Introduction
Conventional therapies have succeeded in controlling
periodontal inflammation but cannot restore the damage to
periodontal tissues (1). In
previous years, tissue engineering technologies have offered a
promising approach for complete periodontal tissue regeneration
(2). A previous study also
indicated that human periodontal ligament stem cells (hPDLSCs)
possess multi-potency and can differentiate into cementoblast-like
cells, adipocytes and collagen-forming cells (3). Thus, hPDLSCs are a promising stem
cell population for complete periodontal tissue regeneration.
Decreased estrogen levels are the primary cause of
osteoporosis in menopausal women. Accordingly, estrogen can be used
to prevent postmenopausal osteoporosis (4). After bone maturation, this tissue is
constantly remodeled throughout adult life (5). Estrogen plays an important role in
maintaining the balance between bone resorption and bone formation
(6). Phytoestrogens are a group of
naturally existing compounds in plants with mammalian estrogen-like
activity (7). Resveratrol, a
well-known phytoestrogen, is a phenolic compound that exists in red
wines, peanuts, mulberries and the berry skins of the majority of
grape cultivars (8). Previous
studies have shown that resveratrol has estrogenic activity
(9,10). It also has several biological
effects including anticancer activity (11), cardiovascular protection (12) and anti-inflammatory properties
(13).
Furthermore, the bone protective activities of
resveratrol have attracted extensive interest (14,15).
One study has demonstrated that resveratrol promotes the
proliferation and differentiation of osteoblastic MC3T3-E1 cells
in vitro (16). Dai et
al (17) reported that
resveratrol stimulates human mesenchymal stem cell (MSC)
proliferation and osteoblast differentiation via ERK-dependent
ERK1/2 activation. Previously, resveratol was also found to enhance
the osteogenesis of human MSCs by upregulating Runt-related
transcription factor 2 (Runx2) gene expression via the
NAD-dependent protein deacetylase sirtuin-1 (SIRT1)/forkhead box
protein O3 (FOXO3A) axis (18).
Tumor necrosis factor-α (TNF-α) plays a critical
role in inflammation, which can be rapidly secreted by macrophages
and T lymphocytes (19).
Accordingly, it is involved in several inflammatory bone diseases
such as rheumatoid arthritis and periodontitis (20,21).
This cytokine can also inhibit the osteogenic differentiation of
MSCs. However, although it has been hypothesized that resveratrol
could positively regulate the osteoblastic differentiation of
hPDLSCs in an inflammatory microenvironment induced by TNF-α, this
has previously not been elucidated.
In the present study, the effect of resveratrol and
TNF-α on hPDLSC proliferation and osteoblast differentiation was
investigated.
Materials and methods
Tissue collection and cell
culture
hPDLSCs were collected from wisdom teeth and
premolars extracted from 20 individuals aged 12–20 years.
Volunteers included 15 males and five females with a mean age of 14
years (age, 12–20 years). The recruitment was carried out between
June and September 2017. All procedures were performed at the
Department of Oral & Maxillofacial Surgery (School of
Stomatology, Shandong University), and written informed consent was
obtained from each participant and the legal guardians of all
children. The extracted teeth were placed in α-Minimal Essential
Medium (α-MEM; Gibco; Thermo Fisher Scientific, Inc.) supplemented
with penicillin (400 U/ml) and streptomycin (400 mg/ml) on ice and
were delivered to the Shandong Provincial Key Laboratory of Oral
Tissue Regeneration immediately. Periodontal membranes were scraped
from the middle third of the healthy non-carious root surface and
cut into 1.0 mm3 fragments with an aseptic scalpel, as
previously described (22). The
fragments were tiled on the bottom of a culture bottle and cultured
in α-MEM supplemented with 20% FBS (Gibco; Thermo Fisher
Scientific, Inc.), 100 U/ml penicillin G and 100 µg/ml
streptomycin. The culture bottle was turned over 4 h later and
incubated at 37°C with 5% CO2. When the cells grew to
80% confluence, cells were digested with 0.25% trypsin. Single cell
colonies were subcultured as previously described (23), and cells at passages 3–5 were used
for the following experiments. hPDLSC morphology was assessed using
an inverted microscope at a magnification of ×100 and ×200.
Cell proliferation assay
The effect of resveratrol on hPDLSC viability was
investigated using a Cell Counting Kit-8 (CCK-8) assay (Dojindo
Molecular Technologies, Inc.), according to the manufacturer's
instructions. hPDLSCs were seeded into 96-well plates at a density
of 3×103 per well. Cells were then cultured with
complete medium (α-MEM containing 10% FBS) for 24 h at 37°C. After
the cells attached to the flask, the culture medium was changed to
complete medium containing TNF-α (1,5 and 10 ng/ml; PeproTech)
and/or Resveratrol (0.1,1 and 10 µM; Sigma-Aldrich; Merck KGaA).
The medium was subsequently changed every three days. On day one,
three and five, the culture medium in each well was substituted for
10% CCK-8 reagent (100 µl). Next, the 96-well plate was incubated
for 2 h in an incubator at 37°C. Absorbance was recorded at a
wavelength of 450 nm using a SPECTROstar Nano ultraviolet
spectrophotometer (Spectro Analytical Instruments GmbH).
Flow cytometry
To analyze cell surface marker proteins, hPDLSCs at
passage 3 were collected using 0.25% trypsin and resuspended in
PBS. In brief, 1×106 cells in suspension were incubated
with fluorescent dye-conjugated monoclonal antibodies at 4°C for 20
min in the dark, washed three times using PBS (Corning, Inc.), and
then analyzed using a flow cytometer and FlowJo®
software (version 10.5.2; BD Biosciences) was used to analyze the
data. The antibodies used for these experiments were CD34-PE (cat.
no. 12-0349-41; 5 µl/test; eBioscience; Thermo Fisher Scientific,
Inc.), CD44-PE (cat. no. 12-0441-81; 0.125 µg/test; Affymetrix;
Thermo Fisher Scientific, Inc.), CD90Thy-1-PE (cat. no. 15-0909-42;
5 µl/test; Affymetrix; Thermo Fisher Scientific, Inc.), CD105-APC
(cat. no. 17-1057-42; 5 µl/test; Affymetrix; Thermo Fisher
Scientific, Inc.) and CD45-PE (cat. no. 12-0451-82; 0.125 µg/test;
eBioscience; Thermo Fisher Scientific, Inc.).
Alkaline phosphatase (ALP) staining
and activity assay
ALP is an osteogenic marker expressed at the early
stage of osteogenic differentiation (24). For ALP staining, hPDLSCs were
cultured in the osteogenic induction medium (Cyagen Science &
Technology Co., Ltd.) for 14 days. The cells were then washed with
PBS and fixed with 4% paraformaldehyde at 4°C for 30 min. A
5-bromo-4-chloro-3-indolyl phosphate (BCIP)/nitro blue tetrazolium
(NBT) ALP staining kit (Beyotime Institute of Biotechnology) was
used for ALP staining according to manufacturer's instructions, and
the stained samples were observed with an inverted fluorescence
microscope (×100). For ALP activity assays, cells were seeded in
6-well plates at the density of 2×105 per well. After 14
days of incubation, hPDLSCs were washed with PBS and scraped into
100 µl of radioimmunoprecipitation assay buffer (Beijing Solarbio
Science & Technology Co., Ltd.) containing 1%
phenylmethylsulphonyl fluoride (Beijing Solarbio Science &
Technology Co., Ltd.). The collected cells were sonicated at 20 KHz
for 15 sec on ice, and the lysates were centrifuged at 12,000 × g
for 15 min at 4°C. Then, ALP activity in each sample was assayed
using the ALP assay kit (Nanjing Jiancheng Bioengineering
Institute) according to manufacturer's instructions. Briefly, 50 µl
of buffer solution, 30 µl of supernatant, and 50 µl of matrix
liquid were added to each well of a 96-well plate and mixed. The
plate was then incubated at 37°C for 15 min. The absorbance was
measured at 520 nm after 150 µl of coloration solution was added to
each well. ALP activity was normalized to total protein amounts
which determined by bicinchoninic acid assay kit (Beijing Solarbio
Science & Technology Co., Ltd), according to manufacturer's
instructions.
Alizarin red staining and calcium
content assay
hPDLSCs were seeded in 6-well plates at
1.5×105 cells per well and cultured for 21 days in
osteogenic medium with or without resveratrol treatment, as
indicated in the experiments (Resveratrol 1 µM; TNF-α 10 ng/ml).
After treatment, the cells were washed three times with PBS and
fixed with 4% paraformaldehyde at room temperature for 30 min.
After three washes with deionized water, hPDLSCs were stained with
1% alizarin red (pH 4.2, Beijing Solarbio Science & Technology
Co., Ltd.) for 10 min at room temperature. Then, 600 µl of 10%
(w/v) cetylpyridinium chloride (Sigma-Aldrich; Merck KGaA) was
added to the stained wells and the absorbance of the extracted dye
was assayed at 562 nm using a spectrophotometer (SPECTROstar
Nano).
Oil Red O staining
For adipogenic induction, hPDLSCs
(1×104/well) were seeded in a 24-well plate. Then, the
cells were exposed to adipogenic induction medium that included
α-MEM containing 10% FBS, 0.2 mmol/l indomethacin (Sigma-Aldrich;
Merck KGaA), 2 µM dexamethasone (Beijing Solarbio Science &
Technology Co., Ltd.), 0.01 g/l insulin (Sigma-Aldrich; Merck KGaA)
and 0.5 mmol/l isobutyl-meth-ylxanthine (Sigma-Aldrich; Merck
KGaA). Following 4 weeks of induction, the lipid droplets were
stained with Oil Red O (cyagen Biosciences, Suzhou, china) for 30
min at room temperature, and then observed with an inverted
fluorescent microscope (Olympus Corporation) at a magnification of
×400.
Reverse transcription-quantitative PCR
(RT-qPCR)
hPDLSCs (2×105/well) were seeded in
6-well plates. Total RNA was isolated after seven, 14 and 21 days
of culture using TRIzol® (Invitrogen; Thermo Fisher
Scientific, Inc.), according to the manufacturer's protocol. cDNA
was synthesized from 1.0 µg of total RNA using RevertAid First
Strand cDNA Synthesis kit (Invitrogen; Thermo Fisher Scientific,
Inc.) according to the manufacturer's instructions. qPCR was
performed utilizing the Roche Light Cycler 480 with a 10 µl
reaction volume. The reaction system contained 5 µl
SYBR® Premix Ex Taq™ (Takara Bio, Inc.), 1 µl cDNA, 0.2
µl of each primer and 3.6 µl RNase-free H2O, following
the manufacturer's protocol. The thermal cycling conditions were as
follows: Incubation at 95°C for 15 min, 45 cycles of denaturation
at 95°C for 5 sec and annealing at 60°C for 35 sec, and a final
extension at 72°C for 3 min. The relative expression levels of
ALP, Runx2, Interleukin (IL)-6 and IL-8 were
normalized to those of GAPDH. The following primers were
used for RT-qPCR: GAPDH forward, 5′-GCACCGTCAAGGCTGAGAAC-3′ and
reverse, 5′-TGGTGAAGACGCCAGTGGA-3′; Runx2 forward,
5′-TCCACACCATTAGGGACCATC-3′ and reverse,
5′-TGCTAATGCTTCGTGTTTCCA-3′; ALP forward,
5′-ATGGGATGGGTGTCTCCACA-3′ and reverse, 5′-CCACGAAGGGGAACTTGTC-3′;
IL-6 forward, 5′-ATAACCACCCCTGACCCAAC-3′ and reverse,
5′-CCCATGCTACATTTGCCGAA-3′ and IL-8 forward,
5′-TCAGAGACAGCAGAGCACAC-3′ and reverse, 5′-GGCAAAACTGCACCTTCACA-3′.
The relative gene expression was calculated using the
2−ΔΔCq method (25),
normalizing to GAPDH levels.
Western blotting
To measure protein levels, hPDLSCs were cultured in
a 6-well plate at a density of 2×105 per well for
further treatment. Cells were cultured with osteogenic induction
medium, osteogenic induction medium + Resveratrol 1 µmol/l,
osteogenic induction medium + TNF-α 10 ng/ml or osteogenic
induction medium + Resveratrol 1 µmol/l + TNF-α 10 ng/ml,
respectively. After 3 weeks of osteogenic induction, cells were
rinsed three times with PBS before being collected with RIPA buffer
(Beyotime Institute of Biotechnology) for 30 min on ice. Then, the
protein concentrations were measured using a bicinchoninic acid
assay (Beijing Solarbio Science & Technology co., Ltd.). Next,
10 µg protein samples were separated using 10% SDS-PAGE gels and
then transferred to PVDF membranes (Bio-Rad Laboratories, Inc.).
The membranes were blocked with 5% non-fat milk for 1 h at room
temperature, incubated with primary antibodies overnight at 4°C,
and then incubated with horseradish peroxidase (HRP)-conjugated
anti-rabbit or anti-mouse IgG (cat. no. SA00001-2 and SA00011; both
at 1:10,000 dilution; Proteintech Group, Inc.) for 1 h at room
temperature. The bands were detected using the chemiluminescent HRP
substrate (EMD Millipore) and scanned utilizing an Amersham Imager
600 (GE Healthcare Life Sciences). Semi-quantitative analysis of
western blotting was performed with the Image J software (version
1.47; National Institutes of Health). GAPDH is used as reference
protein in the study. The primary antibodies and dilution ratios
were as follows: Rabbit anti-human ALP (cat. no. ab108337;
1:20,000; Abcam), rabbit anti-human GAPDH (cat. no. cst2118;
1:1,000; Cell Signaling Technology, Ltd.), rabbit anti-human Runx2
(cat. no. cst12556s; 1:1,000: Cell Signaling Technology, Ltd.),
rabbit anti-human ERK1/2 (cat. no. cst4695; 1:1,000; Cell Signaling
Technology, Ltd.), and rabbit anti-phosphorylated (p)-ERK1/2 (cat.
no. cst4370; 1:1,000; Cell Signaling Technology, Ltd.).
ERK1/2 pathway activation and
inhibition studies
To activate the ERK1/2 pathway, hPDLSCs
(2×105 per well) were seeded in a 6-well plate. The
cells were cultured with α-MEM containing 10% FBS at 37°C for 24 h.
Then, the cells were maintained in α-MEM containing 0.5% FBS for 48
h to synchronize the cells and reduce basal ERK1/2 activity
(26). PD98059 (10−5
mol/l), an inhibitor of the upstream mitogen-activated protein
kinase (MAPK)-ERK 1 pathway (27),
was added to the serum-free culture media 1 h prior to the
application of resveratrol, to assess its effect on ERK1/2 pathway
activity. Then, the lysates were collected for western
blotting.
ELISA
To further clarify the effect of resveratrol (1
µmol/l) or resveratrol (1 µmol/l) plus PD98059 (10−5
mol/l) on TNF-α-induced inflammatory cytokine production, after
TNF-α treatment, the levels of secreted IL-6 and IL-8 within the
conditioned medium were measured using an ELISA kit (IL-6 cat. no.
430507 and IL-8 cat. no. 431507; BioLegend, Inc.) according to the
manufacturer's instructions. The optical density values were
measured using a microplate reader at 450 and 570 nm, and values at
570 nm were subtracted from the absorbance at 450 nm for subsequent
data analysis.
Statistical analysis
All experiments were performed in triplicate. The
significance of differences among the groups was assessed by
one-way ANOVA followed by Tukey's post-hoc test. The significance
of differences among multiple groups at different time points was
assessed by two-way ANOVA followed by Tukey's post-hoc test. Data
were analyzed using GraphPad Prism (version 6; GraphPad Software,
Inc.). All data are presented as the mean ± standard deviation
(SD). P<0.05 was considered to indicate a statistically
significant difference.
Results
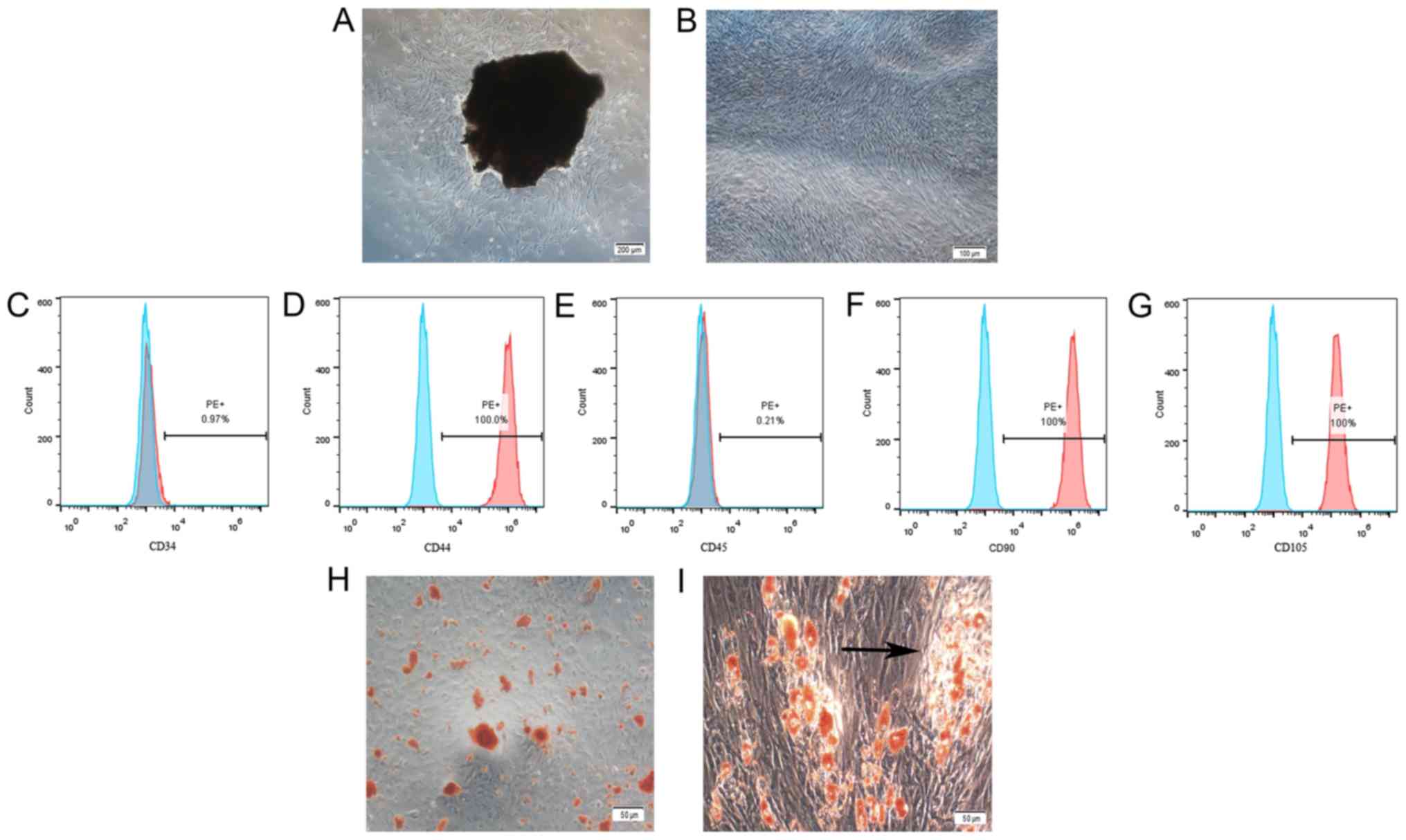
Characterization of hPDLSCs
The characteristics of hPDLSCs were confirmed by
examining their morphology, multi-potency, and expression of cell
surface markers. After seven days of incubation, hPDLSCs displayed
a typical spindle-shaped morphology (Fig. 1A and B). Flow cytometry analysis
showed that the hPDLSCs did not express CD34 or CD45 but expressed
CD44, CD90 and CD105 (Fig. 1C-G).
After three weeks of culture in osteogenic induction medium, some
mineralized nodules were found in the well (Fig. 1C), and lipid droplets were also
observed after four weeks of induction (Fig. 1D).
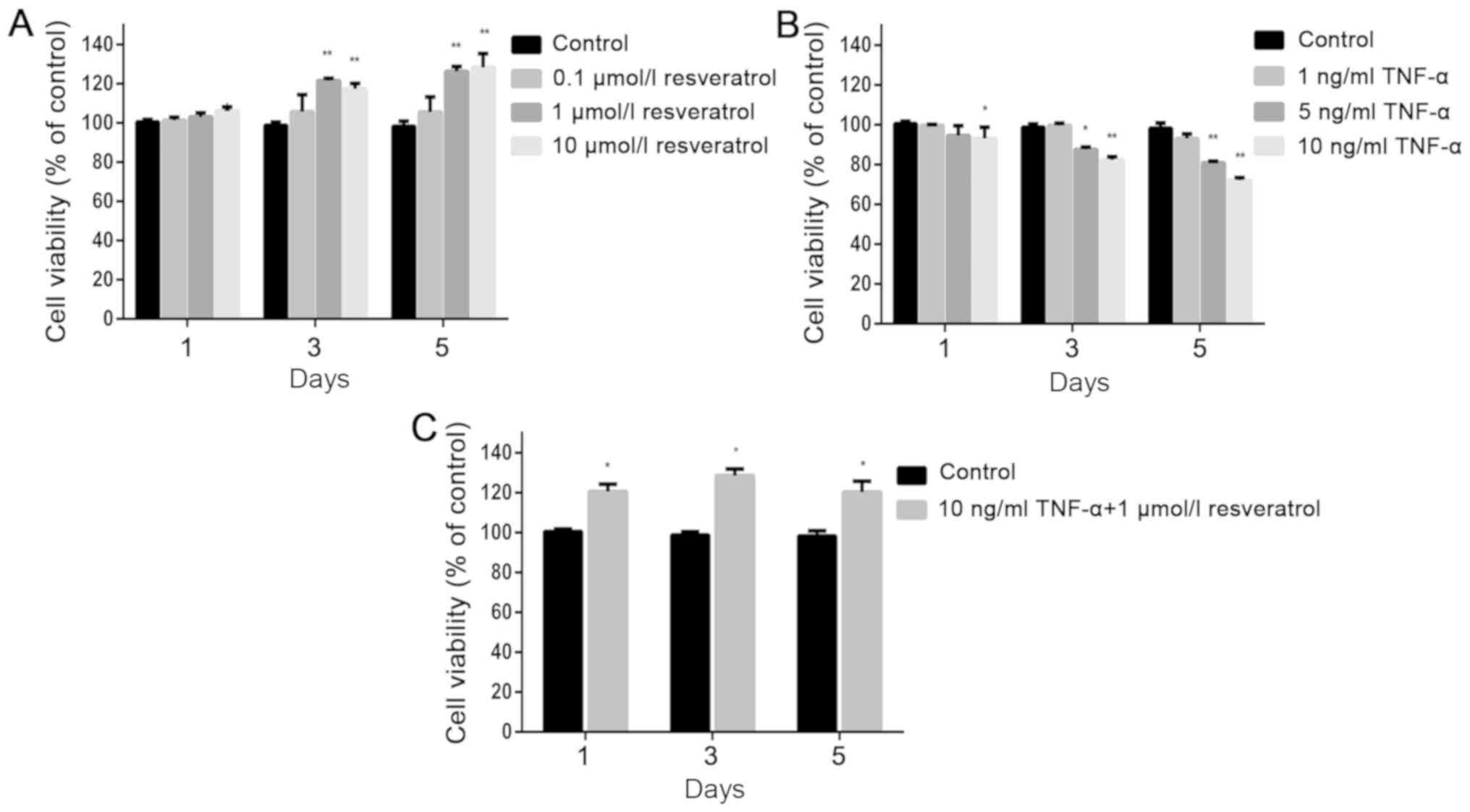
Cell viability of hPDLSCs after
resveratrol and TNF-α treatment
Resveratrol (10−8 to 10−5 M)
was reported to increase human bone marrow-derived mesenchymal stem
cell (BMSC) growth dose-dependently (17). To investigate the effect of
resveratrol and TNF-α on hPDLSC cell viability, cells were treated
with resveratrol and/or TNF-α at different concentrations
(resveratrol: 0.1, 1 and 10 µmol/l; TNF-α: 1, 5 and 10 ng/ml). The
CCK-8 assay was then performed on day one, three, and five. The
CCK-8 assay demonstrated that resveratrol could increase hPDLSC
viability. Furthermore, increases in hPDLSC viability were
identified in the two high-dose groups (1 and 10 µmol/l) on day 3
and 5 (P<0.05; Fig. 2A). As
expected, cell viability decreased after treatment with TNF-α (10
ng/ml) for one day, compared with that in the control group, and
the results were similar when cells were treated with TNF-α for
three or five days. This decrease was significant in the high-dose
group at all three time points (10 ng/ml; Fig. 2B; P<0.05). Moreover, hPDLSC
viability was increased on day one, three and five after
co-treatment with 10 ng/ml TNF-α and 1 µmol/l resveratrol, compared
with that in the control group. These results demonstrated that
resveratrol could alleviate the toxicity of TNF-α in hPDLSCs
(Fig. 2C).
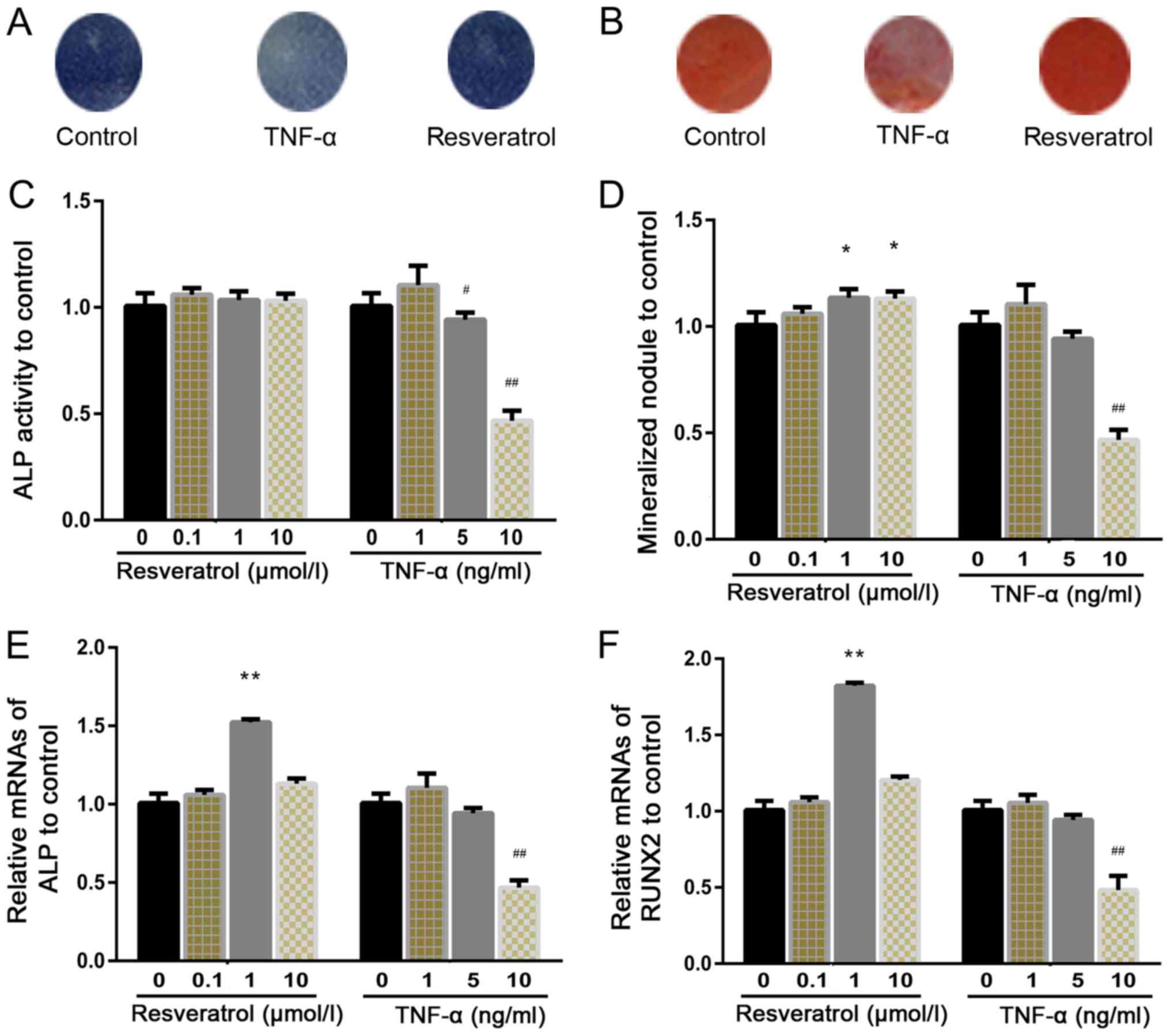
Effects of different doses of
resveratrol and TNF-α on hPDLSC osteogenesis
To examine the effect of resveratrol and TNF-α on
osteogenesis, cells were cultured for 14 days with resveratrol
(0.1, 1 and 10 µmol/l) or TNF-α (1, 5 and 10 ng/ml). After 21 days
of culture, alizarin red staining was carried out (Fig. 3B) and calcium content assays were
performed. The results showed that calcium accumulation in the
TNF-α-treated groups was decreased compared to that in the control
group, and that this decrease was significant in the high-dose
group (10 ng/ml; P<0.01). Furthermore, an increased number of
mineralized nodules could be observed in cells treated with
resveratrol (1 and 10 µmol/l) compared with that in the control
group (Fig. 3D). Cells treated
with TNF-α had a significant decrease in the number of mineralized
nodules, as observed in the high dose group compared with the
control. ALP staining and activity assays were performed as a
standard measurement of osteogenic activity. The results
demonstrated that resveratrol did not have an effect on ALP
staining or activity in hPDLSCs (Fig.
3A and C), whereas TNF-α (1 ng/ml) significantly increased ALP
activity. Furthermore, TNF-α (5 ng/ml) did not alter ALP activity
and TNF-α (10 ng/ml) significantly decreased ALP activity (Fig. 3C).
Furthermore, the mRNA expression of Runx2 and
ALP was assessed by RT-qPCR. Levels of both markers were
significantly increased after treatment with resveratrol (1 µmol/l;
P<0.01) compared with that in the control group. Meanwhile,
ALP and Runx2 mRNA levels were significantly
decreased upon treatment with TNF-α (10 ng/ml) compared with those
in the control groups (Fig. 3E and
F; P<0.01).
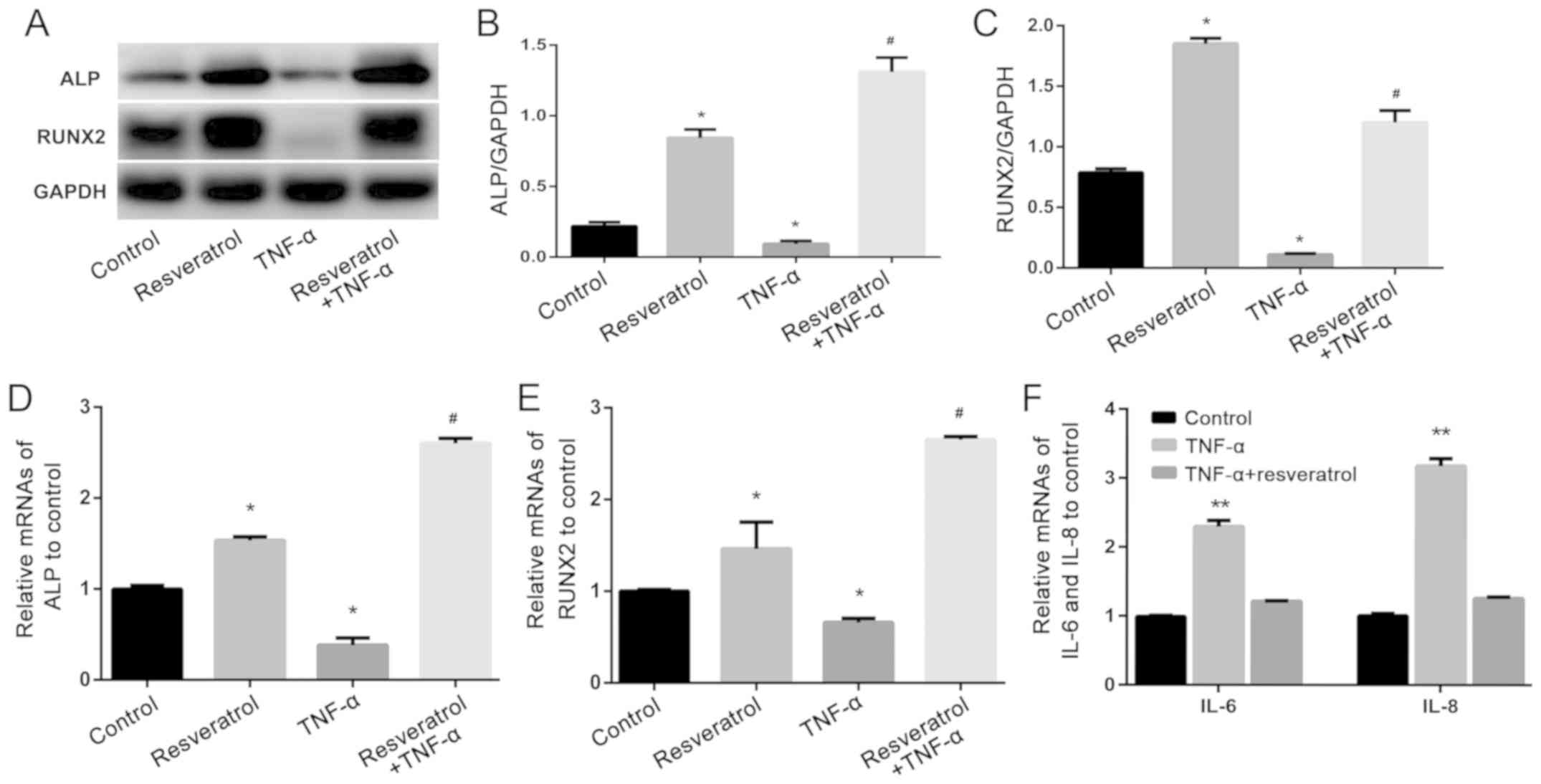
Resveratrol rescues TNF-α induced
inhibition of osteogenesis in hPDLSCs
hPDLSCs were treated with resveratrol (1 µmol/l)
and/or TNF-α (10 ng/ml) during the three weeks of osteogenic
induction. Western blot analysis showed that protein levels of ALP
and Runx2 in the TNF-α-treated group were significantly decreased
compared with those in the control group (Fig. 4A-C; P<0.05). Meanwhile, protein
expression levels in hPDLSCs treated with both TNF-α and
resveratrol were significantly increased compared with those in the
TNF-α-treated group (Fig. 4A-C;
P<0.01). Similar results were found for mRNA expression of
ALP and Runx2 using RT-qPCR (Fig. 4D and E). Taken together, these data
suggested that resveratrol could rescue the TNF-α-induced
inhibition of hPDLSC osteogenesis. Moreover, it was found that
resveratrol and TNF-α could function together to improve bone
formation, by increasing ALP expression at both the mRNA and
protein level, greater than resveratrol alone. Regarding
Runx2 expression at the mRNA level, the combination of
resveratrol and TNF-α had a stronger effect than resveratrol alone,
but the protein levels did not reflect this. However, these changes
are not statistically significant.
Resveratrol attenuates hPDLSC
secretion of inflammatory cytokines induced by TNF-α
To assess cytokine secretion in response to
resveratrol (1 µmol/l) and TNF-α (10 ng/ml), hPDLSCs were treated
with either TNF-α alone or with both TNF-α and resveratrol for 24
h. Data from RT-qPCR showed that the mRNA expression of IL-6
and IL-8 in the TNF-α-treated group was significantly
increased compared with that in the control group (P<0.01).
However, co-treatment did not change mRNA expression of IL-6
and IL-8 compared with that in the control group (Fig. 4F). These results indicated that
resveratrol attenuates the inflammatory effects of TNF-α.
Resveratrol rescues the TNF-α-induced
inhibition of osteogenesis via the ERK1/2 pathway
The MAPK pathway, comprised of serine/threonine
protein kinases, plays an important role in regulating cell
migration, proliferation and differentiation (28,29).
To determine whether the ERK1/2 signaling pathway is involved in
the protective effects of resveratrol on hPDLSC osteogenic
differentiation, the phosphorylation and total levels of ERK in
treated hPDLSCs were measured. Western blot analysis showed that
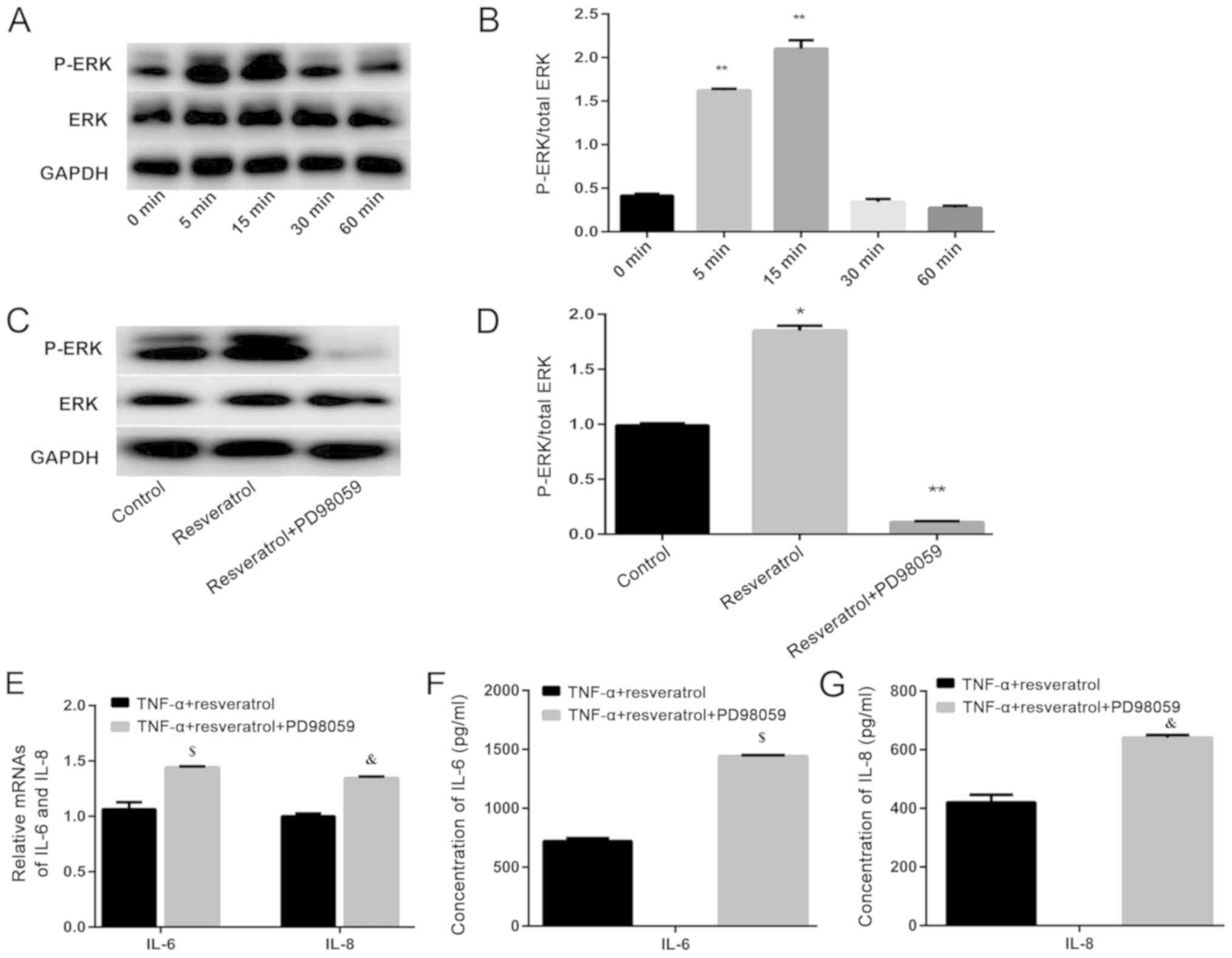
p-ERK gradually increased by 5 min and peaked at 15 min, after
which it returned to basal levels (Fig. 5A and B). Furthermore, PD98059 (10
mmol/l) was added to the serum-free cell culture media 1 h prior to
application of resveratrol to inhibit ERK signaling. As expected,
the protein expression of p-ERK was significantly inhibited by
PD98059, compared with that in the control group (Fig. 5C and D; P<0.01). Results also
showed that the mRNA and protein expression levels of IL-6 and IL-8
increased when PD98059 was added to the co-treated group, compared
with those in the co-treated group alone (Fig. 5E-G). Data indicated that the
anti-inflammatory effects of resveratrol decreased after blocking
the ERK1/2 signaling pathway. Thus, these results together
indicated that resveratrol can activate the ERK1/2 signaling
pathway to attenuate the secretion of inflammatory cytokines by
hPDLSCs upon exposure to TNF-α, this in turn enhances osteogenic
differentiation.
Discussion
Plaque bacteria are the initiating factor of
periodontitis, and the host reaction stimulated by these
microorganisms is the main cause of periodontal tissue destruction
(30). The development of tissue
engineering provides new approaches for dental tissue regeneration
(31). In 2004, Seo et al
(3) successfully isolated and
cultured stem cells from periodontal ligaments of the third molar.
PDLSCs express mesenchymal stem-cell markers, and can differentiate
into cementoblast-like cells, adipocytes and collagen-forming cells
under defined culture conditions (3). Moreover, PDLSCs also have the
capacity to generate a cementum/PDL-like structures when
transplanted into immunocompromised rodents (3). This highlighted that this stem cells
could be used for research on periodontal tissue repair.
In the present study, a comprehensive assessment of
the effect of TNF-α and resveratrol on the proliferation and
osteogenic differentiation of hPDLSCs was carried out. Results
showed that TNF-α could significantly decrease hPDLSC viability and
osteogenic differentiation at a high concentration (10 ng/ml),
whereas co-treatment with TNF-α and resveratrol rescued such
TNF-α-induced inhibitory effects.
TNF-α is a 17-kDa cytokine produced by mononuclear
macrophages and other immune cells (32). It is currently recognized as an
important endogenous inducible pro-inflammatory cytokine and a
critical inflammatory mediator of responses to infectious diseases
(33). Additionally, TNF-α
actively participates in osteoclastogenesis and tissue destruction
observed in periodontal diseases (34). Gilbert et al (35) reported that TNF-α is a potent
inhibitor of osteoblast differentiation in a normal osteoblast-like
cell line (MC3T3-E1 cells). Lacey et al (36) reported that TNF-α inhibits ALP
activity, as well as ALP and Runx2 gene expression
during the osteoblastic differentiation of murine BMSCs.
In addition, Feng et al (37) found that 10 ng/ml of TNF-α could
promote mineralization and mineralization-related gene expression
via the NF-κB signaling pathway in dental pulp stem cells.
Furthermore, in a murine mesenchymal stem cell line (ST2 cells),
levels of Runx2, osteocalcin and ALP were found to be upregulated
in cell cultures treated with TNF-α at lower concentrations (0.01
and 0.1 ng/ml), but were downregulated in cell cultures treated
with TNF-α at higher concentrations of 10 and 100 ng/ml. Long-term
treatment with TNF-α at all concentrations induced inhibitory
effects on mineral nodule formation (38). However, after treatment with TNF-α
at 1 ng/ml, human primary osteoblasts were able to promote the
osteogenesis of adipose tissue-derived MSCs (39). The exact reason for these
contradictory results is unknown, but one possibility is the
different stem cell types used for in vitro studies.
To the best of our knowledge, the effect of TNF-α on
the osteogenic differentiation of periodontal ligament stem cells
has not been previously reported. In the present study, the effect
of different TNF-α concentrations on the proliferation and
osteogenic differentiation of hPDLSCs was assessed. Data showed
that TNF-α (10 ng/ml) could significantly inhibit both processes in
hPDLSCs. Furthermore, the present study found that co-treatment of
hPDLSCs with resveratrol and TNF-α could prevent this inhibitory
effect. Similar protective effects of resveratrol in response to
TNF-α have been were reported in cardiac stem cells (40), fibroblasts (41) and endothelial cells (42).
Previous studies have also shown that resveratrol
possesses potent bone-protective properties. Specifically, Song
et al (43) reported that
resveratrol rescues the inhibitory effects of cyclosporin A on
proliferation and osteoblastic differentiation in murine BMSCs.
Further to this, Boissy et al (44) reported that resveratrol can enhance
the mRNA expression of osteocalcin and osteopontin, two
osteoblastic markers, in immortalized osteoblast-like cells. Other
studies have also confirmed these findings (16–18).
These in vitro findings have been further supported by an
in vivo study. Durbin et al (45) showed that resveratrol consumption
could prevent bone loss in the hind limbs of rats. Moreover, three
months of treatment with resveratrol was found to rescue
ovariectomy-induced bone loss in rats (46,47).
In the present study, the protein expression of ALP and Runx2 were
measured to assess the effect of resveratrol on hPDLSCs. Expression
of both markers at the protein level were increased by resveratrol,
with the relative expression of ALP and Runx2 genes following a
similar trend. These results are consistent with a study on ST2
cells, which showed that resveratrol could significantly enhance
the mRNA expression of Runx2 (48). Although there is evidence
supporting the enhancing effects of resveratrol on the osteogenic
differentiation of hPDLSCs, the underlying mechanism was previously
unclear.
Tseng et al (18) reported a novel mechanism associated
with resveratrol in enhancing the osteogenesis of human MSCs by
upregulating Runx2 gene expression via the SIRT1/FOXO3A
axis. Resveratrol can also promote the osteoblastic differentiation
of multipotent mesenchymal cells via the canonical Wnt signaling
pathway (48). Further to this,
via the ERK/MAPK/nitric oxide synthase (NOS)/cyclic GMP axis,
resveratrol was found to promote osteoblastic differentiation and
osteogenic gene expression in human BMSC cultures (43). To explore the molecular mechanism
underlying the effects of resveratrol on hPDLSCs, the expression
and phosphorylation levels of ERK1/2 were evaluated. The present
study demonstrated that resveratrol could rapidly activate ERK1/2
signaling within 5 min. Moreover, results showed that this was
blocked on addition of PD98059 (10−5 mol/l) 1 h prior to
resveratrol application. A previous study found that resveratrol
stimulates osteoblastic differentiation and osteogenic gene
expression in human MSCs via ERK-dependent ERK1/2 activation
(13). Therefore, it is speculated
that resveratrol can similarly enhance the osteogenic
differentiation of hPDLSCs by activating the ERK1/2 pathway.
The present study was the first to find that
Resveratrol can attenuate the secretion of inflammatory cytokines
mediated by TNF-α. Upon further exploring the underlying
mechanisms, it was found that the mRNA expression of IL-6
and IL-8 significantly increased after addition of PD98059
to the co-treated group (Fig. 5E).
This result is consistent with the protein levels of secreted IL-6
and IL-8, measured by ELISA (Fig. 5F
and G). These results further indicate that the
anti-inflammatory effect of resveratrol might occur via activation
of the ERK1/2 signaling pathway. Previous studies reported that
resveratrol attenuates inflammation in experimental caustic
esophageal burns (49), acute
small intestinal inflammation (50), and tissue damage in a dextran
sulfate sodium-induced colitis rat model (51). Resveratrol also attenuates
inflammation by inhibiting the activity of NF-κB and activating
protein 1 simultaneously. The present study provides a new
molecular mechanism by which resveratrol exerts anti-inflammatory
activity, specifically, by activating the ERK1/2 pathway, improving
the inflammatory microenvironment, and enhancing the osteogenic
differentiation of hPDLSCs.
In conclusion, the present study is the first to
demonstrate that resveratrol not only enhances the osteogenesis of
hPDLSCs by activating the ERK1/2 pathway, but also exerts
anti-inflammatory effects by activating this pathway. Ultimately,
resveratrol decreases the TNF-α-mediated secretion of inflammatory
cytokines and enhances osteogenesis in hPDLSCs. Therefore,
resveratrol may be a good candidate for an anti-inflammatory agent
that can induce bone remodeling in response to an inflammatory
microenvironment. However, further in vivo experiments are
required to evaluate the efficacy and safety of its clinical
application for the treatment of periodontitis.
Acknowledgements
The authors would like to thank the Director of
Shandong Provincial Key Laboratory of Oral Tissue Regeneration for
providing technical support.
Funding
This work was supported by a grant from the National
Natural Science Foundation of China (grant. no. 81371180).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
JZ designed the experiments. JY and XW performed the
experiments. JZ, XW, DM, HG and DZ analyzed the data. JY wrote the
manuscript. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
This study was approved by the Medical Ethical
Committee of the School of Stomatology, Shandong University. Each
participant and the legal guardian of all children provided written
informed consent in accordance with The Declaration of
Helsinki.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
ALP
|
alkaline phosphatase
|
|
BMSCs
|
bone marrow-derived mesenchymal stem
cells
|
|
hPDLSCs
|
human periodontal ligament stem
cells
|
|
MSCs
|
mesenchymal stem cells
|
|
TNF-α
|
tumor necrosis factor-α
|
References
|
1
|
Chen FM, Zhang J, Zhang M, An Y, Chen F
and Wu ZF: A review on endogenous regenerative technology in
periodontal regenerative medicine. Biomaterials. 31:7892–7927.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Han J, Menicanin D, Gronthos S and Bartold
PM: Stem cells, tissue engineering and periodontal regeneration.
Aust Dent J. 59 (Suppl 1):S117–S130. 2014. View Article : Google Scholar
|
|
3
|
Seo BM, Miura M, Gronthos S, Bartold PM,
Batouli S, Brahim J, Young M, Robey PG, Wang CY and Shi S:
Investigation of multipotent postnatal stem cells from human
periodontal ligament. Lancet. 364:149–155. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lindsay R and Tohme JF: Estrogen treatment
of patients with established postmenopausal osteoporosis. Obstet
Gynecol. 76:290–295. 1990.PubMed/NCBI
|
|
5
|
Zaidi M: Skeletal remodeling in health and
disease. Nat Med. 13:791–801. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Falahati-Nini A, Riggs BL, Atkinson EJ,
O'Fallon WM, Eastell R and Khosla S: Relative contributions of
testosterone and estrogen in regulating bone resorption and
formation in normal elderly men. J Clin Invest. 106:1553–1560.
2000. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lampe J and Messina M: Are phytoestrogens
nature's cure for what ails us? A look at the research. Interview
by Nancy I. Hahn. J Am Diet Assoc. 98:974–976. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Jang M, Cai L, Udeani GO, Slowing KV,
Thomas CF, Beecher CW, Fong HH, Farnsworth NR, Kinghorn AD, Mehta
RG, et al: Cancer chemopreventive activity of resveratrol, a
natural product derived from grapes. Science. 275:218–210. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Gehm BD, McAndrews JM, Chien PY and
Jameson JL: Resveratrol, a polyphenolic compound found in grapes
and wine, is an agonist for the estrogen receptor. Proc Natl Acad
Sci USA. 94:14138–14143. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hao XD, Chang J, Qin BY, Zhong C, Chu ZB,
Huang J, Zhou WJ and Sun X: Synthesis, estrogenic activity, and
anti-osteoporosis effects in ovariectomized rats of resveratrol
oligomer derivatives. Eur J Med Chem. 102:26–38. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Savouret JF and Quesne M: Resveratrol and
cancer: A review. Biomed Pharmacother. 56:84–87. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Cao Z and Li Y: Potent induction of
cellular antioxidants and phase 2 enzymes by resveratrol in
cardiomyocytes: Protection against oxidative and electrophilic
injury. Eur J Pharmacol. 489:39–48. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Gómez-Zorita S, Fernández-Quintela A, Lasa
A, Hijona E, Bujanda L and Portillo MP: Effects of resveratrol on
obesity-related inflammation markers in adipose tissue of
genetically obese rats. Nutrition. 29:1374–1380. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ornstrup MJ, Harsløf T, Kjær TN, Langdahl
BL and Pedersen SB: Resveratrol increases bone mineral density and
bone alkaline phosphatase in obese men: A randomized
placebo-controlled trial. J Clin Endocrinol Metab. 99:4720–4729.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Bo S, Gambino R, Ponzo V, Cioffi I, Goitre
I, Evangelista A, Ciccone G, Cassader M and Procopio M: Effects of
resveratrol on bone health in type 2 diabetic patients. A
double-blind randomized-controlled trial. Nutr Diabetes. 8:512018.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Mizutani K, Ikeda K, Kawai Y and Yamori Y:
Resveratrol stimulates the proliferation and differentiation of
osteoblastic MC3T3-E1 cells. Biochem Biophys Res Commun.
253:859–863. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Dai Z, Li Y, Quarles LD, Song T, Pan W,
Zhou H and Xiao Z: Resveratrol enhances proliferation and
osteoblastic differentiation in human mesenchymal stem cells via
ER-dependent ERK1/2 activation. Phytomedicine. 14:806–814. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Tseng PC, Hou SM, Chen RJ, Peng HW, Hsieh
CF, Kuo ML and Yen ML: Resveratrol promotes osteogenesis of human
mesenchymal stem cells by upregulating Runx2 gene expression via
the SIRT1/FOXO3A axis. J Bone Miner Res. 26:2552–2563. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kwan Tat S, Padrines M, Théoleyre S,
Heymann D and Fortun Y: IL-6, RANKL, TNF-alpha/IL-1: Interrelations
in bone resorption pathophysiology. Cytokine Growth Factor Rev.
15:49–60. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Brennan FM, Maini RN and Feldmann M: TNF
alpha-a pivotal role in rheumatoid arthritis? Br J Rheumatol.
31:293–298. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Bartold PM, Cantley MD and Haynes DR:
Mechanisms and control of pathologic bone loss in periodontitis.
Periodontol. 2000(53): 55–69. 2010. View Article : Google Scholar
|
|
22
|
Digirolamo CM, Stokes D, Colter D, Phinney
DG, Class R and Prockop DJ: Propagation and senescence of human
marrow stromal cells in culture: A simple colony-forming assay
identifies samples with the greatest potential to propagate and
differentiate. Br J Haematol. 107:275–281. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Du L, Yang P and Ge S: Stromal
cell-derived factor-1 significantly induces proliferation,
migration, and collagen type I expression in a human periodontal
ligament stem cell subpopulation. J Periodontol. 83:379–388. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Shigetani Y, Ohkura N, Yoshiba K, Ohshima
H, Hosoya A, Yoshiba A and Okiji T: GaAlAs laser-induced pulp
mineralization involves dentin matrix protein 1 and osteopontin
expression. Oral Dis. 22:399–405. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Simmons CA, Matlis S, Thornton AJ, Chen S,
Wang CY and Mooney DJ: Cyclic strain enhances matrix mineralization
by adult human mesenchymal stem cells via the extracellular
signal-regulated kinase (ERK1/2) signaling pathway. J Biomech.
36:1087–1096. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Alessi DR, Cuenda A, Cohen P, Dudley DT
and Saltiel AR: PD 098059 is a specific inhibitor of the activation
of mitogen-activated protein kinase kinase in vitro and in vivo. J
Biol Chem. 270:27489–27494. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Gehart H, Kumpf S, Ittner A and Ricci R:
MAPK signalling in cellular metabolism: Stress or wellness? EMBO
Rep. 11:834–840. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Roy SK, Srivastava RK and Shankar S:
Inhibition of PI3K/AKT and MAPK/ERK pathways causes activation of
FOXO transcription factor, leading to cell cycle arrest and
apoptosis in pancreatic cancer. J Mol Signal. 5:102010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Page RC: The pathobiology of periodontal
diseases may affect systemic diseases: Inversion of a paradigm. Ann
Periodontol. 3:108–120. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Galler KM and D'Souza RN: Tissue
engineering approaches for regenerative dentistry. Regen Med.
6:111–124. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Beutler B, Greenwald D, Hulmes JD, Chang
M, Pan YC, Mathison J, Ulevitch R and Cerami A: Identity of tumour
necrosis factor and the macrophage-secreted factor cachectin.
Nature. 316:552–554. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Suffredini AF, Reda D, Banks SM, Tropea M,
Agosti JM and Miller R: Effects of recombinant dimeric TNF receptor
on human inflammatory responses following intravenous endotoxin
administration. J Immunol. 155:5038–5045. 1995.PubMed/NCBI
|
|
34
|
Brunetti G, Colucci S, Pignataro P,
Coricciati M, Mori G, Cirulli N, Zallone A, Grassi FR and Grano M:
T cells support osteoclastogenesis in an in vitro model derived
from human periodontitis patients. J Periodontol. 76:1675–1680.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Gilbert L, He X, Farmer P, Boden S,
Kozlowski M, Rubin J and Nanes MS: Inhibition of osteoblast
differentiation by tumor necrosis factor-alpha. Endocrinology.
141:3956–3964. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Lacey DC, Simmons PJ, Graves SE and
Hamilton JA: Proinflammatory cytokines inhibit osteogenic
differentiation from stem cells: Implications for bone repair
during inflammation. Osteoarthritis Cartilage. 17:735–742. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Feng X, Feng G, Xing J, Shen B, Li L, Tan
W, Xu Y, Liu S, Liu H, Jiang J, et al: TNF-α triggers osteogenic
differentiation of human dental pulp stem cells via the NF-κB
signalling pathway. Cell Biol Int. 37:1267–1275. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Huang H, Zhao N, Xu X, Xu Y, Li S, Zhang J
and Yang P: Dose-specific effects of tumor necrosis factor alpha on
osteogenic differentiation of mesenchymal stem cells. Cell Prolif.
44:420–427. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Lu Z, Wang G, Dunstan CR and Zreiqat H:
Short-term exposure to tumor necrosis factor-alpha enables human
osteoblasts to direct adipose tissue-derived mesenchymal stem cells
into osteogenic differentiation. Stem Cells Dev. 21:2420–2429.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Gorbunov N, Petrovski G, Gurusamy N, Ray
D, Kim DH and Das DK: Regeneration of infarcted myocardium with
resveratrol-modified cardiac stem cells. J Cell Mol Med.
16:174–184. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zhu X, Liu Q, Wang M, Liang M, Yang X, Xu
X, Zou H and Qiu J: Activation of Sirt1 by resveratrol inhibits
TNF-α induced inflammation in fibroblasts. PLoS One. 6:e270812011.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Bertelli AA, Baccalini R, Battaglia E,
Falchi M and Ferrero ME: Resveratrol inhibits TNF alpha-induced
endothelial cell activation. Therapie. 56:613–616. 2001.PubMed/NCBI
|
|
43
|
Song LH, Pan W, Yu YH, Quarles LD, Zhou HH
and Xiao ZS: Resveratrol prevents CsA inhibition of proliferation
and osteoblastic differentiation of mouse bone marrow-derived
mesenchymal stem cells through an ER/NO/cGMP pathway. Toxicol In
Vitro. 20:915–922. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Boissy P, Andersen TL, Abdallah BM, Kassem
M, Plesner T and Delaissé JM: Resveratrol inhibits myeloma cell
growth, prevents osteoclast formation, and promotes osteoblast
differentiation. Cancer Res. 65:9943–9952. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Durbin SM, Jackson JR, Ryan MJ, Gigliotti
JC, Alway SE and Tou JC: Resveratrol supplementation preserves long
bone mass, microstructure, and strength in hindlimb-suspended old
male rats. J Bone Miner Metab. 32:38–47. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Liu ZP, Li WX, Yu B, Huang J, Sun J, Huo
JS and Liu CX: Effects of trans-resveratrol from Polygonum
cuspidatum on bone loss using the ovariectomized rat model. J Med
Food. 8:14–19. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Zhao H, Li X, Li N, Liu T, Liu J, Li Z,
Xiao H and Li J: Long-term resveratrol treatment prevents
ovariectomy-induced osteopenia in rats without hyperplastic effects
on the uterus. Br J Nutr. 111:836–846. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Zhou H, Shang L, Li X, Zhang X, Gao G, Guo
C, Chen B, Liu Q, Gong Y and Shao C: Resveratrol augments the
canonical Wnt signaling pathway in promoting osteoblastic
differentiation of multipotent mesenchymal cells. Exp Cell Res.
315:2953–2962. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Uguralp S, Irsi C, Aksoy T, Karabulut AB,
Kirimlioglu H and Mizrak B: Resveratrol attenuates inflammation and
stricture formation in experimental caustic esophageal burns.
Pediatr Surg Int. 24:425–430. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Bereswill S, Muñoz M, Fischer A, Plickert
R, Haag LM, Otto B, Kühl AA, Loddenkemper C, Göbel UB and Heimesaat
MM: Anti-inflammatory effects of resveratrol, curcumin and
simvastatin in acute small intestinal inflammation. PLoS One.
5:e150992010. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Larrosa M, Yañéz-Gascón MJ, Selma MV,
González-Sarrías A, Toti S, Cerón JJ, Tomás-Barberán F, Dolara P
and Espín JC: Effect of a low dose of dietary resveratrol on colon
microbiota, inflammation and tissue damage in a DSS-induced colitis
rat model. J Agric Food Chem. 57:2211–2220. 2009. View Article : Google Scholar : PubMed/NCBI
|