Introduction
Hepatocellular carcinoma (HCC) is the third leading
cause of cancer-related mortality worldwide (1). While diverse treatments exist for
HCC, the clinical prognosis of patients with HCC remains poor
(2). Therefore, it is necessary to
investigate the molecular mechanism underlying HCC. Second
mitochondria-derived activator of caspase (SMAC) is a protein
located in the mitochondria that can regulate apoptosis (3). SMAC can be released from mitochondria
into the cytoplasm where it binds to cellular inhibitor of
apoptosis 1 (cIAP1) and inhibits the anti-apoptotic function of
cIAP1, thus promoting apoptosis (4,5). As
a result of previous in-depth studies showing the structure and
function of SMAC protein, the application of SMAC mimetics in the
treatment of various diseases, especially tumors, has become a
novel research hotspot (6–9). SMAC mimetics have been used for the
treatment of cancer, including breast cancer, HCC and lung cancer
(6,7). Birinapant is a SMAC mimetic that can
promote apoptosis of tumor cells by binding to cIAP1 and inducing
cIAP1 degradation (8,9). The caspase-8 inhibitor emricasan can
combine with birinapant to induce necroptosis and treat acute
myeloid leukemia (10).
Additionally, birinapant can act as a nanomicellar carrier of
paclitaxel delivery for cancer therapy (11); and it also has anti-inflammatory
effects (12). Birinapant
attenuates lipopolysaccharide (LPS)-induced liver injury by
inhibiting tumor necrosis factor receptor-associated factor 3
(TRAF3) degradation in Kupffer cells (12). However, whether birinapant affects
HCC growth and the possible underlying molecular mechanism of this
effect are still unknown. In the present study, birinapant and
liver cancer cell lines Huh7, H22 and HepG2 were used to study the
effects and molecular mechanisms of birinapant on the proliferation
and apoptotic rate of liver cancer cells. It was found that
birinapant could promote apoptosis, as well as inhibiting the
proliferation and invasion of liver cancer cells. The molecular
mechanism may be related to the activation of the cIAP1/TRAF3
signaling pathway by birinapant in liver cancer cells.
Materials and methods
Cell culture and treatment
The liver cancer cell lines Huh7, HepG2 and H22, and
the hepatocyte line AML12 were obtained from the Chinese Academy of
Science. All cells were cultured in DMEM (Gibco; Thermo Fisher
Scientific, Inc.) containing 10% FBS (PAN-Biotech GmbH) with 1%
penicillin and 1% streptomycin. Cells were maintained at 37°C in a
humidified atmosphere containing 5% CO2. Huh7, HepG2 and
H22 cells were pretreated in a cell incubator with 300 nmol/ml
birinapant (MedChemExpress) at 37°C for 24 h prior to subsequent
experimentation.
Lentivirus-mediated knockdown of TRAF3
in Huh7 and H22 cells
The lentivirus was obtained from Shanghai GeneChem
Co., Ltd. (shRNA-TRAF3 was inserted into these lentiviruses by
Shanghai GeneChem Co., Ltd). The lentivirus-mediated knockdown of
TRAF3 in Huh7 and H22 cells was performed as previously described
(13). A total of 4 µl of
lentivirus titer (1×108 TU/ml) with shRNA was added to
1×106 Huh7 and H22 cells. Lentivirus without shRNA
served as the negative control, and the blank control group was
Huh7 and H22 cells without the lentivirus vector, these cells were
continuously cultured in a cell incubator at 37°C for 72 h. At 72 h
later, green fluorescence observed under an inverted fluorescent
microscope (×200) and western blotting were used to determine the
efficiency of lentivirus-mediated knockdown.
Establishment of a subcutaneous tumor
model in BALB/c nude mice
Male, 7-week-old BALB/c nude mice (~13 g) were
obtained from the Animal Experimental Center of Chongqing Medical
University. All mice were fed with common feed and sterile water
ad libitum, and housed in 22°C with 50% humidity, with 12 h
light/dark cycles in a cage of 5 mice. In total, 18 mice were
divided into three groups (Control group; shRNA-TRAF3 + Birinapant
group; and Birinapant group) with six mice in each group.
Subcutaneous tumor models were prepared by subcutaneously injecting
2×107 H22 cells into the left armpit of each mouse. In
the birinapant group, nude mice were injected intraperitoneally
with 30 mg/kg body weight birinapant the day after the subcutaneous
injection, and then again 1 week later. In the Control group, mice
were injected with an equivalent volume of saline at the same time
points as birinipant injection. The subcutaneous tumors were
observed every day, and after 15 days the subcutaneous tumor
tissues were removed for subsequent analyses.
Western blotting
After the cells were harvested, the cellular protein
was extracted using a protein extraction kit (Nanjing KeyGen
Biotech Co., Ltd.). Protein concentration was detected by BCA
protein assay kit (Beyotime Institute of Biotechnology) and a
microplate reader (Synergy™ HT; BioTek). Total cellular protein was
mixed with loading buffer and boiled in water for 10 min. In total,
50 µg protein lysate was separated by SDS-PAGE with 10% gels and
transferred to PVDF membranes (EMD Millipore), which were blocked
with 5% skim milk for 1 h at room temperature. The membranes were
then incubated overnight at 4°C with antibodies against cIAP1
(1:1,000; cat. no. ab189193; Abcam), TRAF3 (1:1,000; cat. no.
ab36988; Abcam) and β-actin (1:1,000; cat. no. 4970; Cell Signaling
Technology, Inc.). The membranes were further incubated with the
corresponding horseradish peroxidase (HRP)-conjugated secondary
antibody (1:10,000; cat. no. 7074; Cell Signaling Technology, Inc.)
at 37°C for 1 h. Bands were analyzed and scanned using Quantity One
software (version 5.2.1; Bio-Rad Laboratories, Inc.) after
incubation with enhanced chemiluminescence reagent (EMD Millipore)
at room temperature for 10 sec.
RNA isolation and reverse
transcription-quantitative (RT-q)PCR
Total RNA was isolated from Huh7, H22, HepG2 and
AML12 cells using TRIzol® reagent (Invitrogen; Thermo
Fisher Scientific, Inc.) according to the manufacturer's protocol.
Then, the RNA was reverse transcribed to cDNA with the PrimeScript™
RT Reagent kit (Takara Biotechnology Co., Ltd.) at 37°C for 15 min,
85°C for 5 sec and 4°C hold. RT-qPCR was performed using SYBR-Green
(Takara Biotechnology, Co., Ltd.) and an ABI PRISM 7900 Sequence
Detection system (Applied Biosystems; Thermo Fisher Scientific,
Inc.). The thermocycling conditions were as follows: 95°C for 30
sec for one cycle; 95°C for 5 sec and 60°C for 30 sec for 39
cycles; 65°C for 5 sec and 95°C for 15 sec for one cycle. The gene
expression level of each target gene was normalized to that of
GAPDH for each sample. The primers used were as follows: cIAP1,
forward 5′-GATACGAATGAAAGGCCAAG-3′, reverse
5′-TCTCCAGGTCCAAAATGAAT-3′; TRAF3, forward
5′-CTCCCATCAAGAAGCCACC-3′, reverse 5′-TGTAGGGCCTCTTTGAGGTAA-3′; and
GAPDH, forward 5′-CACCCACTCCTCCACCTTTG-3′ and reverse
5′-CCACCACCCTGTTGCTGTAG-3′. The relative gene expression was
analyzed using the 2−∆∆Cq method (14).
Immunohistochemical staining
Immunohistochemical staining was performed as
previously described (15).
Subcutaneous tumor tissues (n=18) were fixed in 4% formalin at room
temperature for 48 h and embedded in paraffin. Then, sections (4
µm) were cut and mounted on glass slides. The slides were dewaxed
in 100% xylol for 30 min and rehydrated in 100, 95 and 85% graded
alcohol solutions. Subsequently, antigen retrieval was performed in
0.01 M sodium citrate solution at 98°C for 15 min. Endogenous
peroxidase activity was blocked with 5% normal goat serum (OriGene
Technologies, Inc.) at 37°C for 1 h. The subcutaneous tumor tissues
were incubated with a proliferating cell nuclear antigen (PCNA)
antibody (1:100; cat. no. 13110; Cell Signaling Technology, Inc.)
and caspase-3 antibody (1:100; cat. no. ab4051; Abcam) at 4°C
overnight. Then, the slides were washed three times with PBS and
incubated with an anti-rabbit lgG HRP-conjugated secondary antibody
(1:10,000; cat. no. 7074; Cell Signaling Technology, Inc.) for 1 h
at room temperature. The slides were then washed three times with
PBS and 50 µl of DAB solution (Beyotime Institute of Biotechnology)
was added for 3–5 min, followed by another wash. Then, the slides
were stained with hematoxylin at room temperature for 5 min,
followed by another wash. Next, 1% hydrochloric acid alcohol
differentiation was used for 2 sec in order to stain the tissue
blue, and then the sections were dehydrated and sealed with neutral
gum after clearing. After drying, images of the slides were
captured using an upright light microscope (×400). A total of 5
different fields of view were taken from each sample using
magnification, ×400, and the number of brown cells in each field of
view was counted. Statistical analyses were performed using SPSS
17.0 software (SPSS, Inc.).
Cell apoptosis and invasion
assays
Apoptosis assays were performed using an Annexin
V-FITC/PI Cell Apoptosis kit (Nanjing KeyGen Biotech, Co., Ltd.).
In total, 100 µl suspension containing 5×105 cells was
incubated with 5 µl Annexin V and 1 µl PI in the dark at room
temperature for 15 min. The apoptotic rate (early + late apoptotic)
was determined by flow cytometry (BD FACSCalibur; BD Biosciences)
and the software used for analysis was FlowJo™ v10.4.0 (FlowJo
LLC).
Cell invasion assays were performed as described in
previous studies (8,9). In total, 1×105 Huh7 or
1×105 HepG2 cells were resuspended in FBS-free culture
and was added to the upper Transwell chamber containing Matrigel
(BD Biosciences), and 900 µl of culture medium with 10% FBS was
added into the lower chamber. The cells were incubated at 37°C for
24 h and then fixed with 4% paraformaldehyde at room temperature
for 10 min. The cells were removed from the upper surface of the
filter membrane, and the invading cells were stained with a 0.1%
crystal violet solution at room temperature for 10–15 min. Images
were captured and the cells were counted using a light microscope
(×200).
Statistical analysis
Data are presented as the mean ± SD of ≥3
independent experiments. Statistical analyses were performed using
SPSS 17.0 software (SPSS, Inc.). Student's t-test was used for the
comparison of parameters between two groups. One-way ANOVA and
Tukey's test were used to compare the parameters among >2
groups. P<0.05 was considered to indicate a statistically
significant difference.
Results
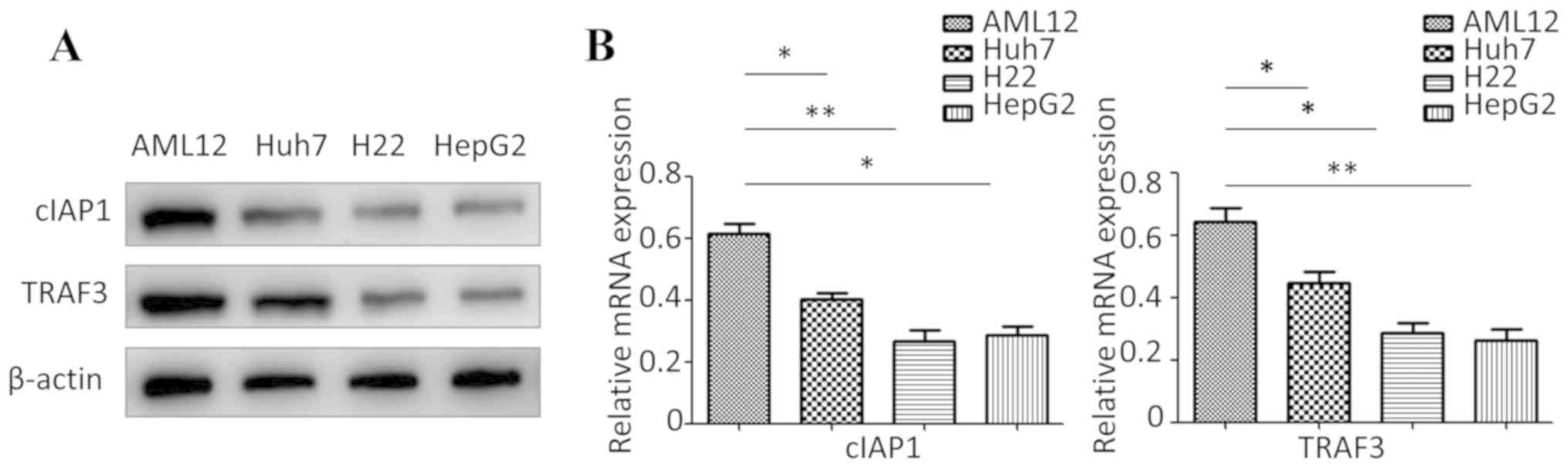
cIAP1 and TRAF3 are expressed at low
levels in Huh7, H22 and HepG2 cells
Western blotting and RT-qPCR were used to detect the
protein and mRNA expression levels of cIAP1 and TRAF3 in Huh7, H22,
HepG2 and AML12 cells. The present results suggested that the
protein and mRNA expression levels of cIAP1 and TRAF3 were
significantly lower in Huh7, H22 and HepG2 cells compared with in
AML12 cells (Fig. 1).
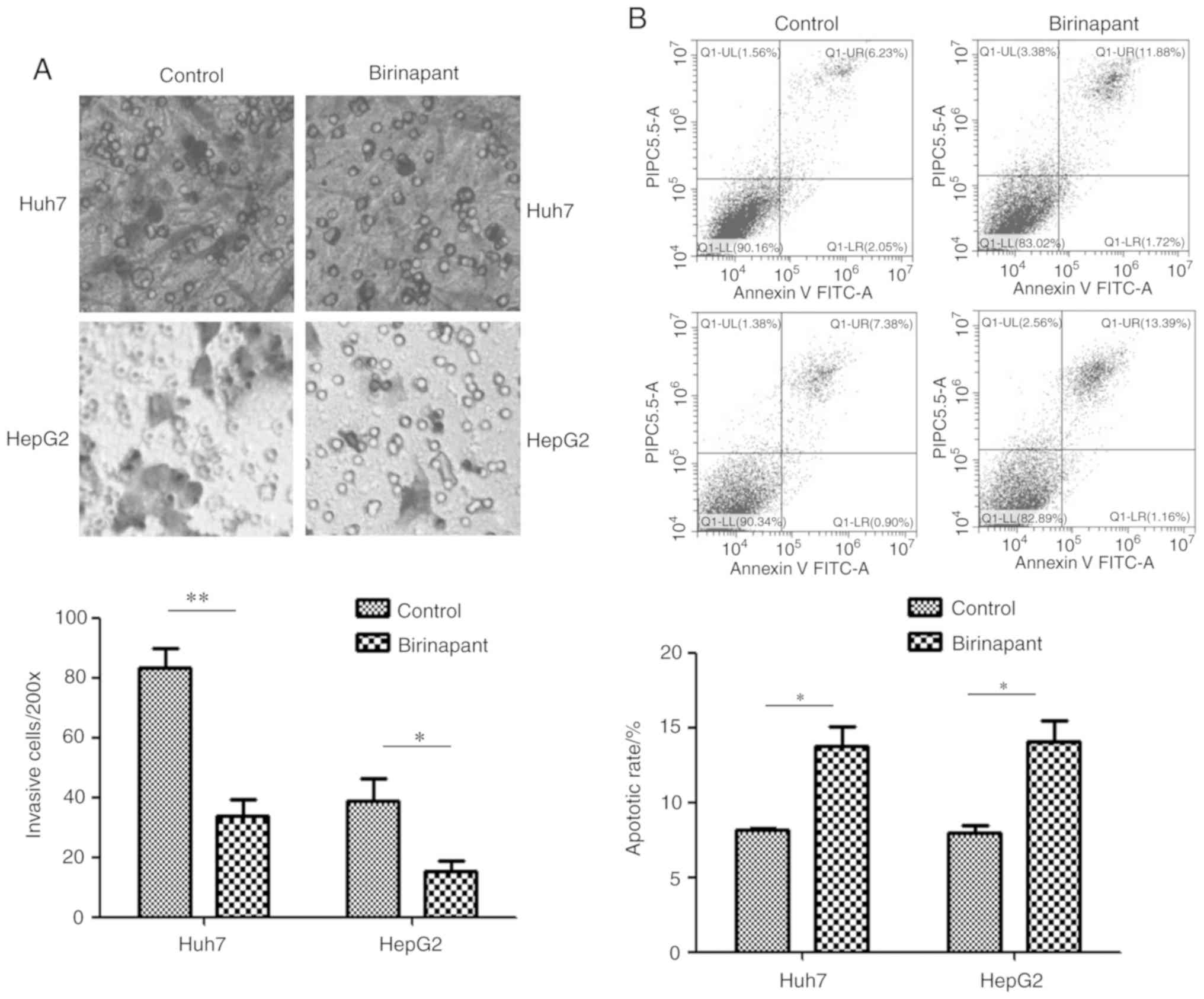
SMAC mimetic birinapant promotes
apoptosis and inhibits invasion of liver cancer cells
The experimental groups used in the present study
included Huh7 and HepG2 cells treated with 300 nmol/ml birinapant
for 24 h, and control cells that did not receive any treatment.
Flow cytometry was used to detect cell apoptosis in the different
groups. Transwell assays were used to detect cell invasion. The
present results suggested that pretreatment with birinapant
promoted apoptosis and inhibited the invasive ability of Huh7 and
HepG2 cells (Fig. 2). H22 cells
were not assessed for cell invasion as they do not adhere to well
walls.
Silencing TRAF3 inhibits
birinapant-mediated apoptosis in liver cancer cells
cIAP1 is the target protein of birinapant, a SMAC
mimetic that regulates the development of several diseases, such as
inflammatory and metabolic diseases, by modulating the cIAP1/TRAF
signaling pathway (6,9,12).
However, whether birinapant-induced apoptosis in liver cancer cells
is related to the cIAP1/TRAF axis is unknown. The present results
suggested that pretreatment with birinapant increased the protein
expression levels of cIAP1 and TRAF3 in Huh7, H22 and HepG2 cells
(Fig. 3A). Therefore, the present
results suggested that birinapant may promote apoptosis in liver
cancer cells by activating the cIAP1/TRAF3 signaling pathway. To
investigate the role of TRAF3 in birinapant-mediated apoptosis in
liver cancer cells, the present study knocked down TRAF3 in liver
cancer cells using a lentivirus (Fig.
3B). There were three different experimental groups: Control
(Con) group, the shRNA-TRAF3 + birinapant (shRNA) group and the
birinapant group. The present results suggested that silencing
TRAF3 expression inhibited birinapant-mediated apoptosis in H22 and
Huh7 cells (Fig. 3C).
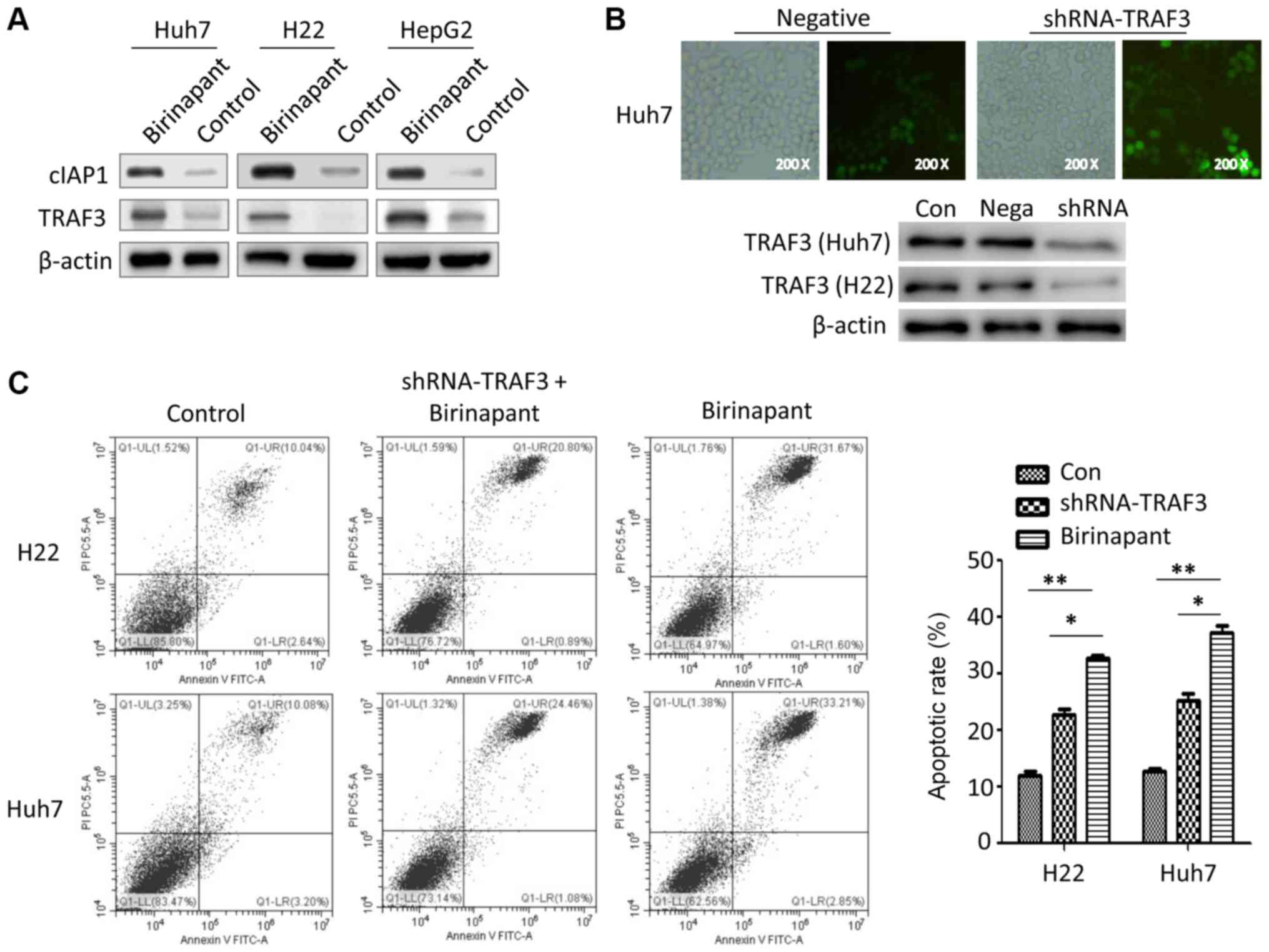 | Figure 3.Role of TRAF3 in birinapant-mediated
apoptosis of liver cancer cells. (A) Western blotting was used to
assess the protein expression levels of cIAP1 and TRAF3 in Huh7,
H22 and HepG2 cells. (B) Green fluorescence (magnification, ×200)
and western blotting were used to investigate the efficiency of
lentivirus-mediated knockdown of TRAF3. (C) Annexin V-FITC
apoptosis assays were used to assess the apoptotic rate in H22 and
Huh7 cells. H22, Control vs. shRNA-TRAF3 + birinapant vs.
birinapant, 12.68 vs. 21.69 vs. 32.27%; Huh7, Control vs.
shRNA-TRAF3 + birinapant vs. birinapant, 13.28 vs. 25.54 vs.
36.06%. *P<0.05, **P<0.01. cIAP1, cellular inhibitor of
apoptosis 1; Con, control; shRNA, short hairpin RNA; Nega, negative
control; TRAF3, tumor necrosis factor receptor-associated factor
3. |
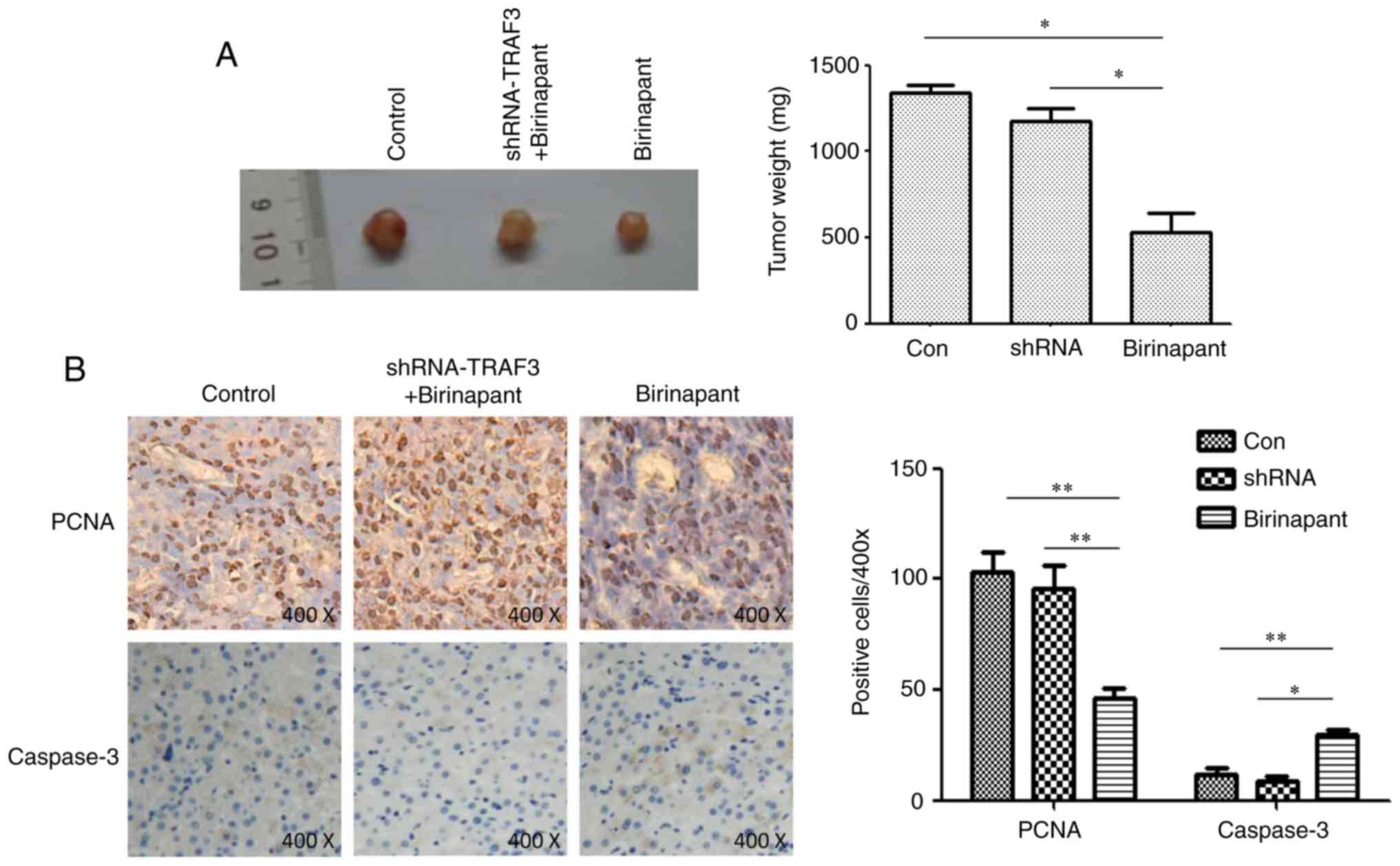
SMAC mimetic birinapant inhibits HCC
growth by activating the cIAP1/TRAF3 signaling pathway
PCNA reflects the proliferative ability of tumor
cells and caspase is a marker of cell apoptosis (16,17).
In the present study, mice were subcutaneously injected with normal
or shRNA-TRAF-infected H22 cells, and treated with saline or
birinapant, and tumor growth was monitored for 15 days.
Immunohistochemistry assays were used to detect the protein
expression levels of PCNA and caspase-3 in the subcutaneous tumors.
Birinapant inhibited the growth of subcutaneous tumors, and
silencing TRAF3 reduced the inhibitory effect of birinapant on
subcutaneous tumor growth (Fig.
4A). The present results suggested that birinapant inhibited
the expression of PCNA and increased the expression of caspase-3,
whereas silencing TRAF3 reversed these effects (Fig. 4B). Collectively, the present
results suggested that the SMAC mimetic birinapant inhibited the
growth of HCC and promoted apoptosis in liver cancer cells,
possibly by activating the cIAP1/TRAF3 signaling pathway.
Discussion
HCC is a serious threat to human health due to a
lack of effective treatments (1,2).
Inducing apoptosis in tumor cells is currently an important
strategy for inhibiting the malignant biological features of cancer
(18,19). SMAC was discovered by Verhagen
and Vaux (20) to localize
in mitochondria and regulate cell apoptosis. SMAC may promote the
apoptosis of tumor cells in several cancer types, including gastric
cancer, ovarian cancer and non-Hodgkin lymphoma (21–23).
This mechanism may be closely related to the fact that upon
stimulation by certain factors, such as interferon and antitumor
drugs, SMAC can be released from the mitochondria into the
cytoplasm to bind cIAPs and inhibit the anti-apoptotic activity of
cIAPs, thus promoting cell apoptosis and further inhibiting tumor
growth (22,23). In addition, SMAC serves an
important role in regulating immunity and inflammation. A previous
in vitro study demonstrated that SMAC inhibits the
LPS-mediated release of inflammatory cytokines from RAW264.7
macrophages by inhibiting the LPS-mediated degradation of TRAF3 and
activation of the MAPK signaling pathway (24).
TRAF3 is expressed by numerous types of cell,
including immune cells such as macrophages, B lymphocytes and T
lymphocytes, and serves important roles in regulating the immune
system (25). TRAF3 functions
mainly via ubiquitination (Ub), including K48-linked Ub and
K63-linked Ub (26). K48
polyubiquitination of TRAF3 induces TRAF3 degradation, which limits
retinoic acid-inducible gene 1-induced type I interferon production
in immune cells (24). TRAF3 is
also subject to post-translational modification with K63-linked
polyubiquitin chains, which is markedly different from K48-linked
polyubiquitination (27). K63
polyubiquitination of TRAF3 does not induce degradation, but
mediates PI3K activation in immune cells (28).
The structure and function of the SMAC protein, and
the application of SMAC mimetics for the treatment of various
tumors, has become a focus in research. SMAC mimetics have been
used for the treatment of several types of cancer, such as breast
cancer, prostate cancer and lung cancer (6,7).
Birinapant, a typical SMAC mimetic, can inhibit the proliferation
of head and neck cancer, myeloma and pancreatic cancer cells
(29,30). However, whether birinapant affects
the growth of HCC and its associated molecular mechanism are still
unknown.
To the best of our knowledge, the present study was
the first to suggest that cIAP1 and TRAF3 were expressed at low
levels in liver cancer cells, and that the SMAC mimetic birinapant
promoted apoptosis and inhibited invasion in liver cancer cells. In
addition, the present results suggested that silencing TRAF3
inhibited birinapant-mediated apoptosis in liver cancer cells and
that birinapant inhibited HCC growth in vivo. Therefore, the
SMAC mimetic birinapant may promote apoptosis, and inhibit the
proliferation and invasion of liver cancer cells. The present
results suggested that the molecular mechanism may be related to
activation of the cIAP1/TRAF3 signaling pathway by birinapant in
liver cancer cells.
Acknowledgements
Not applicable.
Funding
The present study was supported by The Enshi State
Science and Technology Program Project (grant no. 2017-14).
Availability of data and materials
All data generated or analyzed during the present
study are included in this published article.
Authors' contributions
JD and DQ performed most of the experiments and
drafted the manuscript. YZ, QL and YL performed some experiments
and collected the data. JD and JL designed the study. All authors
read and approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by The Research
Ethics Committee of The Central Hospital of Enshi Tujia and Miao
Autonomous Prefecture.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
SMAC
|
second mitochondria-derived activator
of caspase
|
|
HCC
|
hepatocellular carcinoma
|
|
TRAF3
|
tumor necrosis factor
receptor-associated factor 3
|
|
cIAP1
|
cellular inhibitor of apoptosis 1
|
References
|
1
|
Bruix J, Gores GJ and Mazzaferro V:
Hepatocellular carcinoma: Clinical frontiers and perspectives. Gut.
63:844–855. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hartke J, Johnson M and Ghabril M: The
diagnosis and treatment of hepatocellular carcinoma. Semin Diagn
Pathol. 34:153–159. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Boddu P, Carter BZ, Verstovsek S and
Pemmaraju N: SMAC mimetics as potential cancer therapeutics in
myeloid malignancies. Br J Haematol. 185:219–231. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Cossu F, Milani M, Mastrangelo E and Lecis
D: Targeting the BIR domains of inhibitor of apoptosis (IAP)
proteins in cancer treatment. Comput Struct Biotechnol J.
17:142–150. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Pedersen J, LaCasse EC, Seidelin JB,
Coskun M and Nielsen OH: Inhibitors of apoptosis (IAPs) regulate
intestinal immunity and inflammatory bowel disease (IBD)
inflammation. Trends Mol Med. 20:652–665. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chromik J, Safferthal C, Serve H and Fulda
S: Smac mimetic primes apoptosis-resistant acute myeloid leukaemia
cells for cytarabine-induced cell death by triggering necroptosis.
Cancer Lett. 344:101–109. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Nikkhoo A, Rostami N, Hojjat-Farsangi M,
Azizi G, Yousefi B, Ghalamfarsa G and Jadidi-Niaragh F: Smac
mimetics as novel promising modulators of apoptosis in the
treatment of breast cancer. J Cell Biochem. 120:9300–9314. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Abhari BA, McCarthy N, Le Berre M,
Kilcoyne M, Joshi L, Agostinis P and Fulda S: Smac mimetic
suppresses tunicamycin-induced apoptosis via resolution of ER
stress. Cell Death Dis. 10:1552019. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Gerges S, Rohde K and Fulda S: Cotreatment
with Smac mimetics and demethylating agents induces both apoptotic
and necroptotic cell death pathways in acute lymphoblastic leukemia
cells. Cancer Lett. 375:127–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Brumatti G, Ma C, Lalaoui N, Nguyen NY,
Navarro M, Tanzer MC, Richmond J, Ghisi M, Salmon JM, Silke N, et
al: The caspase-8 inhibitor emricasan combines with the SMAC
mimetic birinapant to induce necroptosis and treat acute myeloid
leukemia. Sci Transl Med. 8:339ra692016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Shu X, Zhu Z, Cao D, Zheng L, Wang F, Pei
H, Wen J, Yang J, Li D, Bai P, et al: PEG-derivatized birinapant as
a nanomicellar carrier of paclitaxel delivery for cancer therapy.
Colloids Surf B Biointerfaces. 182:1103562019. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu H, Liao R, He K, Zhu X, Li P and Gong
J: The SMAC mimetic birinapant attenuates
lipopolysaccharide-induced liver injury by inhibiting the tumor
necrosis factor receptor-associated factor 3 degradation in Kupffer
cells. Immunol Lett. 85:79–83. 2017. View Article : Google Scholar
|
|
13
|
Ho PC, Tsui YC, Feng X, Greaves DR and Wei
LN: NF-κB-mediated degradation of the coactivator RIP140 regulates
inflammatory responses and contributes to endotoxin tolerance. Nat
Immunol. 13:379–386. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Liu Z, Mar KB, Hanners NW, Perelman SS,
Kanchwala M, Xing C, Schoggins JW and Alto NM: A NIK-SIX signalling
axis controls inflammation by targeted silencing of non-canonical
NF-κB. Nature. 68:249–253. 2019. View Article : Google Scholar
|
|
16
|
Mi L, Zhou Y, Wu D, Tao Q, Wang X, Zhu H,
Gao X, Wang J, Ling R, Deng J, et al: ACSS2/AMPK/PCNA
pathway-driven proliferation and chemoresistance of esophageal
squamous carcinoma cells under nutrient stress. Mol Med Rep.
20:5286–5296. 2019.PubMed/NCBI
|
|
17
|
Sato Y, Yoshino H, Kazama Y and
Kashiwakura I: Involvement of caspase-8 in apoptosis enhancement by
cotreatment with retinoic acid-inducible gene-I-like receptor
agonist and ionizing radiation in human non-small cell lung cancer.
Mol Med Rep. 18:5286–5294. 2018.PubMed/NCBI
|
|
18
|
Razmkhah M, Abtahi S and Ghaderi A:
Mesenchymal stem cells, immune cells and tumor cells crosstalk: A
sinister triangle in the tumor microenvironment. Curr Stem Cell Res
Ther. 14:43–51. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Jin Y, Meng X, Qiu Z, Su Y, Yu P and Qu P:
Anti-tumor and anti-metastatic roles of cordycepin, one bioactive
compound of Cordyceps militaris. Saudi J Biol Sci.
25:991–995. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Verhagen AM and Vaux DL: Cell death
regulation by the mammalian IAP antagonist Diablo/Smac. Apoptosis.
7:163–166. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Soung YH, Park WS, Nam SW, Lee JY, Yoo NJ
and Lee SH: SMAC/DIABLO mutation is uncommon in gastric and
colorectal carcinomas. Pathology. 38:85–87. 2016. View Article : Google Scholar
|
|
22
|
Cekay MJ, Roesler S, Frank T, Knuth AK,
Eckhardt I and Fulda S: Smac mimetics and type II interferon
synergistically induce necroptosis in various cancer cell lines.
Cancer Lett. 410:228–237. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bhatti IA, Abhari BA and Fulda S:
Identification of a synergistic combination of Smac mimetic and
Bortezomib to trigger cell death in B-cell non-Hodgkin lymphoma
cells. Cancer Lett. 405:63–72. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Tseng PH, Matsuzawa A, Zhang W, Mino T,
Vignali DA and Karin M: Different modes of ubiquitination of the
adaptor TRAF3 selectively activate the expression of type I
interferons and proinflammatory cytokines. Nat Immunol. 11:70–75.
2010. View
Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lin WW, Hostager BS and Bishop GA: TRAF3,
ubiquitination, and B-lymphocyte regulation. Immunol Rev.
266:46–55. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wen J, Bai H, Chen N, Zhang W, Zhu X, Li P
and Gong J: USP25 promotes endotoxin tolerance via suppressing
K48-linked ubiquitination and degradation of TRAF3 in Kupffer
cells. Mol Immunol. 106:53–62. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Guan K, Wei C, Zheng Z, Song T, Wu F,
Zhang Y, Cao Y, Ma S, Chen W, Xu Q, et al: MAVS promotes
inflammasome activation by targeting ASC for K63-linked
ubiquitination via the E3 ligase TRAF3. J Immunol. 194:4880–4890.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Muro I, Fang G, Gardella KA, Mahajan IM
and Wright CW: The TRAF3 adaptor protein drives proliferation of
anaplastic large cell lymphoma cells by regulating multiple
signaling pathways. Cell Cycle. 13:1918–1927. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Eytan DF, Snow GE, Carlson S, Derakhshan
A, Saleh A, Schiltz S, Cheng H, Mohan S, Cornelius S, Coupar J, et
al: SMAC Mimetic birinapant plus radiation eradicates human head
and neck cancers with genomic amplifications of cell death genes
FADD and BIRC2. Cancer Res. 76:5442–5454. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhu X, Shen X, Qu J, Straubinger RM and
Jusko WJ: Multi-scale network model supported by proteomics for
analysis of combined gemcitabine and birinapant effects in
pancreatic cancer cells. CPT Pharmacometrics Syst Pharmacol.
7:549–561. 2018. View Article : Google Scholar : PubMed/NCBI
|