Introduction
Hematologic malignancy is a serious disease that
develops quickly and aggressively, and severely threatens human
health due to its high mortality (1–3). The
incidence of complete remission of hematologic malignancy has
increased recently, with new therapeutic strategies (4). For instance, chimeric antigen
receptor (CAR) T-cells targeting CD19 and therapeutic antibodies
targeting CD20 have been developed and tested in patients with
B-cell lymphoma in preclinical and clinical trials (5). However, there is no effective
treatment for T-cell lymphoma, and the treatment of hematologic
malignancy is relatively limited. Since the 5-year survival rate of
patients with leukemia is between 45–55% (6–8),
developing new therapeutic strategies and finding new agents for
curing hematologic malignancy has become urgent.
Isoferulic acid (IFA), also known as
3-(3-hydroxy-4-methoxyphenyl)-2-propenoic acid, is a natural
compound extracted from Cimicifuga heracleifolia (CH), which
is frequently used in traditional medicine in Asian countries for
treating inflammatory diseases and specific cancers (9,10).
As one of the important active ingredients in CH, IFA has several
therapeutic effects. These include the inhibition of several
inflammatory diseases (11),
elimination of viral infections (12), clearance of reactive oxygen species
(ROS) (13), alleviation of
metabolic diseases (14) and the
reduction of glucose-induced glycation of bovine serum albumin
(11,15). Although IFA affects cell cycle
arrest (16), inhibits tumor cell
proliferation and prompts cell apoptosis (17–19),
whether it inhibits leukemia cells remains to be clarified. In
vitro and in vivo experiments should be carried out to
show whether IFA could become a potential candidate for treating
leukemia.
Leukemia is a hematologic malignancy that generally
originates in the bone marrow, and develops numerous abnormal
leukocytes (20). Abnormal
undifferentiated leukocytes dramatically proliferate, expand and
resist cell apoptosis, resulting in immature cells in the bone
marrow and peripheral blood (21).
Inhibition of tumor cell growth and promotion of cell apoptosis are
two frequent intervention strategies for eliminating cancer cells
(22). Protein kinase B (Akt), a
main downstream signal of PI3K, is an important protein in
promoting cell proliferation, differentiation, migration and
angiogenesis, while also protecting cancer cells against apoptosis
(23–25). Activated Akt promotes cell
proliferation by activating ribosomal protein S6 kinase and
eukaryotic initiation factor 4E (26). It also modulates the cell cycle and
drives the cells to go through both G1/S and G2/M cell cycle
checkpoints (27). Cyclin B-Cdc2
(also known as Cdk1) is an important complex for the regulation of
G2/M transition; it is negatively modulated by Wee1 and myelin
transcription factor 1, and positively regulated by Cdc25B. Both
modulatory cell signaling pathways are precisely controlled by Akt
(28–30). Therefore, interventions that target
Akt-mediated cell signals may be able to inhibit cancer.
In the present study, IFA was found to inhibit cell
growth and promote cell apoptosis in Jurkat, K562 and Raji cell
lines. Leukemia cells were significantly arrested in G2/M phase,
due to the increased phosphorylation of Cdc2 and reduced expression
of Cyclin B1 after treatment with IFA. Furthermore, the latter was
identified to attenuate the phosphorylation of mTOR and Akt. The
results indicated that IFA has an impact on leukemia in
vitro and may be a promising candidate for treating hematologic
malignancy.
Materials and methods
Reagents and antibodies
IFA was ordered from TargetMol. Cell Counting Kit-8
(CCK-8) and trypan blue staining cell viability assay kits were
ordered from Beyotime Institute of Biotechnology. An Annexin
V-FITC/propidium iodide (PI) apoptosis detection kit was purchased
from BestBio Biotechnology. Cleaved poly(ADP-ribose) polymerase
(PARP cat. no. 5625), cleaved caspase-3 (cat. no. 9661), b-actin
(cat. no. 3700), phosphorylated (p)-Cdc2 (Tyr15) (cat. no. 4539),
total-Cdc2 (cat. no. 9116), Cyclin B1 (cat. no. 12231), p-Akt
(Thr308) (cat. no. 13038), total-Akt (cat. no. 4685), p-mTOR
(Ser2448) (cat. no. 5536) and total-mTOR (cat. no. 2983) were
ordered from Cell Signaling Technology, Inc. Horseradish peroxidase
(HRP)-conjugated anti-mouse/rabbit IgG antibody was ordered from
Jackson ImmunoResearch (cat. no. 111-035-003). Other chemical
reagents were purchased from Sigma-Aldrich; Merck KGaA.
Cells and cell culture
Jurkat (acute lymphoid leukemic T cells), K562
(chronic myeloid leukemia), and Raji (Burkitt's lymphoma) cells
were purchased from American Type Culture Collection and maintained
in RPMI-1640 medium with 10% FBS (both Gibco; Thermo Fisher
Scientific, Inc.) at 37°C in a humidified incubator containing 5%
CO2.
Cell viability assay
CCK-8 assay was applied to detect the cell
viability. Briefly, cells were seeded into 96-well plates at 2×104
cells/well for 24 h. IFA at 5, 15 and 45 µM was added for 12, 24
and 48 h. CCK-8 (10 µl) was added and the absorbance at 450 nm was
measured after incubation for 2 h. In addition, the trypan blue
staining cell viability assay kit was used to detect cell
proliferation. Raji, K562 and Jurkat cells were planted into 10-cm
dishes at 1×106 cells/dish. After cell culture for 5 days, at the
point of cell treatment, the cells were collected, stained with
trypan blue within 2 min and counted by a hemocytometer at room
temperature. The IFA-untreated cell group was normalized to 100%
cell viability.
Cell cycle assay
Cell cycle was determined by using PI staining.
Briefly, cells were seeded at a density of 5×105
cells/ml in 12-well plates for 12 h. Then, cells were treated with
IFA at 5, 15 and 45 µM. After incubation for 24 h, cells were
collected and fixed with 70% ethanol overnight at 4°C. After
removing ethanol and neutralizing RNA, PI was used to stain DNA at
4°C. Afterwards, flow cytometry was used to analyze cell cycle
distribution (FACSCalibur, BD Biosciences; FlowJo 7.6, FlowJo
LLC).
Cell apoptosis analysis
Annexin V-FITC/PI analysis (Qiaoxin) was applied to
detect cell apoptosis. Briefly, cells were seeded onto 12-well
plates at 5×105 cells overnight. After treatment with 0,
5, 15 and 45 µM of IFA for 24 h, cells were harvested and rinsed
with PBS. Then, cells were resuspended with binding buffer. The
Annexin V-FITC and PI were incubated with cells for 15 min and 5
min at 4°C, respectively. Finally, flow cytometry was used to
analyze the percentages of apoptotic cells (FACSCalibur; BD
Biosciences; FlowJo 7.6, FlowJo LLC).
Western blot analysis
Cells were treated with IFA at 5, 15 and 45 µM for
24 h. Then, cells were lysed with RIPA Lysis Buffer and the protein
concentration were determined by BCA protein assay and the protein
lysis mixed with loading control (all from Beyotime Institute of
Biotechnology) and heated at 100°C for 5 min. Samples (20 µg) were
loaded on 10% of SDS-PAGE gels and transferred to nitrocellulose
membranes. Before incubation with different antibodies (1:1,000)
overnight at 4°C, the membranes were blocked with 5 % non-fat milk
for 2 h at room temperature. After being washed with TBS-Tween 20
(TBST containing 0.1% Tween 20) five times every 5 min, the
membranes were incubated with HRP-conjugated secondary antibodies
for another 2 h at room temperature and then washed with TBST three
times. The protein bands were visualized using chemiluminescent
substrate reagent (Shanghai Shenger Biotechnology Co., Ltd.). The
semi-quantitative analysis was performed by using ImageJ software
(v1.48U; National Institute of Health).
Statistical analysis
The data were presented as means ± SD based on at
least three independent experiments. The statistical analysis was
performed using Prism 5 (GraphPad Software, Inc). One-way ANOVA
followed by Bonferroni post hoc test were used to determine the
difference between groups. P<0.05 was considered a statistically
significant difference.
Results
IFA displays anti-proliferation
activity in Raji, K562 and Jurkat cell lines
To determine the effect of IFA on proliferation of
leukemia cells, Raji, K562 and Jurkat cells were treated with IFA
for 24 h with 0, 5, 15 and 45 µM, and cell viability was
investigated using CCK-8 and trypan blue staining assays. As
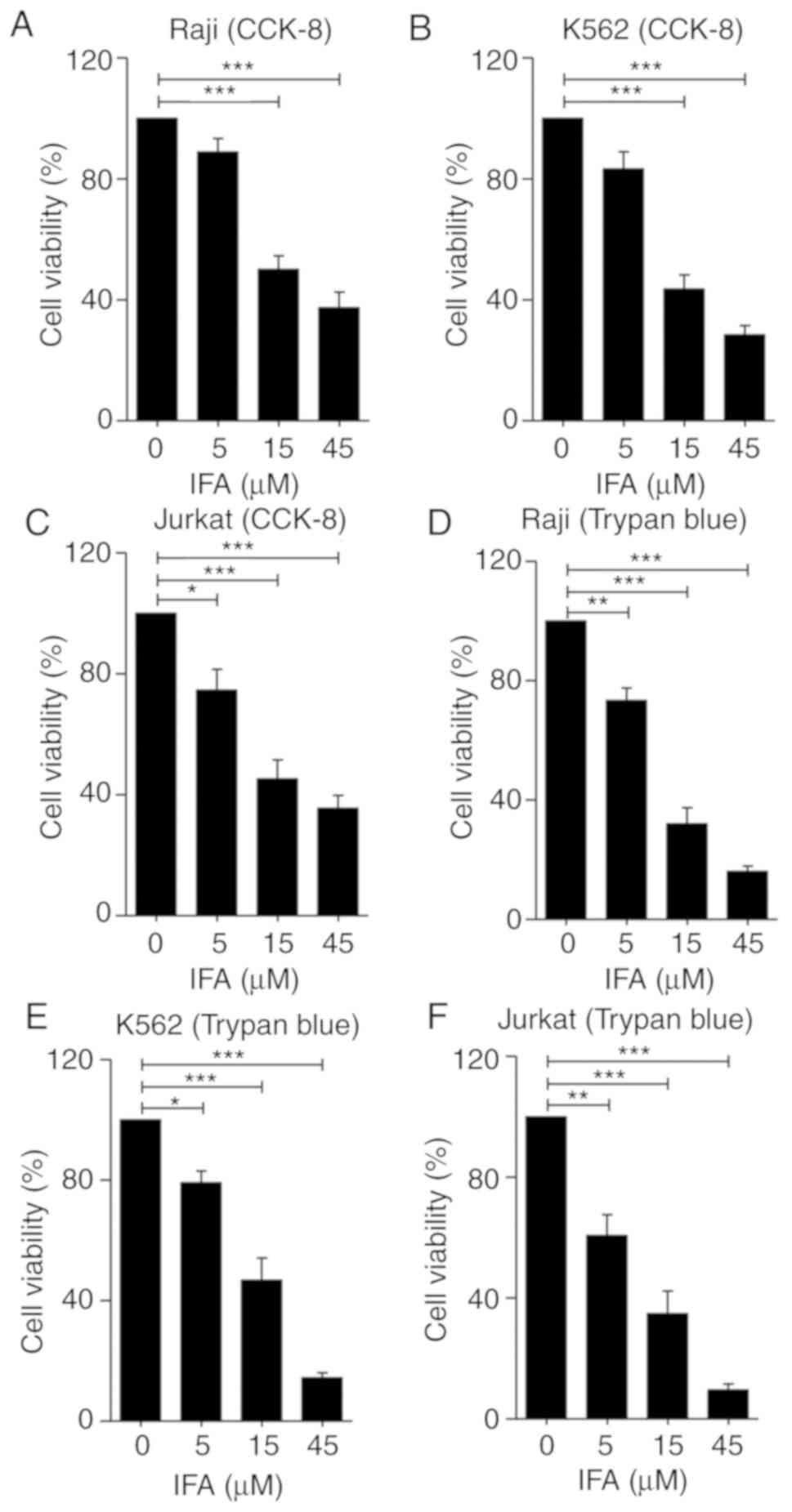
presented in Fig. 1A, IFA showed a
significant inhibitory effect on Raji cell growth in a
dose-dependent manner. After 24 h, the viable cells decreased by
30(±7)%-65(±9)% in the presence of 5–45 µM, compared with that of
the control group. Furthermore, similar inhibitory effects of IFA
on K562 (Fig. 1B) and Jurkat cells
(Fig. 1C) were also detected. To
further confirm the anti-proliferative effect of IFA on leukemia
cells, a trypan blue staining cell viability assay kit was used to
detect the amount of live cells. Raji, K562 and Jurkat cells were
incubated with IFA at 0, 5, 15 and 45 µM for 5 days. The total
amount of cells was calculated and the percentage of cell viability
was presented in Fig. 1D-F.
Similarly, IFA dose-dependently inhibited the proliferation of
Raji, K562 and Jurkat cells (Fig.
1D-F). These results indicated that IFA inhibited leukemia cell
proliferation.
IFA induces cell cycle arrest at G2/M
of Raji, K562 and Jurkat cells
The effect of IFA on cell cycle arrest in Raji, K562
and Jurkat cells was measured by flow cytometry. Raji, K562 and
Jurkat cells were treated with IFA at 0, 5, 15 and 45 µM for 24 h.
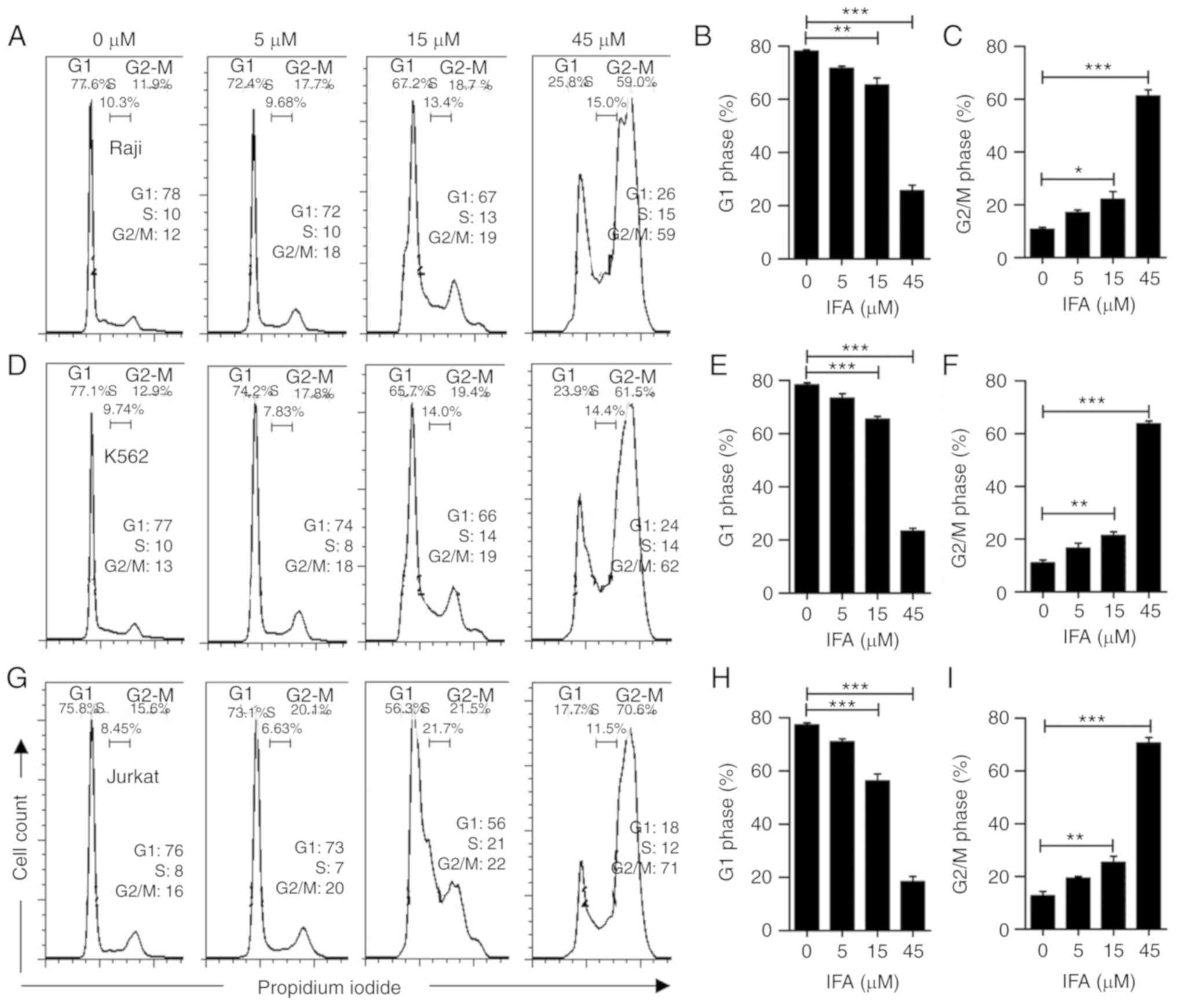
As presented in Fig. 2A, IFA
treatment increased the percentage of Raji cells in G2/M phase in a
dose-dependent manner, and decreased the proportions of Raji cells
in G1 and S phase. Meanwhile, the percentage in G2/M phase
increased from 12% to 59% upon treatment with 45 µM of IFA.
Fig. 2B and C show the
quantification of cell arrest at G1, S and G2/M. It was also found
that IFA had a similar effect on K562 (Fig. 2D-F) and Jurkat cells (Fig. 2G-I). These results revealed that
IFA induced cell cycle arrest at G2/M phase in Raji, K562 and
Jurkat cells.
IFA enhances apoptosis of Raji, K562
and Jurkat cells
It was further determined that significant G2/M cell
cycle arrest led to cell apoptosis in Raji, K562 and Jurkat cells.
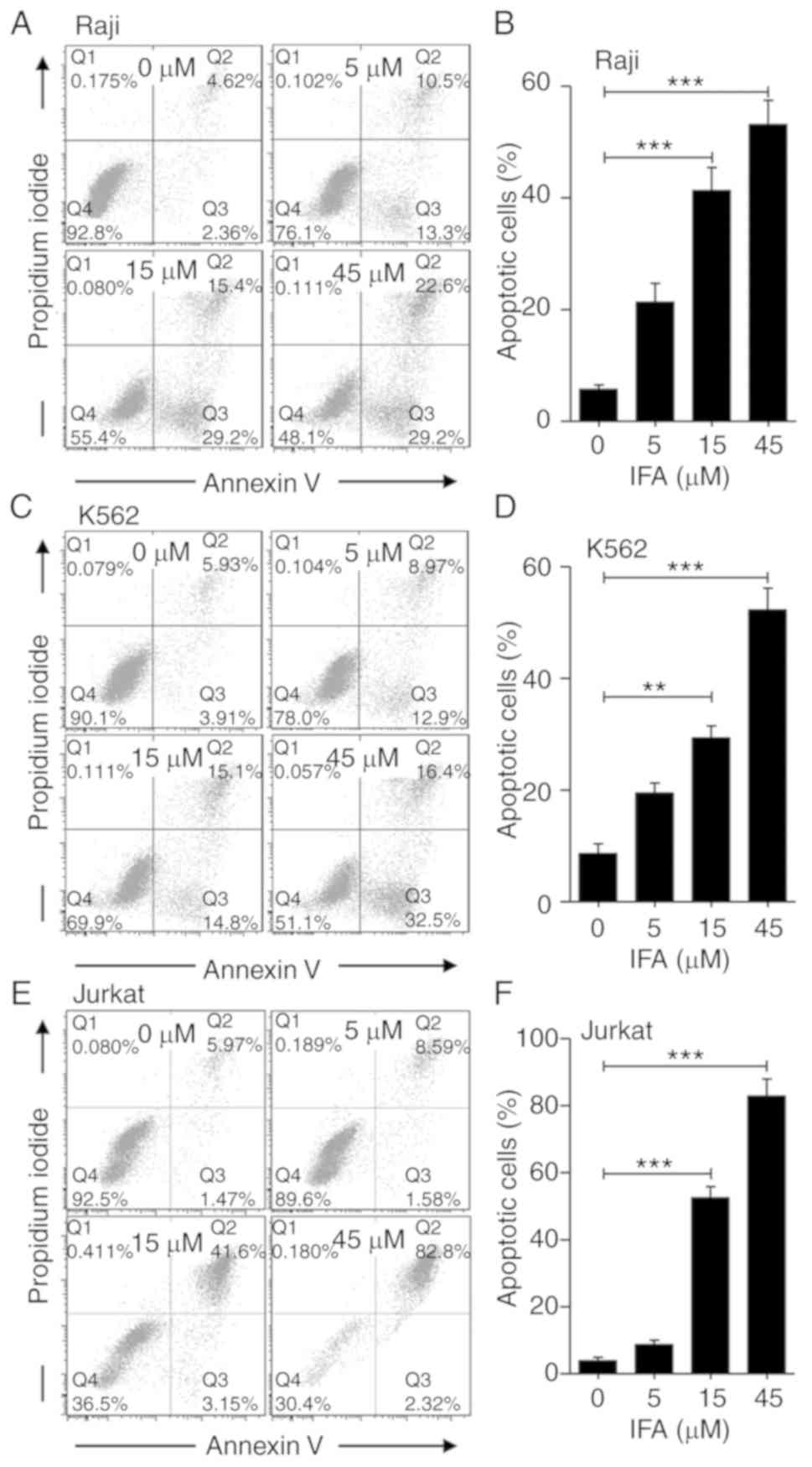
As indicated in Fig. 3, using
Annexin V/PI staining showed that IFA significantly triggered cell
apoptosis in a dose-dependent manner in Raji, K562 and Jurkat
cells,. The proportion of apoptotic cells increased from 7% to 24,
44 and 52% (Fig. 3A and B), from
10% to 22, 30 and 49% (Fig. 3C and
D), and from 7% to 11, 45 and 85% (Fig. 3E and F) after treatment with IFA
for 24 h at 5, 15 and 45 µM in Raji, K562 and Jurkat cells,
respectively. These results suggested that IFA could promote the
apoptosis of leukemia cells compared with control treatment.
IFA induces the expression of
apoptotic proteins
Given that the mitochondrial-related apoptotic
pathway is one of the main pathways of apoptosis (31), the effect of IFA was detected on
the expression of cleaved PARP and cleaved caspase-3, two major
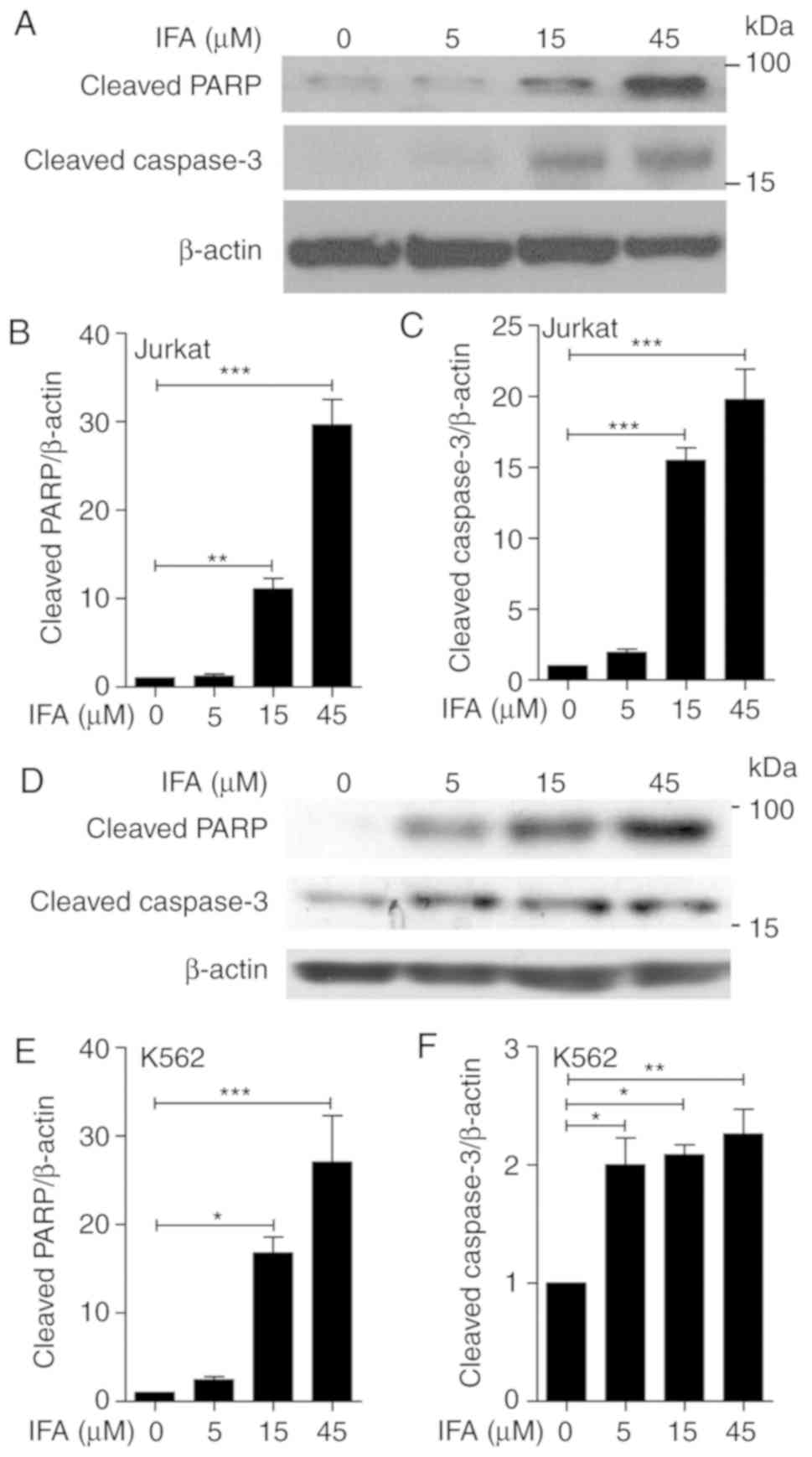
mitochondrial apoptosis-associated proteins. Because IFA displayed
more of an inhibitory effect on Jurkat cells, Jurkat cells were
treated with IFA at 5, 15 and 45 µM for 24 h (Fig. 4A). The expression of cleaved PARP
and cleaved caspase-3 dramatically augmented after incubation with
the indicated concentrations of IFA. Statistical analysis indicated
a dose-dependent manner in IFA-induced expression of cleaved PARP
and cleaved caspase-3 (Fig. 4B and
C). To further confirm the effect of IFA on cell apoptosis,
K562 cells were treated with IFA at 5, 15 and 45 µM for 24 h; in
these cells the cleavage of PARP and cleavage of caspase-3 was
detected (Fig. 4D). Similarly, IFA
dose-dependently increased cleaved PARP and cleaved caspase-3
levels (Fig. 4E and F). These data
showed that IFA enhanced cell apoptosis by inducing the cleavage of
PARP and caspase-3.
IFA modulates the phosphorylation of
Cdc2 and expression of Cyclin B1
To further verify the effect of IFA-induced cell
cycle arrest, the expression of cell cycle associated protein Cdc2
and Cyclin B1 were detected. In Fig.
5A, there was a significant increase in Cdc2 phosphorylation
(Tyr15) and a decrease in the expression of Cyclin B1 after IFA
treatment, compared with the control group in Jurkat cells.
Fig. 5B and C showed that IFA
significantly induced the phosphorylation of Cdc2 and attenuated
the expression of Cyclin B1 in a dose-dependent manner. These
results indicated that G2/M phase arrest induced by IFA was
associated with increased phosphorylation of Cdc2 and the reduction
of Cyclin B1 in Jurkat cells.
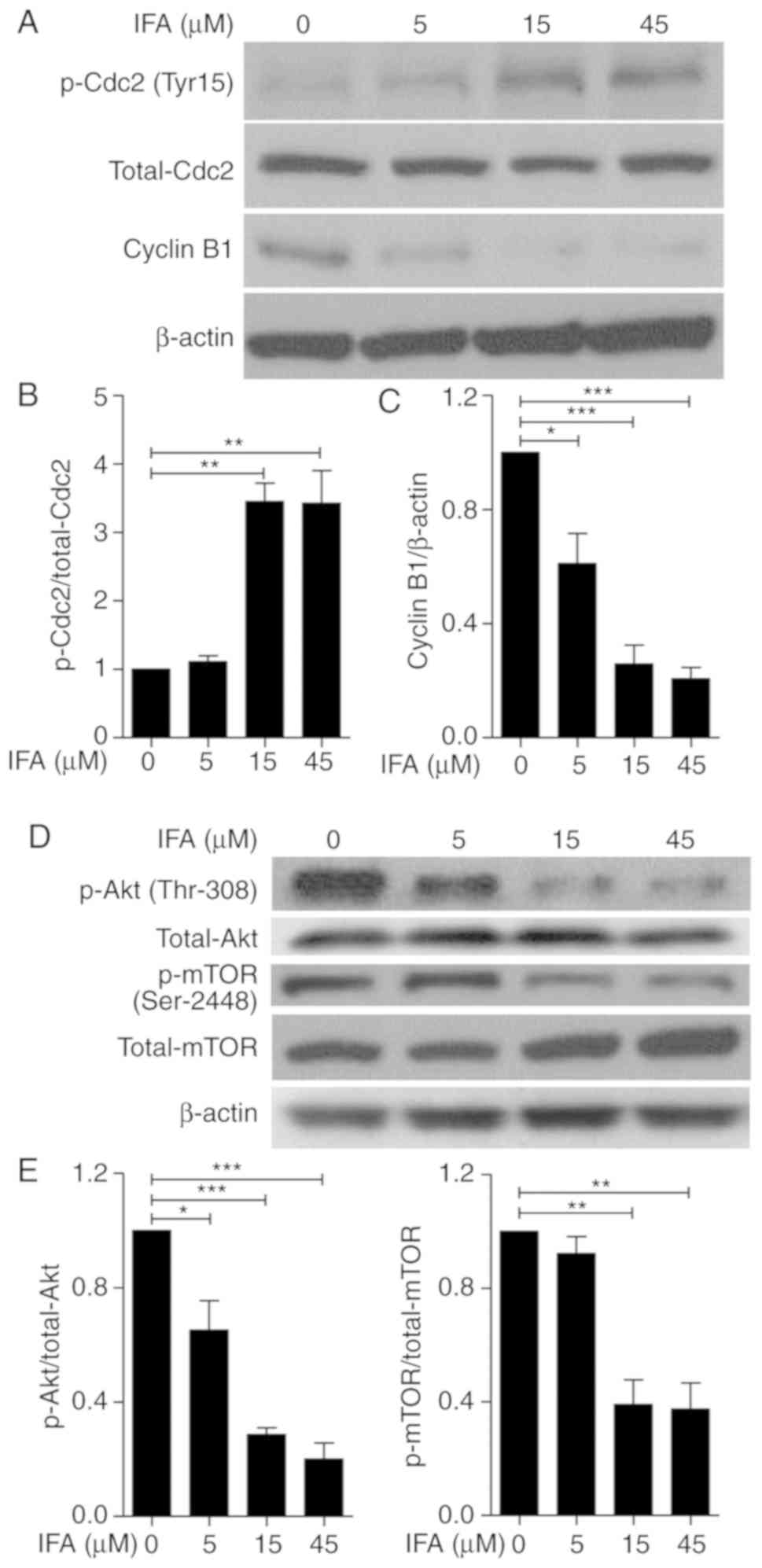 | Figure 5.IFA induces the phosphorylation of
Cdc2, and reduces the expression of Cyclin B1, as well as
phosphorylation of Akt and mTOR in Jurkat cells. Jurkat cells were
incubated with 5, 15 and 45 µM of IFA for 24 h. (A) Phosphorylation
of Cdc2, the expression of Cyclin B1, and (D) the phosphorylation
of Akt and mTOR were measured by western blotting. (B) The ratios
of phosphorylation of Cdc2 to total Cdc2, (C) the expression of
Cyclin B to β-actin, (E) phosphorylation of Akt to total Akt, and
(F) phosphorylation of mTOR to total mTOR were quantified using
ImageJ software. The expression of β-actin was detected as loading
control. Data are expressed as mean ± SD of three independent
experiments. *P<0.05, **P<0.01, ***P<0.001 vs. control
group. IFA, isoferulic acid; p, phosphorylated. |
IFA attenuates Akt/mTOR signaling
Since the Akt/mTOR signaling pathway plays a crucial
role in cancer cell survival, it was further explored whether IFA
induced apoptosis and G2/M phase arrest via the modulation of
Akt/mTOR signaling (28,29). Jurkat cells were treated with the
indicated concentrations of IFA for 24 h. Treatment of IFA
remarkably suppressed the phosphorylation of both Akt (Thr308) and
mTOR (Ser2448; Fig. 5D).
Additionally, a dose-dependent manner was found in the
phosphorylation of Akt and mTOR after IFA treatment (Fig. 5E and F). These results indicated
that IFA inhibited the Akt/mTOR signaling pathway in Jurkat
cells.
Discussion
The presence of IFA leads to the dose-dependent
inhibition of Raji, K562, and Jurkat cell proliferation.
Microtubules are considered to be important targets for cancer
treatment, as disruption of microtubule dynamics would result in
cell cycle arrest, followed by cell apoptosis (32). Paclitaxel is an effective
chemotherapy drug that could inhibit microtubule formation,
resulting in G2/M cell cycle arrest (33). Cell cycle analysis demonstrated
that the induction of G2/M phase arrest by IFA was concomitant with
a decrease in G0/G1 and S phase in Raji, K562 and Jurkat cells. As
is well known, activation of Cdc2, driven by Cyclin B1 binding and
dephosphorylation at Tyr15 by Cdc25C, is an important stage in G2/M
phase transition. Cyclin B1 accumulation is regarded as a marker of
G2/M phase arrest, since it generally accumulates in G2 phase,
reaches its peak at metaphase of cell division and decreases during
anaphase (34). These results
indicated that IFA could induce G2/M phase arrest in Jurkat cells
through Cyclin B1 accumulation and the inhibition of Cdc2
phosphorylation.
It is commonly known that programmed cell death can
regulate cell survival/death balance through blockade of the cell
cycle (35). A number of
chemotherapeutic agents, such as doxorubicin, cisplatin and
etoposide, have been shown to promote cell apoptosis by causing
cell cycle arrest (36). In this
study, IFA was shown to be capable of substantially increasing the
level of apoptosis in Raji, K562 and Jurkat cells. In general, cell
apoptosis can be activated through two signaling pathways, namely
the mitochondrial pathway and the death receptor pathway (37). The mitochondrial pathway (also
called the intrinsic pathway), which is mainly regulated by the
Bcl-2 family, is identified as the leading apoptotic signaling
pathway (36). The mitochondrial
membrane potential decreases following a decrease in the
intracellular Bcl-2/Bax ratio and an increase in the permeability
of the outer mitochondrial membrane, resulting in the release of
cytochrome C (38). The released
cytochrome C in the cytosol can trigger the caspase cascade, after
which PARP is cleaved as a substrate by caspase-3, leading to cell
apoptosis (39). In this study the
apoptosis evoked by IFA was shown to be associated with the
upregulation of cleaved PARP and cleaved caspase-3. This suggested
that IFA could induce apoptosis through the activation of
mitochondria-associated intrinsic apoptosis signaling. Future
studies will be aimed to determine how IFA modulates
mitochondria-mediated apoptosis and which molecule is targeted by
IFA.
Akt/mTOR signaling plays a vital role in the
regulation of tumor cell survival, growth and apoptosis (23,24).
Akt activation could prevent the release of apoptosis-stimulating
factors from mitochondria by promoting the activation of
antiapoptotic proteins (such as Bcl-2) and inhibiting some
proapoptotic proteins (such as Bax and caspase) (40). Rapamycin analogs (rapalogs), the
first generation of allosteric mTORC1 inhibitors, selectively bind
to FK506 binding protein 12 of mTORC1 and inhibit mTOR signals
(41). Rapalogs could reduce the
proliferation of acute myeloid leukemia (AML) cells and
clonogenicity of leukemic progenitor cells in preclinical and
clinical settings (42). However,
their effects were mainly cytostatic, partially affecting
apoptosis, as confirmed in phase 1 and 2 clinical studies (43). The treatment of AML cells with a
combination of PIM inhibitor AZD1897 and Akt inhibitor AZD5363
augmented the blockade of the mTOR axis and led to the induction of
apoptosis (44). Since IFA had an
impact on the inhibition of phosphorylation in the Akt/mTOR signal
pathway, the combination of IFA and Akt inhibitors may be more
effective for treatment of leukemia.
Furthermore, IFA displays various biological
functions potentially beneficial in the treatment of various
different diseases. Pretreatment with IFA could attenuate
methylglyoxal (MG)-induced dysfunction and apoptosis in INS-1
pancreatic β-cells through the mitochondrial survival pathway and
the upregulation of glyoxalase 1 activity (16). IFA derivatives also display varying
degrees of anticancer potency, with some exhibiting excellent
histone deacetylase inhibitory activity (45). IFA could prevent DNA damage and
MG-induced protein glycation through free radical scavenging
activity (46). Dilshara et
al (9) demonstrated that IFA
suppresses the production of not only nitric oxide and
prostaglandin E2, but also their regulatory genes in
lipopolysaccharide-stimulated BV2 microglial cells by inhibiting
PI3K/Akt-dependent NF-κB activity and enhancing nuclear-related
factor 2-mediated heme oxygenase-1 expression. In addition, IFA may
play an important role in preventing diabetic complications by
inhibiting advanced glycation end product formation and
oxidation-dependent protein damage (15). The present findings indicated that
IFA can induce leukemia cell apoptosis and initiate apoptotic
signal pathways, thus suggesting the possibility of IFA having
different targets in different diseases.
In the present study, IFA inhibited p-Akt and
p-mTOR, suggesting that it might inhibit Raji, K562 and Jurkat cell
proliferation and induce cell apoptosis via attenuation of Akt/mTOR
signaling pathway. Taken together, this study indicated the
therapeutic effect of IFA on Raji, K562 and Jurkat cells, thereby
suggesting it as a promising candidate for the treatment of
hematologic malignancy. Conduction of in vivo experiments
will be the next step to determine whether IFA can inhibit leukemia
cell proliferation in mouse models, and whether it can induce
apoptosis in vivo, so as to accentuate the possibility of
clinical application of IFA.
Acknowledgements
Not applicable.
Funding
The present study was funded by Research Grant for
Health Science and Technology of Pudong Municipal Commission of
Health and family Planning of Shanghai (grant no. PW2019A-6),
Shanghai Pudong Hospital Introducing Talent Research Starting Fund
(grant no. 2015YJ-03) and Scientific Research Fund of Shanghai
Pudong Hospital (grant no. 201503).
Availability of data and materials
The analyzed datasets generated during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
ZL and HZ designed research and revised the
manuscript; GF and NZ performed the research analysis and wrote the
manuscript; ZL GF NZ LW and HZ analyzed the data. All authors read
and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that there is no competing
interests.
References
|
1
|
Durinck K, Goossens S, Peirs S, Wallaert
A, Van Loocke W, Matthijssens F, Pieters T, Milani G, Lammens T,
Rondou P, et al: Novel biological insights in T-cell acute
lymphoblastic leukemia. Exp Hematol. 43:625–639. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Egler RA, Ahuja SP and Matloub Y:
L-asparaginase in the treatment of patients with acute
lymphoblastic leukemia. J Pharmacol Pharmacother. 7:62–71. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Liu XY, Yang YF, Wu CT, Xiao FJ, Zhang QW,
Ma XN, Li QF, Yan J, Wang H and Wang LS: Spred2 is involved in
imatinib-induced cytotoxicity in chronic myeloid leukemia cells.
Biochem Biophys Res Commun. 393:637–642. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Nilsson C, Hulegårdh E, Garelius H,
Möllgård L, Brune M, Wahlin A, Lenhoff S, Frödin U, Remberger M,
Höglund M, et al: Secondary acute myeloid leukemia and the role of
allogeneic stem cell transplantation in a population-based setting.
Biol Blood Marrow Transplant. 25:1770–1778. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Han C and Kwon BS: Chimeric antigen
receptor T-cell therapy for cancer: A basic research-oriented
perspective. Immunotherapy. 10:221–234. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Pui CH, Relling MV and Downing JR: Acute
lymphoblastic leukemia. N Engl J Med. 350:1535–1548. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Faderl S, O'Brien S, Pui CH, Stock W,
Wetzler M, Hoelzer D and Kantarjian HM: Adult acute lymphoblastic
leukemia: Concepts and strategies. Cancer. 116:1165–1176. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Cheng W, Zheng T, Wang Y, Cai K, Wu W,
Zhao T and Xu R: Activation of Notch1 signaling by HTLV-1 Tax
promotes proliferation of adult T-cell leukemia cells. Biochem
Biophys Res Commun. 512:598–603. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Dilshara MG, Lee KT, Jayasooriya RG, Kang
CH, Park SR, Choi YH, Choi IW, Hyun JW, Chang WY, Kim YS, et al:
Downregulation of NO and PGE2 in LPS-stimulated BV2 microglial
cells by trans-isoferulic acid via suppression of
PI3K/Akt-dependent NF-κB and activation of Nrf2-mediated HO-1. Int
Immunopharmacol. 18:203–211. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Thiyagarajan G, Muthukumaran P, Sarath
Kumar B, Muthusamy VS and Lakshmi BS: Selective inhibition of PTP1B
by vitalboside a from syzygium cumini enhances insulin sensitivity
and attenuates lipid accumulation via partial agonism to PPARγ: In
vitro and in silico investigation. Chem Biol Drug Des. 88:302–312.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Jairajpuri DS and Jairajpuri ZS:
Isoferulic acid action against glycation-induced changes in
structural and functional attributes of human high-density
lipoprotein. Biochemistry (Mosc). 81:289–295. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sakai S, Kawamata H, Kogure T, Mantani N,
Terasawa K, Umatake M and Ochiai H: Inhibitory effect of ferulic
acid and isoferulic acid on the production of macrophage
inflammatory protein-2 in response to respiratory syncytial virus
infection in RAW264.7 cells. Mediators Inflamm. 8:173–175. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang X, Li X and Chen D: Evaluation of
antioxidant activity of isoferulic acid in vitro. Nat Prod Commun.
6:1285–1288. 2011.PubMed/NCBI
|
|
14
|
Liu IM, Hsu FL, Chen CF and Cheng JT:
Antihyperglycemic action of isoferulic acid in
streptozotocin-induced diabetic rats. Br J Pharmacol. 129:631–636.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Meeprom A, Sompong W, Chan CB and
Adisakwattana S: Isoferulic acid, a new anti-glycation agent,
inhibits fructose- and glucose-mediated protein glycation in vitro.
Molecules. 18:6439–6454. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Meeprom A, Chan CB, Sompong W and
Adisakwattana S: Isoferulic acid attenuates methylglyoxal-induced
apoptosis in INS-1 rat pancreatic β-cell through mitochondrial
survival pathways and increasing glyoxalase-1 activity. Biomed
Pharmacother. 101:777–785. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Nuntanakorn P, Jiang B, Einbond LS, Yang
H, Kronenberg F, Weinstein IB and Kennelly EJ: Polyphenolic
constituents of Actaea racemosa. J Nat Prod. 69:314–318. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kim JY, Kim HY, Jeon JY, Kim DM, Zhou Y,
Lee JS, Lee H and Choi HK: Effects of coronatine elicitation on
growth and metabolic profiles of Lemna paucicostata culture. PLoS
One. 12:e01876222017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Xuan H, Wang Y, Li A, Fu C, Wang Y and
Peng W: Bioactive components of chinese propolis water extract on
antitumor activity and quality control. Evid Based Complement
Alternat Med. 2016:96419652016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Slavin S, Nagler A, Naparstek E,
Kapelushnik Y, Aker M, Cividalli G, Varadi G, Kirschbaum M,
Ackerstein A, Samuel S, et al: Nonmyeloablative stem cell
transplantation and cell therapy as an alternative to conventional
bone marrow transplantation with lethal cytoreduction for the
treatment of malignant and nonmalignant hematologic diseases.
Blood. 91:756–763. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ju J: An increased proportion of apoptosis
in CD4+ T lymphocytes isolated from the peripheral blood
in patients with stable chronic obstructive pulmonary disease.
Tuberc Respir Dis (Seoul). 81:132–137. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Fesik SW: Promoting apoptosis as a
strategy for cancer drug discovery. Nat Rev Cancer. 5:876–885.
2005. View
Article : Google Scholar : PubMed/NCBI
|
|
23
|
Saxena NK, Sharma D, Ding X, Lin S, Marra
F, Merlin D and Anania FA: Concomitant activation of the JAK/STAT,
PI3K/AKT, and ERK signaling is involved in leptin-mediated
promotion of invasion and Migration of hepatocellular carcinoma
cells. Cancer Res. 67:2497–2507. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang Y, Wang SJ, Han ZH, Li YQ, Xue JH,
Gao DF, Wu XS and Wang CX: PI3K/AKT signaling pathway plays a role
in enhancement of eNOS activity by recombinant human angiotensin
converting enzyme 2 in human umbilical vein endothelial cells. Int
J Clin Exp Pathol. 7:8112–8117. 2014.PubMed/NCBI
|
|
25
|
Ahmed MS, El-Senduny F, Taylor J and
Halaweish FT: Biological screening of cucurbitacin inspired estrone
analogs targeting mitogen-activated protein kinase (MAPK) pathway.
Chem Biol Drug Des. 90:478–484. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Aoki M and Fujishita T: Oncogenic roles of
the PI3K/AKT/mTOR axis. Curr Top Microbiol Immunol. 407:153–189.
2017.PubMed/NCBI
|
|
27
|
Massagué J: G1 cell-cycle control and
cancer. Nature. 432:298–306. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chen D, Lin X, Zhang C, Liu Z, Chen Z, Li
Z, Wang J, Li B, Hu Y, Dong B, et al: Dual PI3K/mTOR inhibitor
BEZ235 as a promising therapeutic strategy against
paclitaxel-resistant gastric cancer via targeting PI3K/Akt/mTOR
pathway. Cell Death Dis. 9:1232018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Shimizu Y, Segawa T, Inoue T, Shiraishi T,
Yoshida T, Toda Y, Yamada T, Kinukawa N, Terada N, Kobayashi T, et
al: Increased Akt and phosphorylated Akt expression are associated
with malignant biological features of prostate cancer in Japanese
men. BJU Int. 100:685–690. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yeh YC, Yang CP, Lee SS, Horng CT, Chen
HY, Cho TH, Yang ML, Lee CY, Li MC and Kuan YH: Acute lung injury
induced by lipopolysaccharide is inhibited by wogonin in mice via
reduction of Akt phosphorylation and RhoA activation. J Pharm
Pharmacol. 68:257–263. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Chen YJ, Su JH, Tsao CY, Hung CT, Chao HH,
Lin JJ, Liao MH, Yang ZY, Huang HH, Tsai FJ, et al: Sinulariolide
induced hepatocellular carcinoma apoptosis through activation of
mitochondrial-related apoptotic and PERK/eIF2alpha/ATF4/CHOP
pathway. Molecules. 18:10146–10161. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Checchi PM, Nettles JH, Zhou J, Snyder JP
and Joshi HC: Microtubule-interacting drugs for cancer treatment.
Trends Pharmacol Sci. 24:361–365. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Chang LC, Yu YL, Liu CY, Cheng YY, Chou
RH, Hsieh MT, Lin HY, Hung HY, Huang LJ, Wu YC and Kuo SC: The
newly synthesized 2-arylnaphthyridin-4-one, CSC-3436, induces
apoptosis of non-small cell lung cancer cells by inhibiting tubulin
dynamics and activating CDK1. Cancer Chemother Pharmacol.
75:1303–1315. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Lee MH, Cho Y, Jung BC, Kim SH, Kang YW,
Pan CH, Rhee KJ and Kim YS: Parkin induces G2/M cell cycle arrest
in TNF-α-treated HeLa cells. Biochem Biophys Res Commun. 464:63–69.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Freeman RS, Estus S and Johnson EM Jr:
Analysis of cell cycle-related gene expression in postmitotic
neurons: Selective induction of Cyclin D1 during programmed cell
death. Neuron. 12:343–355. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Montagnoli A, Moll J and Colotta F:
Targeting cell division cycle 7 kinase: A new approach for cancer
therapy. Clin Cancer Res. 16:4503–4508. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Hassan M, Watari H, AbuAlmaaty A, Ohba Y
and Sakuragi N: Apoptosis and molecular targeting therapy in
cancer. Biomed Res Int. 2014:1508452014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Childs AC, Phaneuf SL, Dirks AJ, Phillips
T and Leeuwenburgh C: Doxorubicin treatment in vivo causes
cytochrome C release and cardiomyocyte apoptosis, as well as
increased mitochondrial efficiency, superoxide dismutase activity,
and Bcl-2:Bax ratio. Cancer Res. 62:4592–4598. 2002.PubMed/NCBI
|
|
39
|
Breckenridge DG and Xue D: Regulation of
mitochondrial membrane permeabilization by BCL-2 family proteins
and caspases. Curr Opin Cell Biol. 16:647–652. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Seo BR, Min KJ, Cho IJ, Kim SC and Kwon
TK: Correction: Curcumin significantly enhances dual PI3K/Akt and
mTOR inhibitor NVP-BEZ235-induced apoptosis in human renal
carcinoma caki cells through down-regulation of p53-dependent Bcl-2
expression and inhibition of Mcl-1 protein stability. PLoS One.
11:e01518862016. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Martelli AM, Tazzari PL, Evangelisti C,
Chiarini F, Blalock WL, Billi AM, Manzoli L, McCubrey JA and Cocco
L: Targeting the phosphatidylinositol 3-kinase/Akt/mammalian target
of rapamycin module for acute myelogenous leukemia therapy: From
bench to bedside. Curr Med Chem. 14:2009–2023. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Récher C, Dos Santos C, Demur C and
Payrastre B: mTOR, a new therapeutic target in acute myeloid
leukemia. Cell Cycle. 4:1540–1549. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Perl AE, Kasner MT, Tsai DE, Vogl DT,
Loren AW, Schuster SJ, Porter DL, Stadtmauer EA, Goldstein SC, Frey
NV, et al: A phase I study of the mammalian target of rapamycin
inhibitor sirolimus and MEC chemotherapy in relapsed and refractory
acute myelogenous leukemia. Clin Cancer Res. 15:6732–6739. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Meja K, Stengel C, Sellar R, Huszar D,
Davies BR, Gale RE, Linch DC and Khwaja A: PIM and AKT kinase
inhibitors show synergistic cytotoxicity in acute myeloid leukaemia
that is associated with convergence on mTOR and MCL1 pathways. Br J
Haematol. 167:69–79. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Lu W, Wang F, Zhang T, Dong J, Gao H, Su
P, Shi Y and Zhang J: Search for novel histone deacetylase
inhibitors. Part II: Design and synthesis of novel isoferulic acid
derivatives. Bioorg Med Chem. 22:2707–2713. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Meeprom A, Sompong W, Suantawee T,
Thilavech T, Chan CB and Adisakwattana S: Isoferulic acid prevents
methylglyoxal-induced protein glycation and DNA damage by free
radical scavenging activity. BMC Complement Altern Med. 15:3462015.
View Article : Google Scholar : PubMed/NCBI
|