Introduction
Hepatitis B virus (HBV) is a Hepadnaviridae virus,
which leads to acute and chronic hepatitis in humans (1). HBV, not only causes persistent or
transient infection in the liver, but also infects extrahepatic
tissue, including the pancreas, bile duct, lymphoid system and
kidneys (2). Moreover, HBV
infection is closely associated with chronic glomerulonephritis
(HBV-GN); however, the molecular mechanisms of HBV during HBV-GN
are not completely understood (3,4).
HBV regulatory hepatitis B virus X (HBX) protein is
required for HBV replication (5,6). A
number of studies have reported that HBX promotes cell
proliferation during HBV infection-associated hepatocarcinoma
(7–9). Human renal tubular epithelial cells
(RTECs) are a major part of the renal tubular interstitium and play
an active role in renal inflammation (10). RTEC apoptosis is closely associated
with the dysfunction of the tubular interstitium, which
subsequently leads to the progression of HBV-GN (10). HBX is primarily expressed in RTEC
cells and causes RTEC apoptosis and renal tube lesions in patients
with HBV-GN (11,12). Furthermore, HBX has been reported
to suppress the proliferation of rat RTECs (13), and HBX overexpression accelerates
the cell cycle progression from G1 to S phase in primary
RTECs, leading to cell cycle arrest in the S phase (14). Therefore, identifying a HBX
inhibitor may provide a novel therapeutic for HBV-GN. However, the
precise molecular network of HBX in human RTECs is not completely
understood.
Echinacoside (ECH) is a natural compound extracted
from Cistanche plants, which shows antiapoptotic and
anti-inflammatory effects (15,16).
A previous study reported that ECH induces the proliferation and
prevents the apoptosis of intestinal epithelial cells (17). Furthermore, ECH has been reported
to accelerate bone regeneration by increasing the proliferation of
osteoblast cells (18). ECH can
also decrease HBV replication, antigen expression and inflammation
in rat intestine epithelial cells (16,19).
However, the biological effect of ECH on RTECs has not been fully
elucidated.
Triggering receptor expressed on myeloid cells 2
(TREM2), a member of the transmembrane receptor family, plays a
role in maintaining adequate microglial metabolism during
Alzheimer's disease (20,21). A previous study indicated that
TREM2 overexpression promoted the proliferation of glioma cells
(22). By contrast, TREM2
knockdown decreases the translocation of NF-κB to the nucleus in
degenerative human nucleus pulposus cells (23). Furthermore, TREM2 has been
identified as a novel therapeutic target for human intervertebral
disc degeneration (23). However,
the biological function of TREM2 in human RTECs has not been
reported.
In the present study, HBX overexpression (oeHBX) was
induced in human RTECs (HK-2 cell line) and subsequently, the
effect of ECH on transfected oeHBX cells was investigated. The aim
of the present study was to investigate the functions of HBX and
the potential value of ECH as an effective therapeutic agent for
HBV-GN.
Materials and methods
Cell culture
The human RTEC cell line HK-2 was purchased from
Shanghai Yaji Biotechnology Co., Ltd. HK-2 cells were cultured in
DMEM/F12 media (cat. no. SH30023.01B; HyClone; Cytiva) supplemented
with 10% foetal bovine serum (cat. no. 16000-044; Gibco; Thermo
Fisher Scientific, Inc.), 2 mM L-glutamine and 1%
penicillin/streptomycin (cat. no. P1400-100; Beijing Solarbio
Science & Technology Co., Ltd.) at 37°C with 5%
CO2.
RNA isolation and reverse
transcription-quantitative PCR (RT-qPCR)
Total RNA was extracted from HK-2 cells using
TRIzol® (cat. no. 1596-026; Invitrogen; Thermo Fisher
Scientific, Inc.), according to the manufacturer's protocol. Total
RNA was reverse transcribed into cDNA using the First Strand cDNA
Synthesis kit according to the manufacturer's protocol (cat. no.
K1622; Thermo Fisher Scientific, Inc.). qPCR was performed using
SYBR-Green premix according to the manufacturer's protocol (cat.
no. K0223; Thermo Fisher Scientific, Inc.)on an ABI-7300 real-time
detector (Applied Biosystems; Thermo Fisher Scientific, Inc.). The
conditions of PCR were: 95°C for 10 min followed by 40 cycles of
95°C for 15 sec, 60°C for 45 sec. Three repeats were needed for
each reaction. The following primer pairs were used for the qPCR:
HBX forward, 5′-GGCTGCTAGGTTGTACTG-3′ and reverse,
5′-CAGAGGTGAAGCGAAGTG-3′; TREM2 forward, 5′-TGGCACTCTCACCATTACG-3′
and reverse, 5′-CCTCCCATCATCTTCCTTCAC-3′; and GAPDH forward,
5′-AATCCCATCACCATCTTC-3′ and reverse, 5′-AGGCTGTTGTCATACTTC-3′.
mRNA levels were quantified using the 2−ΔΔCq method and
normalized to the internal reference gene GAPDH (24).
Overexpression and knockdown
To induce the overexpression of HBX and TREM2, the
full-length HBX or TREM2 cDNA sequences were inserted into the
pLVX-puro vector (Clontech Laboratories, Inc.) by double digestion
(HindIII and EcoRI). Subsequently, the recombined
plasmid (1.5 µg) was transfected into HK-2 cells
(2×105). The mock plasmid (an empty vector, 1 µg) was
transiently transfected into HK-2 cells (2×105) as a
negative control (oeNC) using Lipofectamine® 2000 (cat.
no. 11668-027, Invitrogen, Thermo Fisher Scientific, Inc.). ECH
(cat. no. 82854-37-3; Biopurify Phytochemicals, Ltd.) was dissolved
in DMSO (cat. no. D2650; Sigma-Aldrich; Merck KGaA) at the
concentration of 1, 2.5, 5, 10, 20 and 50 mg/l respectively; and
added to the culture of transfected oeHBX or oeTREM2 cells. The
subsequent experimentation began 48 h later.
To knock down TREM2 expression, three small
interfering (si)RNAs (siTREM2-1, siTREM2-2 and siTREM2-3; 20 µM)
targeting TREM2 were synthesized (Shanghai Majorbio Bio-Pharm
Technology Co. Ltd.) and subsequently transfected into HK-2 cells
(2×105) using Lipofectamine® 2000 according
to the manufacturer's protocol (Invitrogen; Thermo Fisher
Scientific, Inc.). A non-specific scrambled siRNA sequence
(5′UUCUCCGAACGUGUCACGU3, 25 nM) si-negative control (NC) was used
as the negative control. The targeted locus and sequences of the
TREM2 siRNAs are provided in Table
SI. The subsequent experimentation was performed 48 h
later.
Western blotting
Total protein was extracted from HK-2 cells using
RIPA lysis buffer (cat. no. BYL40825; JRDUN Biotech Co., Ltd.).
Total protein was quantified using the Bicinchoninic Acid assay kit
(cat. no. PICPI23223; Thermo Fisher Scientific, Inc.) according to
the manufacturer's protocol and 20 µg protein/lane was separated
via SDS-PAGE on a 15% gel. The densitometry of samples were
determined using a microplate reader (DNM-9602, Pulangxin).
Subsequently, the separated proteins were transferred onto a PVDF
membrane. Following blocking with 5% non-fat dry milk for 1 h at
room temperature, the membrane was incubated with primary
antibodies (Table SII) at 4°C
overnight and re-incubated with the secondary antibody anti-mouse
IgG (1:1,000, cat. no. A0216, Beyotime Institute of Biotechnology)
for 1 h at 37°C. The immunoreactive protein bands were visualized
using an ECL system (Cytiva). Blots were performed in triplicate.
GAPDH and H3 were used as the loading controls for
cytoplasmic and nuclear proteins, respectively.
Cell proliferation
The Cell Counting Kit-8 (CCK-8; cat. no. CP002; SAB
Biotherapeutics, Inc.)assay was performed to quantify cell
proliferation. Briefly, a total of 3×104 cells/well were
seeded into 96-well plates and CCK-8 reagent was added, according
to the manufacturer's protocol. Subsequently, the optical density
(OD) 450 value of each well was determined by a microplate reader
at 0, 12, 24 and 48 h. The assay was performed in triplicate.
Cell apoptosis
Briefly, an Annexin V-FITC Apoptosis Detection kit
(cat. no. C1063; Beyotime Institute of Biotechnology) was used to
examine the number of apoptotic HK-2 cells, according to the
manufacturer's protocol. After 48 h post-transfection, the cells
were performed using a flow cytometer (Accuri C6, BD Biosciences)
and analyzed using FlowJo software version 7.6.5 (FlowJo LLC).
Three repeats were performed for each sample.
Luciferase reporter assay
The 3′-UTR sequence of TREM2 was inserted into the
firefly luciferase reporter plasmid pGL3-Enhancer-luc2
(pGL3-Enhancer-luc2-pTREM2; E1910, Promega Corporation) by double
digestion using XhoI and HindIII. Subsequently, the
pGL3-Enhancer-luc2-pTREM2 plasmid (1.5 µg) was transfected into
oeHBX and oeNC cells (2×105) using
Lipofectamine® 2000 (cat. no. 11668-027, Invitrogen,
Thermo Fisher Scientific, Inc.). Additionally, the NF-κB inhibitor
PDTC (10 µM; cat. no. S1809; Beyotime Institute of Biotechnology)
was dissolved in DMSO (cat. no. D2650; Sigma-Aldrich; Merck KGaA)
and added to the oeHBX cell culture to knock down the expression of
NF-κB. Following incubation for 48 h at room temperature, the cells
were collected. Subsequently, the firefly and Renilla
luciferase activities were determined using the Dual-Luciferase
Reporter Assay system (cat. no. E1910; Promega Corporation),
according to the manufacturer's protocol. Firefly luciferase
activity was normalized to Renilla luciferase activity.
Three repeats were performed for each sample.
Statistical analysis
Statistical analyses were performed using GraphPad
Prism software (version 7.0; GraphPad Software, Inc.). Data are
presented as the mean ± SD. Data were analysed using one-way ANOVA
followed by Tukey's post hoc test. P<0.05 was considered to
indicate a statistically significant difference.
Results
ECH suppresses the function of HBX in
HK-2 cells
To assess the function of HBX, HBX overexpression
was induced in HK-2 cells. The relative mRNA and protein levels of
HBX were significantly upregulated in oeHBX cells compared with
oeNC cells (Fig. 1A and B).
Subsequently, oeHBX cells were cultured with different
concentrations of ECH (1, 2.5, 5, 10, 20 and 50 mg/l; E1, E2.5, E5,
E10, E20 and E50, respectively). HBX overexpression significantly
reduced the proliferation of HK-2 cells, and this decrease was
reversed by ECH in a dose-dependent manner at 24 and 48 h (Fig. 1C).
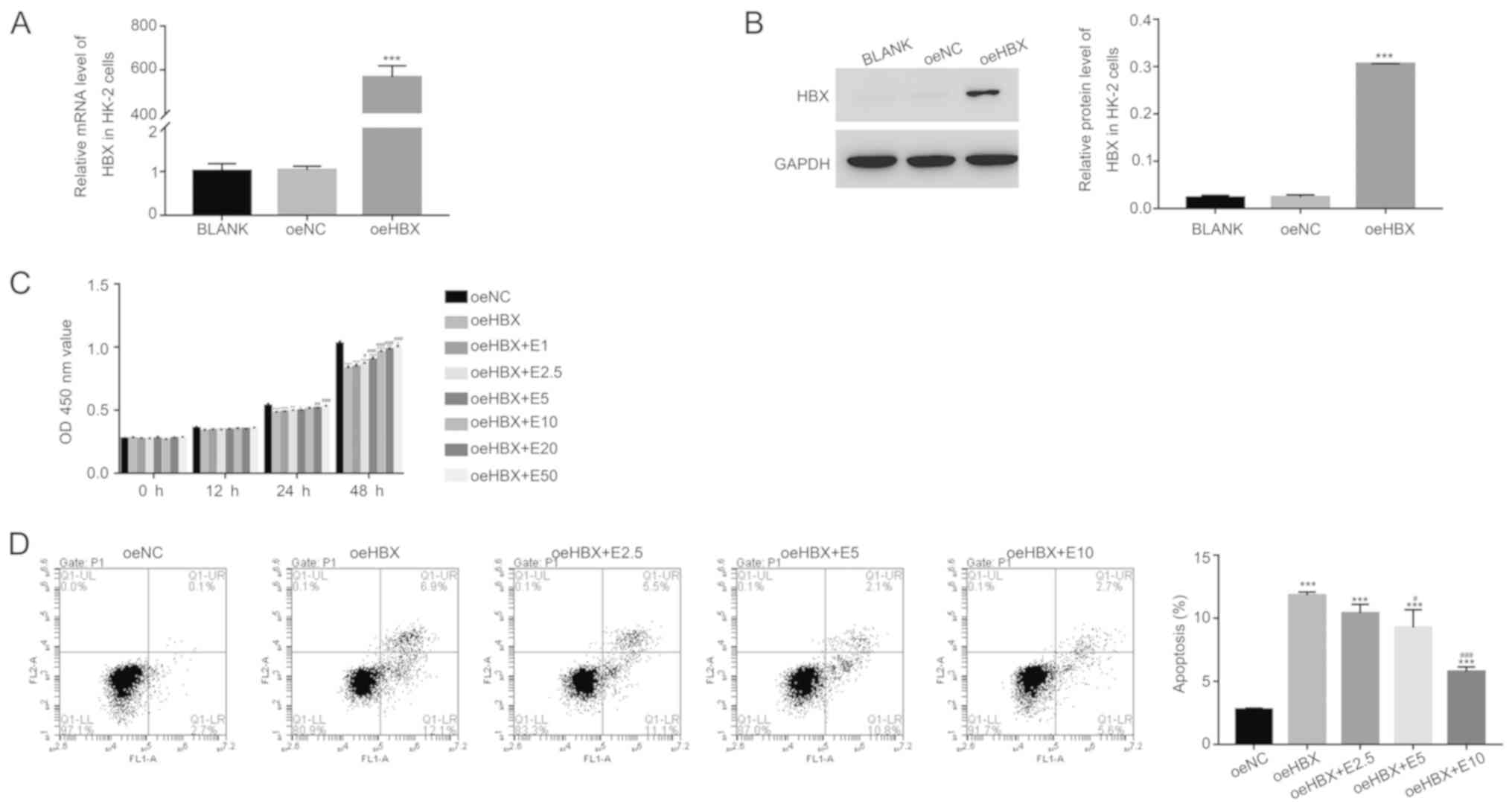 | Figure 1.ECH suppresses the function of HBX in
HK-2 cells. Relative (A) mRNA and (B) protein expression levels of
HBX in HK-2 cells. (C) ECH increased the cell proliferation rate of
oeHBX cells. (D) ECH reduced the apoptotic rate of oeHBX cells.
*P<0.05, **P<0.01 and ***P<0.001 vs. oeNC.
#P<0.05, ##P<0.01 and
###P<0.001 vs. oeHBX. ECH, echinacoside; HBX,
hepatitis B virus X; oe, overexpression; NC, negative control; E1,
1 mg/l ECH; E2.5, 2.5 mg/l ECH; E5, 5 mg/l ECH; E10, 10 mg/l ECH;
E20, 20 mg/l ECH; E50, 50 mg/l ECH; OD, optical density. |
Furthermore, the proliferation rate of oeHBX cells
displayed no significant difference compared with E1-treated cells
(Fig. 1C). By contrast, E10
treatment significantly increased the proliferation rate of oeHBX
cells compared with E20 and E50 treatment which exhibited no
significant difference (Fig. 1C).
Therefore, the effect of ECH on apoptosis was examined in E2.5, E5
and E10-treated cells. HBX overexpression significantly increased
the apoptosis of HK-2 cells, and the apoptosis rate of oeHBX cells
decreased with ECH treatment in a dose-dependent manner (Fig. 1D). Overall, the results suggested
that ECH inhibited the function of HBX in HK-2 cells in a
dose-dependent manner.
TREM2 expression is inhibited by ECH
in transfected oeHBX cells
RT-qPCR and western blotting were used to examine
the relative mRNA and protein expression levels of TREM2 in oeHBX
cells, respectively. TREM2 mRNA and protein expression levels were
significantly upregulated in oeHBX cells compared with oeNC cells
(Fig. 2A and B). However, ECH
decreased the expression levels of TREM2 in transfected oeHBX cells
in a dose-dependent manner (Fig. 2A
and B). Therefore, the results suggested that HBX expression
was positively associated with TREM2 expression, and further
indicated the suppressive effect of ECH on HBX in HK-2 cells.
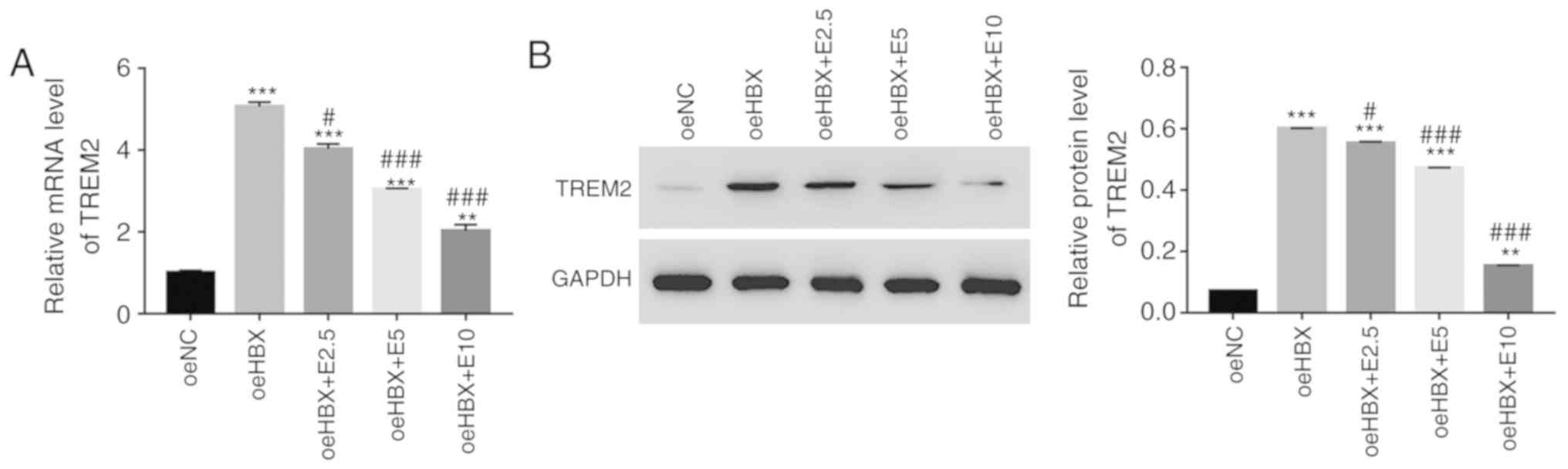 | Figure 2.ECH suppresses the expression of
TREM2 in oeHBX cells. (A) Relative mRNA expression levels of TREM2
were reduced by ECH in oeHBX cells. (B) Relative protein expression
levels of TREM2 were reduced by ECH in oeHBX cells. **P<0.01 and
***P<0.001 vs. oeNC. #P<0.05 and
###P<0.001 vs. oeHBX. ECH, echinacoside; TREM2,
triggering receptor expressed on myeloid cells 2; oe,
overexpression; HBX, hepatitis B virus X; NC, negative control; E1,
1 mg/l ECH; E2.5, 2.5 mg/l ECH; E5, 5 mg/l ECH; E10, 10 mg/l ECH;
E20, 20 mg/l ECH; E50, 50 mg/l ECH. |
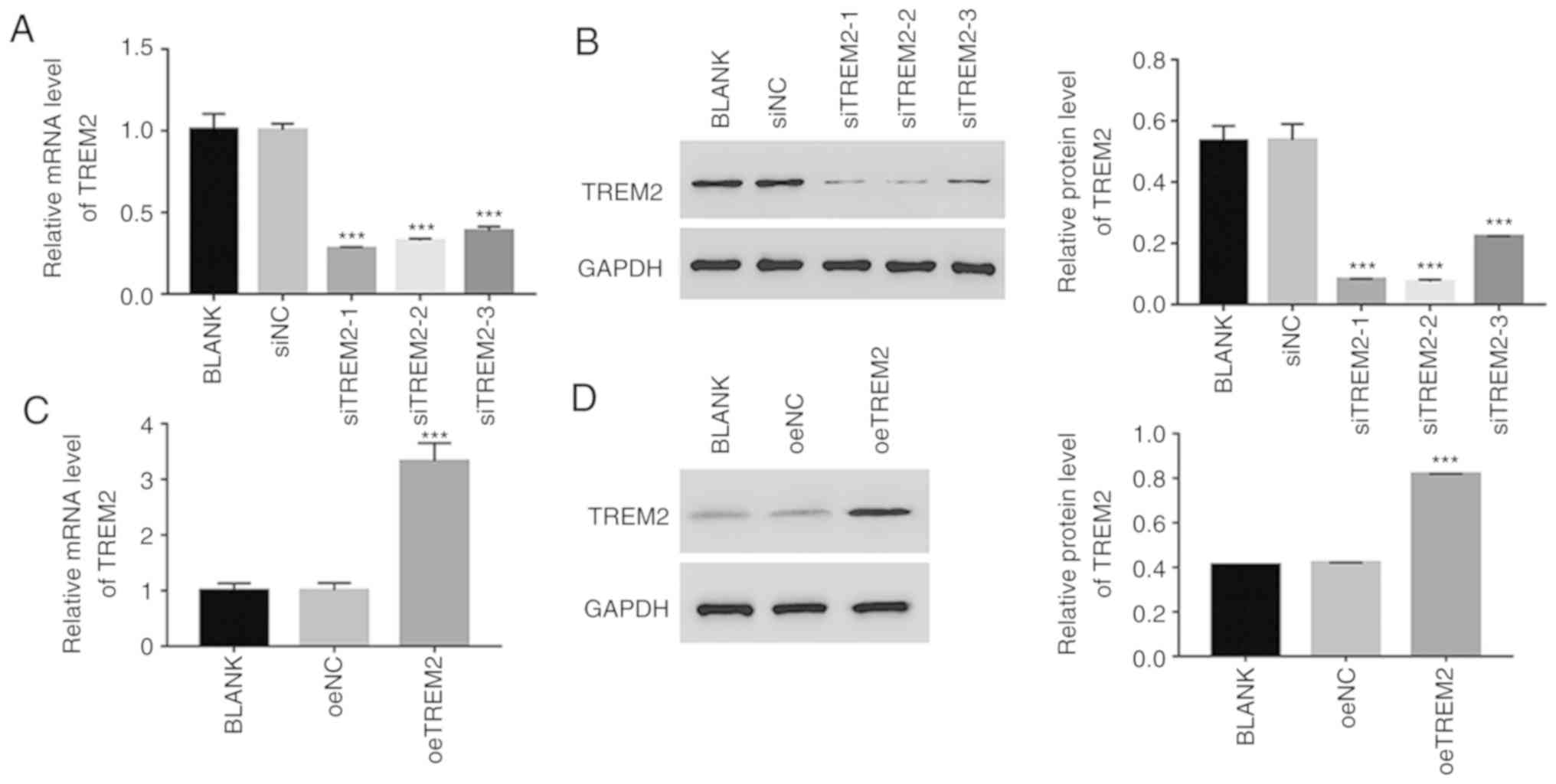
Knockdown and overexpression of TREM2
in HK-2 cells
To further assess the function of TREM2, TREM2
knockdown and overexpression were induced in HK-2 cells using RNA
interference and a lentiviral vector, respectively. For the
knockdown assay, three siRNAs targeting the human gene TREM2
(siTREM2-1, siTREM2-2 and siTREM2-3) were transfected into HK-2
cells, and a non-specific siRNA was used as a negative control
(siNC). All three TREM2 siRNAs successfully knocked down the
endogenous expression of TREM2 in HK-2 cells (Fig. 3A and B). However, the lowest
relative TREM2 mRNA and protein expression levels were observed in
siTREM2-1 and siTREM2-2-transfected cells (Fig. 3A and B), therefore, siTREM2-1 and
siTREM2-2-transfected cells were chosen for subsequent
experimentation.
For the overexpression experiments, a plasmid
containing the full-length human TREM2 cDNA sequence was
constructed and subsequently transfected into HK-2 cells (oeTREM2).
A mock plasmid served as a negative control (oeNC). The level of
TREM2 expression was significantly increased in oeTREM2 cells
compared with oeNC cells (Fig. 3C and
D). Therefore, oeTREM2 cells were used for subsequent
experimentation.
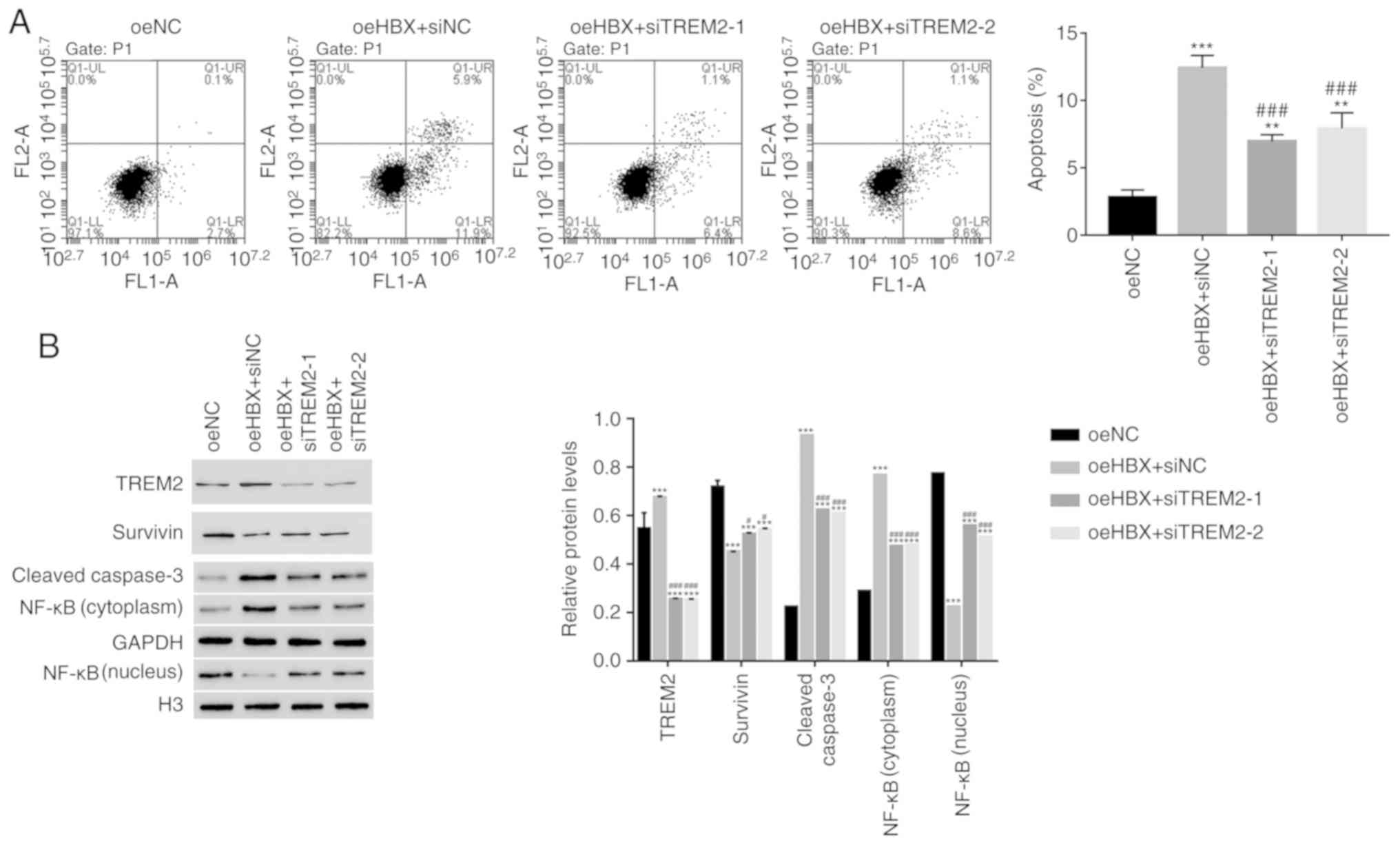
TREM2 silencing decreases the
apoptosis of oeHBX cells
The apoptosis profile of oeHBX cells transfected
with siTREM2 or siNC was assessed. The apoptosis rate of oeHBX +
siNC cells was significantly higher compared with oeNC cells.
However, the apoptosis rate was significantly decreased in oeHBX +
siTREM2-1 or siTREM2-2 cells compared with oeHBX + siNC cells
(Fig. 4A). The results suggested
that TREM2 knockdown inhibited the function of HBX during HK-2 cell
apoptosis.
Survivin belongs to the inhibitor of apoptosis
protein family, which inhibits apoptotic activity and suppresses
cell death (25). Targeting
Survivin has been identified as a novel approach for the treatment
of renal cell carcinoma (26).
NF-κB is also an apoptosis inhibitor that plays a role in
antiapoptotic tumor processes (27). Furthermore, caspase3 activation is
closely associated with apoptosis (28). In the present study, the protein
content of TREM2, Survivin and cleaved caspase3 in HK-2 cells was
quantified. The level of TREM2 expression was significantly
decreased in oeHBX + siTREM2-1/2 cells compared with oeHBX + siNC
cells (Fig. 4B). The level of
Survivin expression was significantly increased in oeHBX +
siTREM2-1/2 cells compared with oeHBX cells. Additionally, the
level of cleaved caspase3 expression was significantly decreased in
oeHBX + siTREM2-1/2 cells compared with oeHBX + siNC cells
(Fig. 4B). Furthermore, oeHBX
significantly decreased the translocation of NF-κB to the nucleus,
and siTREM2-1/2 transfection significantly increased NF-κB nuclear
translocation (Fig. 4B). Taken
together, the results suggested that TREM2 was a downstream factor
of HBX in HK-2 cells, therefore, HBX might promote apoptosis by
regulating TREM2 expression in human RTECs.
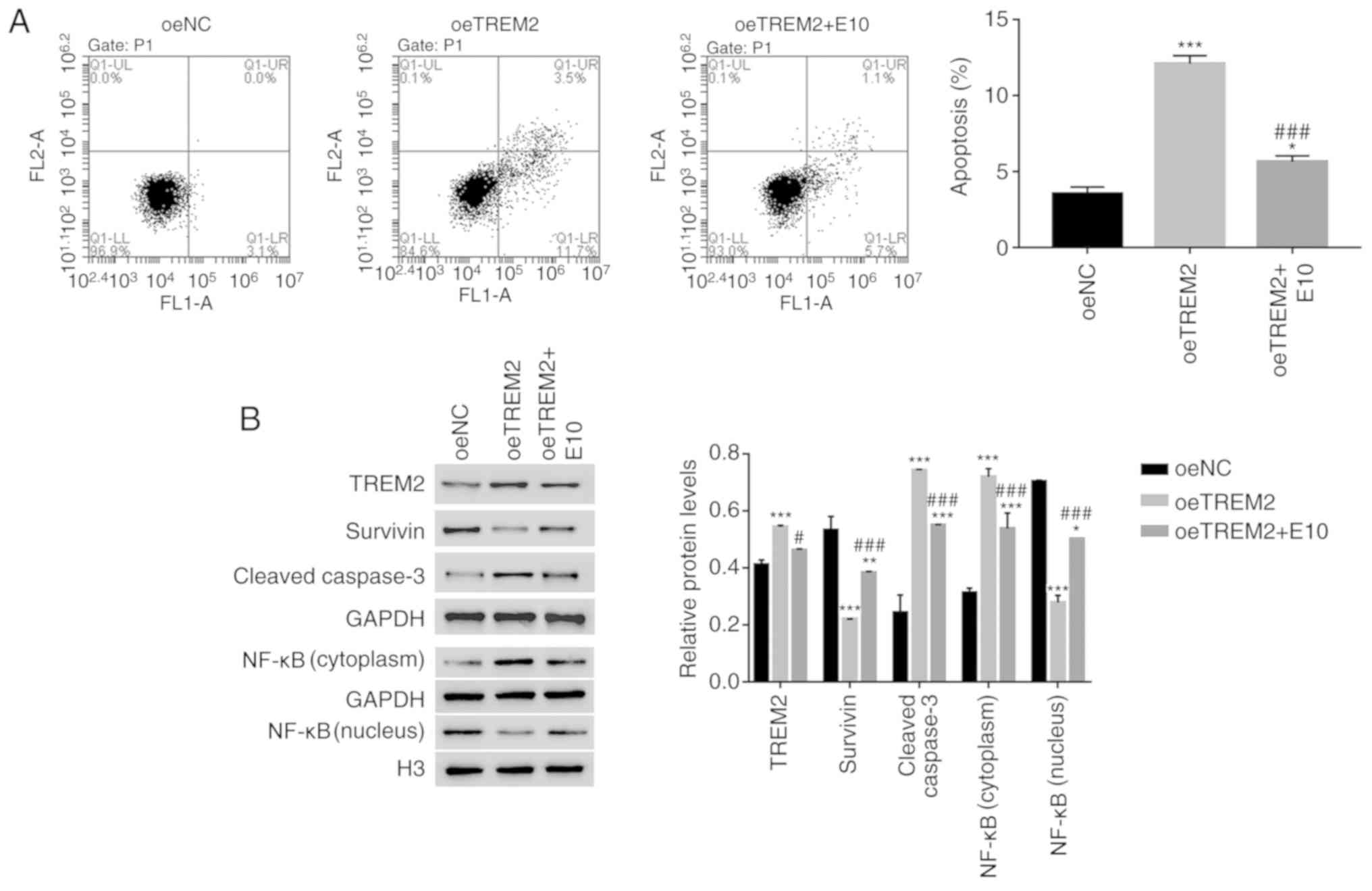
TREM2 overexpression suppresses the
translocation of NF-κB to the nucleus in HK-2 cells
The effect of ECH on oeTREM2-transfected cells was
analysed. TREM2 overexpression promoted the apoptosis of HK-2 cells
(Fig. 5A). The apoptosis of
oeTREM2 cells was significantly decreased by ECH treatment (E10;
Fig. 5A). Furthermore, ECH
significantly decreased the expression of TREM2 in oeTREM2 cells.
TREM2 overexpression significantly decreased the expression levels
of Survivin, however, this decrease in expression was reversed by
ECH. Additionally, the protein expression levels of cleaved
caspase3 were significantly increased in oeTREM2 cells compared
with oeNC cells; an effect which was significantly decreased by ECH
treatment. Furthermore, ECH promoted the translocation of NF-κB to
the nucleus in oeTREM2 cells (Fig.
5B). Taken together, the results indicated that ECH also
suppressed the function of TREM2 in human RTEC cells.
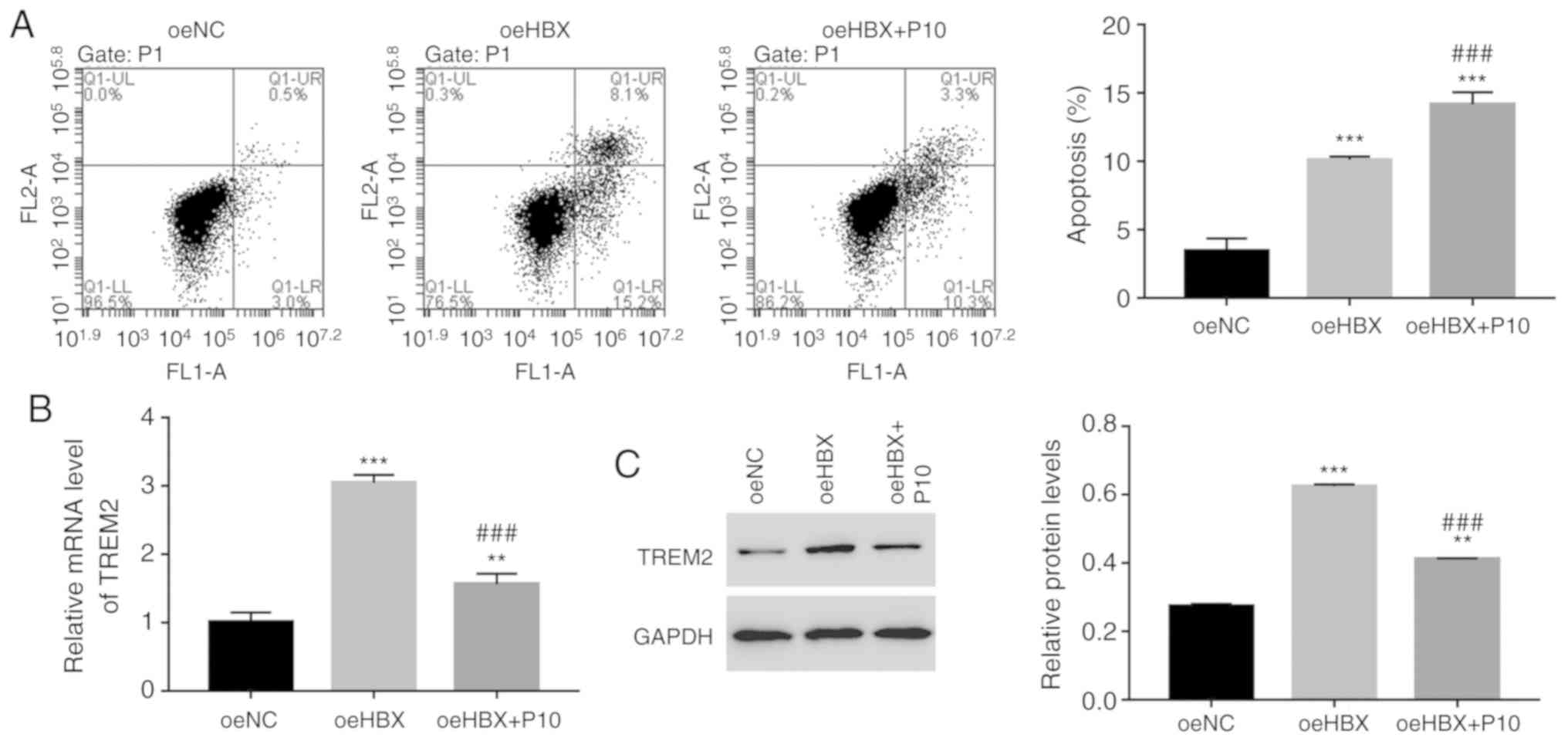
HBX functions are suppressed by the
NF-κB inhibitor PDTC in human RTECs
To further assess the relationship between HBX and
NF-κB, HK-2 cells were cultured with a specific NF-κB inhibitor,
PDTC (10 µmol/l; P10). The cell apoptosis rate of oeHBX cells was
significantly increased compared with oeNC cells (Fig. 6A). However, oeHBX + PDTC cells
displayed a significantly increased apoptosis rate compared with
oeHBX cells. Furthermore, the relative mRNA and protein expression
levels of TREM2 were decreased in oeHBX + PDTC cells compared with
oeHBX cells (Fig. 6B). Therefore,
the results suggested that NF-κB negatively regulated TREM2
expression in oeHBX-transfected cells.
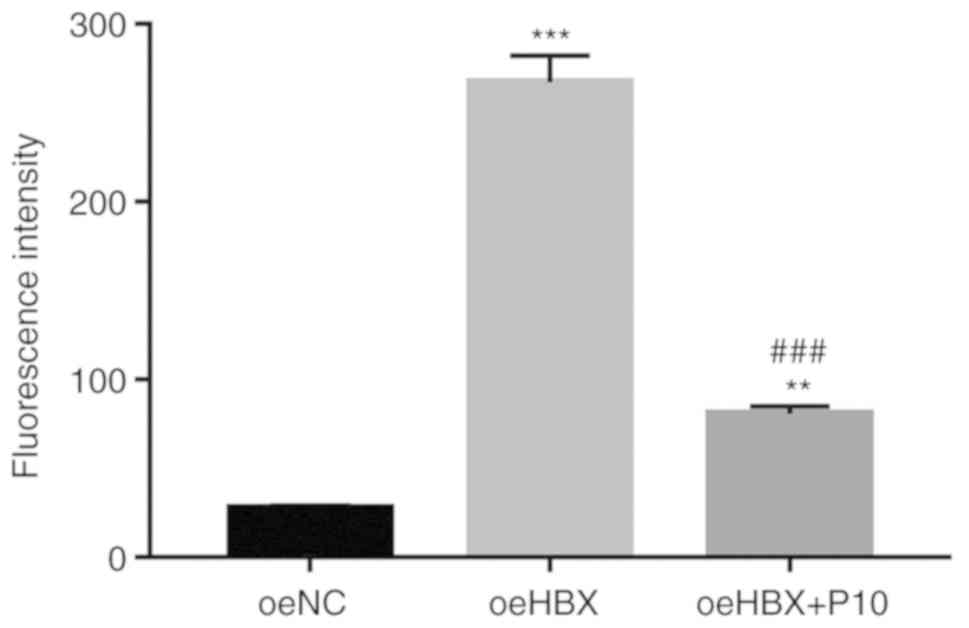
NF-κB inhibitor PDTC suppresses TREM2 promoter
activity in oeHBX cells. A luciferase reporter (pGL3-Enhancer-luc2)
containing the wild-type promoter sequence of TREM2
(pGL3-Enhancer-luc2-pTREM2) was constructed and subsequently
transfected into oeNC, oeHBX and oeHBX + P10 cultured cells.
The luciferase activity of the reporter vector
containing the wild-type TREM2 promoter was significantly increased
in oeHBX cells compared with oeNC cells. However, PDTC
significantly suppressed the luciferase activity of the reporter in
oeHBX cells (Fig. 7). Overall, the
results indicated that PDTC inhibited the transcription of TREM2 by
suppressing the promoter activity of TREM2 in oeHBX cells.
Discussion
HBX is primarily expressed in RTECs of patients with
HBV-GN, and functions as a determinant of viral pathogenesis
(11,29). Therefore, increasing knowledge of
the function of the HBX molecule network is a critical step in
developing a novel approach for the treatment of HBV-GN. The aim of
the present study was to further examine the function of HBX and to
explore its potential molecular network in human RTECs.
HBX inhibits the proliferation of RTECs (11). In the present study,
antiproliferation and pro-apoptosis functions of HBX in human RTECs
were identified, suggesting that HBX suppressed the development of
human RTECs.
A previous study indicated that ECH inhibits HBV
replication and antigen expression (19). In the present study, ECH treatment
disrupted the function of HBX in HK-2 cells, therefore, HBX may be
affected by ECH in HK-2 cells. Furthermore, the results suggested
that ECH may serve as a potential therapeutic agent for HBV-GN.
TREM2 is an innate immune receptor that plays a role
in the inflammatory response (30). In the present study, HBX expression
was positively associated with TREM2 expression in human RTECs.
Moreover, TREM2 knockdown significantly decreased the apoptosis
rate of oeHBX cells, and ECH treatment reduced the effect of
oeTREM2 on cell apoptosis. Taken together, these results suggested
that TREM2 was targeted by HBX in HK-2 cells. Therefore, ECH may
inhibit the apoptosis of human RTECs by blocking the activity of
the HBX/TREM2 signalling pathway.
A previous report demonstrated that NF-κB, not only
plays a role in cell apoptosis, but is also involved in the
regulation of immune responses and inflammation (31). Furthermore, TREM2 is mediated by
the NF-κB-sensitive microRNA-34a during Alzheimer's disease and
macular degeneration (32,33). In the present study, the apoptotic
rate of oeHBX cells was significantly increased in the presence of
PDTC. To the best of our knowledge, the present study suggested for
the first time that, the NF-κB inhibitor PDTC suppressed the
promoter activity of TREM2. Furthermore, the results suggested that
TREM2 negatively regulated the translocation of NF-κB to the
nucleus in HK-2 cells, but was positively associated with cleaved
caspase3 expression levels. TREM2 played an opposite role in HK-2
cells to its role in human degenerative nucleus pulposus and glioma
cells (22,23). Therefore, the present study
suggested that TREM2 might have different functions in different
types of human cells.
Taken together, the present study, not only
indicated that NF-κB was a novel component of the HBX/TREM2
signalling pathway, but also suggested that NF-κB negatively
regulates TREM2 expression in human RTECs. However, the major
limitations of the present study were the lack of in vivo
experiments and clinical data. Therefore, further in vivo
and clinical studies are required to confirm the findings of the
present study.
In the present study, the function of ECH was
investigated and the results suggested the potential effects of ECH
on the signalling pathways in human RTECs. Furthermore, the present
study enhanced the existing knowledge of the biological function of
ECH in human RTEC cells and also suggested its potential as a novel
therapeutic agent for HBV-GN.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
This research was supported by the Backbone Training
Project of Traditional Chinese Medicine in Yangpu District,
Shanghai (grant no. YP18ZY03).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
DG designed this project and wrote the manuscript;
YZ and QW performed the experiments; LZ analyzed the data and
edited diagrams. LW provided technical assistance. All authors
reviewed and approved the final manuscript.
Ethics approval and consent to
participate
This research was approved by the ethics committee
of Yangpu District KongJiang Hospital.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Karayiannis P: Hepatitis B virus:
virology, molecular biology, life cycle and intrahepatic spread.
Hepatology international. 11:1–9. 2017. View Article : Google Scholar
|
|
2
|
Seeger C and Mason WS: Hepatitis B virus
biology. Microbiol Mol Biol Rev. 64:51–68. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Usama E, Ana Maria S, W Ray K and Fervenza
FC: Treatment of hepatitis B virus-associated nephropathy. Nephron
Clin Pract. 119:c41–c49. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zhang Y, Li J, Peng W, Yu G, Wang L, Chen
J and Zheng F: HBV-Associated Postinfectious Acute
Glomerulonephritis: A Report of 10 Cases. PloS one.
11:e01606262016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Slagle BL and Bouchard MJ: Hepatitis B
Virus X and Regulation of Viral Gene Expression. Cold Spring Harb
Perspect Med. 6:a0214022016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Guerrieri F, Belloni L, D'Andrea D,
Pediconi N, Le Pera L, Testoni B, Scisciani C, Floriot O, Zoulim F,
Tramontano A, et al: Genome-wide identification of direct HBx
genomic targets. BMC genomics. 18:1842017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Shi T, Hua Q, Ma Z and Lv Q:
Downregulation of miR-200a-3p induced by hepatitis B Virus X (HBx)
Protein promotes cell proliferation and invasion in
HBV-infection-associated hepatocarcinoma. Pathol Res Pract.
213:1464–1469. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Tian Y, Xiao X, Gong X, Peng F, Xu Y,
Jiang Y and Gong G: HBx promotes cell proliferation by disturbing
the cross-talk between miR-181a and PTEN. Sci Rep. 7:400892017.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Idrissi ME, Hachem H, Koering C, Merle P,
Thénoz M, Mortreux F and Wattel E: HBx triggers either cellular
senescence or cell proliferation depending on cellular phenotype. J
Viral Hepat. 23:130–138. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang X, Wang L, Zhu N, Zhou Y, Gu LJ and
Yuan WJ: Hepatitis B virus X protein modulates renal tubular
epithelial cell-induced T-cell and macrophage responses. Immunol
Cell Biol. 94:266–273. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
He P, Zhang D, Li H, Yang X, Li D, Zhai Y,
Ma L and Feng G: Hepatitis B virus X protein modulates apoptosis in
human renal proximal tubular epithelial cells by activating the
JAK2/STAT3 signaling pathway. Int J Mol Med. 31:1017–1029. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yang Y, Wang X, Zhang Y and Yuan W:
Hepatitis B virus X protein and proinflammatory cytokines synergize
to enhance TRAIL-induced apoptosis of renal tubular cells by
upregulation of DR4. Int J Biochem Cell Biol. 97:62–72. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
He P, Zhou G, Qu D, Zhang B, Wang Y and Li
D: HBx inhibits proliferation and induces apoptosis via Fas/FasL
upregulation in rat renal tubular epithelial cells. J Nephrol.
26:10332013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Han W, Luo M, He M, Zhu Y, Zhong Y, Ding
H, Hu G, Liu L, Chen Q and Lu Y: HBx gene transfection affects the
cycle of primary renal tubular epithelial cells through regulating
cyclin expression. Mol Med Rep. 18:1947–1954. 2018.PubMed/NCBI
|
|
15
|
Sun GD, Li CY, Cui WP, Guo QY, Dong CQ,
Zou HB, Liu SJ, Dong WP and Miao LN: Review of Herbal Traditional
Chinese Medicine for the Treatment of Diabetic Nephropathy. J
Diabetes Res. 2016:57498572016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li L, Wan G, Han B and Zhang Z:
Echinacoside alleviated LPS-induced cell apoptosis and inflammation
in rat intestine epithelial cells by inhibiting the mTOR/STAT3
pathway. Biomed Pharmacother. 104:622–628. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Jia Y, Guan Q, Guo Y and Du C:
Echinacoside stimulates cell proliferation and prevents cell
apoptosis in intestinal epithelial MODE-K cells by up-regulation of
transforming growth factor-β1 expression. J Pharmacol Sci.
118:99–108. 2012. View Article : Google Scholar
|
|
18
|
Li F, Yang Y, Zhu P, Chen W, Qi D, Shi X,
Zhang C, Yang Z and Li P: Echinacoside promotes bone regeneration
by increasing OPG/RANKL ratio in MC3T3-E1 cells. Fitoterapia.
83:1443–1450. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Dai LH, Shen YM, Wu YH, Yu XP, Hu HJ, Mi
YJ and Chen JJ: Effect of echinacoside on replication and antigen
expression of hepatitis B virus. Zhongguo Zhong Yao Za Zhi.
40:30472015.(In Chinese). PubMed/NCBI
|
|
20
|
Cantoni C, Bollman B, Licastro D, Xie M,
Mikesell R, Schmidt R, Yuede CM, Galimberti D, Olivecrona G, Klein
RS, et al: TREM2 regulates microglial cell activation in response
to demyelination in vivo. Acta Neuropathol. 129:429–447. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ulland TK, Song WM, Huang SC, Ulrich JD,
Sergushichev A, Beatty WL, Loboda AA, Zhou Y, Cairns NJ, Kambal A,
et al: TREM2 Maintains Microglial Metabolic Fitness in Alzheimer's
Disease. Cell. 170:649–663.e613. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang XQ, Tao BB, Li B, Wang XH, Zhang WC,
Wan L, Hua XM and Li ST: Overexpression of TREM2 enhances glioma
cell proliferation and invasion: a therapeutic target in human
glioma. Oncotarget. 7:2354–2366. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bai M, Yin HP, Zhao J, Li Y and Wu YM:
Roles of TREM2 in degeneration of human nucleus pulposus cells via
NF-κB p65. J Cell Biochem. 119:8784–8796. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta DeltaC(T)) Method. Methods. 25:4022001. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Jaiswal PK, Goel A and Mittal RD:
Survivin: A molecular biomarker in cancer. Indian J Med Res.
141:389–397. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Carew JS, Espitia CM, Zhao W, Mita MM,
Mita AC and Nawrocki ST: Targeting survivin inhibits renal cell
carcinoma progression and enhances the activity of temsirolimus.
Mol Cancer Ther. 14:14042015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ohshima K, Sugihara M, Haraoka S, Suzumiya
J, Kanda M, Kawasaki C, Shimazaki K and Kikuchi M: Possible
immortalization of Hodgkin and Reed-Sternberg cells: telomerase
expression, lengthening of telomere, and inhibition of apoptosis by
NF-kappaB expression. Leuk Lymphoma. 41:367–376. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yang LQ, Fang DC, Wang RQ and Yang SM:
Effect of NF-κB, survivin, Bcl-2 and Caspase3 on apoptosis
ofgastric cancer cells induced by tumor necrosis factor related
apoptosis inducing ligand. World J Gastroenterol. 10:22–25. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Bouchard MJ and Schneider RJ: The
enigmatic X gene of hepatitis B virus. J Virol. 78:12725–12734.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhong L, Chen XF, Wang T, Wang Z, Liao C,
Wang Z, Huang R, Wang D, Li X, Wu L, et al: Soluble TREM2 induces
inflammatory responses and enhances microglial survival. J Exp Med.
214:597–607. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Dolcet X, Llobet D, Pallares J and
Matias-Guiu X: NF-κB in development and progression of human
cancer. Virchows Arch. 446:475–482. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhao Y, Bhattacharjee S, Jones BM, Dua P,
Alexandrov PN, Hill JM and Lukiw WJ: Regulation of TREM2 expression
by an NF-кB-sensitive miRNA-34a. Neuroreport. 24:318–323. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Bhattacharjee S, Zhao Y, Dua P, Rogaev EI
and Lukiw WJ: MicroRNA-34a-mediated down-regulation of the
microglial-enriched triggering receptor and phagocytosis-sensor
TREM2 in age-related macular degeneration. PLoS One.
11:e01502112016. View Article : Google Scholar : PubMed/NCBI
|