Introduction
Gastric cancer (GC) is one of the most common causes
of cancer-associated mortality worldwide and is a major health
problem in China with an increasing incidence and mortality rate
according to a survey in 2015 (1,2).
Although advances have been made in the detection and clinical
treatment of GC in previous decades, marked variation in survival
rates are found between patients diagnosed at different tumor
stages. Patients diagnosed during the later stage often have
metastatic disease and are no longer able to receive surgical
treatment, leaving chemotherapy as the only available option
(3). However, chemotherapy is
often associated with a low response rate, high toxicity and drug
resistance in a large number of patients (4–6).
Therefore, there is an urgency for identifying and developing novel
compounds to optimize therapeutic options and improve the prognosis
of patients with GC.
Signal transducer and activator of transcription 3
(STAT3) is the most studied member of the STAT family and is
constitutively activated in various malignancies, including GC
(7). Under normal physiological
conditions, cells exhibit transient STAT3 phosphorylation, which
lasts for only a relatively short period of time; however, once the
tumor-related signaling pathways are dysregulated, this process
becomes constitutive (8).
Following activation, STAT3 undergoes phosphorylation-induced
dimerization, nuclear translocation and binding to its target
genes, leading to the transcriptional activation of downstream
target genes that regulate tumor cell proliferation and progression
(3,9). There is substantial evidence
demonstrating that the phosphorylation of STAT3 is abnormally
activated in GC and that the constitutive activation of STAT3 is
positively correlated with a poor prognosis and metastasis,
indicating that it may serve as a negative prognostic factor
(7,10–12).
As a consequence, inhibitors targeting the activation of STAT3
offer promise in suppressing cancer proliferation and mobility and
are being widely investigated in GC and several other types of
cancer that contain activated STAT3 (13,14).
However, the majority of STAT3 inhibitors fail to demonstrate
satisfactory ability to suppress tumor growth and/or have high
toxicity (15). Therefore, it
remains important to identify novel inhibitors of STAT3 activation
that are effective in treating GC, while producing minimal side
effects.
In addition, the dysregulation of mitogen-activated
protein kinase (MAPK) signaling is closely associated with cell
growth, progression and apoptosis. The three most widely studied
MAPKs are extracellular signal-related kinases (ERKs), p38 MAPKs
and c-Jun NH2-terminal kinases (JNKs) (16). Increasing evidence suggests that
the activation of ERK, p38 and JNK MAPK signaling is involved in
cancer initiation and progression, indicating that they may be
promising therapeutic targets (17,18).
BP-1-102 was designed as a STAT3 inhibitor and is an
analog of S3I-201, which functions by disrupting STAT3
homodimerization (19). BP-1-102
inhibits the STAT3 Src homology 2 (SH2) domain, binds STAT3 with an
affinity of 504 nM, and disrupts STAT3:STAT3 complex formation with
a half maximal inhibitory concentration (IC50) of 6.8
µM, which is a notable improvement from that of S3I-201
(IC50=86 µM); however, BP-1-102 has minimal or no effect
on other STATs, including STAT1 and STAT5 (20,21).
The antitumor effect of BP-1-102 has been evaluated in human
pancreatic, breast, prostate, liver and lung cancer in
vitro, and in human breast and non-small cell lung tumor
xenografts in vivo through either tail vein or oral
administration (20,21); however, its role in GC has not been
reported. Therefore, in the present study, in vitro
experiments were conducted to investigate whether BP-1-102 exerts
an antitumor effect on GC cells with constitutively activated STAT3
and to examine the molecular mechanisms involved.
Materials and methods
Cell lines and culture conditions
Five human gastric cancer cell lines (AGS, HGC-27,
MKN28, MGC803 and SGC7901) were obtained from the Institute of
Cellular Biology (Chinese Academy of Science, Shanghai, China) and
cultured in RPMI-1640 medium (Gibco; Thermo Fisher Scientific,
Inc., Waltham, MA, USA) supplemented with 10% fetal bovine serum
(FBS; Gibco; Thermo Fisher Scientific, Inc.) and 1%
penicillin/streptomycin (Thermo Fisher Scientific, Inc.) in a
humidified 37°C incubator with 5% CO2.
Reagents and antibodies
The novel STAT3 inhibitor BP-1-102 was obtained from
Selleck Chemicals, LLC (Houston, TX, USA) and dissolved in sterile
dimethyl sulfoxide (DMSO; Sigma-Aldrich; Merck KGaA, Darmstadt,
Germany) and stored at −20°C. The primary antibodies for STAT3
(cat. no. ab68153, monoclonal, raised in rabbit, 1:2,000),
phosphorylated (p-)STAT3 (Y705; cat. no. ab76315, monoclonal,
raised in rabbit, 1:5,000), JNK (cat. no. ab208035, monoclonal,
raised in rabbit, 1:1,000), p38 MAPK (cat. no. ab170099,
monoclonal, raised in rabbit, 1:1,000), p-JNK (Y185/Y185/Y223; cat.
no. ab76572, monoclonal, raised in rabbit, 1:5,000) and p-p38 MAPK
(T180/Y182; cat. no. ab195049, monoclonal, raised in rabbit,
1:1,000) were purchased from Abcam (Cambridge, UK). The antibodies
against p44/42 MAPK (ERK1/2; cat. no. 4695, monoclonal, raised in
rabbit, 1:1,000), p-p44/42 MAPK (p-ERK1/2, T202/Y204; cat. no.
4377, monoclonal, raised in rabbit, 1:1,000), c-Myc (cat. no. 9402,
polyclonal, raised in rabbit, 1:1,000), cyclin D1 (cat. no. 2922,
polyclonal, raised in rabbit, 1:1,000), survivin (cat. no. 2803,
polyclonal, raised in rabbit, 1:1,000), cleaved-PARP (c-PARP, cat.
no. 5625, polyclonal, raised in rabbit, 1:1,000), cleaved-caspase 3
(c-caspase 3, cat. no. 9661, polyclonal, raised in rabbit, 1:1,000)
and BIM (cat. no. 2933, polyclonal, raised in rabbit, 1:1,000) were
purchased from Cell Signaling Technology, Inc. (Beverly, MA, USA).
GAPDH (cat. no. HRP-60004, 1:1,000) and HRP-conjugated secondary
antibodies (cat. no. SA00001-2, 1:5,000) were purchased from
ProteinTech Group, Inc. (Wuhan, China).
Cell viability assay
The AGS (3×103 cells/well) and HGC-27
(2×103 cells/well) cells were seeded into 96-well
plates, exposed to DMSO vehicle (1 µM) or various concentrations of
BP-1-102 (2, 4 and 6 µM in 1 µM DMSO). The maximum final
concentration of DMSO was ≤0.1% in the cell culture medium.
Following incubation for 24, 48 and 72 h at 37°C, a Cell Counting
Kit-8 (CCK8; Dojindo Molecular Technologies, Inc., Kumamato, Japan)
was used to assess cell viability following the manufacturer's
protocol, and the absorbance at a wavelength of 450 nm was measured
using a microplate enzyme-linked immunosorbent assay reader
(Bio-Rad Laboratories, Inc., Hercules, CA, USA).
Colony formation assay
The AGS (5×102 cells/well) and HGC-27
(8×102 cells/well) cells were seeded in 6-well culture
plates, treated with different concentrations of BP-1-102 (2, 4 and
6 µM in 1 µM DMSO) or DMSO vehicle (1 µM). Following culture for
~14 days, the colonies were fixed with 95% ethanol, stained with
0.1% crystal violet for 30 min and washed with phosphate-buffered
saline, following which colony numbers were counted using an
inverted microscope (magnification, ×200; Zeiss GmbH, Jena,
Germany).
Flow cytometry
Apoptosis was evaluated using the Annexin
V-fluorescein isothiocyanate (FITC)/propidium iodide (PI) Detection
kit (BD Biosciences, San Jose, CA, USA). The AGS cells
(2×105 cells/well) were seeded into 6-well plates and
incubated overnight, following which the cells were treated with
the different concentrations of BP-1-102 for 8 h. The cells were
then harvested and resuspended in 500 µl of 1X binding buffer
solution, incubated with Annexin V-FITC (5 µl) and PI (5 µl) at 4°C
for 15 min. Subsequently, the samples were analyzed within 1 h by
flow cytometry (BD Biosciences) and BD CellQuest Pro software
(version 2.0, BD Pharmingen; BD Biosciences).
For cell cycle analysis, the BP-1-102-pretreated
cells were trypsinized, fixed in 75% ethanol, incubated at 4°C
overnight, and then centrifuged at 800 × g for 5 min at room
temperature. The cells were then re-suspended in PI and RNase A
solution for 30 min at room temperature in the dark and evaluated
using flow cytometry (BD Biosciences) and BD CellQuest Pro software
(BD Pharmingen; BD Biosciences) within 1 h.
Transwell assay for migration and
invasion
For the migration and invasion assays, the cells
were pre-exposed to BP-1-102 (6 µM in 1 µM DMSO) or DMSO vehicle (1
µM) for 8 h and 24-well Transwell® plates with 8-µm pore
polycarbonate filters (Costar; Corning Incorporated, Corning, NY,
USA) were used. The cells were harvested in serum-free RPMI-1640
medium at a density of 5×104 cells in 200 µl and seeded
into the upper chambers, which had either been coated with Matrigel
(BD Biosciences) or left uncoated. Subsequently, 700 µl of
RPMI-1640 complete medium was added to the lower chambers.
Following incubation for 18 h, the cells that had penetrated the
membrane into the lower chamber were fixed with 95% ethanol and
stained with 0.1% crystal violet. Images were then captured with a
light microscope under ×200 magnification.
Western blot analysis
The cells (5×105 cells/well) were
cultured in six-well plates. Following treatment with BP-1-102 (6
µM) for various durations (0, 3, 5 and 8 h) or at various
concentrations of BP-1-102 (0, 2, 4 and 6 µM) for 8 h, the cells
were harvested and quantified as previously described (22). Samples containing 20 µg of protein
were subjected to electrophoresis on 10 or 12% SDS-PAGE gels and
transferred onto a polyvinylidene difluoride membrane (EMD
Millipore, Billerica, MA, USA). The membranes were blocked using
blocking buffer (10% skimmed milk in Tris-buffered saline
containing 0.1% Tween-20) for 1 h, incubated with primary
antibodies overnight at 4°C, and then incubated with anti-rabbit
HRP-conjugated secondary antibodies. Finally, the immunoreactive
protein bands were visualized using an enhanced chemiluminescence
western blotting kit (Bio-Rad Laboratories, Inc.) following the
manufacturer's protocol.
Statistical analysis
Experimental data from experiments conducted in
triplicate are presented as the mean ± standard deviation.
Statistical analyses between two groups were performed using
Student's t-test, and multiple comparisons were made using one-way
analysis of variance followed by Dunnett's test (GraphPad Prism
5.01; GraphPad Software, Inc., San Diego, CA, USA). P<0.05 was
considered to indicate a statistically significant difference.
Results
BP-1-102 inhibits the growth of AGS
cells
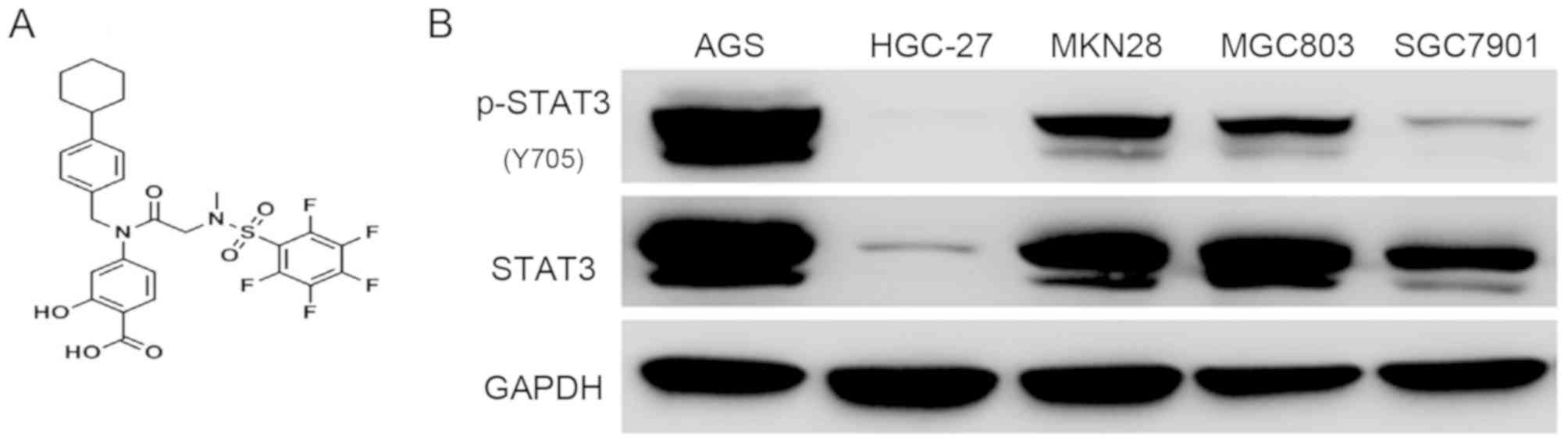
The chemical structure of BP-1-102 is shown in
Fig. 1A. To evaluate the antitumor
effect of BP-1-102, five GC cell-lines (AGS, HGC-27, MKN28, MGC803
and SGC7901) were used to examine the expression status and the
phosphorylation level of STAT3. The AGS cells exhibited a high
level of STAT3 Y705 phosphorylation, which is the most studied
activated-form of STAT3, whereas the HGC-27 cells exhibited the
lowest expression profile (Fig.
1B). Based on these results, the AGS and HGC-27 cell lines were
selected for subsequent experiments. The AGS and HGC-27 cells were
exposed to various concentrations of BP-1-102 to evaluate its
effect on the proliferation of GC cells using a CCK8 assay.
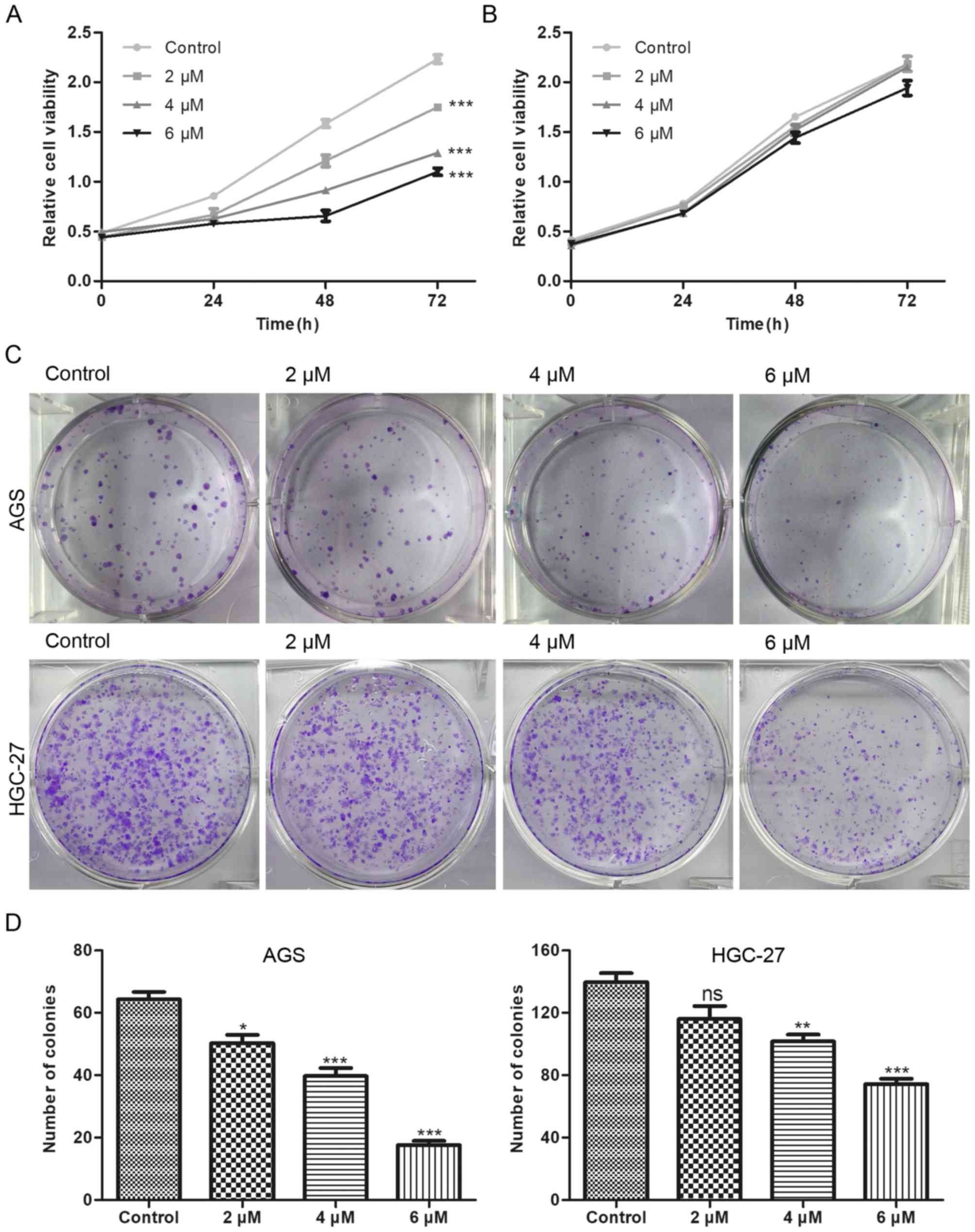
Compared with the control group, BP-1-102 treatment
dose-dependently suppressed the proliferation of AGS cells
(Fig. 2A) but had no such
inhibitory effect on HGC-27 cells (Fig. 2B). Furthermore, the results of the
colony formation assays showed that the BP-1-102-treated AGS cells
formed smaller and fewer colonies compared with those in the
control group. BP-1-102 was less effective towards HGC-27 cells
than AGS cells (Fig. 2C and D).
These results indicated that BP-1-102 exerted a tumor suppressive
role in GC cells lines and that this effect was enhanced by high
expression levels of p-STAT3 (Y705). Therefore, the AGS cell line
was selected for subsequent experiments.
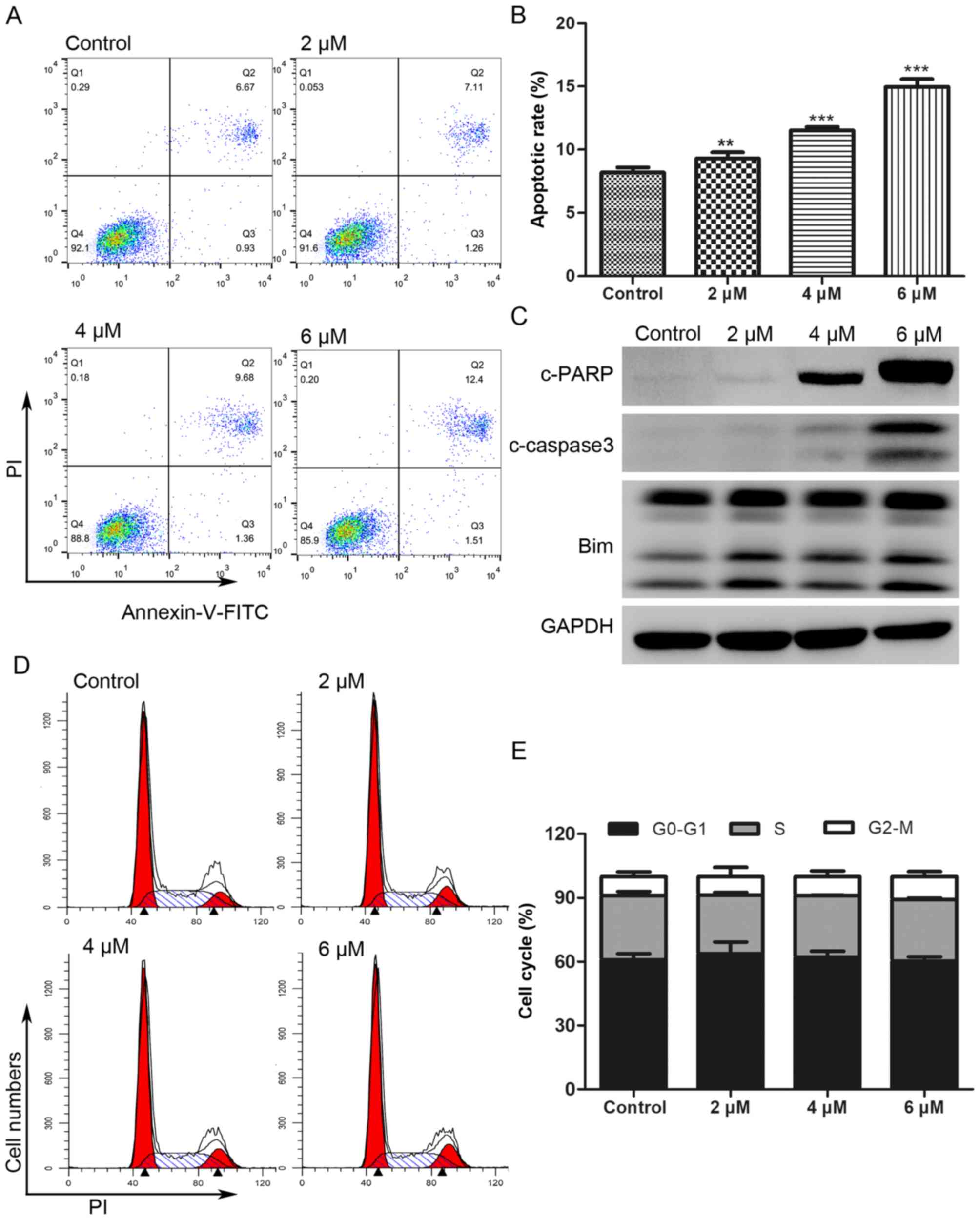
BP-1-102 induces apoptosis in AGS
cells and has little effect on cell cycle
The Annexin V-FITC/PI double staining assay was
performed to investigate whether the antitumor effect of BP-1-102
was associated with the induction of apoptosis. As shown in
Fig. 3A and B, the proportion of
apoptotic AGS cells (Annexin V-FITC positive) increased from 7.6%
in the untreated control cells to 8.37, 11.04 and 13.91% following
treatment for 8 h with 2, 4 and 6 µM of BP-1-102, respectively. The
expression of apoptotic-related proteins was analyzed by western
blotting. As shown in Fig. 3C,
BP-1-102 treatment markedly increased the expression of
cleaved-poly (ADP-ribose) polymerase (PARP), cleaved-caspase 3, and
B-cell lymphoma 2 (Bcl-2)-interacting mediator of cell death (Bim)
in a dose-dependent manner. BP-1-102 treatment had no significant
influence on the AGS cell cycle distribution (Fig. 3D and E).
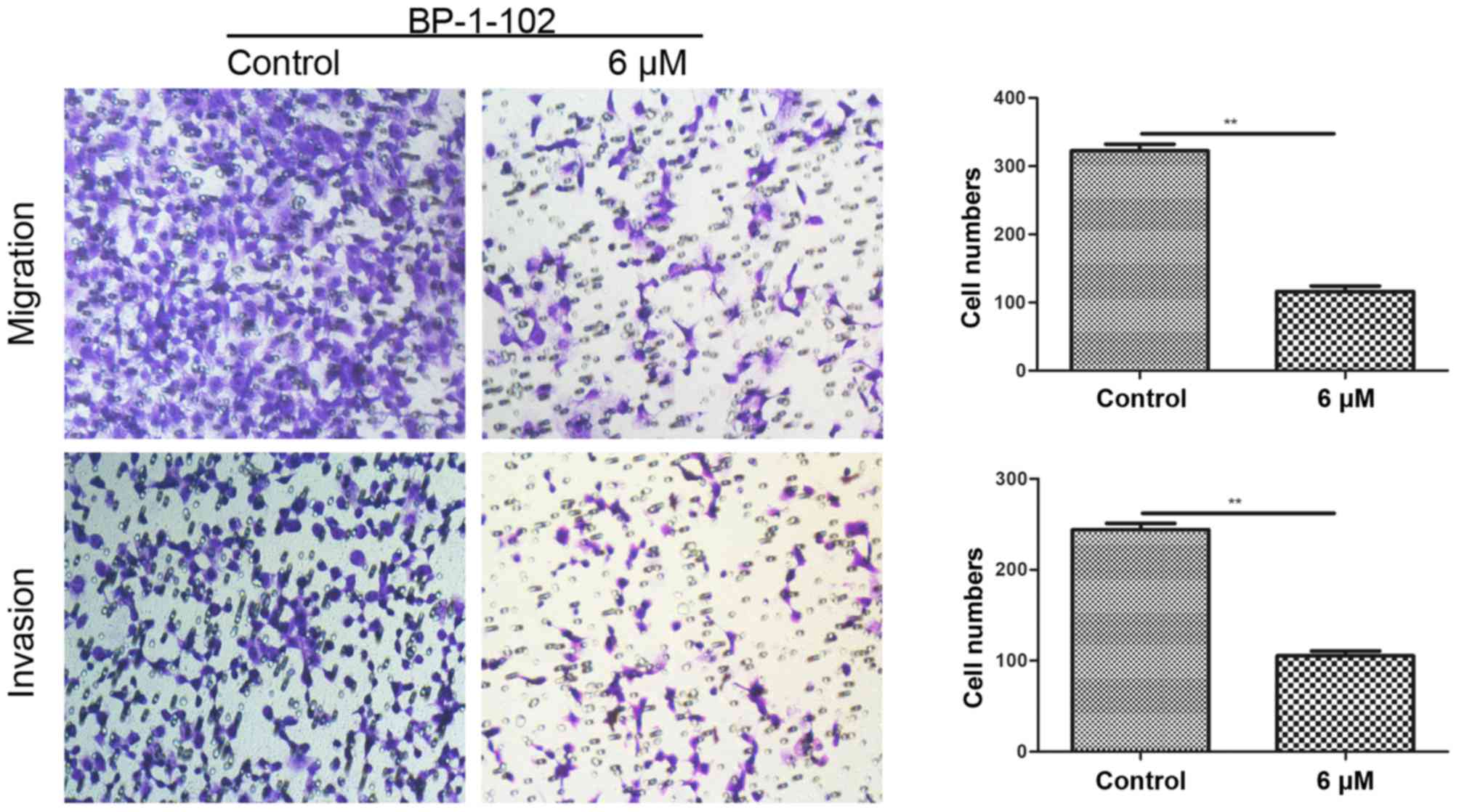
BP-1-102 inhibits the mobility of AGS
cells
Transwell assays were conducted to assess the effect
of BP-1-102 on tumor cell migration and invasion. As shown in
Fig. 4, the number of penetrating
cells was significantly lower in the BP-1-102-treated group (6 µM)
compared with that in the control group (untreated). These data
indicate the involvement of BP-1-102 in suppressing the mobility of
AGS cells.
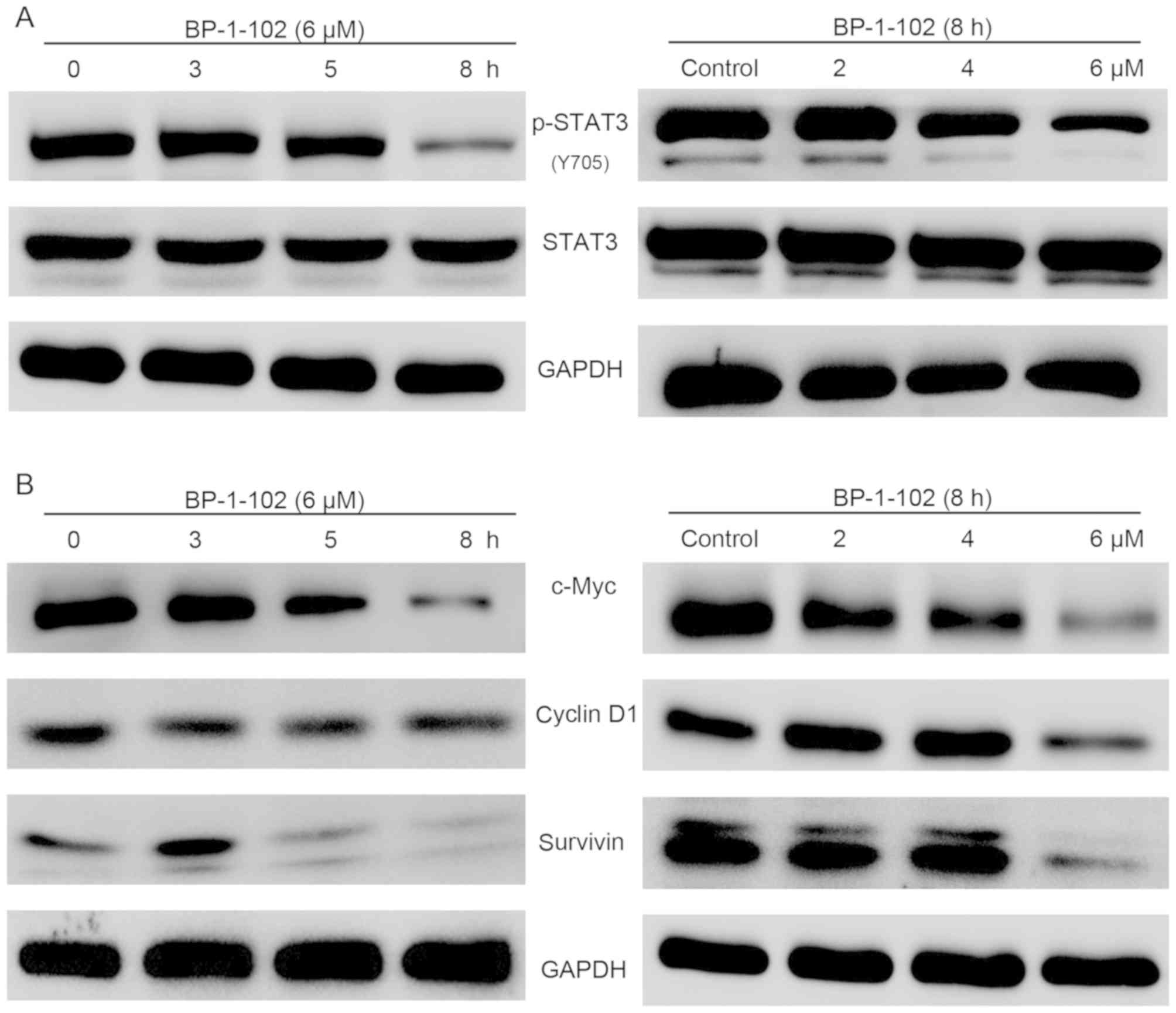
BP-1-102 inhibits the phosphorylation
of STAT3 and its target genes in a time-dependent and
dose-dependent manner
The previous findings demonstrated that BP-1-102
inhibited cell growth, migration and invasion ability and induced
apoptosis. To gain insight into the potential mechanism of BP-1-102
activity in AGS cells, the activation of STAT3 and its target genes
were examined using western blot analysis. As shown in Fig. 5A, the expression of p-STAT3 (Y705)
was markedly suppressed by BP-1-102 treatment in a time- and
dose-dependent manner, whereas the expression levels of total STAT3
remained unchanged. The STAT3 signaling pathway regulates cell
proliferation and survival by modulating the expression of various
target genes. The results showed that BP-1-102 also time- and
dose-dependently downregulated the expression of c-Myc, cyclin D1
and survivin (Fig. 5B).
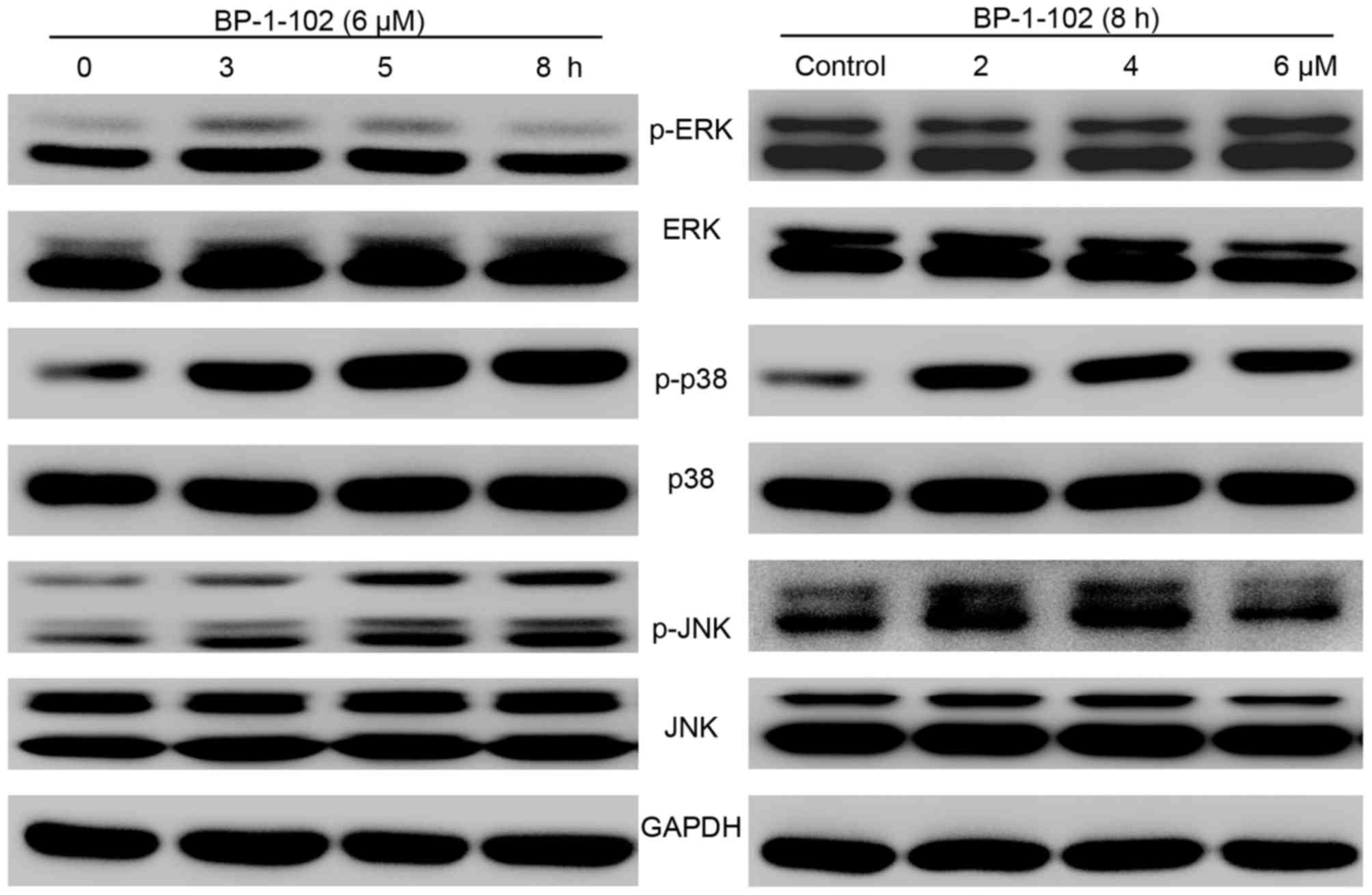
BP-1-102 modulates the expression of
MAPKs
As BP-1-102 inhibited cell growth and mobility and
induced apoptosis, the possible mechanisms involved were
investigated by evaluating the effect of BP-1-102 treatment on the
elementary activation status of MAPKs. As shown in Fig. 6, following exposure of the AGS
cells to BP-1-102, the phosphorylation of p38 and JNK was
upregulated, whereas the phosphorylation of ERK was decreased, in a
time- and dose-dependent manner relative to the total levels of
p38, JNK and ERK, which served as internal controls.
Discussion
STAT3 is pivotal in signal transduction and
transcription, and regulates the transcriptional activity of
downstream target genes involved in cell proliferation (c-Myc and
cyclin D1), anti-apoptotic genes (Bcl-2, Bcl-extra large and
survivin) and genes involved in invasion (matrix
metalloproteinase-2 and −9) (23,24).
Although the basal expression levels differ in tissues from
different organs, STAT3 is frequently overexpressed or
constitutively activated in human solid tumors compared with
matched normal tissues (25).
Previously, constitutively activated STAT3 was reported to occur in
various types of cancer, including prostate, renal, breast, ovarian
and liver cancer, and is correlated with poor overall survival
rates of patients with lung cancer or GC (26–28).
In particular, a previous study of GC found that the overexpression
levels of p-STAT3 and total STAT3 were correlated with poor overall
survival rate (29). Therefore,
although the role of STAT3 in tumorigenesis remains to be fully
elucidated, the inhibition of STAT3 continues to be an attractive
strategy for the clinical treatment of tumors with activated
STAT3.
Several strategies have been established for the
inhibition of STAT3, including the inhibition of upstream
activators, inhibiting the phosphorylation of STAT3 itself, and
suppressing the nuclear translocation of STAT3; notably a large
number of inhibitors targeting STAT3 have been identified over the
last few decades (14,30). Specifically, several non-peptide
STAT3-SH2 domain inhibitors, including S3I-201, STATTIC and STA-21,
have been found to be effective in suppressing cell growth in
vitro and in vivo (31–33).
S3I-201 (NSC 74859) was effective in inhibiting the formation of
STAT3 dimers (IC50=86 µM) and the expression of STAT3
target genes (c-Myc, cyclin D1 and survivin) in breast cancer
(33). In liver cancer, it was
found that S3I-201 was not only able to inhibit tumor growth but
also significantly enhanced the antitumor effects of cetuximab and
doxorubicin (34–36). STATTIC can selectively inhibit
STAT3 dimerization (IC50=5.1 µM) and nuclear
translocation in breast and liver cancer (31). In addition, it has been found that
STATTIC enhances chemo- and radio-sensitivity in patients with head
and neck squamous cell carcinoma (37,38).
STA-21 was found to reduce the survival of breast carcinoma cells
with hyperactivated STAT3; however, it exerted little effect on
cells with a low expression of activated STAT3 (32). STA-21 has been also reported to be
involved in regulating cell differentiation and inflammation
(39–41). However, although present inhibitors
have an acceptable antitumor effect, they are neither effective at
low concentrations nor orally bioavailable. For these reasons,
although substantial progress has been made, further progress is
necessary, and novel inhibitors with higher efficiency, improved
bioavailability and fewer side effects are required. BP-1-102 is a
novel orally bioavailable STAT3 inhibitor that functions by
directly interacting with STAT3 at a relatively low concentration
(IC50=6.4 µM) and has been shown to have significant
antitumor effects in several tumors (19); however, its potential use in GC
treatment has not been investigated previously.
In the present study, cell growth, colony formation,
apoptosis, cell cycle and cell mobility were evaluated in
vitro to determine the effect of BP-1-102 on the biological
characteristics of AGS cells. It was found for the first time, to
the best of our knowledge, that BP-1-102 exhibited superior
suppressive effects on the cell proliferation and colony formation
capacities of AGS cells with a higher expression level of p-STAT3
(Y705) than HGC-27 cells with lower expression levels of p-STAT3
(Y705). In addition, the expression status of STAT3 and several
STAT3-regulated genes were examined. The results revealed that the
constitutive activation of STAT3 was inhibited by BP-1-102 in a
time- and dose-dependent manner. Subsequently, the expression
levels of c-Myc and cyclin D1, two proteins associated with the
inhibition of cell proliferation, decreased when the cells were
exposed to BP-1-102. Apoptosis assays demonstrated that the
proportion of apoptotic cells increased upon incubation of the
cells with BP-1-102, which can be attributed to suppression of the
anti-apoptotic protein surviving (42). Additionally, several
apoptotic-related proteins were measured via western blotting. When
exposed to BP-1-102, the expression of cleaved-PARP,
cleaved-caspase 3 and pro-apoptotic Bim were markedly increased in
the AGS cells in a dose-dependent manner. However, although a
significant decrease in the expression of cyclin D1, an important
target in regulating cell cycle (43), was observed, the cell cycle
distribution of AGS cells remained unchanged when pretreated with
BP-1-102 for 8 h. In addition, based on the results of the
Transwell assays, the migration and invasion capacities of AGS
cells were markedly suppressed by BP-1-102 treatment. The detailed
mechanisms underlying these effects of BP-1-102 on AGS cells remain
to be elucidated.
The dysregulation of MAPK is critical in tumor
development (18,44). ERK, the most widely studied MAPK,
has been shown to be a major regulator in cell growth and the
activation of p38 and JNK MAPKs are reported to be positively
correlated with apoptosis (16,45,46).
In the present study, BP-1-102 was found to activate JNK and p38
MAPK and to inactivate ERK in AGS cells in a time- and
dose-dependent manner. The results suggest that BP-1-102 may also
act as a MAPK regulator in AGS cells. However, future
investigations are warranted to further examine the roles of MAPKs
in the effects of BP-1-102 on growth suppression and apoptosis
induction in AGS cells. There were several limitations to the
present study. For example, the mechanism was based on a single
cell line and in vivo experiments are required to further
evaluate the antitumor effect and toxicity of BP-1-102. Further
experiments, including in vivo experiments are to be
performed in the future to validate its antitumor potential in
GC.
In conclusion, the present study is the first, to
the best of our knowledge, to demonstrate that BP-1-102 is a potent
inhibitor of STAT3 signaling and a potential regulator of MAPKs in
GC cells. BP-1-102 suppressed the proliferation, migration and
invasion capacities of AGS human gastric cancer cells and induced
their apoptosis. These findings suggest that BP-1-102 may be a
potential therapeutic agent. Future in vivo experiments are
warranted to further evaluate this possible approach, and to
determine the efficacy and safety of BP-1-102 in treating patients
with GC with hyperactivated STAT3.
Acknowledgements
Not applicable.
Funding
This study was supported by grants from the National
Natural Science Foundation of China (grant nos. 81201089 and
81272676) and the Natural Science Foundation of Zhejiang province,
China (grant nos. LY15H160026 and LY15H160012).
Availability of data and materials
The data used and/or analyzed in the present study
are available from the corresponding author on reasonable
request.
Authors' contributions
XXJ, JT, ZQL, XFY and LST conceived and designed the
study. XXJ, JT, MJW, STC, ZZX, YHW and HHW performed the data
analysis. XXJ and JT wrote the paper. All the authors reviewed and
edited the manuscript. All authors read and approved the
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
Statistics, 2017. CA Cancer J Clin. 67:7–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chen W, Zheng R, Baade PD, Zhang S, Zeng
H, Bray F, Jemal A, Yu XQ and He J: Cancer statistics in China,
2015. CA Cancer J Clin. 66:115–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Jiang C, Chen X, Alattar M, Wei J and Liu
H: MicroRNAs in tumorigenesis, metastasis, diagnosis and prognosis
of gastric cancer. Cancer Gene Ther. 22:291–301. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ajani JA: Chemotherapy for gastric
carcinoma: New and old options. Oncology (Williston Park).
12:44–47. 1998.PubMed/NCBI
|
|
5
|
Tannock IF: Cancer: Resistance through
repopulation. Nature. 517:152–153. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kessler DA, Austin RH and Levine H:
Resistance to chemotherapy: Patient variability and cellular
heterogeneity. Cancer Res. 74:4663–4670. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Judd LM, Bredin K, Kalantzis A, Jenkins
BJ, Ernst M and Giraud AS: STAT3 activation regulates growth,
inflammation, and vascularization in a mouse model of gastric
tumorigenesis. Gastroenterology. 131:1073–1085. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bromberg JF, Wrzeszczynska MH, Devgan G,
Zhao Y, Pestell RG, Albanese C and Darnell JE Jr: Stat3 as an
oncogene. Cell. 98:295–303. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Li Y, Rogoff HA, Keates S, Gao Y,
Murikipudi S, Mikule K, Leggett D, Li W, Pardee AB and Li CJ:
Suppression of cancer relapse and metastasis by inhibiting cancer
stemness. Proc Natl Acad Sci USA. 112:1839–1844. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hajimoradi M, Mohammad Hassan Z, Ebrahimi
M, Soleimani M, Bakhshi M, Firouzi J and Samani FS: STAT3 is
overactivated in gastric cancer stem-like cells. Cell J.
17:617–628. 2016.PubMed/NCBI
|
|
11
|
Deng JY, Sun D, Liu XY, Pan Y and Liang H:
STAT-3 correlates with lymph node metastasis and cell survival in
gastric cancer. World J Gastroenterol. 16:5380–5387. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gong W, Wang L, Yao JC, Ajani JA, Wei D,
Aldape KD, Xie K, Sawaya R and Huang S: Expression of activated
signal transducer and activator of transcription 3 predicts
expression of vascular endothelial growth factor in and angiogenic
phenotype of human gastric cancer. Clin Cancer Res. 11:1386–1393.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Cafferkey C and Chau I: Novel STAT 3
inhibitors for treating gastric cancer. Expert Opin Investig Drugs.
25:1023–1031. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Furtek SL, Backos DS, Matheson CJ and
Reigan P: Strategies and approaches of targeting STAT3 for cancer
treatment. ACS Chem Biol. 11:308–318. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lai PS, Rosa DA, Magdy Ali A, Gómez-Biagi
RF, Ball DP, Shouksmith AE and Gunning PT: A STAT inhibitor patent
review: Progress since 2011. Expert Opin Ther Pat. 25:1397–1421.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Boutros T, Chevet E and Metrakos P:
Mitogen-activated protein (MAP) kinase/MAP kinase phosphatase
regulation: Roles in cell growth, death, and cancer. Pharmacol Rev.
60:261–310. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Meng X and Zhang S: MAPK cascades in plant
disease resistance signaling. Annu Rev Phytopathol. 51:245–266.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kim EK and Choi EJ: Compromised MAPK
signaling in human diseases: An update. Arch Toxicol. 89:867–882.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Page BD, Fletcher S, Yue P, Li Z, Zhang X,
Sharmeen S, Datti A, Wrana JL, Trudel S, Schimmer AD, et al:
Identification of a non-phosphorylated, cell permeable, small
molecule ligand for the Stat3 SH2 domain. Bioorg Med Chem Lett.
21:5605–5609. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang X, Yue P, Page BD, Li T, Zhao W,
Namanja AT, Paladino D, Zhao J, Chen Y, Gunning PT and Turkson J:
Orally bioavailable small-molecule inhibitor of transcription
factor Stat3 regresses human breast and lung cancer xenografts.
Proc Natl Acad Sci USA. 109:9623–9628. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Resetca D, Haftchenary S, Gunning PT and
Wilson DJ: Changes in signal transducer and activator of
transcription 3 (STAT3) dynamics induced by complexation with
pharmacological inhibitors of Src homology 2 (SH2) domain
dimerization. J Biol Chem. 289:32538–32547. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zheng L, Xu C, Guan Z, Su X, Xu Z, Cao J
and Teng L: Galectin-1 mediates TGF-β-induced transformation from
normal fibroblasts into carcinoma-associated fibroblasts and
promotes tumor progression in gastric cancer. Am J Transl Res.
8:1641–1658. 2016.PubMed/NCBI
|
|
23
|
Luwor RB, Stylli SS and Kaye AH: The role
of Stat3 in glioblastoma multiforme. J Clin Neurosci. 20:907–911.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang X, Yue P, Fletcher S, Zhao W,
Gunning PT and Turkson J: A novel small-molecule disrupts Stat3 SH2
domain-phosphotyrosine interactions and Stat3-dependent tumor
processes. Biochem Pharmacol. 79:1398–1409. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhuang S: Regulation of STAT signaling by
acetylation. Cell Signal. 25:1924–1931. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Tong M, Wang J, Jiang N, Pan H and Li D:
Correlation between p-STAT3 overexpression and prognosis in lung
cancer: A systematic review and meta-analysis. PLoS One.
12:e01822822017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yu S, Li G, Wang Z, Wang Z, Chen C, Cai S
and He Y: The prognostic value of pSTAT3 in gastric cancer: A
meta-analysis. J Cancer Res Clin Oncol. 142:649–657. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Aggarwal BB, Sethi G, Ahn KS, Sandur SK,
Pandey MK, Kunnumakkara AB, Sung B and Ichikawa H: Targeting
signal-transducer-and-activator-of-transcription-3 for prevention
and therapy of cancer: Modern target but ancient solution. Ann N Y
Acad Sci. 1091:151–169. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Pan YM, Wang CG, Zhu M, Xing R, Cui JT, Li
WM, Yu DD, Wang SB, Zhu W, Ye YJ, et al: STAT3 signaling drives
EZH2 transcriptional activation and mediates poor prognosis in
gastric cancer. Mol Cancer. 15:792016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Gelain A, Mori M, Meneghetti F and Villa
S: Signal transducer and activator of transcription protein 3
(STAT3): An update on its direct inhibitors as promising anticancer
agents. Curr Med Chem. Jul 19–2018.(Epub ahead of print).
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Schust J, Sperl B, Hollis A, Mayer TU and
Berg T: Stattic: A small-molecule inhibitor of STAT3 activation and
dimerization. Chem Biol. 13:1235–1242. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Song H, Wang R, Wang S and Lin J: A
low-molecular-weight compound discovered through virtual database
screening inhibits Stat3 function in breast cancer cells. Proc Natl
Acad Sci USA. 102:4700–4705. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Siddiquee K, Zhang S, Guida WC, Blaskovich
MA, Greedy B, Lawrence HR, Yip ML, Jove R, McLaughlin MM, Lawrence
NJ, et al: Selective chemical probe inhibitor of Stat3, identified
through structure-based virtual screening, induces antitumor
activity. Proc Natl Acad Sci USA. 104:7391–7396. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Hu QD, Chen W, Yan TL, Ma T, Chen CL,
Liang C, Zhang Q, Xia XF, Liu H, Zhi X, et al: NSC 74859 enhances
doxorubicin cytotoxicity via inhibition of epithelial-mesenchymal
transition in hepatocellular carcinoma cells. Cancer Lett.
325:207–213. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Chen W, Shen X, Xia X, Xu G, Ma T, Bai X
and Liang T: NSC 74859-mediated inhibition of STAT3 enhances the
anti-proliferative activity of cetuximab in hepatocellular
carcinoma. Liver Int. 32:70–77. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Lin L, Amin R, Gallicano GI, Glasgow E,
Jogunoori W, Jessup JM, Zasloff M, Marshall JL, Shetty K, Johnson
L, et al: The STAT3 inhibitor NSC 74859 is effective in
hepatocellular cancers with disrupted TGF-beta signaling. Oncogene.
28:961–972. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhang Q, Zhang C, He J, Guo Q, Hu D, Yang
X, Wang J, Kang Y, She R, Wang Z, et al: STAT3 inhibitor stattic
enhances radiosensitivity in esophageal squamous cell carcinoma.
Tumour Biol. 36:2135–2142. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Pan Y, Zhou F, Zhang R and Claret FX:
Stat3 inhibitor Stattic exhibits potent antitumor activity and
induces chemo- and radio-sensitivity in nasopharyngeal carcinoma.
PLoS One. 8:e545652013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Ahmad SF, Ansari MA, Nadeem A, Zoheir KMA,
Bakheet SA, Alsaad AMS, Al-Shabanah OA and Attia SM: STA-21, a
STAT-3 inhibitor, attenuates the development and progression of
inflammation in collagen antibody-induced arthritis. Immunobiology.
222:206–217. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Park JS, Kwok SK, Lim MA, Kim EK, Ryu JG,
Kim SM, Oh HJ, Ju JH, Park SH, Kim HY and Cho ML: STA-21, a
promising STAT-3 inhibitor that reciprocally regulates Th17 and
Treg cells, inhibits osteoclastogenesis in mice and humans and
alleviates autoimmune inflammation in an experimental model of
rheumatoid arthritis. Arthritis Rheumatol. 66:918–929. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Takaishi M, Yokogawa M, Miyoshi K,
Nakajima K, DiGiovanni J and Sano S: STA-21, a stat3 inhibitor,
induces differentiation of normal human epidermal keratinocytes and
human keratinocyte derived cell lines. J Invest Dermatol. 130
(Suppl):S97. 2010.
|
|
42
|
Ambrosini G, Adida C and Altieri DC: A
novel anti-apoptosis gene, survivin, expressed in cancer and
lymphoma. Nat Med. 3:917–921. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Baldin V, Lukas J, Marcote MJ, Pagano M
and Draetta G: Cyclin D1 is a nuclear protein required for cell
cycle progression in G1. Genes Dev. 7:812–821. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Burotto M, Chiou VL, Lee JM and Kohn EC:
The MAPK pathway across different malignancies: A new perspective.
Cancer. 120:3446–3456. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Chen F: JNK-induced apoptosis,
compensatory growth, and cancer stem cells. Cancer Res. 72:379–386.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Scuteri A, Galimberti A, Maggioni D,
Ravasi M, Pasini S, Nicolini G, Bossi M, Miloso M, Cavaletti G and
Tredici G: Role of MAPKs in platinum-induced neuronal apoptosis.
Neurotoxicology. 30:312–319. 2009. View Article : Google Scholar : PubMed/NCBI
|