10:2322016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ullrich C, Pirchl M and Humpel C:
Hypercholesterolemia in rats impairs the cholinergic system and
leads to memory deficits. Mol Cell Neurosci. 45:408–417. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Cunha S, Amaral MH, Lobo JM and Silva AC:
Therapeutic strategies for Alzheimer's and Parkinson's diseases by
means of drug delivery systems. Curr Med Chem. 23:3618–3631. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Acharya NK, Goldwaser EL, Forsberg MM,
Godsey GA, Johnson CA, Sarkar A, DeMarshall C, Kosciuk MC, Dash JM,
Hale CP, et al: Sevoflurane and Isoflurane induce structural
changes in brain vascular endothelial cells and increase
blood-brain barrier permeability: Possible link to postoperative
delirium and cognitive decline. Brain Res. 1620:29–41. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ben-Zvi A, Lacoste B, Kur E, Andreone BJ,
Mayshar Y, Yan H and Gu C: Mfsd2a is critical for the formation and
function of the blood-brain barrier. Nature. 509:507–511. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Berger JH, Charron MJ and Silver DL: Major
facilitator superfamily domain-containing protein 2a (MFSD2A) has
roles in body growth, motor function, and lipid metabolism. PLoS
One. 7:e506292012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Guerriero F, Sgarlata C, Francis M,
Maurizi N, Faragli A, Perna S, Rondanelli M, Rollone M and Ricevuti
G: Neuroinflammation, immune system and Alzheimer disease:
Searching for the missing link. Aging Clin Exp Res. 29:821–831.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Barrientos RM, Hein AM, Frank MG, Watkins
LR and Maier SF: Intracisternal interleukin-1 receptor antagonist
prevents postoperative cognitive decline and neuroinflammatory
response in aged rats. J Neurosci. 32:14641–14648. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Jolkkonen J, Puurunen K, Koistinaho J,
Kauppinen R, Haapalinna A, Nieminen L and Sivenius J:
Neuroprotection by the alpha2-adrenoceptor agonist,
dexmedetomidine, in rat focal cerebral ischemia. Eur J Pharmacol.
372:31–36. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Mantz J, Josserand J and Hamada S:
Dexmedetomidine: new insights. Eur J Anaesthesiol. 28:3–6. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Venn RM, Bryant A, Hall GM and Grounds RM:
Effects of dexmedetomidine on adrenocortical function, and the
cardiovascular, endocrine and inflammatory responses in
post-operative patients needing sedation in the intensive care
unit. Br J Anaesth. 86:650–656. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Goyagi T: Dexmedetomidine reduced
sevoflurane-induced neurodegeneration and long-term memory deficits
in neonatal rats. Int J Dev Neurosci. 75:19–26. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Akpinar H, Naziroglu M, Ovey IS, Cig B and
Akpinar O: The neuroprotective action of dexmedetomidine on
apoptosis, calcium entry and oxidative stress in cerebral
ischemia-induced rats: Contribution of TRPM2 and TRPV1 channels.
Sci Rep. 6:371962016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhang C, Li J, Wang J, Song X, Zhang J, Wu
S, Hu C, Gong Z and Jia L: Antihyperlipidaemic and hepatoprotective
activities of acidic and enzymatic hydrolysis exopolysaccharides
from Pleurotus eryngii SI-04. BMC Complement Altern Med.
17:4032017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tomas Pereira I and Burwell RD: Using the
spatial learning index to evaluate performance on the water maze.
Behav Neurosci. 129:533–539. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hovens IB, Schoemaker RG, van der Zee EA,
Absalom AR, Heineman E and van Leeuwen BL: Postoperative cognitive
dysfunction: Involvement of neuroinflammation and neuronal
functioning. Brain Behav Immun. 38:202–210. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hu N, Wang C, Zheng Y, Ao J, Zhang C, Xie
K, Li Y, Wang H, Yu Y and Wang G: The role of the
Wnt/β-catenin-Annexin A1 pathway in the process of
sevoflurane-induced cognitive dysfunction. J Neurochem.
137:240–252. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Vlisides P and Xie Z: Neurotoxicity of
general anesthetics: an update. Curr Pharm Des. 18:6232–6240. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Bilotta F, Gelb AW, Stazi E, Titi L,
Paoloni FP and Rosa G: Pharmacological perioperative brain
neuroprotection: A qualitative review of randomized clinical
trials. Br J Anaesth. 110 (Suppl 1):i113–i120. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Di Cataldo V, Geloen A, Langlois JB,
Chauveau F, Theze B, Hubert V, Wiart M, Chirico EN, Rieusset J,
Vidal H, et al: Exercise does not protect against peripheral and
central effects of a high cholesterol diet given ad libitum
in old ApoE−/− mice. Front Physiol. 7:4532016.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Vorhees CV and Williams MT: Morris water
maze: Procedures for assessing spatial and related forms of
learning and memory. Nat Protoc. 1:848–858. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Varatharaj A and Galea I: The blood-brain
barrier in systemic inflammation. Brain Behav Immun. 60:1–12. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Elwood E, Lim Z, Naveed H and Galea I: The
effect of systemic inflammation on human brain barrier function.
Brain Behav Immun. 62:35–40. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhao Z and Zlokovic BV: Blood-brain
barrier: A dual life of MFSD2A? Neuron. 82:728–730. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Paeschke N, von Haefen C, Endesfelder S,
Sifringer M and Spies CD: Dexmedetomidine prevents
lipopolysaccharide-induced microRNA expression in the adult rat
brain. Int J Mol Sci. 18:E18302017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wang D, Xu X, Wu YG, Lyu L, Zhou ZW and
Zhang JN: Dexmedetomidine attenuates traumatic brain injury: Action
pathway and mechanisms. Neural Regen Res. 13:819–826. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Tanabe K, Matsushima-Nishiwaki R, Kozawa O
and Iida H: Dexmedetomidine suppresses interleukin-1beta-induced
interleukin-6 synthesis in rat glial cells. Int J Mol Med.
34:1032–1038. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Terrando N, Monaco C, Ma D, Foxwell BM,
Feldmann M and Maze M: Tumor necrosis factor-alpha triggers a
cytokine cascade yielding postoperative cognitive decline. Proc
Natl Acad Sci USA. 107:20518–20522. 2010. View Article : Google Scholar : PubMed/NCBI
|