Introduction
Esophageal cancer is a malignant tumor associated
with poor prognosis and high mortality rates (1). The incidence of esophageal cancer
exhibits marked geographic variation, appearing to be low in
Western Africa, and high in Japan, Southeastern Africa and Northern
China (2). Esophageal squamous
cell carcinoma (ESCC) is the dominant histological type of
esophageal cancer in China and it is associated with a particularly
high morbidity in certain areas, including the south of Hebei in
Northern China (3). The 5-year
survival rate of patients with ESCC with advanced-stage or
metastatic disease is <20% (4).
Tobacco and alcohol use have been demonstrated to be independent
risk factors for ESCC, but the exact pathogenetic mechanism remains
to be elucidated. An improved understanding of the mechanisms
underlying ESCC pathogenesis may identify promising molecular
biomarkers for the early diagnosis and prevention of this
malignancy.
Aberrant protein phosphorylation is a prerequisite
for the occurrence and progression of several tumors, and it is one
of the hallmarks of cancer cells (5). Protein kinases and protein
phosphatases serve key roles in regulating cellular signal
transduction. Protein phosphatases include protein serine/threonine
phosphatases, protein tyrosine phosphatases (PTPs),
tumor-suppressive metal-dependent protein phosphatases,
tumor-suppressive phosphoprotein phosphatases, tumor-suppressive
PTPs, receptor type and tumor-suppressive PTPs, non-receptor type
(PTPN). Protein phosphatases were initially considered to be tumor
suppressors, but several have been demonstrated to serve purely as
oncogenes, whereas others may serve as tumor suppressors and
oncogenes, according to the cellular environment or other
unidentified factors (6). PTPNs
are represented by 17 members and are absolutely specific to
phospho-tyrosine. They consist of a highly conserved catalytic
domain and variable regulatory domain arrays, acting by subcellular
targeting or directly regulating phosphatase activity (7). A number of PTPNs serve as tumor
suppressors by inhibiting Janus kinase/signal transducer and
activator of transcription (JAK/STAT) signaling, receptor tyrosine
kinase-mediated growth factor signaling, or cell motility/invasion.
Furthermore, certain PTPNs promote tumor-suppressive Hippo
signaling and inhibit tyrosine-protein kinase-ABL1-mediated
transformation through Yes-associated protein 1 (8).
Protein tyrosine phosphatase, non-receptor type 6
(PTPN6) (also known as HCP, HCPH or SHP1), is encoded by 17 exons
and has 2 Src homology 2 domains, is primarily expressed in
hematopoietic cells, and serves as a key regulator of multiple
signaling pathways in hematopoietic cells. PTPN6 has been
demonstrated to interact with and dephosphorylate a variety of
phospho-proteins involved in hematopoietic cell signaling pathways.
Plutzky et al (9) first
identified that PTPN6 is located in a chromosomal region that is
frequently damaged in childhood leukemia. Oka et al
(10) observed the loss of mRNA
and protein expression of PTPN6 in natural killer T-cell lymphomas
and 95% of several other types of malignant lymphomas, while only
60% of less malignant forms were negative. The loss of expression
of PTPN6 is likely associated with malignant transformation and
increased invasiveness. In subsequent studies, promoter
hypermethylation of PTPN6 was identified in several hematological
malignancies (11,12) and in solid tumors, including
nasopharyngeal carcinoma (13) and
breast ductal carcinoma (14). The
PTPN6 gene has two promoter regions that are 7 kb apart, and has 3
different transcripts. The longer transcript, driven by the P1
promoter, is expressed primarily in non-hematopoietic cells,
whereas the shorter transcript, driven by the P2 promoter (P2), is
only expressed in cells of the hematopoietic lineage (15). PTPN6 expression driven by P1 in
non-hematopoietic cells is low compared with the expression
regulated by P2 in hematopoietic cells (16). It has been suggested that
downregulation of PTPN6 is primarily due to DNA hypermethylation of
CpG islands in the PTPN6 P2 (10,11).
However, the mechanism and methylation status of PTPN6 in ESCC have
not yet been fully elucidated. The aims of the present study were
to investigate the expression of PTPN6 in ESCC tissues and
esophageal cancer cell lines, elucidate the role of CpG
hypermethylation in the inactivation of PTPN6, and improve the
understanding of the functional and prognostic significance of
PTPN6 in ESCC tumorigenesis and progression.
Materials and methods
Cell culture and treatment
A total of 5 human esophageal cancer cell lines
(Eca109, Kyse150, Kyse170, Yes-2 and TE1) and a human normal
esophageal epithelial cell (HEEpiC) line were purchased from
American Type Culture Collection (Manassas, VA, USA). All the cell
lines were cultured in RPMI-1640 medium (Invitrogen; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) supplemented with 10% fetal
bovine serum (FBS; Gibco; Thermo Fisher Scientific, Inc.) at 37°C
in a humidified atmosphere of 5% CO2, and were assessed
by reverse transcription-quantitative polymerase chain reaction
(RT-qPCR) analysis for mycoplasma contamination. All the cell lines
were seeded prior to drug treatment. Cells (1.5×105/ml)
were treated with 5 µM DNA methyltransferase inhibitor
5-aza-2′-deoxycytidine (5-Aza-dC; Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany) for the first 48 h and, subsequently, the
medium containing 5-Aza-dC was changed every 24 h. Control cells
were cultured in RPMI-1640 medium with no drug treatment.
Patients and specimens
A total of 71 primary ESCC samples and corresponding
adjacent normal tissues were collected by surgical resection
between January 2008 and January 2011 at the Department of Thoracic
Surgery of the Fourth Hospital of Hebei Medical University
(Shijiazhuang, China). The study was approved by the Ethics
Committee of the Fourth Hospital of Hebei Medical University, and
conformed to all relevant ethical regulations for human research
subjects in accordance with Declaration of Helsinki. All the
participants signed a written informed consent form. The patients
comprised 51 males and 20 females, with a median age of 62 years
(range, 39–78 years) (Table I).
Freshly removed ESCC and paired adjacent non-cancerous esophageal
tissues were divided into two groups, one of which was fixed in
formalin at room temperature and embedded in paraffin, and the
other was frozen and stored at −80°C for DNA and RNA isolation.
Clinical data and clinicopathological characteristics were
collected from medical records. The subjects were interviewed for
information on demographic and exogenous risk factors, including
smoking, alcohol consumption and family history.
 | Table I.Clinicopathologic characteristics of
esophageal squamous cell carcinoma cases. |
Table I.
Clinicopathologic characteristics of
esophageal squamous cell carcinoma cases.
| Group | N (%) |
|---|
| Age, years |
|
<62 | 30 (42.3) |
| ≥62 | 41 (57.7) |
| Sex |
| Male | 51 (71.8) |
|
Female | 20 (28.2) |
| TNM stage |
| I | 4 (5.6) |
| II | 24 (33.8) |
|
III | 38 (53.5) |
| IV | 5 (7.0) |
| Pathological
differentiation |
|
Well | 7 (9.9) |
|
Moderate | 30 (42.3) |
|
Poor | 34 (47.9) |
| Depth of
invasion |
|
T1/2 | 28 (39.4) |
|
T3/4 | 43 (60.6) |
| LN metastasis |
|
Negative (N0) | 22 (31.0) |
|
Positive (N1/2/3) | 49 (69.0) |
| Family history of
UGIC |
|
Negative | 47 (66.2) |
|
Positive | 24 (33.8) |
RT-qPCR analysis
Total RNA was extracted from cell lines and frozen
tumor tissues using TRIzol® reagent (Thermo Fisher
Scientific, Inc.). The RT-for-PCR kit (Invitrogen; Thermo Fisher
Scientific, Inc.) was used to synthesize single-stranded cDNA
according to the protocol of the manufacturer. The mRNA expression
levels were quantified using primers, cDNA template and Power
SYBR-Green PCR Master Mix (Promega Corporation, Madison, WI, USA),
according to the protocol of Power SYBR-Green PCR Master Mix The
primers used for PTPN6 are listed in Table II. The PCR cycle conditions were:
94°C for 30 sec, followed by 40 cycles of 94°C for 10 sec, 60°C for
30 sec and 72°C for 1 min. The data were analyzed by the
2−ΔΔCq method (17) and
the human GAPDH gene was used as an endogenous control.
| Table II.Primer sequences and reaction
conditions of PTPN6 used. |
Table II.
Primer sequences and reaction
conditions of PTPN6 used.
| PCR types | Gene | Primer
sequence | Annealing
temperature, °C | Product size,
bp |
|---|
| RT-qPCR | PTPN6 | F:
5′-GGCCTGGACTGTGACATTGA-3′ | 56 | 188 |
|
|
| R:
5′-ATGTTCCCGTACTCCGACTC-3′ |
|
|
|
| GAPDH | F:
5′-AGGTGAAGGTCGGAGTCAACG-3′ | 56 | 104 |
|
|
| R:
5′-AGGGGTCATTGATGGCAACA-3′ |
|
|
| BGS | PTPN6 | F:
5′-AGGGTTGTGGTGAGAAATTAATTAG-3′ | 58 | 222 |
|
|
| R:
5′-TTACACACTCCAAACCCAAATAATAC-3′ |
|
|
| BS-MSP | Methylation | F:
5′-GAACGTTATTATAGTATAGCGTTC-3′ | 60 | 158 |
|
|
| R:
5′-TCACGCATACGAACCCAAACG-3′ |
|
|
|
| Unmethylation | F:
5′-GTGAATGTTATTATAGTATAGTGTTTGG-3′ | 59 | 158 |
|
|
| R:
5′-TTCACACATACAAACCCAAACAAT-3′ |
|
|
Western blot analysis of PTPN6 protein
expression in ESCC cell lines
Total protein from cultured cell lines was extracted
using radioimmunoprecipitation assay reagent supplemented with
protease inhibitors (Thermo Fisher Scientific, Inc.). The protein
was quantified using BCA Protein Assay kit (Beyotime Biotechnology,
Shanghai, China). Protein samples (20 µg/lane) were prepared for
western blot analysis with 15% SDS-PAGE gels and transferred to a
polyvinylidene fluoride membrane (EMD Millipore, Billerica, MA,
USA). To detect the expression of PTPN6, the membranes were
incubated at 4°C overnight with the specific primary antibodies
(1:1,000; rabbit anti-human monoclonal antibody; cat. no. ab32559;
Abcam, Cambridge, UK). Subsequently, the secondary antibody (goat
anti-rabbit IgG-HRP; 1:2,000; cat. no. sc-2004; Santa Cruz
Biotechnology, Inc., Dallas, TX, USA) incubated with the membrane
at room temperature for 1 h. To ensure equal loading in all the
lanes, anti-β actin (1:1,000; cat. no. ab119716; Abcam) was used as
the control. Consequently, the protein bands were analyzed by the
Image Lab software version 4.1 (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA).
Immunohistochemical staining for the
PTPN6 protein in ESCC tissues
PTPN6 protein expression was determined by
immunostaining using the streptavidin-peroxidase method in tumor
samples and corresponding adjacent normal sections. Specimens were
embedded in paraffin and cut into 4-µm sections. Then descending
alcohol series were used to deparaffinization and rehydration were
used by descending alcohol series. Antigen retrieval was performed
in a pressure cooker at 100°C for 5 min in Tris-EDTA buffer (pH
9.0). Rabbit anti-human monoclonal antibody for PTPN6 (1:100
dilution; cat. no., ab32559; Abcam) was applied to investigate the
protein expression of PTPN6 at 4°C overnight. Following an
overnight incubation, specimens were subjected to the
streptavidin-peroxidase (SP) method using a standard SP kit (cat.
no. PV-9001; OriGene Technologies, Inc., Beijing, China) according
to the manufacturer's protocol. PBS (pH 9.0) was used as negative
control of the primary antibody. The slides were examined using a
light microscope (Olympus BX41; Olympus Corporation, Tokyo, Japan;
magnification, ×200 and ×400) and scored by experienced
pathologists in a double-blinded manner.
DNA extraction and sodium bisulfite
treatment
Genomic DNA was isolated from esophageal cancer cell
lines, frozen ESCC tumor samples and corresponding normal tissues
using a DNA extraction kit (Shanghai Generay Biotech Co. Ltd.,
Shanghai, China). To assess the DNA methylation patterns, DNA was
bisulfite-modified using an Epitect Fast Bisulfite Conversion kit
(Qiagen GmbH, Hilden, Germany), which converts unmethylated
cytosine residues to thymine, whereas methylated cytosine residues
remain unaffected.
Methylated CpG site distribution via
bisulfite genomic sequencing (BGS) assay
To analyze the DNA methylation pattern of the PTPN6
P2, a BGS assay was used to detect the methylated CpG site
distribution in the esophageal cancer cell lines. Subsequently, the
online MethPrimer program was used to detect the distribution of
CpG islands (URL: http://www.urogene.org/methprimer/). A pair of primers
(from-167 to-326 bp) was designed by Sangon Biotech Co., Ltd.
(Shanghai, China) to recognize sodium bisulfite-converted genomic
DNA. The primer sequence for BGS: Sense
5′-AGGGTTGTGGTGAGAAATTAATTAG-3′, and antisense
5′-TTACACACTCCAAACCCAAATAATAC-3′. The PCR products were purified
using the QIAEXII Gel Extraction kit (Qiagen GmbH) and cloned into
pGEM-T vectors (Promega Corporation). Up to 10 clones for each
specimen were analyzed by bisulfite sequencing.
Methylation analysis of PTPN6 via
bisulfite conversion-specific and methylation-specific polymerase
chain reaction (BS-MSP) assay
The PTPN6 P2 was analyzed by the BS-MSP method as
described above using bisulfite-treated genomic DNA. According to
the distribution of the primary methylated CpG sites by the BGS
assay, the MSP primers were designed by Sangon Biotech Co., Ltd.,
and the reaction conditions were summarized in Table II. According to the manufacturer's
recommendations, genomic DNA methylated in vitro by CpG
methyltransferase (Sss I) (New England BioLabs, Inc., Ipswich, MA,
USA) and water blanks were applied as positive and negative
controls, respectively. The BS-MSP products were analyzed on 2%
agarose gel with ethidium bromide staining. All reactions were
performed in duplicate for each of the samples.
Cell transfection
To determine the overexpression of PTPN6, Eca109 and
Yes-2 cells in the logarithmic growth phase were cultured in 6-well
plates. When the density of Eca109 and Yes-2 cells reached to 80%,
the cells were transfected with PTPN6 expression plasmid
(pcDNA3.1-PTPN6) or the empty vector (pcDNA3.1-EV) (Sangon Biotech
Co., Ltd.) as control at a final concentration of 2.5 µg/µl using
Lipofectamine® 2000 transfection reagent (Invitrogen;
Thermo Fisher Scientific, Inc.) according to the manufacturer's
instructions. Following transfection, the cells were incubated in
RPMI-1640 medium for 4–6 h, followed by replacement with RPMI-1640
supplemented with 10% FBS. After 24 h, the transfected cells were
extracted for subsequent experimentation.
Cell proliferation
The cells (1.0×105) were seeded into
96-well plates for the cell proliferation assays. The proliferation
of Eca109 and Yes-2 cells transfected with PTPN6 was determined by
MTS assay. The absorbance was measured at a wavelength of 492 nm,
followed by incubation for 4 h in a humidified incubator containing
5% CO2 at 37°C. The proliferation rates were determined
at 0, 24, 48, 72 and 96 h after transfection. All the experiments
were performed in triplicate.
Colony formation assay
For the colony formation assay, 2,500 cells were
seeded in 6-well plates and incubated with RPMI-1640 medium
containing 10% FBS for 1 week. Colonies (>50 cells) were fixed
in methanol for 15 min (at room temperature) and dyed with 0.5%
crystal violet solution for 20 min, and the colony number was
counted under an inverted microscope (DWI40CCB; Leica, Wetzlar,
Germany; magnification, ×100).
Wound healing assay
Cells (5.0×105) in the logarithmic growth
phase were inoculated in 6-well plates. Following transfection for
24 h, scratch wounds were created using a 200 µl pipette tip. The
detached cells were removed by washing with PBS 3 times. RPMI-1640
medium was then added to the plates and images were observed after
culture for 0, 12 and 24 h. The inverted microscope (Leica
Microsystems GmbH, Wetzlar, Germany; magnification, ×100) was
applied to measure the relative migration distance.
Cell invasion assay
The invasion of PTPN6-transfected Eca109 and Yes-2
cells was measured in 24-well Transwell chambers (Corning
Incorporated, Corning, NY, USA). The Transwell chambers were coated
with 20 µl Matrigel at 4°C and incubated at 37°C for 4 h. After 24
h transfection, 5,000 cells/well were seeded in the upper chambers,
and the lower chambers were filled with RPMI-1640 medium
supplemented with 10% FBS. Following incubation at 37°C for 24 h,
invading cells located in the lower chamber were fixed in 4%
paraformaldehyde at room temperature for 15 min and stained with
0.1% crystal violet at room temperature for 30 min. The number of
cells that had invaded through the membrane to the lower surface
was observed in 5 microscopic fields per filter under the inverted
microscope (Leica Microsystems GmbH; magnification, ×100). The
experiments were performed in triplicate.
Statistical analysis
Statistical analysis was performed with SPSS 22.0
software package (IBM Corp., Armonk, NY, USA). The RT-qPCR results
are presented as the mean ± standard deviation. Student's t-test
was applied to compare the expression means between different
continuous variables. Pearson's χ2 test was applied to
assess the status of gene methylation. For prognostic analysis of
PTPN6 protein expression and methylation, survival curves were
constructed using the Kaplan-Meier method and the log-rank or the
Breslow tests. One-way analysis of variance was adopted to measure
the comparison of multiple groups (the function of PTPN6 in
esophageal cancer cell lines), and within-group variations were
performed by Student Newman-Keuls test. All statistical tests were
two-sided and P<0.05 was considered to indicate a statistically
significant difference.
Results
mRNA and protein expression of PTPN6
is decreased in esophageal cancer cell lines and ESCC tissues
The mRNA expression of PTPN6 was first detected in 5
esophageal cancer cell lines and normal human esophageal epithelial
cells. As demonstrated in Fig. 1A,
the mRNA expression of PTPN6 was markedly decreased in the ESCC
TE1, Eca109, Kyse150, Kyse170 and Yes-2 cell lines compared with
that in the normal esophageal epithelial HEEpiC cells. This result
was additionally confirmed by the results of western blot analysis
(Fig. 1B).
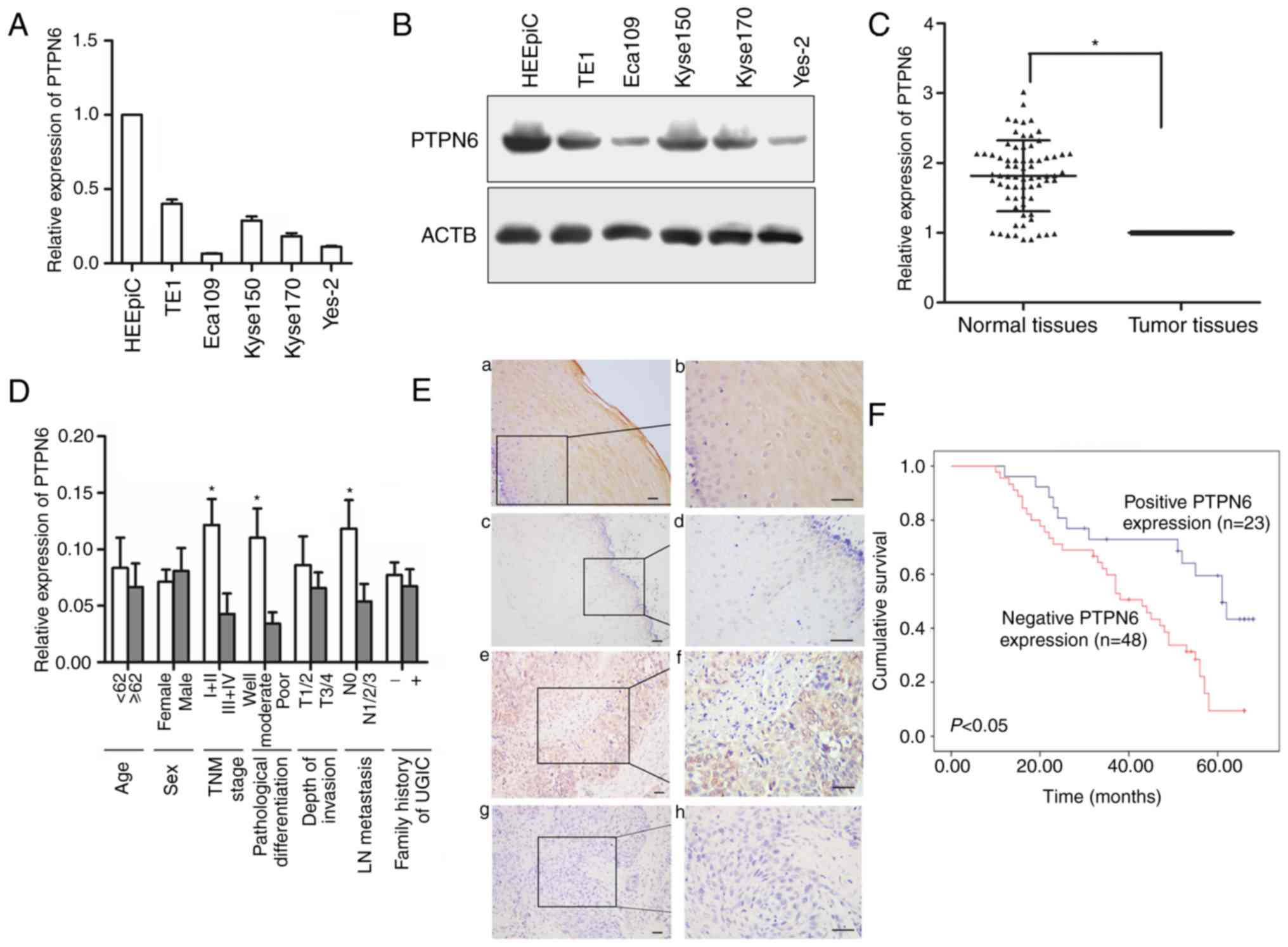 | Figure 1.Expression of PTPN6 in human
esophageal cancer cell lines and ESCC tissues. (A) Relative mRNA
expression of PTPN6 in 5 esophageal cancer cell lines and human
normal esophageal epithelial cells detected by reverse
transcription-quantitative polymerase chain reaction analysis. (B)
Protein expression of PTPN6 in esophageal cancer cell lines and
human normal esophageal epithelial cells detected by western blot
analysis. (C) Relative mRNA expression of PTPN6 in normal tissues
and corresponding ESCC tumor tissues; *P<0.05. (D) Relative mRNA
expression of PTPN6 in different subgroups. *P<0.05 vs. TNM:
I+IIstage vs. III+IV stage; Well + moderate differentiation vs.
poor differentiation; Positive lymph node metastasis vs. negative.
lymph node metastasis. (E) Streptavidin-peroxidase
immunohistochemical staining of PTPN6 in ESCC tumor tissues and
corresponding normal tissues (SP9001): (a and b) Positive staining
of PTPN6 in normal tissues at (a) magnification, ×200 and (b)
magnification, ×400; (c and d) negative staining of PTPN6 in normal
tissues at (c) magnification, ×200 and (d) magnification, ×400; (e
and f) positive staining of PTPN6 in ESCC tissues at (e)
magnification, ×200 and (f) magnification, ×400; (g and h) negative
staining of PTPN6 in ESCC tissues at (g) magnification, ×200 and
(h) magnification ×400. (F) Direct association between negative
PTPN6 protein expression and poor patient survival. PTPN6, protein
tyrosine phosphatase, non-receptor type 6; ESSC, esophageal
squamous cell carcinoma; TNM, tumor node metastasis; LN, lymph
node; UGIC, upper gastrointestinal cancer. |
The mRNA expression of PTPN6 in ESCC tumor tissues
was significantly decreased compared with that in the corresponding
normal tissues (P<0.01; Fig.
1C). PTPN6 mRNA expression was associated with Tumor Node
Metastasis stage (18),
pathological differentiation and lymph node (LN) metastasis
(Fig. 1D). Immunohistochemical
staining was used to assess the protein expression of PTPN6 in
tumor tissues and corresponding normal tissues. Protein expression
of PTPN6 was detected primarily in the cytoplasm and the nucleus of
tumor or normal cells (Fig. 1E).
The protein expression of PTPN6 in tumor tissues (32.4%, 23/71) was
markedly decreased compared with that in corresponding normal
tissues (77.5%, 55/71; P<0.01; Table III). When stratified for
clinicopathological characteristics, PTPN6 protein expression was
identified to be significantly associated with
tumor-node-metastasis (TNM) stage, pathological differentiation and
LN metastasis (P<0.05; Table
IV).
| Table III.Protein expression and methylation
status of PTPN6 in ESCC tumor tissues and corresponding normal
tissues. |
Table III.
Protein expression and methylation
status of PTPN6 in ESCC tumor tissues and corresponding normal
tissues.
|
|
| Protein
expression | Methylation
frequency |
|---|
|
|
|
|
|
|---|
| Group | N | n (%) | P-value | n (%) | P-value |
|---|
| Normal tissues | 71 | 55 (77.5) | <0.001 | 12 (16.9) | <0.001 |
| Tumor tissues | 71 | 23 (32.4) |
| 45 (63.4) |
|
| Table IV.Immunohistochemical staining
characteristics and methylation status of PTPN6 in ESCC
tissues. |
Table IV.
Immunohistochemical staining
characteristics and methylation status of PTPN6 in ESCC
tissues.
|
|
| Protein
expression | Methylation
frequency |
|---|
|
|
|
|
|
|---|
| Group | N | n (%) | P-value | n (%) | P-value |
|---|
| Age, years |
|
<62 | 30 | 12 (40.0) | 0.241 | 17 (56.7) | 0.315 |
|
≥62 | 41 | 11 (26.8) |
| 28 (68.3) |
|
| Sex |
|
Male | 51 | 15 (29.4) | 0.391 | 35 (68.6) | 0.143 |
|
Female | 20 | 8 (40.0) |
| 10 (50.0) |
|
| TNM stage |
|
I+II | 28 | 16 (57.1) | <0.001 | 12 (42.9) | 0.004 |
|
III+IV | 43 | 7 (16.3) |
| 33 (76.7) |
|
| Pathological
differentiation |
|
Well/moderate | 37 | 21 (56.7) | <0.001 | 18 (48.6) | 0.007 |
|
Poor | 34 | 2 (5.9) |
| 27 (79.4) |
|
| Depth of
invasion |
|
T1/2 | 28 | 11 (39.3) | 0.317 | 14 (50.0) | 0.059 |
|
T3/4 | 43 | 12 (27.9) |
| 31 (72.1) |
|
| LN metastasis |
|
Negative (N0) | 22 | 11 (50.0) | 0.034 | 9 (40.9) | 0.008 |
|
Positive (N1/2/3) | 49 | 12 (24.5) |
| 36 (61.0) |
|
| Family history of
UGIC |
|
Negative | 47 | 17 (36.2) | 0.341 | 30 (63.8) | 0.912 |
|
Positive | 24 | 6 (25.0) |
| 15 (62.5) |
|
Downregulation of PTPN6 is associated
with poor ESCC patient survival
The 5-year survival rate in the positive and
negative PTPN6 expression ESCC groups was 47.8 and 20.8%,
respectively (P<0.05; log-rank test). As presented in Fig. 1F, patients with ESCC negative for
protein expression of PTPN6 exhibited poor survival.
Upregulation of PTPN6 by 5-Aza-dC
treatment in esophageal cancer cell lines
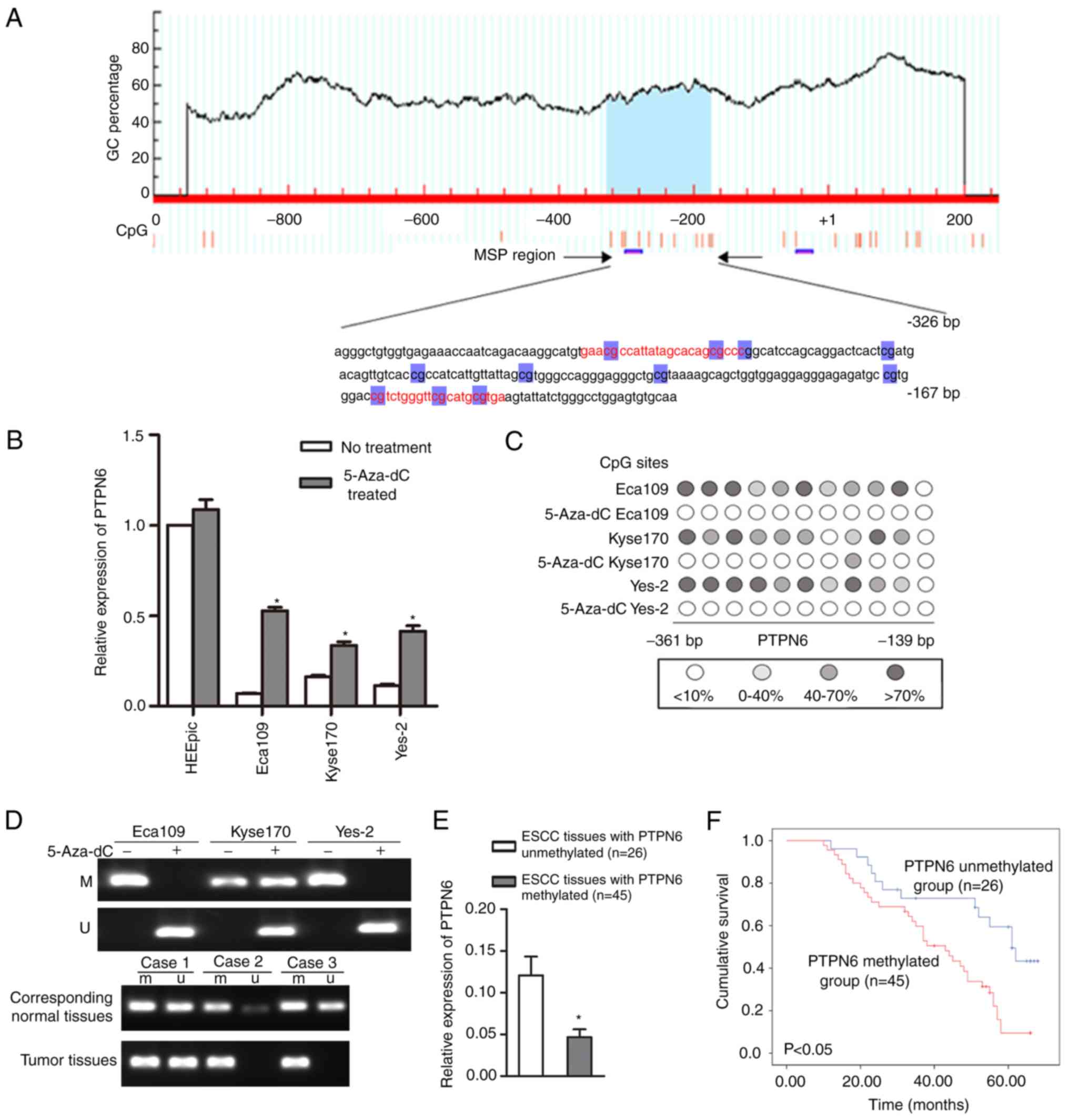
As demonstrated in Fig.
2A, the online MethPrimer program was used to detect the
distribution of CpG islands in the PTPN6 promoter region and
genomic sequence. A total of 1 CpG island was identified to be
located in the promoter region. The Eca109, Kyse170 and Yes-2 cell
lines, which exhibited a relatively low PTPN6 expression, were
subsequently treated with 5-Aza-dC. As indicated in Fig. 2B, the mRNA expression level of
PTPN6 was markedly upregulated in these 3 esophageal cancer cell
lines following treatment with 5-Aza-dC, suggesting that aberrant
methylation may be one of the mechanisms leading to PTPN6 silencing
in esophageal cancer cell lines.
Methylation analysis of PTPN6 in
esophageal cancer cell lines and tumor tissues
The methylation status of the CpG sites in the
promoter region of PTPN6 was first verified by BGS assay in
esophageal cancer cell lines, and frequent hypermethylation of the
CpG sites in the promoter region of PTPN6 was detected in Eca109,
Kyse170 and Yes-2 cells (Fig. 2C).
In particular, fully methylated PTPN6 in the Eca109 and Yes-2 cell
lines was detected by the BS-MSP assay (Fig. 2D). Following treatment with
5-Aza-dC, the aberrant methylation status of the cells was reversed
in the 3 cell lines. The frequency of PTPN6 methylation in ESCC
tumor tissues (63.4%, 45/71) was significantly higher compared with
that in corresponding normal tissues (16.9%, 12/71; P<0.05;
Table III and Fig. 2D). When stratified for
clinicopathological characteristics, the methylation frequency of
PTPN6 was associated with TNM stage, pathological differentiation
and LN metastasis (P<0.05). However, the methylation status of
PTPN6 in ESCC tumor tissues was not associated with age or sex
(P>0.05; Table IV).
Association between PTPN6 expression
and methylation status
As demonstrated in Fig.
2E, the mRNA expression level of PTPN6 in ESCC tissues with
PTPN6 methylation was significantly decreased compared with that in
ESCC tissues with unmethylated PTPN6 (P<0.05). Similarly, the
protein expression of PTPN6 in ESCC tissues with PTPN6 methylation
was significantly decreased compared with that in ESCC tissues with
unmethylated PTPN6 (P<0.05; Table
V).
| Table V.Association between PTPN6 protein
expression and methylation status in patients with esophageal
squamous cell carcinoma. |
Table V.
Association between PTPN6 protein
expression and methylation status in patients with esophageal
squamous cell carcinoma.
|
| Protein
expression |
|---|
|
|
|
|---|
| Group | + | − | P-value |
|---|
| Methylation
status |
|
| <0.001 |
| M | 6 | 39 |
|
| U | 17 | 9 |
|
Promoter hypermethylation of PTPN6 is
associated with poor ESCC patient survival
As demonstrated in Fig.
2F, PTPN6 methylation was identified to be negatively
associated with ESCC patient survival. In patients with ESCC with
hypermethylation of PTPN6, the 5-year survival rate was 17.8%
compared with 50.0% in patients with ESCC with unmethylated PTPN6
(P<0.05; log-rank test).
Upregulation of PTPN6 inhibits
esophageal cancer cell proliferation and invasion in vitro
The function of PTPN6 was then investigated in
esophageal cancer cell lines. The construct containing PTPN6
transcripts (pcDNA3.1-PTPN6) was transfected into Eca109 and Yes-2
cells. As indicated in Fig. 3A and
B, significant upregulation of PTPN6 was detected in
pcDNA3.1-PTPN6-transfected Eca109 and Yes-2 cells. Transfection of
PTPN6 led to a marked inhibition of Eca109 and Yes-2 cell
proliferation, as detected by the MTS assay (Fig. 3C). The results were additionally
verified with the colony formation assay (Fig. 3D). Furthermore, the wound healing
assay was performed, starting 24 h after pcDNA3.1-PNPN6
transfection. Overexpression of PTPN6 effectively decreased the
area of the scratch covered (Fig.
3E). Similarly, the Transwell assay confirmed a decrease in the
migration ability of pcDNA3.1-PTPN6-transfected cells (Fig. 3F). These results indicate that
PTPN6 inhibited the proliferation, migration and invasion of Eca109
and Yes-2 cells in vitro.
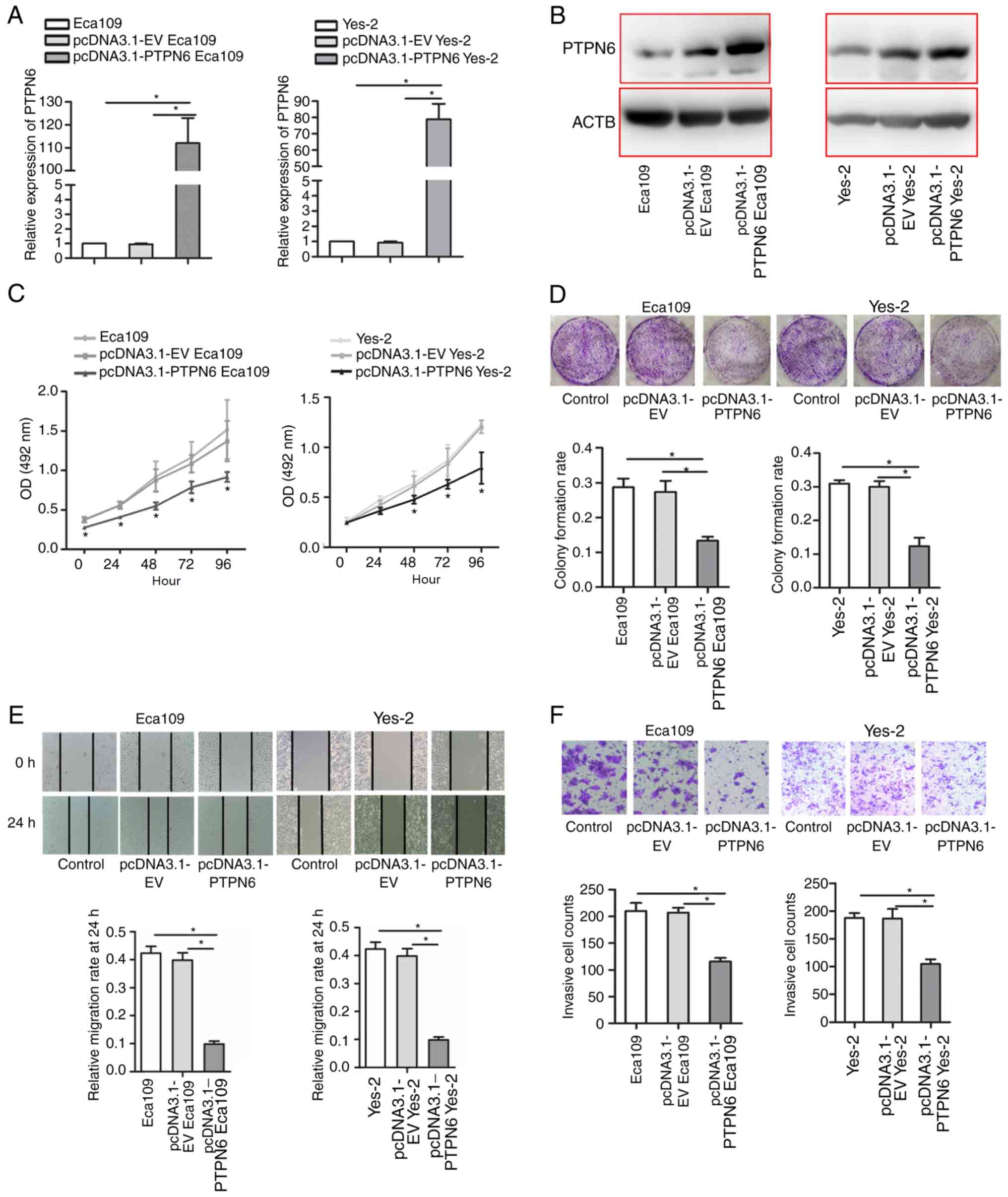 | Figure 3.Functional analysis of PTPN6 in human
esophageal cancer cell lines. (A) Overexpression of PTPN6 was
detected by reverse transcription-quantitative polymerase chain
reaction in PTPN6-transfected Eca109 and Yes-2 cells compared with
empty vector-transfected cells. *P<0.05. (B) Overexpression of
PTPN6 was detected by western blot analysis in PTPN6-transfected
Eca109 and Yes-2 cells compared with empty vector-transfected
cells. (C) Overexpression of PTPN6 inhibited Eca109 and Yes-2 cell
proliferation, as detected by the MTS assay; *P<0.05 vs. empty
vector. (D) Colony formation assay of PTPN6 cells transiently
overexpressing PTPN6. *P<0.05. (E) Upregulation of PTPN6
inhibited Eca109 and Yes-2 cell migration, as detected by the wound
healing assay. *P<0.05. Magnification, ×200 (F) Overexpression
of PTPN6 inhibited Eca109 and Yes-2 cell invasiveness, as detected
by the Transwell invasion assay. The results were determined by
counting the cells that had penetrated through the Matrigel-coated
Transwell chambers (8-µm pore size). *P<0.05 vs. EV.
Magnification, ×100. PTPN6, protein tyrosine phosphatase,
non-receptor type 6; ACTB, β-actin; OD, optical density; EV, empty
vector. |
Discussion
The PTPN6 gene is located on human chromosome 12p13
and encodes a Mr 68,000 non-receptor type protein-tyrosine
phosphatase. The PTPN6 gene has been considered as a candidate
tumor suppressor in hematological and solid malignancies, and
promoter methylation may be an important epigenetic mechanism
involved in silencing its expression. However, the detailed roles
of PTPN6 and its promoter methylation status in the pathogenesis of
primary ESCC remain elusive. In the present study, significant
downregulation of PTPN6 and frequent hypermethylation of the CpG
sites within the P2 were detected in esophageal cancer cell lines
and ESCC tissues. The mRNA expression level of PTPN6 was
significantly upregulated in 5-Aza-dC-treated esophageal cancer
cells. In addition, the methylation status and expression of PTPN6
were associated with TNM stage, pathological differentiation and LN
metastasis in patients with ESCC. Additional study verified that
aberrant hypermethylation of the P2 exhibited higher tumor
specificity and was associated with the expression level of PTPN6.
Survival analysis demonstrated that downregulation and
hypermethylation of PTPN6 were associated with poor ESCC patient
survival. Furthermore, upregulation of PTPN6 inhibited the
proliferation and invasion of esophageal cancer cells in
vitro.
Genomic DNA methylation is an important epigenetic
event in humans, and the alterations of methylation patterns may
serve important roles in tumorigenesis (19–21).
As aberrant DNA methylation is one of the earliest molecular
changes during the transformation process from normal to cancerous
cells (22), detection of an
aberrant DNA methylation pattern may have potential applications in
the early detection of malignancies. Transcriptional silencing of
PTPN6 due to promoter hypermethylation has been previously
demonstrated in several hematopoietic cell lines, leukemia and
lymphoma (10,23,24).
A high frequency of promoter hypermethylation was also observed in
endometrial carcinoma, and was identified to be associated with
patient age and tumor differentiation. PTPN6 promoters were
completely methylated in endometrial carcinoma cell lines, and this
methylation status was reversed by 5-Aza-dC treatment (25). In the present study, downregulation
of PTPN6 was detected in esophageal cancer cell lines and ESCC
tissues, and P2 hypermethylation may be one of the important
epigenetic mechanisms silencing this gene in ESCC. Furthermore, the
expression and methylation status of PTPN6 were associated with TNM
stage, pathological differentiation and LN metastasis in patients
with ESCC, and were associated with patient survival, indicating
that detection of the P2 methylation status may be a promising
biomarker for predicting the prognosis of ESCC.
It has been suggested that epigenetic silencing of
PTPN6 in myeloproliferative neoplasms and K562 cells causes
constitutive activation of JAK/STAT signaling (26). The reversal of PTPN6 expression by
5-Aza-dC treatment caused decreased expression levels of p-STAT3,
p-JAK3 and JAK3, but not of the STAT3 protein (27). The JAK/STAT signaling pathway is
one of the most important signaling cascades that regulate immune
response, cell growth, differentiation and other cellular
biological activities (28). The
silence of PTPN6 may result in JAK or STAT activation in cancer
cells (11,24). However, the role of PTPN6 in ESCC
has not been fully elucidated. In the present study, it was
confirmed that upregulation of PTPN6 inhibited the proliferation
and invasion of esophageal cancer cells in vitro, indicating
that PTPN6 may serve as a tumor suppressor gene by inhibiting the
proliferation and invasion of cancer cells. However, the specific
regulated pathway of PTPN6 in ESCC requires additional
investigation.
In summary, PTPN6 may serve as tumor suppressor gene
in ESCC and inhibit esophageal cancer cell proliferation and
invasion. The P2 is frequently methylated in esophageal cancer
cells and ESCC tissues, and this may be one of the epigenetic
mechanisms implicated in PTPN6 silencing in ESCC. Furthermore,
PTPN6 may serve as a potential prognostic marker for predicting
survival in patients with ESCC.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data generated and analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
LL conducted the analyses, participated in the
overall conceptualization of the study, wrote the final manuscript
and performed the computational analyses. JL conceptualized and
supervised the study. SZ and XL participated in the analysis of
results. All authors have read and approved the final version of
this manuscript.
Ethics approval and consent to
participate
The study was approved by the Ethics Committee of
The Fourth Hospital of Hebei Medical University, and conformed to
all relevant ethical regulations for human research subjects. All
the participants signed a written informed consent form.
Patient consent for publication
All the participants signed a written informed
consent form.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
P2
|
promoter 2
|
|
ESCC
|
esophageal squamous cell carcinoma
|
|
PTPs
|
protein tyrosine phosphatases
|
|
PTPN6
|
protein tyrosine phosphatase,
non-receptor type 6
|
References
|
1
|
Lambert R and Hainaut P: The
multidisciplinary management of gastrointestinal cancer.
Epidemiology of oesophagogastric cancer. Best Pract Res Clin
Gastroenterol. 21:921–945. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Enzinger PC and Mayer RJ: Esophageal
cancer. N Engl J Med. 349:2241–2252. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Guohong Z, Min S, Duenmei W, Songnian H,
Min L, Jinsong L, Hongbin L, Feng Z, Dongping T, Heling Y, et al:
Genetic heterogeneity of oesophageal cancer in high-incidence areas
of southern and northern China. PLoS One. 5:e96682010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Pennathur A, Gibson MK, Jobe BA and
Luketich JD: Oesophageal carcinoma. Lancet. 381:400–412. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Meeusen B and Janssens V: Tumor
suppressive protein phosphatases in human cancer: Emerging targets
for therapeutic intervention and tumor stratification. Int J
Biochem Cell Biol. 96:98–134. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Elson A: Stepping out of the shadows:
Oncogenic and tumor-promoting protein tyrosine phosphatases. Int J
Biochem Cell Biol. 96:135–147. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tonks NK: Protein tyrosine
phosphatases-from housekeeping enzymes to master regulators of
signal transduction. FEBS J. 280:346–378. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kleppe M, Soulier J, Asnafi V, Mentens N,
Hornakova T, Knoops L, Constantinescu S, Sigaux F, Meijerink JP,
Vandenberghe P, et al: PTPN2 negatively regulates oncogenic JAK1 in
T-cell acute lymphoblastic leukemia. Blood. 117:7090–7098. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Plutzky J, Neel BG, Rosenberg RD, Eddy RL,
Byers MG, Jani-Sait S and Shows TB: Chromosomal localization of an
SH2-containing tyrosine phosphatase (PTPN6). Genomics. 13:869–872.
1992. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Oka T, Yoshino T, Hayashi K, Ohara N,
Nakanishi T, Yamaai Y, Hiraki A, Sogawa CA, Kondo E, Teramoto N, et
al: Reduction of hematopoietic cell-specific tyrosine phosphatase
SHP-1 gene expression in natural killer cell lymphoma and various
types of lymphomas/leukemias: Combination analysis with cDNA
expression array and tissue microarray. Am J Pathol. 159:1495–1505.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chim CS, Fung TK, Cheung WC, Liang R and
Kwong YL: SOCS1 and SHP1 hypermethylation in multiple
myeloma: Implications for epigenetic activation of the Jak/STAT
pathway. Blood. 103:4630–4635. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Amin HM, Hoshino K, Yang H, Lin Q, Lai R
and Garcia-Manero G: Decreased expression level of SH2
domain-containing protein tyrosine phosphatase-1 (Shp1) is
associated with progression of chronic myeloid leukaemia. J Pathol.
212:402–410. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Challouf S, Ziadi S, Zaghdoudi R, Ksiaa F,
Ben Gacem R and Trimeche M: Patterns of aberrant DNA
hypermethylation in nasopharyngeal carcinoma in Tunisian patients.
Clin Chim Acta. 413:795–802. 2010. View Article : Google Scholar
|
|
14
|
Hachana M, Trimeche M, Ziadi S, Amara K
and Korbi S: Evidence for a role of the Simian Virus 40 in human
breast carcinomas. Breast Cancer Res Treat. 113:43–58. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Banville D, Stocco R and Shen SH: Human
protein tyrosine phosphatase 1C (PTPN6) gene structure: Alternate
promoter usage and exon skipping generate multiple transcripts.
Genomics. 27:165–173. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tsui FW, Martin A, Wang J and Tsui HW:
Investigations into the regulation and function of the SH2
domain-containing protein-tyrosine phosphatase, SHP-1. Immunol Res.
35:127–136. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2−ΔΔCT method. Methods. 25:402–408. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yamasaki M, Miyata H, Miyazaki Y,
Takahashi T, Kurokawa Y, Nakajima K, Takiguchi S, Mori M and Doki
Y: Evaluation of the nodal status in the 7th edition of the
UICC-TNM classification for esophageal squamous cell carcinoma:
Proposed modifications for improved survival stratification: Impact
of lymph node metastases on overall survival after esophagectomy.
Ann Surg Oncol. 21:2850–2856. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Warnecke PM and Bestor TH: Cytosine
methylation and human cancer. Curr Opin Oncol. 12:68–73. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Parris TZ, Kovács A, Hajizadeh S, Nemes S,
Semaan M, Levin M, Karlsson P and Helou K: Frequent MYC
coamplification and DNA hypomethylation of multiple genes on 8q in
8p11-p12-amplified breast carcinomas. Oncogenesis. 3:e952014.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Vasiljević N, Scibior-Bentkowska D,
Brentnall AR, Cuzick J and Lorincz AT: Credentialing of DNA
methylation assays for human genes as diagnostic biomarkers of
cervical intraepithelial neoplasia in high-risk HPV positive women.
Gynecol Oncol. 132:709–714. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
El-Osta A, Baker EK and Wolffe AP:
Profiling methyl-CpG specific determinants on transcriptionally
silent chromatin. Mol Biol Rep. 28:209–215. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Oka T, Ouchida M, Koyama M, Ogama Y,
Takada S, Nakatani Y, Tanaka T, Yoshino T, Hayashi K, Ohara N, et
al: Gene silencing of the tyrosine phosphatase SHP1 gene by
aberrant methylation in leukemias/lymphomas. Cancer Res.
62:6390–6394. 2002.PubMed/NCBI
|
|
24
|
Chim CS, Wong KY, Loong F and Srivastava
G: SOCS1 and SHP1 hypermethylation in mantle cell
lymphoma and follicular lymphoma: Implications for epigenetic
activation of the Jak/STAT pathway. Leukemia. 18:356–358. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sheng Y, Wang H, Liu D and Zhang C, Deng
Y, Yang F, Zhang T and Zhang C: Methylation of tumor suppressor
gene CDH13 and SHP1 promoters and their epigenetic regulation by
the UHRF1/PRMT5 complex in endometrial carcinoma. Gynecol Oncol.
140:145–151. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhang MY, Fung TK, Chen FY and Chim CS:
Methylation profiling of SOCS1, SOCS2, SOCS3, CISH and
SHP1 in Philadelphia-negative myeloproliferative neoplasm. J
Cell Mol Med. 17:1282–1290. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Han Y, Amin HM, Frantz C, Franko B, Lee J,
Lin Q and Lai R: Restoration of shp1 expression by
5-AZA-2′-deoxycytidine is associated with downregulation of
JAK3/STAT3 signaling in ALK-positive anaplastic large cell
lymphoma. Leukemia. 20:1602–1609. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Niwa Y, Kanda H, Shikauchi Y, Saiura A,
Matsubara K, Kitagawa T, Yamamoto J, Kubo T and Yoshikawa H:
Methylation silencing of SOCS-3 promotes cell growth and migration
by enhancing JAK/STAT and FAK signalings in human hepatocellular
carcinoma. Oncogene. 24:6406–6417. 2005. View Article : Google Scholar : PubMed/NCBI
|