Introduction
Cervical cancer is the second most frequent cancer
in women with ~530,000 novel cases each year (1). It is well established that persistent
infection with high-risk oncogenic human papillomavirus (HPV) is
the main etiological agent and initial step in cervical
carcinogenesis (2). Although, the
best characterized target of the E6 protein of HPV is p53, HPV E6
binds to a diverse range of cellular proteins by altering its
stability in a proteasome-dependent manner (3,4). In
p53 degradation, the central pocket of E6 directly binds to the
LxxLL motif of the ubiquitin ligase E6AP, which induces a
conformational change in HPV E6 allowing the formation of a complex
with p53 (5,6). In this ternary complex, named
E6/E6AP/p53, p53 is polyubiquitinated by E6AP and subsequently
degraded by a proteasome (7,8).
Therefore, p53 responses such as those resulting from DNA damage,
which includes cell cycle arrest, DNA damage repair and apoptosis,
are deficient. Hence, the cell is more susceptible to the
acquisition of genomic instability and finally cell transformation
(9). Therefore, the reactivation
of p53 has been suggested to be an effective strategy for cancer
therapy in wild-type p53 retaining tumor cells, as its reactivation
can result in cell cycle arrest and/or apoptosis by activating or
repressing the transcription of apoptotic genes and cell cycle
regulatory genes (10).
Flavonoids are widely distributed in plants and are
associated with a diverse range of biological activities that
include inhibition of phosphoinositide-3-kinase, reactive oxygen
species (ROS) generation, DNA damage, cell cycle arrest and
apoptosis, as well as p53 restoration (11–13).
For instance, the catechin-like flavonoid, epigallocatechin gallate
(EGCG) increased the level of the p53 protein accompanied by a
reduction of HPV E6 protein in HeLa and CaSki cells (14). Furthermore, the flavonoid luteolin
and flavonoid-like synthetic compounds inhibited the binding
between HPV E6 and E6AP in vitro and induced an increased
expression of the p53 and p21 proteins in cervical cancer cells
(15).
Several studies have demonstrated the anticancer
activity of quercetin, a polyphenolic flavonoid, against a number
of types of cancer cells, such as hepatocellular carcinoma cells
where quercetin inhibited the cell proliferation through cell cycle
arrest, apoptosis and DNA fragmentation, together with an increase
of the total p53 protein and p53 phosphorylation (16). In addition, in melanoma cells,
quercetin induced apoptosis by a p53/Bax-dependent mechanism and
was correlated with an increase in ROS (17). However, a common mechanism for
quercetin-induced p53 restoration has not been well established in
HPV-positive cervical cancer cells.
In the present study, it was reported that quercetin
arrested the cell cycle in G2 phase and triggered apoptosis in
cervical cancer cells. Also, it was observed that quercetin
promoted the activation of p53 by an increase of total p53 protein
and its nuclear localization, together with the increase of
expression of its transcriptional targets including Bax and p21.
However, quercetin did not decrease the expression of HPV E6, the
agent responsible for the decrease of p53 in these cells. In
addition, the molecular docking results predict that quercetin
would be able to interrupt the association of E6 with E6AP by
binding to the E6 pocket and therefore preventing the formation of
the p53 binding cleft on E6 and finally p53 degradation.
Materials and methods
Cell lines and treatments
Human cervical cancer cells expressing HPV-16 (SiHa
cells), HPV 18 (HeLa cells) were obtained from the American Type
Culture Collection (Manassas, VA, USA) and human foreskin
fibroblasts (HFF cells) were kindly provided by Dr. Ramón González
(CIDC, UAEM, Cuernavaca, Mor, México). All the cells were
maintained in Dulbecco's Modified Eagle's Medium High Glucose (DMEM
HG, Caisson Labs, UT, USA) supplemented with 10% (v/v) Fetal Bovine
Serum (Biowest LLC, MO, USA) at 37°C in a humidified atmosphere of
5% CO2. Treatment with quercetin or taxol (Sigma
aldrich; St. Louis, MO, USA) did not exceed 0.5% of DMSO.
Cell viability
Cell viability was measured using
[3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium]
inner salt MTS assay (Promega, Madison WI, USA). Briefly, a total
of 4X103 SiHa, HeLa or HFF cells per well were plated in
a 96-well plate and allowed to grow during overnight. Cells were
exposed to increasing concentrations of quercetin by triplicate for
48 h. Subsequently, 20 µl of MTS reagent was added into each well
containing the untreated and treated cells in 100 µl DMEM HG and
incubated at 37°C for 3 h. Then the absorbance values were measured
at 490 nm in an automatic microplate reader (Promega, Madison, WI,
USA). Data were analyzed, and cell viability rate was calculated in
GraphPad PRISM version 6.01 statistical program and the
IC50 values were determined by regression analysis.
Flow cytometry
HeLa and SiHa cells were treated with quercetin at
IC50, whilst HFF cells were exposed to 500 µM quercetin
during 48 h. The cells were separately treated with 5 nM taxol (as
G2/M control). Control and treated cells were harvested,
centrifuged and fixed in cold 70% ethanol. Fixed cells were
incubated with 10 µg/ml ribonuclease A and 10 µg/ml propidium
iodide during 30 min on ice. Then 10,000 events were acquired in
flow cytometer (FACSCalibur; Beckman Coulter, Inc., Brea, CA, USA).
Obtained data were analyzed using the FlowJo Software (Tree Star,
Inc., Ashland, OR, USA) to generate DNA content frequency
histograms. The experiments were conducted by triplicate in three
independent experiments and the statistical analysis was performed
using ANOVA and subsequent Dunnett test with P<0.05 considered
to indicate a statistically significant difference.
Mitotic nuclei staining
HeLa and SiHa cells were grown on slide and treated
with quercetin and taxol as described in flow cytometry assay. The
cells were fixed with 4% paraformaldehyde in PEM buffer (PIPES 100
mM pH 6.9, EGTA 5 mM, MgCl2 2 mM) for 15 min followed 4%
paraformaldehyde in NaHCO3 for 45 min. The cells were
permeabilized with 0.1% Triton in PBS 1X for 10 min. Nuclei were
stained with DAPI 0.2 µg/ml, at 37°C for 10 min. Cell images were
observed using a fluorescence microscope and acquired by
NIS-Elements software. At least 200 nuclei were counted from
different fields acquired in distinct days to determine the
percentage of mitotic nuclei.
Epifluorescence microscopy
HeLa or SiHa cells were exposed to their respective
quercetin IC50. HFF cells were exposed at 140 µM
quercetin, the highest concentration employed in cervical cancer
cells. After 72 h, attached cells on slide were stained with
acridine orange/ethidium bromide (AO/EB, 1:1 molecular relation).
The images were immediately captured using an epifluorescence
microscope to determinate quercetin-induced apoptotic traits.
Different controls were included in the experiment: Cells not
treated, as negative control; cell treated at 1 mM
H2O2 for 3 h, as apoptosis control; and cells
immersed in 80–90°C water for 15 sec, as necrosis control. It is
well known that AO can pass through cell membrane, but EB cannot.
Necrotic cells stain red but have a nuclear morphology resembling
that of viable cells. Apoptotic cells appear green, and
morphological changes such as formation of apoptotic bodies are
observed. The criteria for identification are as follows: Viable
cells appear to have green nucleus with intact structure; early
apoptosis cells exhibit a bright green nucleus showing condensation
of chromatin; late apoptosis appears as dense orange areas of
chromatin condensation; and orange intact nucleus depicts secondary
necrosis (18,19). The experiments were conducted by
triplicate in three independent experiments.
Immunofluorescence of p53 protein
50,000 HeLa or SiHa cells were grown on glass
coverslips in a 24-well cell culture plate and allowed to attach
during overnight. Upon quercetin IC50 treatment, both
HeLa and SiHa cells were fixed and permeabilized as mentioned in
the Mitotic nuclei staining section. The cells were then incubated
with the primary antibody mouse anti-p53 (1:250; DO-1, Santa Cruz
Biotechnology, Dallas, TX, USA) overnight at 4°C. The secondary
antibody anti-mouse Alexa 488 (1:1,000) was added and incubated for
two h at 37°C to visualize the p53 localization. The nuclei were
stained with DAPI (0.2 µg/ml). The images were acquired in confocal
microscopy. The acquisitions for DAPI and Alexa488 were made
sequentially to avoid crossover of fluorescence emission. These
experiments were conducted by duplicated in two different days.
Western blot analysis
Following quercetin treatment, HeLa and SiHa cells
were lysed with RIPA buffer (Tris-HCl 50 mM, NaCl 150 mM, NP40 1%,
sodium deoxycholate 0.5%, SDS 0.1%) containing protease inhibitor
cocktail (Sigma, MO, USA). Protein concentration was determined by
bicinchoninic acid method using Pierce™ BCA Protein
Assay kit (Pierce biotechnology, IL, USA). Equal protein amounts
from all samples were loaded and resolved in SDS-PAGE 12% and
transferred to PVDF membrane. The membranes were blocked with 5%
milk in TBS-Tween-20 for 1 h, and then incubated overnight at 4°C
with the following primary antibodies: p53 (DO-1, 1:1,000
dilution), HPV18 E6 (G-7, 1:200 dilution), HPV16/18 E6 (C1P5, 1:100
dilution), Bax (2D2, 1:500 dilution), Bcl-2 (C-2, 1:500), from
Santa Cruz Biotechnology. The proteins were visualized using the
ChemiDoc-XRS Imaging System (Bio-Rad Laboratories, Inc. CA, USA),
using supersignal west femto maximum sensitivity substrate (Thermo
Fischer Scientific, IL, USA) as a developer. GAPDH antibody primary
(GA1R) from Santa Cruz Biotechnology was used as the loading
control. The experiment was executed in triplicate and
quantification was performed via densitometric analysis using
ImageJ TM software. The densitometric data were normalized with the
loading control. The level of the expression of the proteins was
determined in function to the respective control.
RT-PCR
To determinate the level of transcript of p21 gene,
total RNA from SiHa and HeLa cells were obtained following 48 h of
quercetin IC50 exposure. 50 ng of DNase I-treated total
RNAs were retro-transcribed and PCR amplified in one step as
follow: cDNA synthesis, 50°C for 30 min; retrotranscriptase
inactivation, 95°C for 15 min; 35 cycles of DNA amplification at
95°C for 30 sec, 52°C for 30 sec, 72°C for 1 min; final extension,
72°C for 5 min, in Mastercycler gradient Serial No. 5331
(Eppendorf, Hamburg, Germany) by using One-Step RT-PCR Hot-Start
kit (Thermo Fisher, CA, USA). Specific oligonucleotides of p21
(GenBank reference sequence: NM_001291549) (forward sequence,
5′→3′TTAGCAGCGGAACAAGGAGT and reverse sequence,
5′→3′AGAAACGGGAACCAGGACAC; product size, 252 bp) and of the loading
control, GAPDH (GenBank reference sequence, NM_002046.6) (forward
sequence, 5′→3′CAACGACCACTTTGTCAAGC and reverse sequence,
5′-GGTGGTCCAGGGGTCTTACT-3′; product size, 115 bp). RT-PCR reactions
were resolved in agarose gel 1.2% and visualized in ChemiDoc XRS
Imaging System (Bio-Rad Laboratories, Inc, CA, USA). The
densitometric analysis was performed as mentioned in the Western
blot section.
Caspase 3/7 activity assay
4×103 HeLa or SiHa cells were plated in 96-well
plate and allowed overnight. Then, cells were treated at
IC50 of quercetin for 72 h. After treatment, the
caspasas 3/7 activity was tested by Caspase-Glo 3/7 Assay from
Promega, following manufacturer's instructions. The results were
represented as relative units of luminescence and represented in
graphs. The statistical analysis was performed by One-way ANOVA
with P<0.05 considered to indicate a statistically significant
difference.
Molecular docking
The HPV16 E6 crystal structure bound to LxxLL E6AP
motif and fusioned to Maltose Binding Protein (MBP) was taken from
PDB (ID: 4GIZ) (20). Missing
hydrogens from E6 were added using charmm-gui server. The energy
from E6 structure was minimized to avoid any bad contacts with 100
steepest descent steps using charmm version 42 with charmm36
parameters. Marvin was used for drawing, displaying and
characterizing chemical structures of quercetin, luteolin, CAF24
and C170. Minimized E6 structure was used as target to conduct
1,000 independent blind docking runs. The initial pdbqt files and
the configuration files were set using Pymol and Autodock Vina
Plugin for Pymol. The grid center was set on (3.76, 60.0, 30.29)
with a size of 50×40×60Å using 1Å of spacing size. The analysis was
made using local unix scripts. Final structure images were made
using VMD molecular software. Protein-ligand interactions maps were
made with Maestro.
Statistical analysis
Data were analyzed using the GraphPad Prism version
6.01 statistical program (GraphPad Software, Inc.). Statistical
differences between groups were evaluated using one-way analysis of
variance followed by the Dunnett's test or t-test. P<0.05 was
considered to indicate a statistical significance.
Results
Cytotoxic effect of quercetin in
cervical cancer cells
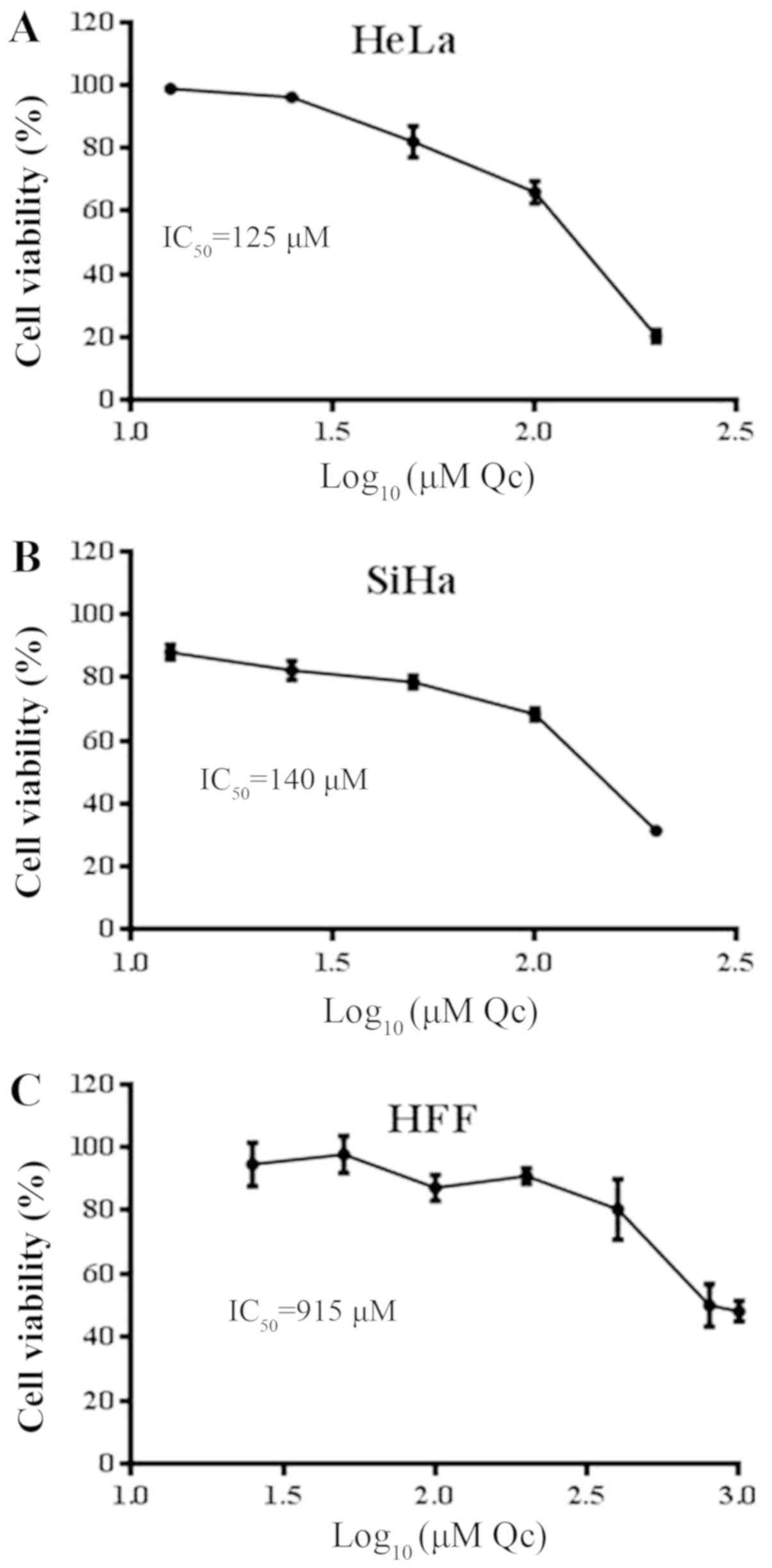
To determine the cytotoxic effect of quercetin on
HeLa and SiHa cells, a cell viability assay was performed following
48 h of quercetin exposure. The results demonstrated that quercetin
decreased the viability of the HeLa and SiHa cells at a similar
concentration, with an IC50 of 125 and 140 µM,
respectively (Fig. 1A and B).
Nevertheless, in HFF cells quercetin required ~1,000 µM to reach
the cytotoxic effects demonstrated in cervical cancer cells
(Fig. 1C). This result
demonstrates that quercetin exhibits selective toxicity for the
cervical cancer cells, HeLa and SiHa, compared with the HFF
cells.
Induction of cell cycle arrest by
quercetin in cervical cancer cells
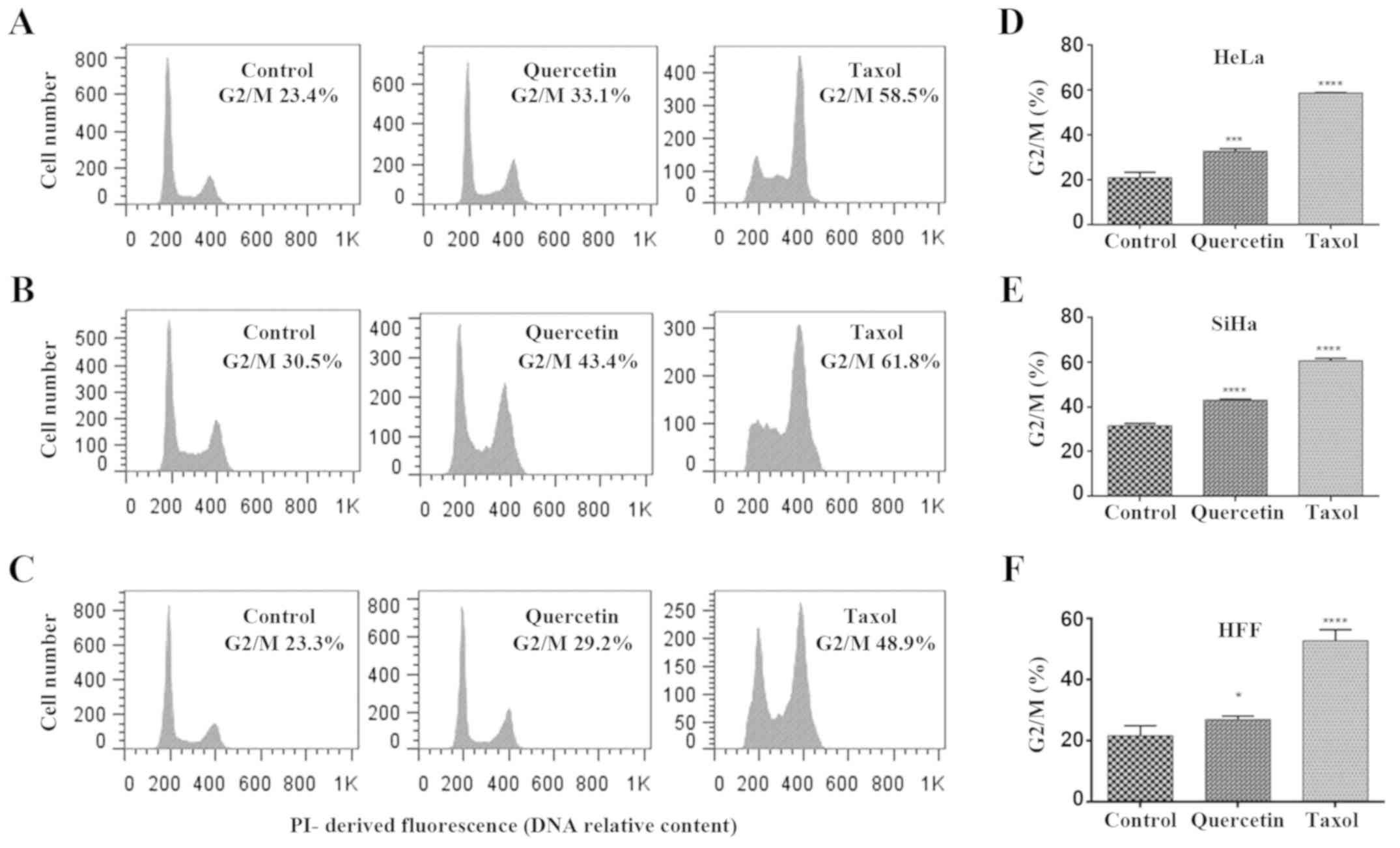
Quercetin has demonstrated multiple biological
activities, including cell cycle arrest. To investigate whether the
quercetin-induced toxicity is due to its effect on cell
proliferation, cell cycle analysis was performed by flow cytometry
(Fig. 2). The results revealed
that quercetin did not induce significant changes in HFF cells at
140 µM (data not shown), whilst at a high concentration (500 µM) a
barely significant increase in the cell population of G2/M phase
(with 4n DNA content) was reached (Fig. 2C and D). Furthermore, quercetin at
IC50 induced a significant increase of G2/M phases in
HeLa and SiHa cells (Fig. 2A-F),
as did treatment with taxol (used as a positive control). Taxol is
a known mitosis blocker due to its ability to bind tubulin,
resulting in stabilization of microtubules, which blocks
metaphase/anaphase progression (21). However, quercetin does not have a
unique mechanism of cell cycle arrest. To discriminate between
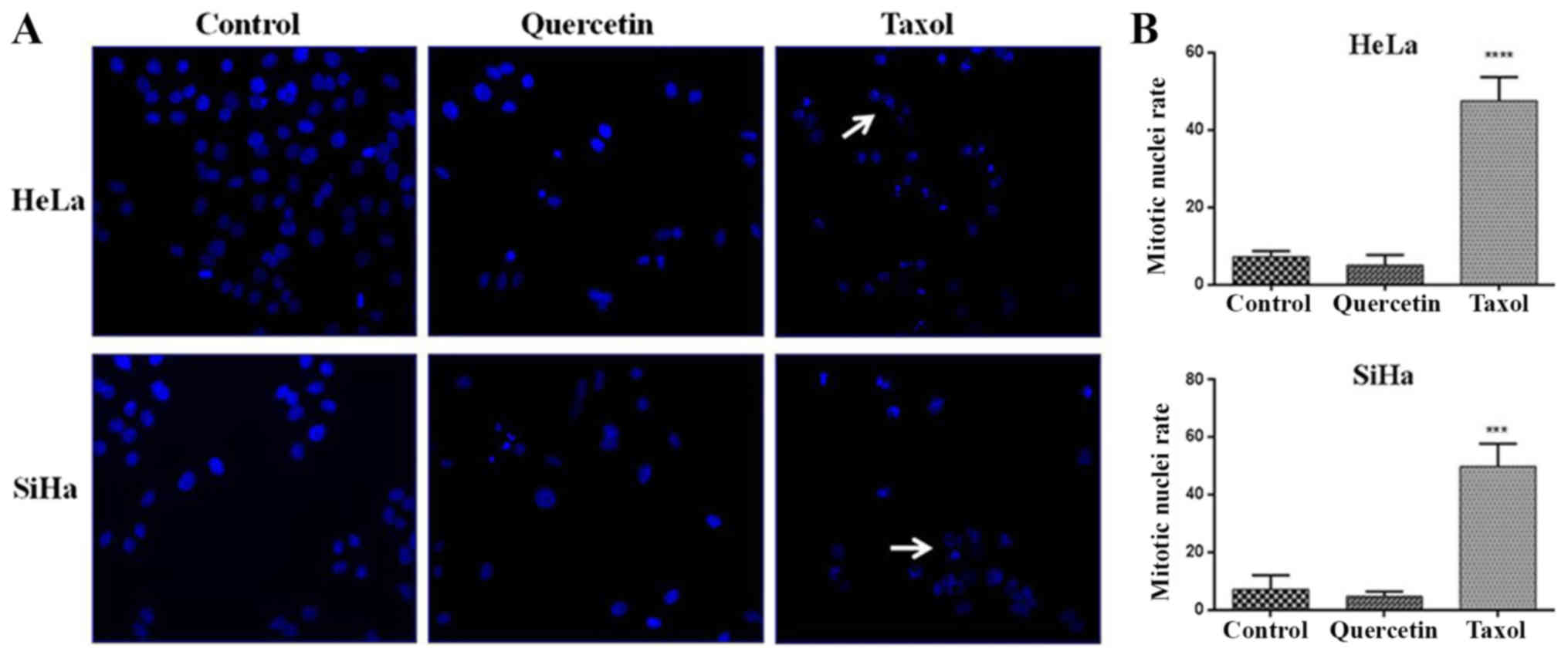
quercetin-induced G2 and M phase arrest in HeLa and SiHa cells,
DAPI-stained nuclei were analyzed by epifluorescence microscopy
following 48 h of quercetin treatment. Following exposure to taxol,
a significant increase of mitotic nuclei in HeLa and SiHa cells was
observed, whilst significant changes were not detected when HeLa
and SiHa cells were challenged with quercetin (Fig. 3A and B). These results indicate
that quercetin arrested in the G2 phase rather than the M phase of
the cell cycle in cervical cancer cells.
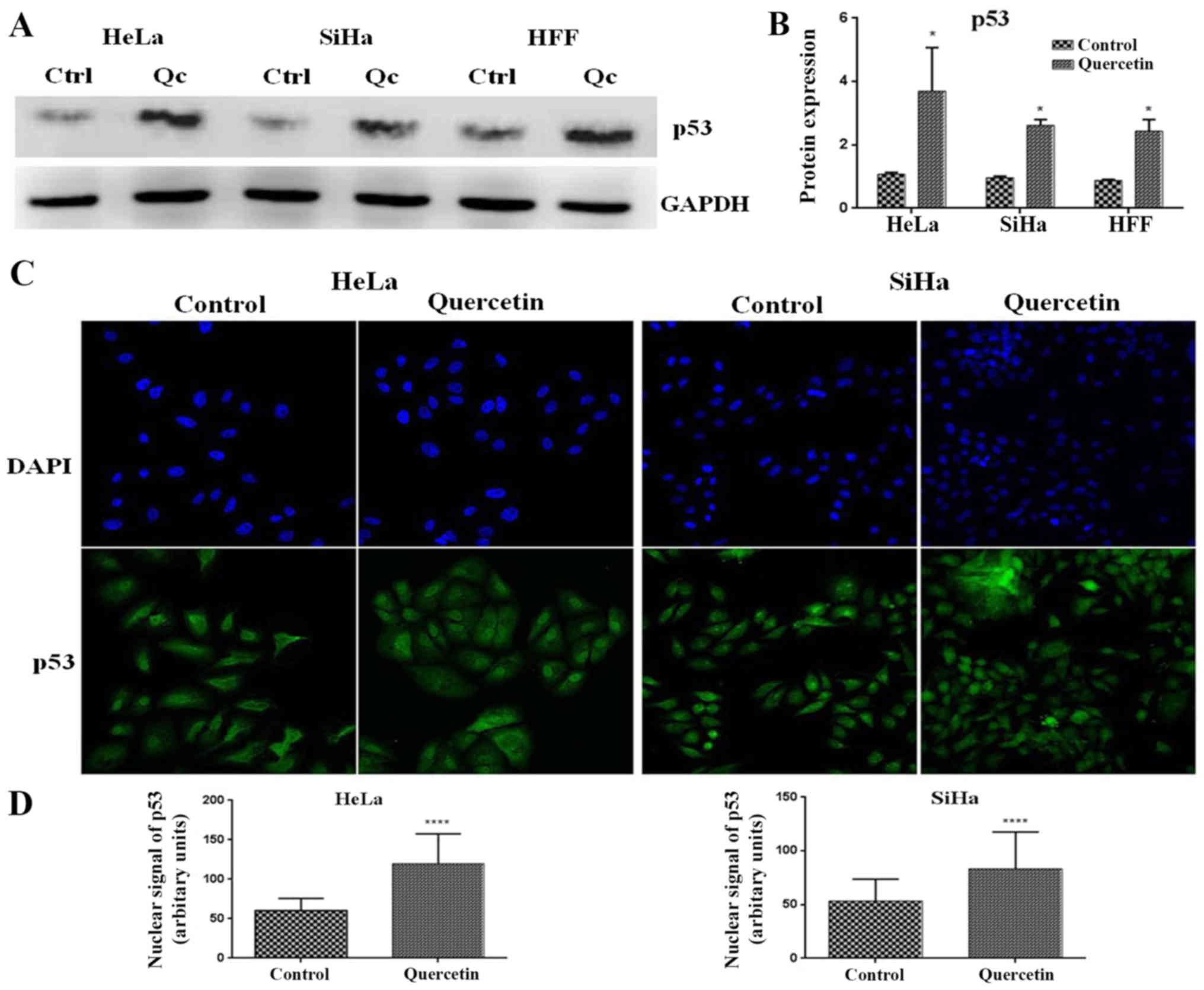
Effect of quercetin on p53 expression
and its cellular localization in cervical cancer cells
p53 is a master cell cycle regulator, which is able
to induce the expression of genes involved in different checkpoints
of the cell cycle. To investigate whether quercetin modified the
expression of p53 in cervical cancer cells, western blotting was
performed. It was observed that quercetin significantly increased
the level of p53 in HeLa, SiHa and HFF cells (Fig. 4A and B). Furthermore, it is well
known that p53 nuclear translocation is necessary for its
transcriptional activity and cell cycle regulation effects.
Notably, it was observed that quercetin significantly increased the
p53 nuclear signal in HeLa and SiHa cells (Fig. 4C and D). These results suggest that
quercetin promotes the nuclear activities of p53 in HeLa and SiHa
cells.
Upregulation of target genes of
p53
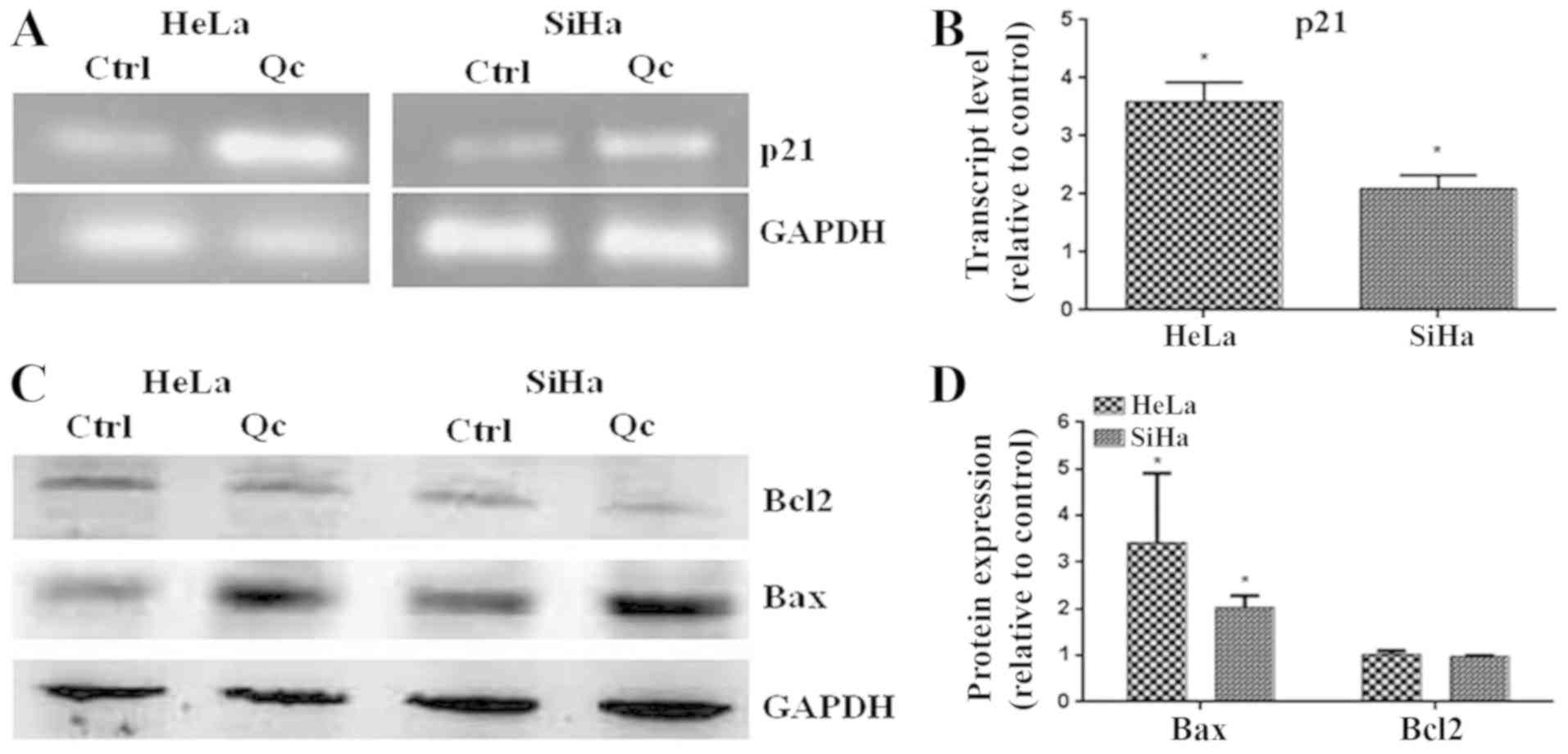
To demonstrate the activity of p53 induced by
quercetin, the level of transcript of p21 and the levels of
pro-apoptotic Bax protein, two well-known target genes of p53, were
evaluated. The results demonstrated that quercetin increased the
level of the transcript of p21 in HeLa and SiHa cells (Fig. 5A and B). additionally, quercetin
significantly increased the level of the pro-apoptotic Bax protein
(Fig. 5C and D). These results
suggest that quercetin promotes the transcriptional activity of
p53.
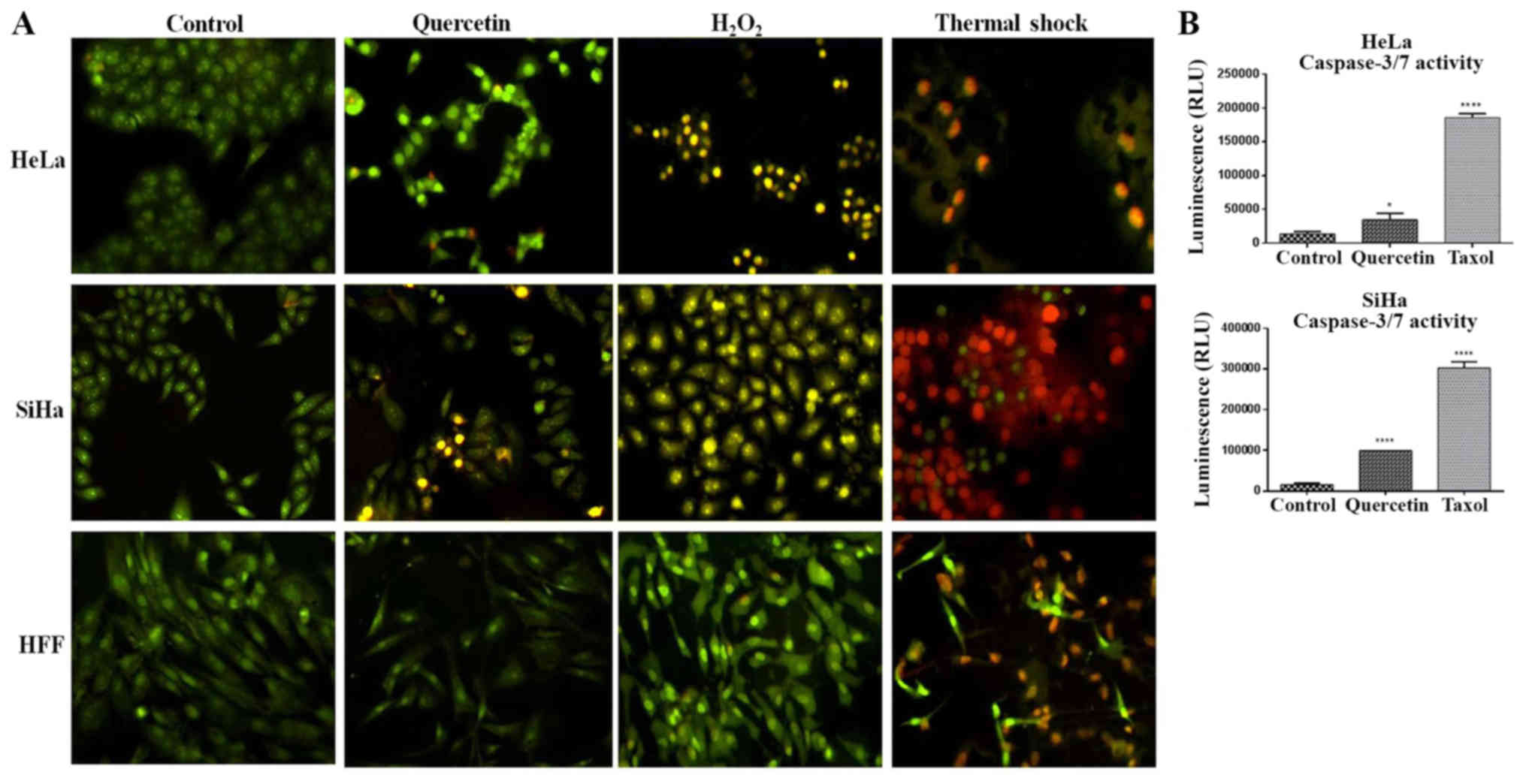
Induction of apoptosis by quercetin in
cervical cancer cells
Due to quercetin increasing the expression of p53
and Bax proteins, two known apoptotic genes, the effect of
quercetin in the induction of apoptosis in cervical cancer cells
was evaluated. Therefore, apoptotic morphological traits were
analyzed using AO/EB staining, as well as measuring the activity of
caspases 3/7. As presented in Fig.
6A, the nuclei of the untreated cells were homogenously stained
with AO in comparison to quercetin-treated cells or the apoptosis
positive control (H2O2), which displayed
intensely nuclei stained, presumably due to the chromatin
condensation. Furthermore, cells treated with quercetin, as in the
apoptosis control, exhibited membrane blebbing and apoptotic
bodies, which were more evident in the SiHa cells compared with the
HeLa cells. In addition, quercetin significantly induced the
activation of caspases 3/7 in HeLa and SiHa cells (Fig. 6B). These results together with the
augmentation of the Bax/Bcl-2 ratio in HeLa and SiHa cells
(Fig. 5C and D), an early event of
mitochondrial outer membrane perforation and subsequent apoptosis,
indicate that quercetin triggers apoptosis in these cells.
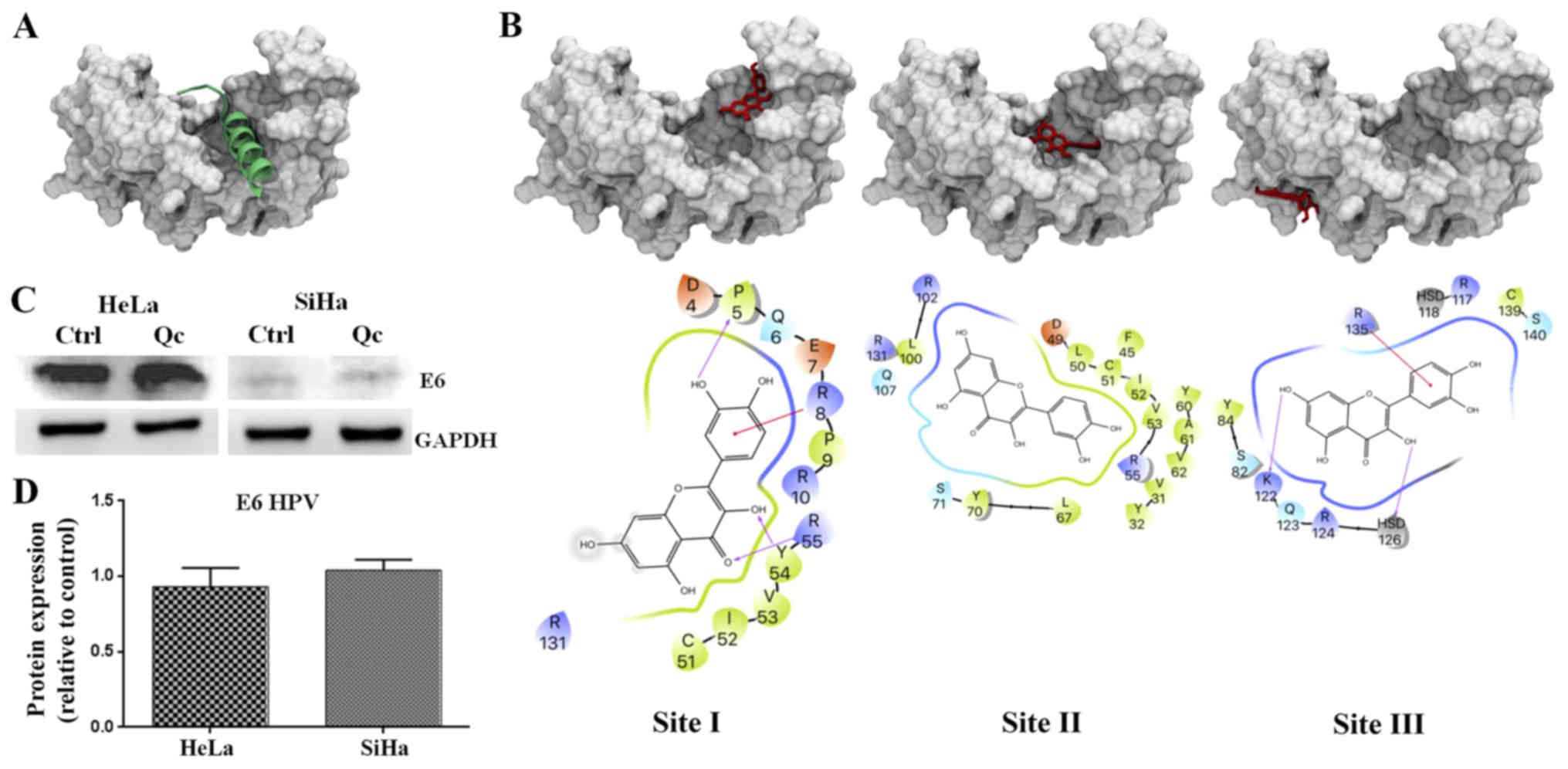
Quercetin-induced p53 activation is E6
HPV protein expression-independent
In HPV positive cervical cancer cells, the stability
of p53 is totally E6 dependent (22). Therefore quercetin, which caused an
activation of p53, would also be expected to induce a decrease in
E6 expression. However, western blotting demonstrated that HPV E6
expression remains constant despite the increase of p53 in HeLa and
SiHa cells (Fig. 7C). An
explanation for the activation of p53 by quercetin in cervical
cancer cells could be associated with the disruption of the E6/E6AP
complex assembly rather than downregulation of E6 expression. This
assumption is supported by the report that flavonoids structurally
similar to quercetin were capable of interrupting E6-E6AP
association in vitro (15).
Quercetin is predicted to bind to the
E6AP binding pocket in HPV E6
To predict the binding between quercetin and E6
protein, the quercetin ligand was subjected to 1,000 independent
molecular blind docking assays against the E6 protein structure
(PDB ID: 4GIZ) (20). The same
operation was performed for flavonoids luteolin, CAF24 and C170
(reference ligands), which were demonstrated by Cherry et al
in 2013 to be capable of binding specifically to HPV E6 protein to
disrupt the association with E6AP (Fig. 7A) (15). The results predicted that all
ligands, including quercetin, were able to interact with the E6
pocket, necessary for the binding with E6AP. Docking predicted that
quercetin could bind in three sites on the E6 protein (Fig. 7B). One of them (known as site II)
was the E6 pocket site where quercetin bound with the lowest energy
(−7.08±-0.18; Fig. 7B, middle
panel). Notably, quercetin in site I (Fig. 7B, left panel) made contact with the
Pro5, Gln6, Glu7, Arg8, Pro9 and Arg10 residues of E6 (Table I) that were previously described to
participate in E6-p53 interactions (20). Quercetin in site II made contact
with Asp49 of E6 which establishes polar interactions with His115
in the L1 loop of p53. Also, in site II quercetin contacted
residues Leu50, L100 and Arg102 of E6 that impact on the
architecture of E6 necessary for competent interaction with p53,
which are discussed below. These results predict that quercetin
would be able to interrupt the association of E6 with E6AP by
binding to the E6 pocket and therefore preventing the formation of
the p53 binding cleft on E6 and finally p53 degradation. Taken
together, these results suggest that quercetin reactivates p53,
preventing the formation of the E6/E6AP complex, leading to cell
cycle arrest in the G2 phase and apoptosis.
 | Table I.Summary of the contacts of each
ligand with the residues of the HPV E6 protein (PDB ID: 4GIZ). |
Table I.
Summary of the contacts of each
ligand with the residues of the HPV E6 protein (PDB ID: 4GIZ).
| Ligand | Site 1 | Site 2 | Site 3 | Energy (kcal/mol)
in site 2 |
|---|
| Quercetin | D4, P5, Q6, E7, R8,
P9, R10, C51, I52, V53, Y54, R55, R131. | V31, Y32, F45, D49,
L50, C51, I52, V53, R55, Y60, A61, V62, L67, Y70, S71, L100, R102,
Q107, R131. | S82, Y84, R117,
H118, K122, Q123, R124, H126, R135, C139, S140. | −7.08±-0.18 |
| CAF24 | P5, Q6, E7, R8, P9,
R10, C51, I52, V53, Y54, R55. | R10, K11, V31, Y32,
F45, D49, L50, C51, V53, R55, A61, V62, L67, Y70, S71, I73, S74,
R77, H78, R102, Q107, I128, R131, T133. | K72, I73, E75, Y76,
Y79, S82, R124, H126, R135. | −7.99±-0.18 |
| C170 | * | P5, R8, P9, R10,
K11, V31, Y32, F45, D49, L50, C51, I52, V53, Y54, R55, Y60, A61,
V62, C66, L67, F69, Y70, S71, I73, S74, E75, R77, H78, L100, R102,
C103, I104, Q107, I128, R129, R131, T133. | * | −6.8±-0.17 |
| Luteolin | D4, P5, Q6, E7, R8,
P9, R10 C51, I52, V53, Y54, R55, R131. | R10, K11, V31, Y32,
F45, R48, D49, L50, C51, I52, V53, Y60, A61, V62, C66, L67, Y70,
S71, S74, Q75, H78, L100, R102, G107, R131. | S82, Y84, R117,
H118, K122, G123, R124, H126, R135, S140. | −6.82±-0.19 |
Discussion
Currently vaccination against most prevalent
oncogenic HPVs as well as cervical cancer screening employed mainly
in developed countries, has reduced the incidence and the mortality
associated with this malignancy (23,24).
However, cervical cancer is still one of the most common types of
cancer in women worldwide. Aside from chemotherapy there are no
specific treatments for HPV. The development of safe and effective
drugs with minimal side effects is required for cervical cancer. In
fact, the results of the present study demonstrated that quercetin
possesses a selective toxicity towards cervical cancer derived
cells infected with HPV16 (SiHa cells) and HPV18 (HeLa cells),
[which are responsible for ~70% of all cervical cancer cases
worldwide (1)] compared with HFF
cells (considered normal cells), which required around a 7–8 fold
higher concentration to exhibit the cytotoxic effects presented in
the cervical cancer cell lines. In general, the quercetin
IC50 in cervical cancer cells lines resulted high, which
is a common feature for flavonoids (13,25).
Indeed, further in vivo studies need to be performed to
verify the pharmacokinetic of quercetin, as well as the synthesis
of analogs to enhance its potency in future works.
Quercetin at high concentrations (>40 µM) is able
to act as a prooxidant molecule causing DNA damage and resulting in
cell cycle arrest and/or p53-dependent or independent mitochondrial
apoptosis (12,25). Flow cytometry assays demonstrated
that in SiHa and HeLa cells quercetin induced an increase in the
G2/M phase cell population, whilst in HFF cells, the increase in
the G2/M phase was barely significant and required a ~3-fold higher
concentration of quercetin, which indicated a selectivity to
cervical cancer cells in comparison with HFF cells. Furthermore,
this arrest in G2/M phase did not correlate with an increase of the
mitotic nuclei rate in HeLa and SiHa cells following quercetin
treatment, suggesting that the arrest occurred in the G2 phase.
Furthermore, the G2 phase cell cycle arrest was accompanied by the
restoration of p53, by the increase of total p53 protein and its
nuclear localization in HeLa and SiHa cells following quercetin
treatment. In fact, a common feature between all these cell types
is the status of the wild type p53, whose restoration could
activate the p53 responses, such as cell cycle arrest and
apoptosis, which has been widely studied (23).
The best-characterized p53 target gene is p21, for
which the promoter contains two highly conserved p53-responsive
elements on which p53 directly binds to activate p21 transcription
(26). In response to DNA damage,
p53 induces p21 transcription and then the p21 protein can
associate and inhibit the mitosis-promoting factor, cyclin
dependent kinase1-Cyclin B1 to arrest in the G2 phase (27). In the present study, an
accumulation of the p21 transcript in HeLa and SiHa cells was
observed following exposure to quercetin, demonstrating the
reactivation of p53 as a transcriptional factor. Therefore, the
upregulation of p21 together with the reactivation of p53 strongly
suggest its participation in the G2 phase cell cycle arrest.
In addition to the increase of the p21 transcript by
p53, the activation of p53 by quercetin also induced high levels of
the pro-apoptotic protein Bax. Notably, mitochondrial apoptosis
regulated by p53 can occur by activating or repressing the
transcription of pro-apoptotic (Bax, Bid, and Noxa) or survival
(Bcl-2 and Bcl-xL) genes, respectively (28–30).
In addition, p53 can also trigger mitochondrial apoptosis through
activation of Bak or inhibition of Bcl-2 by direct association. The
increase of the Bax/Bcl-2 ratio results in the formation of the
mitochondria outer membrane pore, cytochrome release and the
subsequent activation of caspases 3/7 (31,32).
Furthermore, quercetin induced caspases 3/7 activity, as well as
morphological cell traits associated to apoptosis in HeLa and SiHa
cervical cancer cells. This result matches with a previous study in
which quercetin induced mitochondrial apoptosis (33).
p53 inactivation by HPV E6 is a general feature in
cervical cancer. Therefore, identifying the inhibition of HPV E6 in
cervical cancer is of interest in order to reactivate p53 and
sensitize cervical cancer cells to apoptosis. Notably, the increase
of the p53 protein in HeLa and SiHa cells did not alter the
expression of the HPV E6 protein. This result contrasts with the
finding reported by Qiao et al (14) in which they demonstrated that
treatment with the flavonoid EGCG in HeLa and CaSki cells caused an
increase of p53 accompanied by a decrease of HPV E6 protein.
Furthermore, it corresponds to the published data by Malecka et
al (34) in which the
flavonoid gossypetin with only an additional hydroxyl in the C8
position compared with quercetin, disrupted the E6/E6AP
association. Similar results were previously described by Cherry
et al (15), which clearly
demonstrated that in vitro, luteolin (lacking a hydroxyl in
the C3 position with respect to quercetin) and synthetic flavonoids
disrupted the binding of HPV 16 E6 and the core 70 amino acids
containing the E6 binding region (LxxLL) of E6AP and inhibited the
E6-induced p53 degradation in rabbit reticulocyte lysate.
The crystallographic structure of the HPV-16 E6
protein (PDB ID: 4GIZ) in complex with the LxxLL peptide of E6AP
and p53 core was solved. It was demonstrated that the LxxLL motif
E6AP binds to a conserved pocket of the E6 protein causing a
conformational change of E6 to allow it to competently interact
with p53 (20), since neither E6
nor E6AP are separately able to associate with p53 (8). Based on the solved structure of
E6/E6AP/p53 and E6/E6AP, the critical E6 residues to interact with
E6AP and p53 were revealed and the functionality in p53 degradation
was demonstrated by mutagenesis (6,20).
In the present study it was demonstrated that quercetin in site I
from the E6 structure established contact with Pro5, Gln6, Arg8
and, Arg10 residues that bring together the N-terminal arm and α1
helix of E6 and N-terminal arm, β1 and β10 of p53. Moreover, E6
Glu7 residue establishes direct contacts with p53, while E6 Pro5,
Arg8 and Pro9 residues modulate the conformation of the N-terminal
region of E6. In fact, mutations in Pro5, Arg8 or Pro9 impair
ternary complex assembly and p53 degradation (35,36).
In addition, in site II corresponding to a hydrophobic pocket of E6
where the LxxLL motif E6AP binds, quercetin binding was predicted
to be more energetically favorable. In site II quercetin
established contacts with Asp49 residue whose substitution to
alanine impairs p53 ternary complex assembly and p53 degradation
suggesting its essential role in this process. Also, in site II,
quercetin made contact with Leu50, L100, Arg102 and Arg131 of E6.
Point mutations in the hydrophobic residue Leu50 have been
demonstrated to decrease LxxLL peptide binding and p53 degradation.
Hydrophobic interactions were established between the E6 residue
Leu100 and p53 core residues Leu114 and Trp146. Surrounding the
hydrophobic pocket, the charged residue Arg102 holds together the
E6 N domain, E6C domain, and the LxxLL motif of E6AP. Arg131
provides structurally equivalent contacts with the E6AP peptide.
Point mutations of these arginine residues lead to markedly
decreased LxxLL peptide binding and p53 degradation (20). These multiple contacts that
quercetin was predicted to establish with critical residues of E6,
which are established interactions with E6AP and p53, suggest that
quercetin could disrupt the E6AP/E6/p53 complex formation and
prevent p53 degradation. However, further in vitro or in
vivo binding assays need to be performed to confirm the
separation of p53 and E6/E6AP.
Finally, the present study proposes that quercetin
can reactivate p53 and induce G2 phase cell cycle arrest and
apoptosis in HPV-positive human cervical cancer-derived cells, by
inhibiting the interaction of E6/E6AP.
Acknowledgements
The authors thank Dr Ramón González from CIDC and Dr
Patricia García López INCan for the relevant suggestions and
criticisms to the project.
Funding
Aldo F. Clemente Soto acknowledges fellowship 379417
from CONACYT. Partial support from UAEM (grants nos.
SI-DGDI-UAEM/13/289) and PAPIIT-UNAM (IN210316) is
acknowledged.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
AFC-S and LG-M conceived the original idea and
planned the experiments of the project. AFC-S participated in the
experiments. AFC-S and ES-V acquired and analyzed the confocal
microscopy images. CM-P carried out the molecular docking. JNS-C
supervised the cell viability and flow cytometry assay. All authors
analyzed and interpreted the data. OP-Z supervised the project.
AFC-S wrote the manuscript with the support of LG-M. All authors
contributed and approved the final version of the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Marth C, Landoni F, Mahner S, McCormack M,
Gonzalez-Martin A and Colombo N; ESMO Guidelines Committee, :
Cervical cancer: ESMO clinical practice guidelines for diagnosis,
treatment and follow-up. Ann Oncol. 28 Suppl_4:iv72–iv83. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Dürst M, Gissmann L, Ikenberg H and zur
Hausen H: A papillomavirus DNA from a cervical carcinoma and its
prevalence in cancer biopsy samples from different geographic
regions. Proc Natl Acad Sci USA. 80:3812–3815. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Gardiol D, Kühne C, Glaunsinger B, Lee SS,
Javier R and Banks L: Oncogenic human papillomavirus E6 proteins
target the discs large tumour suppressor for proteasome-mediated
degradation. Oncogene. 18:5487–5496. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Nakagawa S and Huibregtse J: Human
scribble (Vartul) is targeted for ubiquitin-mediated degradation by
the high-risk papillomavirus E6 proteins and the E6AP
ubiquitin-protein ligase. Mol Cell Biol. 20:8244–8253. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ansari T, Brimer N and Vande Pol SB:
Peptide interactions stabilize and restructure human papillomavirus
type 16 E6 to interact with p53. J Virol. 86:11386–11391. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Martinez-Zapien D, Ruiz FX, Poirson J,
Mitschler A, Ramirez J, Forster A, Cousido-Siah A, Masson M, Vande
Pol S, Podjarny A, et al: Structure of the E6/E6AP/p53 complex
required for HPV-mediated degradation of p53. Nature. 529:541–545.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Scheffner M, Huibregtse JM, Vierstra RD
and Howley PM: The HPV-16 E6 and E6-AP complex functions as an
ubiquitin-protein ligase in the ubiquitination of p53. Cell.
75:495–505. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Huibregtse JM, Scheffner M and Howley PM:
A cellular protein mediates association of p53 with the E6
oncoprotein of human papillomavirus types 16 or 18. EMBO J.
10:4129–4135. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hurlin PJ, Kaur P, Smith PP, Perez-Reyes
N, Blanton RA and McDougall JK: Progression of human papillomavirus
type 18-immortalized human keratinocytes to a malignant phenotype.
Proc Natl Acad Sci USA. 88:570–574. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chen J: The cell-cycle arrest and
apoptotic functions of p53 in tumor initiation and progression.
Cold Spring Harb Perspect Med. 6:a0261042016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Gulati N, Laudet B, Zohrabian VM, Murali R
and Jhanwar-Uniyal M: The antiproliferative effect of quercetin in
cancer cells is mediated via inhibition of the PI3K-Akt/PKB
pathway. Anticancer Res. 26:1177–1181. 2006.PubMed/NCBI
|
|
12
|
Metodiewa D, Jaiswal AK, Cenas N,
Dickancaité E and Segura-Aguilar J: Quercetin may act as a
cytotoxic prooxidant after its metabolic activation to semiquinone
and quinoidal product. Free Radic Biol Med. 26:107–116. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Srivastava S, Somasagara RR, Hegde M,
Nishana M, Tadi SK, Srivastava M, Choudhary B and Raghavan SC:
Quercetin, a natural flavonoid interacts with DNA, arrests cell
cycle and causes tumor regression by activating mitochondrial
pathway of apoptosis. Sci Rep. 6:240492016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Qiao Y, Cao J, Xie L and Shi X: Cell
growth inhibition and gene expression regulation by
(−)-epigallocatechin-3-gallate in human cervical cancer cells. Arch
Pharm Res. 32:1309–1315. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Cherry JJ, Rietz A, Malinkevich A, Liu Y,
Xie M, Bartolowits M, Davisson VJ, Baleja J and Androphy EJ:
Structure based identification and characterization of flavonoids
that disrupt human papillomavirus-16 E6 function. PLoS One.
8:e845062013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tanigawa S, Fujii M and Hou DX:
Stabilization of p53 is involved in quercetin-induced cell cycle
arrest and apoptosis in HepG2 cells. Biosci Biotechnol Biochem.
72:797–804. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Thangasamy T, Sittadjody S, Lanza-Jacoby
S, Wachsberger PR, Limesand KH and Burd R: Quercetin selectively
inhibits bioreduction and enhances apoptosis in melanoma cells that
overexpress tyrosinase. Nutr Cancer. 59:258–268. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yang GY, Liao J, Kim K, Yurkow EJ and Yang
CS: Inhibition of growth and induction of apoptosis in human cancer
cell lines by tea polyphenols. Carcinogenesis. 19:611–616. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Mohan S, Bustamam A, Ibrahim S, Al-Zubairi
AS, Aspollah M, Abdullah R and Elhassan MM: In vitro
ultramorphological assessment of apoptosis on CEMss induced by
linoleic acid-rich fraction from Typhonium flagelliforme tuber.
Evid Based Complement Alternat Med 2011. 4218942011.
|
|
20
|
Zanier K, Charbonnier S, Sidi AO, McEwen
AG, Ferrario MG, Poussin-Courmontagne P, Cura V, Brimer N, Babah
KO, Ansari T, et al: Structural basis for hijacking of cellular
LxxLL motifs by papillomavirus E6 oncoproteins. Science.
339:694–698. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Jordan MA, Toso RJ, Thrower D and Wilson
L: Mechanism of mitotic block and inhibition of cell proliferation
by taxol at low concentrations. Proc Natl Acad Sci USA.
90:9552–9556. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hengstermann A, Linares LK, Ciechanover A,
Whitaker NJ and Scheffner M: Complete switch from Mdm2 to human
papillomavirus E6-mediated degradation of p53 in cervical cancer
cells. Proc Natl Acad Sci USA. 98:1218–1223. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Van Kriekinge G, Castellsagué X, Cibula D
and Demarteau N: Estimation of the potential overall impact of
human papillomavirus vaccination on cervical cancer cases and
deaths. Vaccine. 32:733–739. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Peirson L, Fitzpatrick-Lewis D, Ciliska D
and Warren R: Screening for cervical cancer: A systematic review
and meta-analysis. Syst Rev. 2:352013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wätjen W, Michels G, Steffan B, Niering P,
Chovolou Y, Kampkötter A, Tran-Thi QH, Proksch P and Kahl R: Low
concentrations of flavonoids are protective in rat H4IIE cells
whereas high concentrations cause DNA damage and apoptosis. J Nutr.
135:525–531. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Laptenko O, Beckerman R, Freulich E and
Prives C: p53 binding to nucleosomes within the p21 promoter in
vivo leads to nucleosome loss and transcriptional activation. Proc
Natl Acad Sci USA. 108:10385–10390. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Charrier-Savournin FB, Château MT, Gire V,
Sedivy J, Piette J and Dulic V: p21-mediated nuclear retention of
cyclin B1-Cdk1 in response to genotoxic stress. Moll Biol Cell.
15:3965–3976. 2004. View Article : Google Scholar
|
|
28
|
Miyashita T, Krajewski S, Krajewska M,
Wang HG, Lin HK, Liebermann DA, Hoffman B and Reed JC: Tumor
suppressor p53 is a regulator of bcl-2 and bax gene expression in
vitro and in vivo. Oncogene. 9:1799–1805. 1994.PubMed/NCBI
|
|
29
|
Sax JK, Fei P, Murphy ME, Bernhard E,
Korsmeyer SJ and El-Deiry WS: BID regulation by p53 contributes to
chemosensitivity. Nat Cell Biol. 4:842–849. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Nakano K and Vousden KH: PUMA, a novel
proapoptotic gene, is induced by p53. Mol Cell. 7:683–694. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Erster S, Mihara M, Kim RH, Petrenko O and
Moll UM: In vivo mitochondrial p53 translocation triggers a rapid
first wave of cell death in response to DNA damage that can precede
p53 target gene activation. J Biol Chem. 24:6728–6741. 2004.
|
|
32
|
Tomita Y, Marchenko N, Erster S,
Nemajerova A, Dehner A, Klein C, Pan H, Kessler H, Pancoska P and
Moll UM: WT p53, but not tumor-derived mutants, bind to Bcl2 via
the DNA binding domain and induce mitochondrial permeabilization. J
Biol Chem. 281:8600–8606. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Vidya Priyadarsini R, Senthil Murugan R,
Maitreyi S, Ramalingam K, Karunagaran D and Nagini S: The flavonoid
quercetin induces cell cycle arrest and mitochondria-mediated
apoptosis in human cervical cancer (HeLa) cells through p53
induction and NF-κB inhibition. Eur J Pharmacol. 649:84–91. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Malecka KA, Fera D, Schultz DC,
Hodawadekar S, Reichman M, Donover PS, Murphy ME and Marmorstein R:
Identification and characterization of small molecule human
papillomavirus E6 inhibitors. ACS Chem Biol. 9:1603–1612. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Mietz JA, Unger T, Huibregtse JM and
Howley PM: The transcriptional transactivation function of
wild-type p53 is inhibited by SV40 large T-antigen and by HPV-16 E6
oncoprotein. EMBO J. 11:5013–5020. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Cooper B, Schneider S, Bohl J, Jiang Yh,
Beaudet A and Vande Pol S: Requirement of E6AP and the features of
human papillomavirus E6 necessary to support degradation of p53.
Virology. 306:87–99. 2003. View Article : Google Scholar : PubMed/NCBI
|