Introduction
Melanoma is one of the most common skin cancers, but
its prognosis after metastasis remains poor (1). As reported in a previous cohort
study, the morbidity of melanoma in the United States, United
Kingdom and Sweden will continue to increase until at least 2022
(2). Traditional treatments,
including radiotherapy and chemotherapy, have low efficacy in
advanced melanoma patients (3). In
contrast, the discovery of BRAF inhibitors developed for BRAF gene
mutations in melanoma, such as vemurafenib and dabrafenib, has
largely extended progression-free-survival (PFS) compared with the
PFS seen with chemotherapy. However, melanoma has been found to be
highly susceptible to acquired BRAF inhibitor resistance (4,5).
Thus, it is imperative to explore methods that overcome resistance
to BRAF inhibitors. The progress obtained in recent years reveals
that the mechanisms underlying resistance to BRAF inhibitors are
multiple, complicated and unpredictable (6). To date, a combination of
mitogen-activated protein kinase kinase inhibitors and BRAF
inhibitors has been shown to be clinically effective (7). However, the combination of these two
targeted drugs can only partially impede BRAF inhibitor resistance
(8). Therefore, the development of
novel agents is urgently required to overcome BRAF inhibitor
resistance.
Modern drug discovery programmes are gradually
placing more focus on the screening of plants or other natural
products, given their ease of availability, cost-effective nature
and safer toxicity profiles. Juglone (5-hydroxy-1,4-naphthoquinone)
is a natural naphthoquinone that is isolated from the roots,
leaves, woods and fruits of walnut trees. It is not only a drug
used to treat infections, but also an anti-tumour agent (9,10).
It has been reported that juglone exerts cytotoxic and genotoxic
effects against B16F10 melanoma cells (11). It has also displayed
radiation-sensitizing potential (12). Our previous study demonstrated that
juglone potentiated tumour necrosis factor-related
apoptosis-inducing ligand-induced apoptosis in human melanoma cells
(13). The cytotoxic and
sensitizing effects of juglone can be attributed to mechanisms such
as the induction of reactive oxygen species (ROS) (11,13,14).
It has been documented that inducing ROS production might reverse
the resistance to BRAF inhibition (15). Therefore, it was hypothesized that
juglone might eradicate melanoma resistance to BRAF inhibitors
through the induction of ROS. In the present study, the effect of
juglone on BRAF-induced cytotoxicity was examined, and it was
demonstrated that the underlying mechanisms were associated with
ROS and the p38-p53 pathway.
Materials and methods
Chemicals and reagents
BRAF inhibitor PLX4032 was purchased from Selleck
Chemicals, and juglone was purchased from Sigma-Aldrich; Merck
KGaA. They were each dissolved in dimethyl sulfoxide (DMSO), and
the final concentration of DMSO for cell culture was <0.1%
(v/v). DMSO,
3-(4′,5′-dimethylthiazol-2′-yl)-2,5-diphenyl-tetrazolium bromide
(MTT) and N-acetyl-L-cysteine (NAC) were obtained from
Sigma-Aldrich; Merck KGaA. 2′,7′-dichlorofluorescein diacetate
(DCFH-DA), 4% paraformaldehyde, 5-ethynyl-29-deoxyuridine (EdU)
cell proliferation kit with Alexa Fluor® 488, and
5,5′,6,6′-tetrachloro-1,1′,3,3′-tetraethyl-midacarbocyanine iodide
(JC-1) staining kit were purchased from Beyotime Institute of
Biotechnology. JNK inhibitor (SP600125) and p38 inhibitor
(SB203580) were obtained from MedChemExpress. The
Annexin-V-phycoerythrin (PE) Apoptosis Detection kit (cat. c559763)
was supplied by BD Biosciences. The antibodies against β-actin
(cat. no. ab8226), poly(ADP-ribose) polymerase (PARP; cat. no.
ab191217), survivin (cat. no. ab76424), Bcl-2 (cat. no. ab32124),
Bax (cat. no. ab32503), phosphorylated (P)-p38 (cat. no. ab195049),
p38 (cat. no. ab170099), P-p53 (cat. no. ab33889) and p53 (cat. no.
ab179477) were obtained from Abcam, and the antibody against
cytochrome c (cat. no. 4280) was purchased from Cell
Signaling Technology, Inc.
Cell lines and culture
Human melanoma SK-MEL-5 and A375 cell lines were
obtained from American Type Culture Collection and maintained in
DMEM (Invitrogen; Thermo Fisher Scientific, Inc.) containing 4 mM
L-glutamine, 3.7 g/l sodium bicarbonate, 4.5 g/l glucose and 10%
foetal bovine serum (Invitrogen; Thermo Fisher Scientific, Inc.).
The cells were maintained in a 5% CO2 humidified
incubator at 37°C. Both SK-MEL-5 and A375 cells carrying the
BRAFV600E mutation are sensitive to vemurafenib treatment (16,17).
A SK-MEL-5 subline with acquired resistance (SK-MEL-5R) to PLX4032
was generated by continuous exposure of parental SK-MEL-5 cells to
gradually increasing concentrations of PLX4032 (from 0.1 µM up to 1
µM) over a period of 3 months. The fold resistance was
intermittently evaluated by cell viability assays and it was found
that SK-MEL-5R showed higher cell viability compared with SK-MEL-5
when treated with different concentrations of PLX4032 (data not
shown). A375 cells with acquired resistant to PLX4032 (A375R) were
generated by the same method, with the concentration of PLX4032
increasing from 0.1 µM up to 2 µM, as previously described
(18).
Cell viability assays
The cytotoxic effects of juglone and/or PLX4032 on
SK-MEL-5R and A375R cells were assessed by MTT assays. Briefly,
4,000 cells in 200 µl cell culture medium/well were seeded into
96-well plates and incubated overnight at 37°C. After NAC (0 and 2
mM), SB203580 (0 and 10 µM) or SP600125 (0 and 10 µM) pre-treatment
for 1 h at 37°C, the cells were treated with juglone (0, 5 or 7.5
µM) and/or PLX4032 (0, 5 or 10 µM) for 24 h at 37°C. DMEM
containing corresponding amounts of 0.1% DMSO were used as vehicle
controls. Then, 20 µl MTT solution (5 mg/ml) was added to each
well, and the cells were incubated for 2 h at 37°C. Subsequently,
the formazan crystals that formed were dissolved with 100 µl DMSO,
and the optical density (OD) was measured at 570 nm on a microplate
spectrophotometer (Infinite 200 PRO; Tecan Group, Ltd.). The cell
viability was determined using the following formula: Cytotoxicity
(%)=(OD treatment/OD vehicle control) ×100. For crystal violet
staining, 2×105 cells/well were seeded into 6-well
plates, followed by exposure to juglone (0 and 5 µM) and/or PLX4032
(0 and 10 µM) for 72 h at 37°C. Then, the cells were fixed with 10%
formalin for 10 min at room temperature and stained with 0.05%
crystal violet solution (CV) in distilled water for 30 min at room
temperature. Finally, the CV was removed, cells were washed twice
with distilled water and images were captured using a Flatbed
Scanner (Canon LiDE 220; Canon Inc.).
Cell proliferation assays
To assess the cell proliferation, an EdU assay kit
was used according to the manufacturer's instructions. Briefly,
2×105 cells/well were seeded in a 6-well plate and
treated with juglone (0 and 5 µM) and/or PLX4032 (0 and 10 µM) for
24 h, followed by incubation with cell culture medium containing 10
µM EdU at 37°C for 2 h. Then, the cells were fixed with 4%
paraformaldehyde for 15 min and permeabilized with 0.3% Triton
X-100 in PBS for 15 min. After washing with PBS, cells were
incubated with click additive solution (supplied with the kit) for
30 min in the dark at room temperature. Finally, cells were washed
with 0.3% Triton X-100 and incubated with Hoechst 33342 for 10 min
at room temperature. After washing with PBS 3 times for 5 min each
time, the images were captured using a fluorescence microscope
(Axio Vert. A1; Zeiss GmbH) at 400× magnification and the
subsequent images were analyzed using ImageJ software (v1.52 g;
National Institutes of Health). The data comprised three
replicates.
Apoptosis assays
Apoptosis was measured using flow cytometry. Cells
were seeded into 6-well culture plates at a density of
2×105 cells/well, and then treated with juglone (0 and 5
µM) and/or PLX4032 (0 and 10 µM) for 24 h at 37°C. Next, cells were
harvested, washed with PBS twice and resuspended in 1X binding
buffer (supplied with kit). Finally, the cells were incubated with
Annexin V-PE and 7-AAD in the dark at room temperature for 15 min,
and analysed using a flow cytometer (Attune NxT; Thermo Fisher
Scientific, Inc.). The data were analysed using FlowJo software
V6.0 (Tree Star, Inc.).
Western blot analysis
Cells were harvested following the pre-treatment
with NAC (0 and 2 mM), SB203580 (0 and 10 µM) or SP600125 (0 and 10
µM) for 1 h at 37°C and treated with juglone (0 and 5 µM) and/or
PLX4032 (0 and 10 µM) for 6 h at 37°C. The cells were then lysed in
RIPA buffer (Beyotime Institute of Biotechnology). The mixture was
centrifuged at 12,000 × g at 4°C for 15 min, and the supernatants
were collected. The protein concentrations were determined using a
bicinchoninic acid protein assay kit (Bio-Rad Laboratories, Inc.).
Next, 30 µg cellular proteins were separated on a 10 or 12%
SDS-polyacrylamide gel and electroblotted onto a PVDF membrane.
Membranes were blocked with 5% milk in Tween-20/Tris-buffered
saline (TBST) for 1 h at room temperature, prior to incubation
overnight at 4°C with 1:1,000 dilution of primary antibodies
against β-actin, PARP, survivin, Bcl-2, Bax, p-p38, p38, p-p53,
p53, JNK, p-JNK and cytochrome c. Membranes were washed with
TBST, followed by 1 h incubation at room temperature in 1:4,000
dilution of horseradish peroxidise-conjugated secondary antibodies
(cat. nos. 7074 and 7076; Cell Signaling Technology, Inc.). After
washing again, the blots were developed using Supersignal West
Femto Chemiluminescent substrate (Thermo Fisher Scientific, Inc.).
Images were acquired by the ChemiDoc™ MP Imaging system (Bio-Rad
Laboratories, Inc.). The band intensities were semi-quantified
using ImageJ software and normalized to β-actin as the loading
control. The western blot assays were repeated three times.
Evaluation of ROS
ROS were detected using the fluorescent probe
DCFH-DA. Briefly, the cells were seeded into 6-well culture plates
at a density of 2×105 cells/well, and then treated with
juglone (0 and 5 µM) and/or PLX4032 (0 and 10 µM) for 2 h. The
cells were then washed with PBS, incubated in serum-free medium,
and loaded with DCFH-DA (10 µM). Following incubation in the dark
at 37°C for 30 min, the cells were washed with PBS and vortexed in
700 µl lysis buffer (90% DMSO:10% PBS)/well for 15 min. The cell
lysis buffer was transferred into a black 96-well Immuno Plate
(Thermo Fisher Scientific, Inc.), and fluorescence was detected
using a fluorescence microplate reader (Infinite 200 PRO; Tecan
Group, Ltd.) at an excitation wavelength of 485 nm, and an emission
wavelength of 535 nm.
Analysis of mitochondrial membrane
potential (ΔΨM)
ΔΨM was evaluated using a JC-1 kit.
Briefly, 1×106 cells seeded in a 6-well plate were
cultured with juglone (0 and 5 µM) and/or PLX4032 (0 and 10 µM) for
3 h. Then, the cells were labelled with JC-1 and incubated for 20
min at 37°C, according to the manufacturer's instructions. Cells
were washed twice, resuspended and detected by flow cytometer
(Attune NxT; Thermo Fisher Scientific, Inc.); the PE and FITC
detection channels were selected at emission wavelengths of 530 and
585 nm, and an excitation wavelength of 488 nm. The data were
analysed by FlowJo software V6.0 (Tree Star, Inc.). Mitochondrial
carbonyl cyanide 3-chlorophenylhydrazone (supplied in kit) was used
as a positive control. The percentage of depolarized mitochondrial
was calculated as previously described (19).
Statistical analysis
All results are presented as the mean ± standard
deviation of at least three independent experiments. Statistical
analysis was performed by SPSS Statistics 21.0 software (IBM
Corp.). Student's t-test was used for comparisons between two
groups, and one-way ANOVA and subsequent Tukey's post-hoc analysis
was applied for comparison of more than two independent groups.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Juglone sensitizes melanoma cells to
PLX4032-induced cytotoxicity
MTT was used to determine the cytotoxic effects of
juglone and/or PLX4032 on melanoma cells. As shown in Fig. 1A and C, the combination of juglone
and PLX4032 in SK-MEL-5R and A375R melanoma cells induced a much
higher cytotoxicity than juglone or PLX4032 alone (P<0.01). In
addition, according to the MTT results, the combination index (CI)
of juglone and PLX4032 cotreatment in SK-MEL-5R cells was
determined using the Chou-Talalay method (20). The CI values varied from 0.34–3.65,
which suggested a strong synergistic effect between juglone and
PLX4032 (Fig. 1B). Crystal violet
staining confirmed that the combination of juglone and PLX4032
further induced cytotoxicity to melanoma cells, compared with
either juglone alone or PLX4032 alone (Fig. 1D). EdU assays also suggested that
cell proliferation was further inhibited after treatment with a
combination of PLX4032 and juglone (Fig. 1E and F).
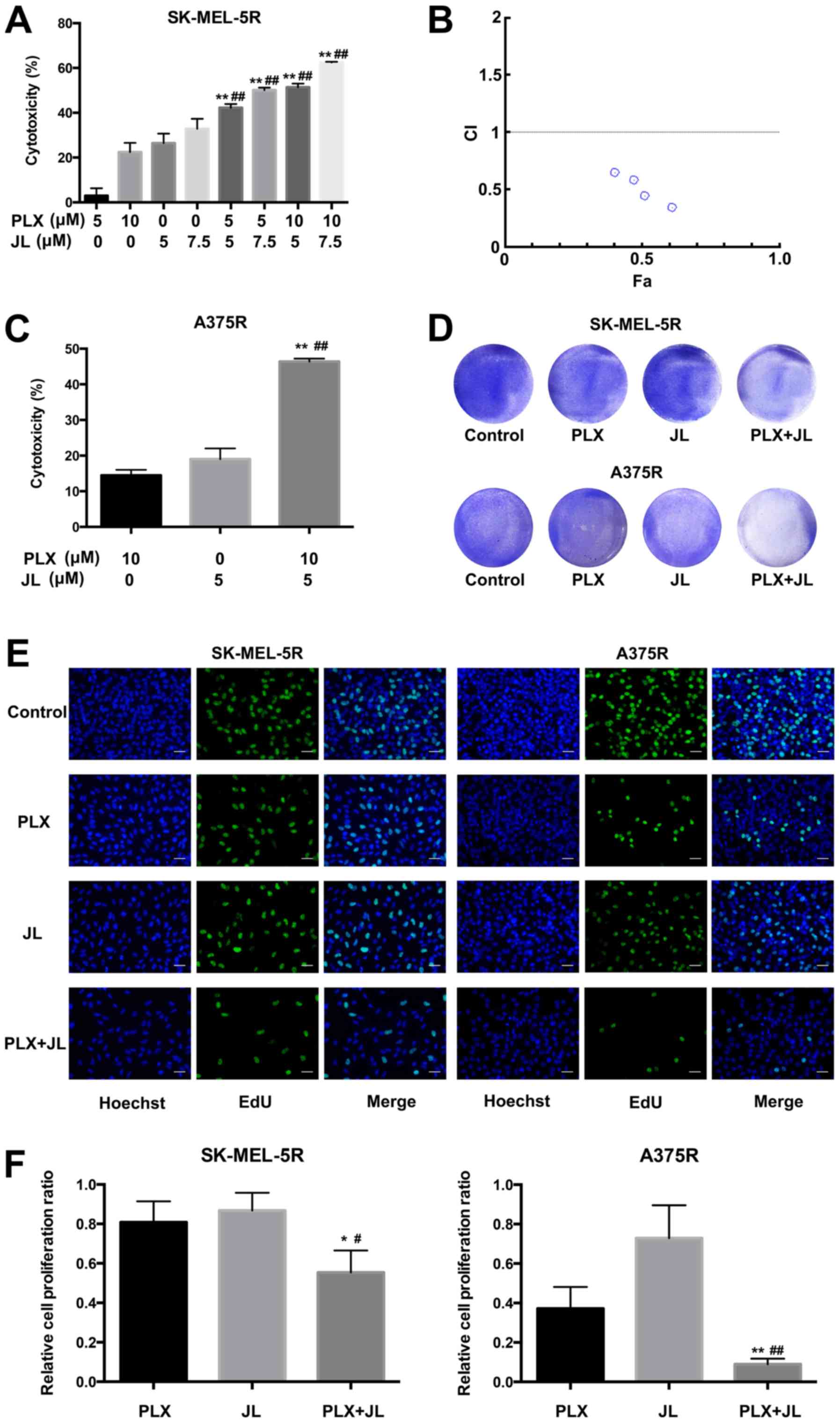 | Figure 1.Juglone sensitizes melanoma cells to
PLX4032-induced cytotoxicity. (A) MTT assay of SK-MEL-5R cells;
SK-MEL-5R cells were treated with juglone (0, 5 and 7.5 µM) and/or
PLX4032 (0, 5 and 10 µM) for 24 h. **P<0.01 vs. same
concentration PLX alone, ##P<0.01 vs. same
concentration JL alone. (B) CI of juglone and PLX4032. (C) MTT
assay of A375R cells; A375R cells were treated with juglone (0 and
5 µM) and/or PLX4032 (0 and 10 µM) for 48 h. **P<0.01 vs. PLX,
##P<0.01 vs. JL. (D) Crystal violet staining;
SK-MEL-5R and A375R cells were exposed to juglone (0 and 5 µM)
and/or PLX4032 (0 and 10 µM) for 72 h. The cells were then fixed
and stained with crystal violet solution. (E) EdU assay; cells were
double-stained with EdU (green) and Hoechst 33342 (blue). Scale
bar, 20 µm. (F) Relative cell proliferation rates were analyzed
according to the EdU assay results. **P<0.01, *P<0.05 vs.
PLX; ##P<0.01, #P<0.05 vs. JL. CI,
combination index; EdU, 5-ethynyl-29-deoxyuridine; JL, juglone;
PLX, PLX4032. |
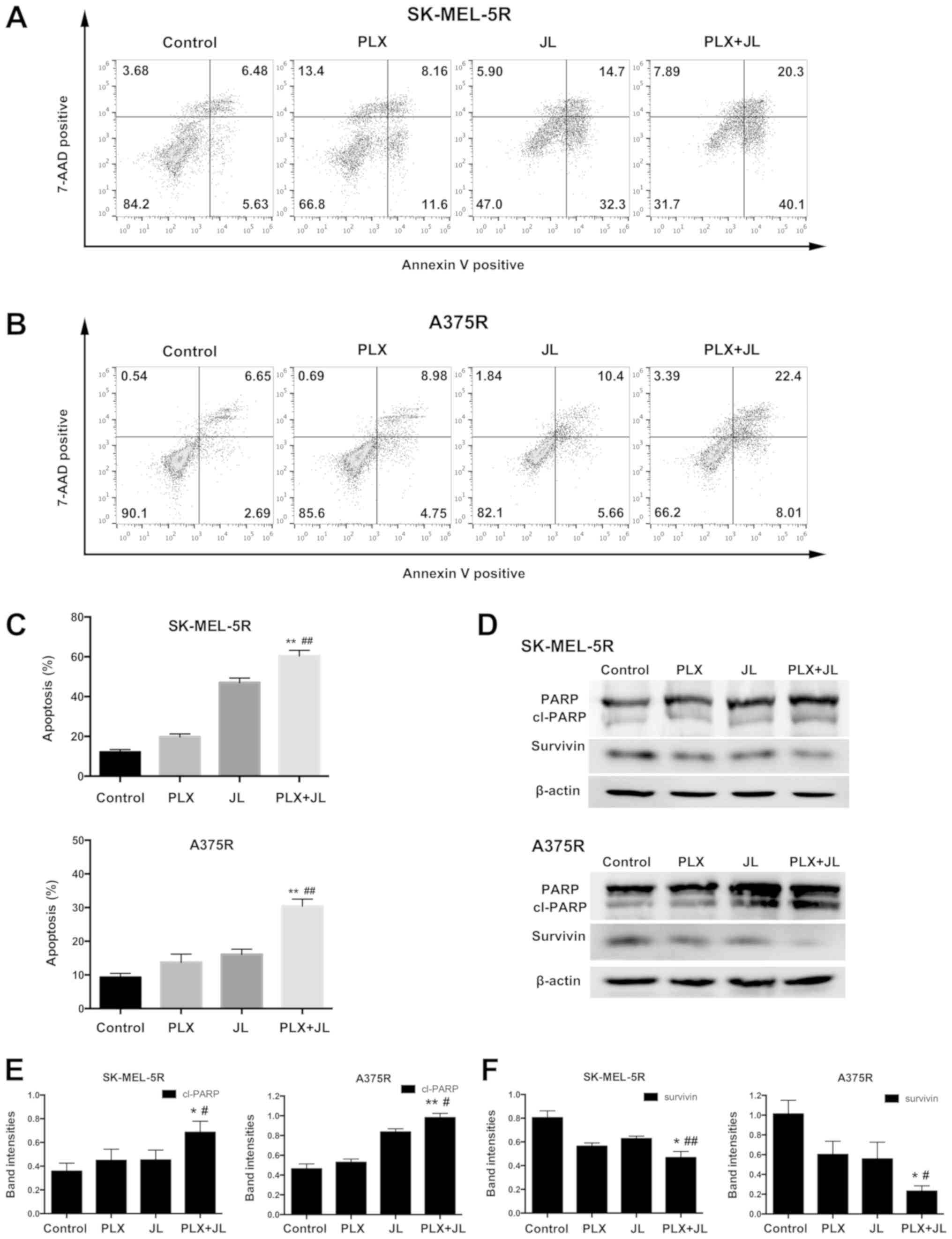
Juglone potentiates PLX4032-induced
apoptosis in melanoma
To further demonstrate the synergistic effect of
juglone and PLX4032, flow cytometry analysis was performed to
measure the percentages of both Annexin
V+/7-AAD− (early apoptotic cells) and Annexin
V+/7-AAD+ (late apoptotic/necrotic cells)
cells. In SK-MEL-5R cells, PLX4032 alone induced only minor
apoptosis. However, the combination of juglone and PLX4032
exhibited a much higher rate of apoptosis than either PLX4032 or
juglone alone (Fig. 2A and C;
P<0.01). Similar results were observed in A375R cells (Fig. 2B and C; P<0.01).
PARP is one of the terminal pro-apoptotic proteins.
The cleaved forms of PARP are its active forms (21). Survivin, on the other hand,
inhibits apoptosis (22,23). As demonstrated by western blotting,
the protein expression of cleaved PARP in the juglone and PLX4032
combination group was the highest, while the protein expression of
survivin was the lowest, as compared with levels in either the
PLX4032 or juglone alone groups (Fig.
2D-F).
Juglone enhances PLX4032-induced
apoptosis through the mitochondrial pathway
ΔΨM decline is one of the earliest
indicators of intrinsic apoptosis. In the present study, the
fluorescent probe JC-1 was used to determine the effect of juglone
on ΔΨM in SK-MEL-5R cells. When the mitochondrial
potential is normal, JC-1 forms a polymer in the mitochondrial
matrix and exhibits red fluorescence with an emission of 590 nm;
however, when the ΔΨM is downregulated, JC-1 changes
into a monomeric form that yields green fluorescence with an
emission of 530 nm, so the PE and FITC channel were selected to
measure the fluorescence intensity of JC-1 dye in the cells
(24). As shown in Fig. 3A and B, juglone caused a marked
decrease in ΔΨM compared with the vehicle control.
Moreover, the combination of juglone and PLX4032 further decreased
the ΔΨM, and the percentage of cells with low potential
was increased from 44.5% in the juglone alone group to 55.8% when
cotreated with juglone and PLX4032.
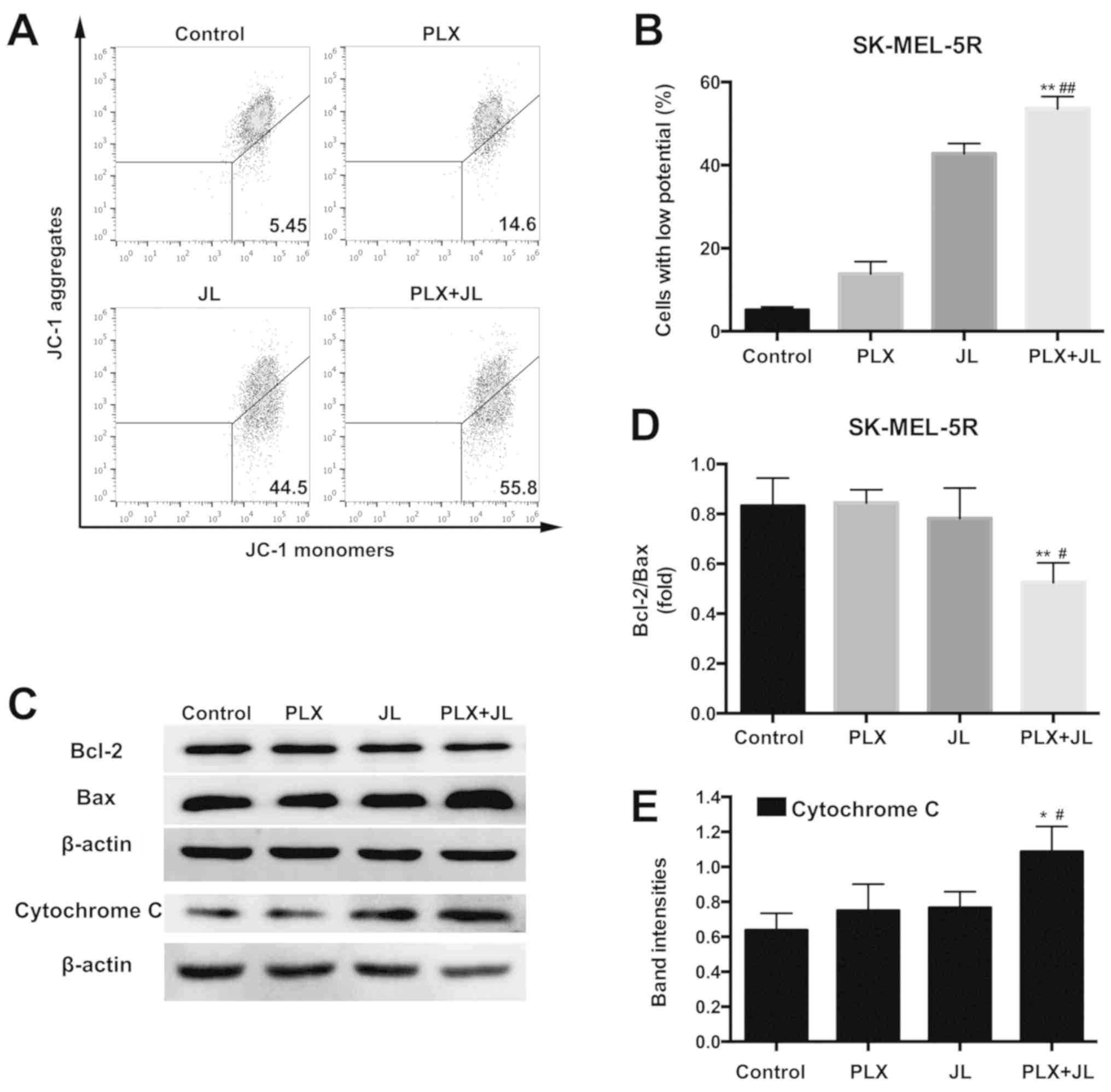 | Figure 3.Juglone enhances PLX4032-induced
mitochondrial apoptosis. (A) Mitochondrial membrane potential
analysed by JC-1 fluorescence with flow cytometry. (B)
Quantification of cells with low potential. **P<0.01 vs. PLX
alone; ##P<0.01 vs. JL alone. (C) Representative
western blot images of Bcl-2, Bax and cytochrome c. (D)
Bcl-2/Bax band intensity ratio. **P<0.01 vs. PLX alone;
#P<0.05 vs. JL alone. (E) Semi-quantification of band
intensities of cytochrome c. *P<0.05 vs. PLX;
#P<0.05 vs. JL. JC-1,
5,5′,6,6′-etrachloro-1,1′,3,3′-tetraethyl-imidacarbocyanine iodide;
PLX, PLX4032; JL, juglone. |
The pro-apoptotic protein Bax and anti-apoptotic
protein Bcl-2 serve central roles in the mitochondria-dependent
apoptotic pathway; Bax can be translocated to the outer
mitochondrial membrane following a death signal, where it promotes
a permeabilization that favors the release of different apoptogenic
factors, such as cytochrome c (25). In addition, the ability of Bax to
form channels on the mitochondrial membrane can be inhibited by
Bcl-2 (26). As demonstrated by
western blot analysis (Fig. 3C),
the protein expression of Bax was notably increased after
cotreatment with juglone and PLX4032, while no changes were seen in
the juglone or PLX4032 only groups. A reduction in the Bcl-2/Bax
ratio was observed after the combination drug treatment in
SK-MEL-5R cells (Fig. 3D). The
combination of PLX4032 and juglone also increased the protein
expression of cytochrome c (Fig. 3C and E).
Juglone increases the cellular ROS
level and activated p38, p53 and JNK in SK-MEL-5R cells
Since mitochondrial apoptosis is associated with the
generation of ROS (27), the
production of intracellular ROS was examined using the
oxidant-sensitive fluorescent probe DCFH-DA. The amount of ROS was
analysed in cells treated with juglone and/or PLX4032 using a
fluorescence microplate reader. Both PLX4032 or juglone alone
increased the level of ROS. However, the level of ROS was
significantly higher in the juglone and PLX4032 cotreatment group
compared with the juglone or PLX4032 alone groups (Fig. 4A).
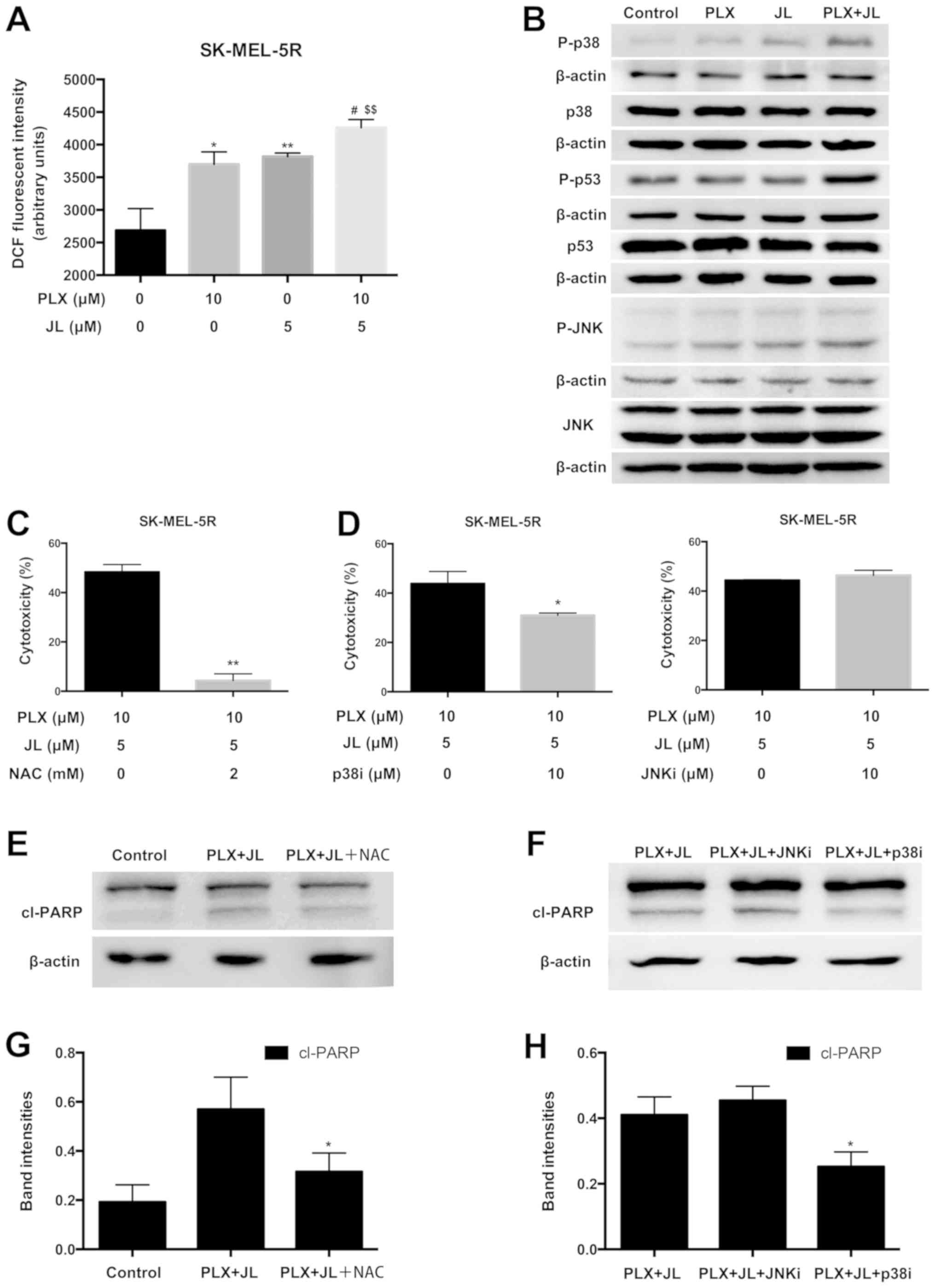 | Figure 4.Juglone in combination with PLX4032
increases the production of ROS and activates p38, p53 and JNK. (A)
Intracellular ROS levels measured by DCFH-DA. SK-MEL-5R cells were
treated with juglone (0 and 5 µM) and/or PLX4032 (0 and 10 µM) for
2 h, fluorescence intensity was monitored by a fluorescence
microplate reader. **P<0.01, *P<0.05 vs. control;
#P<0.05 vs. PLX; $$P<0.01, vs. JL. (B)
Representative western blot images of p-p38, p38, p-p53, p53, p-JNK
and JNK. (C) NAC pre-treatment partially reverses cytotoxic effect
of juglone. SK-MEL-5R cells were pre-treated with NAC (0 and 2 mM)
for 1 h, followed by juglone and PLX4032 cotreatment for 24 h. The
cytotoxicity was measured using MTT assays. **P<0.01 vs. NAC
non-pre-treatment. (D) p38i partially reversed the cytotoxic effect
of juglone and PLX4032. SK-MEL-5R cells were pre-treated with p38i
(0 and 10 µM) or JNKi (0 and 10 µM) for 1 h, followed by juglone
and PLX4032 cotreatment for 24 h. Cytotoxicity was measured using
MTT assays. *P<0.05. (E) Representative western blot images of
cl-PARP from cells pre-treated with NAC. (F) Representative western
blot images of cl-PARP from cells pre-treated with p38i and JNKi.
(G) Semi-quantification of band intensities of cl-PARP from part
(E). (H) Semi-quantification of band intensities of cl-PARP from
part (F). *P<0.05 vs. PLX and JL cotreatment. NAC,
N-acetyl-L-cysteine; PARP, poly(ADP-ribose) polymerase; DCFH-DA,
2′,7′-dichlorofluorescein diacetate; DCF,
2′,7′-dichlorofluorescein; JL, juglone; ROS, reactive oxygen
species; P-, phosphorylated; Cl-, cleaved; p38i, p38 inhibitor;
JNKi, JNK inhibitor. |
Previous evidence has indicated that ROS can also
mediate cell death via key modulators, such as p38
mitogen-activated protein kinase (MAPK) and p53 (28,29).
To test whether the p38-p53 pathway was activated by juglone and
PLX4032 cotreatment, western blotting was used to measure the
protein expression of p38, p53 and their phosphorylated
counterparts. As demonstrated in Fig.
4B, in SK-MEL-5R cells, juglone and PLX4032 cotreatment
markedly increased p38 and p53 phosphorylation compared with
treatment of PLX4032 or juglone alone. JNK (Thr183/Tyr185)
activation was also observed after juglone and PLX4032
cotreatment.
NAC and p38 inhibitor pre-treatment
partially reverse juglone and PLX4032 cotreatment-induced
cytotoxicity
After cells were pre-treated with NAC (a ROS
scavenger), or SB203580 (a p38 inhibitor), juglone and PLX4032
cotreatment-induced cytotoxicity was partially reversed (Fig. 4C and D). Furthermore, as
demonstrated by western blotting, the protein expression of cleaved
PARP in the NAC or p38 inhibitor pre-treatment group was lower than
the protein expression in the juglone/PLX4032 combination group
without NAC or p38 inhibitor pre-treatment (Fig. 4E-H; P<0.05). However,
pre-treatment with SP600125 (a JNK inhibitor), failed to reverse
the cytotoxicity induced by juglone and PLX4032 cotreatment
(Fig. 4D). This was consistent
with the inability of JNK inhibitor to reduce the levels of cl-PARP
(Fig. 4F and H). These results
suggested that ROS production and p38 activation play important
roles in juglone-induced PLX4032 sensitization.
Discussion
It has been reported that juglone displays
anti-tumour activities in a variety of tumour cell lines (11,14,30),
and that treatment of B16F10 melanoma cells with juglone results in
a concentration-dependent reduction in cell viability. For example,
after 24 h exposure to 5 µM juglone, cytotoxicity was increased by
only ~20% compared with the vehicle control (11). This is in accordance with the
finding in the present study that juglone at a low concentration
possesses weak activity against BRAF-mutant melanoma cells.
Juglone has displayed potent cytotoxic properties
against chemo-resistant and radio-resistant tumour cell lines both
in vitro and in vivo (12,31).
In addition, it has been considered as a sensitizing agent for
overcoming trastuzumab resistance in human breast cancer SKBR3
cells (32). Combination therapy
with a BRAF inhibitor and juglone might provide a novel strategy
for overcoming BRAF inhibitor resistance. In the present study, to
the best of our knowledge, it was reported for the first time that
juglone enhanced the response to a BRAF inhibitor in BRAF
inhibitor-resistant melanoma cells.
Apoptosis is a form of complex signalling-controlled
cell death. The two main pathways of apoptosis are the extrinsic
pathway and the intrinsic pathway, which is also called the
mitochondrial pathway. The intrinsic pathway that initiates
apoptosis involves a diverse array of non-receptor-mediated
stimuli. These produce intracellular signals that cause changes in
the inner mitochondrial membrane (33). The anti-cancer effects of juglone
can be partially attributed to mitochondrial apoptosis (30,34).
Juglone has been found to markedly alter the levels of Bcl-2 and
Bax, thereby releasing cytochrome c into the cell cytoplasm,
and inducing subsequent caspase activation and activation of PARP
(30,34). In the present study, it was found
that juglone potentiated BRAF inhibitor-induced apoptosis in both
A375R and SK-MEL-5R cells, which was accompanied by a decline in
ΔΨM, and a reduction of the Bcl-2/Bax ratio.
ROS are one of the stimuli that can induce the
mitochondrial apoptotic pathway. An increase in ROS leads to the
opening of the mitochondrial membrane permeability transition pore,
which destroys the integrity of the mitochondrial membrane,
inducing ΔΨM loss (33). Meanwhile, damaged mitochondria
produce more ROS and accelerate apoptosis. It was revealed that the
cytotoxicity of juglone is generally attributed to its ability to
induce redox cycling with consequent ROS, leading to oxidative
stress-mediated cell death (35).
Elevated ROS levels have been observed after juglone treatment in
multiple human cancer lines, including glioma, gastric cancer and
melanoma cells (14,30,36).
In the present study, juglone alone significantly increased the
levels of ROS in BRAF inhibitor-resistant melanoma cells. Moreover,
the level of ROS was higher in the juglone and PLX4032 cotreatment
group than in the single agent treatment groups. Pre-treatment with
NAC, a ROS scavenger, partially reversed juglone and PLX4032
combination-induced cytotoxicity. Therefore, it was concluded that
the synergistic effects of juglone and a BRAF inhibitor in melanoma
cells were partially mediated by ROS.
In addition to disrupting the mitochondrial
membrane, ROS act as secondary messengers to trigger apoptotic
signals (37). The downstream
molecules affected by ROS include MAPK, PI3K/AKT, phospholipase γ1,
NF-κB and Janus kinase, among which the MAPK pathway is a complex
family that transmits extracellular signals to the intracellular
environment. MAPK pathways include ERK, JNK and p38. Unlike ERK,
both JNK and p38 mediate tumour cell apoptosis. JNK and p38 and are
also more stress-responsive than ERK, and are often termed
stress-activated protein kinases (38). p53 is a direct downstream effector
of p38 (28,39), which causes cell cycle arrest and
induces cellular senescence and apoptosis (40). In the present study, it was
demonstrated that the p38-p53 pathway was activated after juglone
and PLX4032 cotreatment.
A previous study has demonstrated that juglone
induced HeLa cells to undergo apoptosis by activating the JNK/c-Jun
pathway (41). Previously, a JNK
activator was reported to exert anti-tumour activity in
vemurafenib-resistant melanoma cells, and this indicated that the
induction of apoptosis through the activation of the JNK pathway
might represent a novel strategy to overcome resistance to
vemurafenib (42). In the present
study, it was demonstrated that juglone and PLX4032 cotreatment
activated JNK. However, unlike p38 inhibition, inhibition of JNK
could not reverse the juglone-induced enhanced sensitivity to
PLX4032. Therefore, this suggested that p38, and not JNK, plays a
crucial role in the induction of apoptosis following juglone and
PLX4032 cotreatment. Juglone has been reported to possess low
cytotoxicity in human peripheral blood mononuclear cells (43). However, it is unclear whether
juglone combined with a BRAF inhibitor is toxic in normal cells.
Further in vivo studies are necessary to determine the
efficacy and safety of juglone and BRAF inhibitor cotreatment in
BRAF inhibitor-resistant melanoma.
In conclusion, juglone potentiated the BRAF
inhibitor-induced apoptosis in BRAF inhibitor-resistant melanoma
cells, and these effects were partially mediated through ROS and
the p38-p53 pathway, thereby suggesting the potential of juglone as
a sensitizer to BRAF inhibitors in the treatment of melanoma.
Acknowledgements
Not applicable.
Funding
The present study was supported by grants from the
National Natural Science Foundation of China (grant no. 81673917)
and Shanghai Science and Technology Committee (grant no.
13JC1401401).
Availability of data and materials
The datasets used and/or analysed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
JX and JW designed the research and interpreted the
data; ZL and XL performed the experiments and wrote the manuscript.
ML, JC and SH analyzed the data. All authors read and approved the
final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2019. CA Cancer J Clin. 69:7–34. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Whiteman DC, Green AC and Olsen CM: The
growing burden of invasive melanoma: Projections of incidence rates
and numbers of new cases in six susceptible populations through
2031. J Invest Dermatol. 136:1161–1171. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Flaherty KT, Puzanov I, Kim KB, Ribas A,
McArthur GA, Sosman JA, O'Dwyer PJ, Lee RJ, Grippo JF, Nolop K and
Chapman PB: Inhibition of mutated, activated BRAF in metastatic
melanoma. N Engl J Med. 363:809–819. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sosman JA, Kim KB, Schuchter L, Gonzalez
R, Pavlick AC, Weber JS, McArthur GA, Hutson TE, Moschos SJ,
Flaherty KT, et al: Survival in BRAF V600-mutant advanced melanoma
treated with vemurafenib. N Engl J Med. 366:707–714. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Obenauf AC, Zou Y, Ji AL, Vanharanta S,
Shu W, Shi H, Kong X, Bosenberg MC, Wiesner T, Rosen N, et al:
Therapy-induced tumour secretomes promote resistance and tumour
progression. Nature. 520:368–372. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Luebker SA and Koepsell SA: Diverse
mechanisms of BRAF inhibitor resistance in melanoma identified in
clinical and preclinical studies. Front Oncol. 9:2682019.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Long GV, Stroyakovskiy D, Gogas H,
Levchenko E, de Braud F, Larkin J, Garbe C, Jouary T, Hauschild A,
Grob JJ, et al: Combined BRAF and MEK inhibition versus BRAF
inhibition alone in melanoma. N Engl J Med. 371:1877–1888. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Long GV, Weber JS, Infante JR, Kim KB,
Daud A, Gonzalez R, Sosman JA, Hamid O, Schuchter L, Cebon J, et
al: Overall survival and durable responses in patients with BRAF
V600-mutant metastatic melanoma receiving dabrafenib combined with
trametinib. J Clin Oncol. 34:871–878. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Sugie S, Okamoto K, Rahman KW, Tanaka T,
Kawai K, Yamahara J and Mori H: Inhibitory effects of plumbagin and
juglone on azoxymethane-induced intestinal carcinogenesis in rats.
Cancer Lett. 127:177–183. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Polonik SG, Prokof'eva NG, Agafonova IG
and Uvarova NI: Antitumor and immunostimulating activity of
5-hydroxy-1, 4-naphthoquinone (juglone) O-and S-acetylglycosides.
Pharm Chem J. 37:397–398. 2003. View Article : Google Scholar
|
|
11
|
Aithal KB, Kumar SM, Rao NB, Udupa N and
Rao SB: Juglone, a naphthoquinone from walnut, exerts cytotoxic and
genotoxic effects against cultured melanoma tumor cells. Cell Biol
Int. 33:1039–1049. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Aithal KB, Kumar S, Rao BN, Udupa N and
Rao SBS: Tumor growth inhibitory effect of juglone and its
radiation sensitizing potential: In vivo and in vitro studies.
Integr Cancer Ther. 11:68–80. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liu X, Chen Y, Zhang Y, Du J, Lv Y, Mo S,
Liu Y, Ding F, Wu J and Li J: Juglone potentiates TRAIL-induced
apoptosis in human melanoma cells via activating the ROS-p38-p53
pathway. Mol Med Rep. 16:9645–9651. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Xu HL, Yu XF, Qu SC, Qu XR, Jiang YF and
Sui da Y: Juglone, from Juglans mandshruica Maxim, inhibits growth
and induces apoptosis in human leukemia cell HL-60 through a
reactive oxygen species-dependent mechanism. Food Chem Toxicol.
50:590–596. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Corazao-Rozas P, Guerreschi P, Jendoubi M,
André F, Jonneaux A, Scalbert C, Garçon G, Malet-Martino M,
Balayssac S, Rocchi S, et al: Mitochondrial oxidative stress is the
Achille's heel of melanoma cells resistant to Braf-mutant
inhibitor. Oncotarget. 4:1986–1998. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Feng Z, Kochanek S, Close D, Wang L,
Srinivasan A, Almehizia AA, Iyer P, Xie XQ, Johnston PA and Gold B:
Design and activity of AP endonuclease-1 inhibitors. J Chem Biol.
8:79–93. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Liu F, Cao J, Wu J, Sullivan K, Shen J,
Ryu B, Xu Z, Wei W and Cui R: Stat3-targeted therapies overcome the
acquired resistance to vemurafenib in melanomas. J Invest Dermatol.
133:2041–2049. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Guerriero L, Palmieri G, De Marco M, Cossu
A, Remondelli P, Capunzo M, Turco MC and Rosati A: The
anti-apoptotic BAG3 protein is involved in BRAF inhibitor
resistance in melanoma cells. Oncotarget. 8:80393–80404. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Liu L, Li T, Tan J, Fu J, Guo Q, Ji H and
Zhang Y: NG as a novel nitric oxide donor induces apoptosis by
increasing reactive oxygen species and inhibiting mitochondrial
function in MGC803 cells. Int Immunopharmacol. 23:27–36. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chou TC: Drug combination studies and
their synergy quantification using the Chou-Talalay method. Cancer
Res. 70:440–446. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Soldani C and Scovassi AI:
Poly(ADP-ribose) polymerase-1 cleavage during apoptosis: An update.
Apoptosis. 7:321–328. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ambrosini G, Adida C, Sirugo G and Altieri
DC: Induction of apoptosis and inhibition of cell proliferation by
survivin gene targeting. J Biol Chem. 273:11177–11182. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Shin S, Sung BJ, Cho YS, Kim HJ, Ha NC,
Hwang JI, Chung CW, Jung YK and Oh BH: An anti-apoptotic protein
human survivin is a direct inhibitor of caspase-3 and-7.
Biochemistry. 40:1117–1123. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Tang C, Liang J, Qian J, Jin L, Du M, Li M
and Li D: Opposing role of JNK-p38 kinase and ERK1/2 in hydrogen
peroxide-induced oxidative damage of human trophoblast-like JEG-3
cells. Int J Clin Exp Pathol. 7:959–968. 2014.PubMed/NCBI
|
|
25
|
Renault TT and Manon S: Bax: Addressed to
kill. Biochimie. 93:1379–1391. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Antonsson B, Conti F, Ciavatta A,
Montessuit S, Lewis S, Martinou I, Bernasconi L, Bernard A, Mermod
JJ, Mazzei G, et al: Inhibition of Bax channel-forming activity by
Bcl-2. Science. 277:370–372. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Simon HU, Haj-Yehia A and Levi-Schaffer F:
Role of reactive oxygen species (ROS) in apoptosis induction.
Apoptosis. 5:415–418. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Perfettini JL, Castedo M, Nardacci R,
Ciccosanti F, Boya P, Roumier T, Larochette N, Piacentini M and
Kroemer G: Essential role of p53 phosphorylation by p38 MAPK in
apoptosis induction by the HIV-1 envelope. J Exp Med. 201:279–289.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wu GS: The functional interactions between
the MAPK and p53 signaling pathways. Cancer Biol Ther. 3:156–161.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ji YB, Qu ZY and Zou X: Juglone-induced
apoptosis in human gastric cancer SGC-7901 cells via the
mitochondrial pathway. Exp Toxicol Pathol. 63:69–78. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Segura-Aguilar J, Jönsson K, Tidefelt U
and Paul C: The cytotoxic effects of 5-OH-1,4-naphthoquinone and
5,8-diOH-1,4-naphthoquinone on doxorubicin-resistant human leukemia
cells (HL-60). Leuk Res. 16:631–637. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Sajadimajd S and Yazdanparast R:
Sensitizing effect of juglone is mediated by down regulation of
Notch1 signaling pathway in trastuzumab-resistant SKBR3 cells.
Apoptosis. 22:135–144. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Elmore S: Apoptosis: A review of
programmed cell death. Toxicol Pathol. 35:495–516. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Xu HL, Yu XF, Qu SC, Zhang R, Qu XR, Chen
YP, Ma XY and Sui DY: Anti-proliferative effect of Juglone from
Juglans mandshurica Maxim on human leukemia cell HL-60 by inducing
apoptosis through the mitochondria-dependent pathway. Eur J
Pharmacol. 645:14–22. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Ahmad T and Suzuki YJ: Juglone in
oxidative stress and cell signaling. Antioxidants (Basel). 8(pii):
E912019. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Sidlauskas K, Sidlauskiene R, Li N and
Liobikas J: 5-Hydroxy-1,4-naphthalenedione exerts anticancer
effects on glioma cells through interaction with the mitochondrial
electron transport chain. Neurosci Lett. 639:207–214. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Ziech D, Franco R, Pappa A and
Panayiotidis MI: Reactive oxygen species (ROS)-induced genetic and
epigenetic alterations in human carcinogenesis. Mutat Res.
711:167–173. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Martindale JL and Holbrook NJ: Cellular
response to oxidative stress: Signaling for suicide and survival. J
Cell Physiol. 192:1–15. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Smeenk L, Van Heeringen SJ, Koeppel M,
Gilbert B, Janssen-Megens E, Stunnenberg HG and Lohrum M: Role of
p53 serine 46 in p53 target gene regulation. PLoS One.
6:e175742011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Levine AJ and Oren M: The first 30 years
of p53: Growing ever more complex. Nat Rev Cancer. 9:749–758. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Lu Z, Chen H, Zheng XM and Chen ML:
Experimental study on the apoptosis of cervical cancer Hela cells
induced by juglone through c-Jun N-terminal kinase/c-Jun pathway.
Asian Pac J Trop Med. 10:572–575. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Graziani G, Artuso S, De Luca A, Muzi A,
Rotili D, Scimeca M, Atzori MG, Ceci C, Mai A, Leonetti C, et al: A
new water soluble MAPK activator exerts antitumor activity in
melanoma cells resistant to the BRAF inhibitor vemurafenib. Biochem
Pharmacol. 95:16–27. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Montenegro RC, Araújo AJ, Molina MT,
Marinho Filho JD, Rocha DD, Lopéz-Montero E, Goulart MO, Bento ES,
Alves AP, Pessoa C, et al: Cytotoxic activity of naphthoquinones
with special emphasis on juglone and its 5-O-methyl derivative.
Chem Biol Interact. 184:439–448. 2010. View Article : Google Scholar : PubMed/NCBI
|