Introduction
Diabetes is a chronic metabolic disease and has
become a major global health issue, which is attributed to diet,
smoking, lifestyle and environmental changes (1). According to statistics, there are an
increasing number of cases of diabetes worldwide, and it is
predicted that the global diabetes population will increase to
591.9 million individuals by 2035 (2). Diabetes can lead to the damage of
multiple organs including eyes, heart and kidney, and diabetic
cardiomyopathy (DCM) is a primary complication of the disease
(3). Currently, drug treatment is
still the main therapeutic strategies, however, diet control,
regular exercise, hyperbaric oxygen therapy and stem cell therapy
can also reduce the diabetic complications (1).
Myocardial fibrosis is a major pathological process
of DCM and the characteristic is an imbalance of extracellular
matrix leading to excessive deposition of collagens, which alters
the cardiac structure and impairs the systolic and diastolic
function of the heart, eventually leading to heart failure
(4). A recent study demonstrated
that continuous hyperglycemia leads to high activation of cardiac
fibroblasts (CFs) and induces CF differentiation into
myofibroblasts, resulting in cardiac extracellular matrix unbalance
and myocardial fibrosis in the heart tissue (5–7).
However, the underlying mechanisms of myocardial fibrosis in DCM
remain unclear.
Calcium sensitive receptor (CaSR) is a member of the
C family of the G protein-coupled receptor superfamily, which is
composed of 1,078 amino acids and consists of four major regions
(8–10). CaSR is widely expressed in
prokaryotic and eukaryotic cells, where it regulates the release of
parathyroid hormone and maintains the homeostasis of calcium and
other metal ions (11,12).
The ubiquitin proteasome signaling pathway is a
predominant protein degradation pathway (13), and its activation is regulated by
numerous proteins, such as the Smad ubiquitin regulatory factor 2
(Smurf2) that regulates the transforming growth factor
(TGF)-β1/Smads signaling pathway (14). Autophagy is the primary metabolic
process of eukaryotic organisms (15). During autophagy, substances in the
cytoplasm are phagocytized by autophagosomes, which are spherical
structures with bilayer membranes, and are transported to lysosomes
for degradation. After binding to endosomes or lysosomes,
autophagosomes and their contents are degraded (16) and can therefore provide energy for
the synthesis of macromolecules. Numerous studies have indicated
that autophagy can improve heart diseases. For example,
TGF-β1 treatment causes fibrogenesis and increases
autophagy in human atrial fibroblasts (17). Furthermore, exogenous
H2S treatment promotes the clearance of ubiquitin
aggregates via autophagy in a type 2 diabetes model mice (18).
Previous studies have indicated that CaSR is
associated with a variety of heart and lung diseases (19,20).
The preliminary results indicated that high glucose (HG)-treated
CFs displayed significantly upregulated CaSR expression and an
increased number of autophagosomes. To explore the functional role
and underlying mechanisms of CaSR in HG-induced myocardial
fibrosis, a CaSR agonist and inhibitor were used in the present
study.
Materials and methods
Isolation and culture of neonatal rat
CFs
Primary CFs were isolated from neonatal Wistar rat
(3 days of age). All experiments were approved by the Mudanjiang
Medical University Medical Science Ethics Committee. As previously
described (8,21,22),
neonatal rats were sacrificed via 2% isoflurane inhalation and
subsequent cervical dislocation, the heart was rapidly excised, cut
into pieces (0.5 mm3) in PBS solution and digested using
trypsin for 8 min under sterile conditions at 37.5°C. Dulbecco's
modified Εagles medium (DMEM; Gibco; Thermo Fisher Scientific,
Inc.) was added to terminate the digestion process. The
aforementioned step was repeated 8 times. Cells were obtained by
centrifugation for 10 min at 800 × g at 4°C. Following incubation
for 2 h, the unattached cells were discarded, the attached cells
(CFs) were plated in a petri dish with DMEM containing 10% FBS
(Gibco; Thermo Fisher Scientific, Inc.) and 1%
penicillin-streptomycin (Gibco; Thermo Fisher Scientific, Inc.) and
placed in an incubator at 37°C with 5% CO2. CFs were
passaged every 3 days and CFs at passage 3 were treated with HG (40
mM), a CaSR agonist (R568; 5 µM; cat. no. sc-361302; Santa Cruz
Biotechnology, Inc.), a CaSR inhibitor (Calhex231; 3 µM; cat. no.
sc-207394; Santa Cruz Biotechnology, Inc.) for 24 or 48 h according
to different experiments. CFs were transfected with Smurf2-small
interfering (si)RNA (cat. no. sc-41676; Santa Cruz Biotechnology,
Inc.) for 12 h. CFs cultured with DMEM containing 5.5 mM glucose
composed the control group.
Western blotting
Total proteins were extracted from CFs by RIPA
buffer (Beyotime Institute of Biotechnology) at 4°C and quantified
using the BCA assay kit (Beyotime Institute of Biotechnology).
Proteins (20 µg) were separated by SDS-PAGE (10 or 14%) and
transferred onto PVDF membranes. After blocking with 5% non-fat
dried milk for 2 h at room temperature, membranes were incubated
overnight at 4°C with primary antibodies (1:1,000) against CaSR,
collagen I/III (Col-I/III), p62, Beclin1, microtubule associated
protein 1 light chain 3 α-I/II (LC3-I/II), SKI like proto-oncogene
(SnoN), α-smooth muscle actin (α-SMA), β-actin (cat. nos. sc-47741,
59772/271249, 28359, 48341, 398822, 136958, 130617 and 47778,
respectively; Santa Cruz Biotechnology, Inc.), Smurf2, smad2,
phosphorylated (p)-smad2, smad3, p-smad3, transforming growth
factor (TGF-β1), MMP9 (cat. nos. 12024, 5339, 18338,
9513, 9520, 3709 and 3852, respectively; Cell Signaling Technology,
Inc.), MMP2 and smad7 (cat. nos. 10373-2-AP and 25840-1-AP,
respectively; ProteinTech Group, Inc.). Membranes were then
incubated with the secondary antibodies anti-rat or anti-rabbit
immunoglobulin G (1:10,000; cat. nos. bs-0295M-HRP or bs-0296R-HRP;
BIOSS) for 1 h at room temperature. Bands were detected using ECL
Western Blotting Substrate kit (Beyotime Institute of
Biotechnology). Relative expression levels were normalized to
endogenous control β-actin using Gel-Pro-Analyser Software Version
6.3 (Media Cybernetics, Inc.).
Immunoprecipitation
CFs were seeded into 35-mm culture dishes
(1×106/dish) and cultured at 37°C for 48 h. After
treatments, CFs were harvested and treated with lysed in lysis
buffer plus 1% PMSF (Roche Diagnostics) for 30 min at 4°C. Cells
were centrifuged at 4°C at 3,000 × g for 25 min. Subsequently,
cells were incubated with ubiquitin specific antibody (1:1,000;
cat. no. sc-8017; Santa Cruz Biotechnology, Inc.) overnight at 4°C.
Protein A/G Magnetic Beads (Selleck Chemicals) were used for
binding according to the manufacturer's protocol. Following
incubation for 2 h, the beads were eluted, centrifuged and the
supernatant was collected. Subsequently, western blotting was
performed as aforementioned to detect the level of protein
ubiquitination.
Cell proliferation detection by EdU
and cell cycle analysis
Cell proliferation was detected using an EdU kit
(Guangzhou RiboBio Co., Ltd.). Briefly, CFs were seeded into
96-well plates (5×103/well) and treated with HG (40 mM),
R568 (5 µM) or Calhex231 (3 µM) for 24 h. Subsequently, cells were
stained using the EdU kit according to the manufacturers'
instructions and the nucleus was stained with Hoechst at 37°C for
10 min. Stained cells were observed using an EVOS M50000
fluorescence microscope (magnification, ×200; Thermo Fisher
Scientific, Inc.).
For cell cycle analysis, CFs were seeded into 6-well
plates (1×105 cells per well) and treated with HG (40
mM), R568 (5 µM) or Calhex231 (3 µM) for 48 h. Subsequently, CFs
were washed three times with cold PBS, fixed with 70% ethanol at
4°C overnight and centrifuged at 800 × g at room temperature for 3
min. Cells were then treated with staining buffer, suspended and
incubated with RNaseA (cat. no. C1052; Beyotime Institute of
Biotechnology). CFs were stained with PI (50 µg/ml; Beyotime
Institute of Biotechnology) for 30 min at 37°C. Cell cycle phase
analysis was detected by flow cytometry using a BD C6 flow
cytometer (BD Biosciences) and data analysis of the cytometric
files was performed using BD ModFit LT version 2.0 (Verity Software
House).
Detection of cell viability by the
cell counting kit (CCK)-8 assay
CFs were seeded (2×103 cells/well) into
96-well plates and treated with HG (40 mM), R568 (5 µM) or
Calhex231 (3 µM). Cell viability was detected at 0, 12, 24, 36, 48,
60 or 72 h using a CCK-8 assay kit (cat. no. AR1199; Wuhan Boster
Biological Technology, Ltd.) according to the manufacturer's
protocol. The absorbance was read at 450 nm on a microplate
reader.
Transmission electron microscopy
(TEM)
To observe autophagosomes of CFs, ultrastructural
analysis was performed as previously described (23,24).
Briefly, CFs were fixed with 2.5% glutaraldehyde at 4°C overnight,
and with 1% osmium tetroxide for 2 h at room temperature.
Subsequently, CFs were dehydrated using a graded ethanol series
(50, 70, 90 and 100%), embedded in epoxy resin, stained with 3%
uranyl acetate and 3% lead citrate for 15 min at room temperature
and observed using a H-7650 electron microscopy (magnification,
×10,000). The number of autophagosomes in each field was calculated
using the Adobe Photoshop CS5 (version 12.0.3; Adobe Systems, Inc.)
and ImageJ (version 1.46r; National Institutes of Health) software
(25,26).
Cell autophagy analysis and
transfection with Smurf2-siRNA
CFs were seeded in a confocal dish (3×105
cells/well; Thermo Fisher Scientific, Inc.). Cell autophagy was
determined using a CYTO-ID autophagy detection kit (Enzo Life
Sciences, Inc.) according to the manufacturer's protocol.
LC3II-positive punctate pattern was observed using a BX61
fluorescence microscope (magnification, ×200; Olympus Corporation).
The number of autophagosomes was quantified using ImageJ software
(version 1.48u; National Institutes of Health).
For the transfection, CFs were seeded into a 35-mm
dish (2×105 per dish) and maintained for 24 h in culture
medium without antibiotics before transfection. After being washed
3 times with PBS, transfection was performed using
Lipofectamine® 3000 transfection reagent (Thermo Fisher
Scientific, Inc.) in Opti-MEM medium and control siRNA
(5′-CCCTTAAAGTTCATTCGGG-3′) or Smurf2-siRNA
(5′-GCGTTTGATGCGAGGCATA-3′) at a final concentration of 300 nM
(Santa Cruz Biotechnology, Inc.) for 12 h. After transfection, CFs
were cultured in regular medium for 24 h and then the cells were
subjected to further analysis.
Measurement of intracellular
calcium
CFs were treated with HG (40 mM), HG + R568 (40 mM +
5 µM) or HG + Calhex231 (40 mM + 3 µM). Subsequently, Fluo-3 AM (5
mM; cat. no. ab145254) was added to each group in the dark for 30
min at 37°C. CFs were washed with Ca2+ free Tyrode's
solution and subsequently observed using a BX61 fluorescence
microscope (magnification, ×200; Olympus Corporation).
ELISA
CF culture media was collected to detect Col-I/III
and TGF-β1 levels using ELISA detection kits (cat. nos.
EK0411, EK0424 and EK0513, respectively; Boster Biological
Technology) according to the manufacturer's protocols.
Statistical analysis
All experiments were performed at least 3 times
independently. Data were presented as the means ± standard error of
the mean. Statistical analysis was performed by a two-tailed
Student's t-test or one-way ANOVA followed by the Bonferroni
multiple comparisons test using SPSS 18.0 software (SPSS, Inc.).
P<0.05 was considered to indicate a statistically significant
difference.
Results
Effects of CaSR on Col-I/III and MMPs
expression in CFs and on CF proliferation
Following 48 h culture, the expression of CaSR,
α-SMA, Col-I, Col-III, MMP2 and MMP9 was significantly increased in
the HG group compared with control group. Furthermore, treatment
with R568 and Calhex231 enhanced or attenuated the effects of HG,
respectively (Fig. 1A and B). In
CF supernatant, the concentration of Col-I/III was significantly
higher in the HG and HG + R568 groups, but significantly lower in
the HG + Calhex231 group (Fig.
1C).
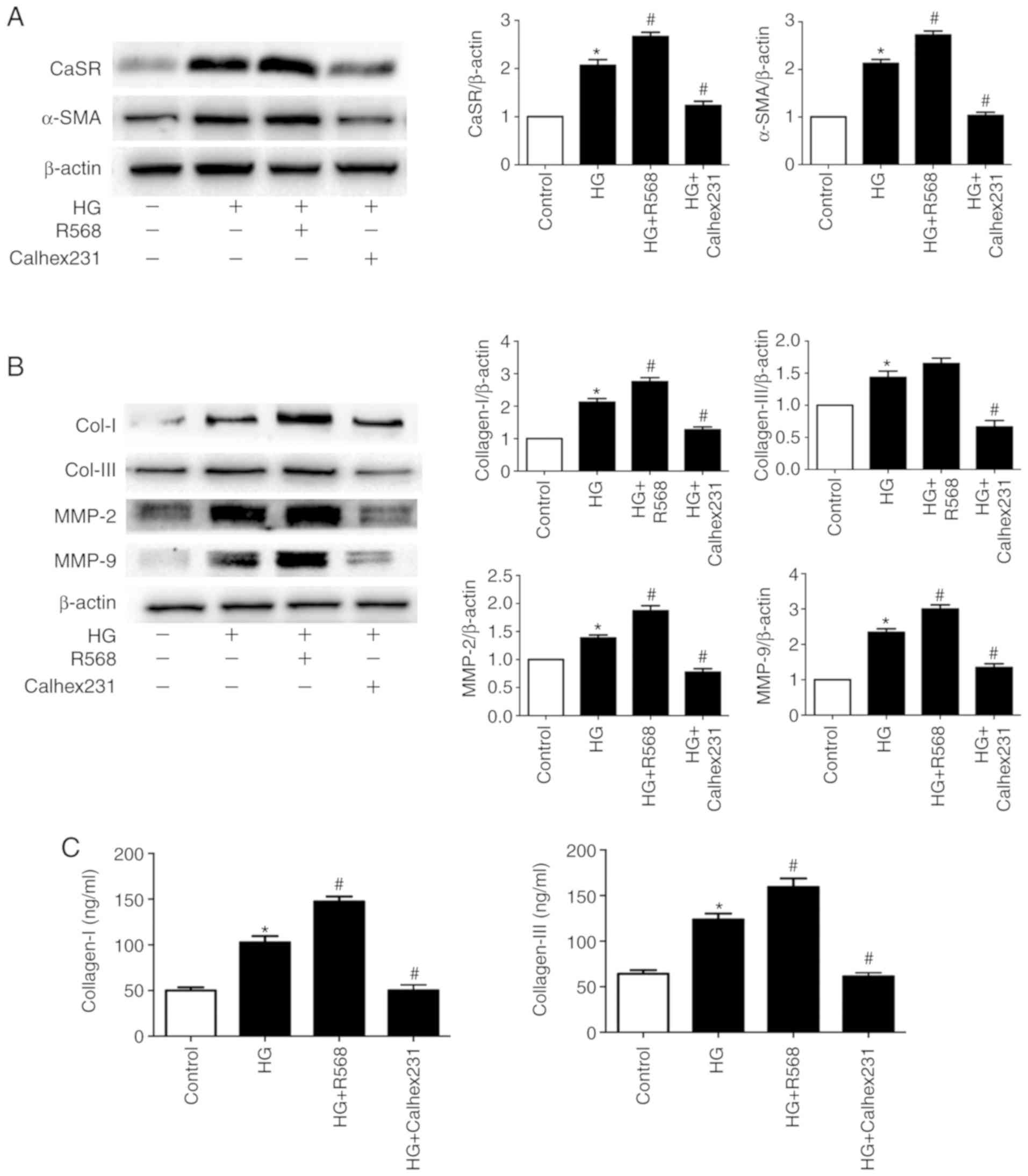 | Figure 1.Effects of CaSR on Col-I/II and MMPs
in CFs treated with HG. CFs in the control group were cultured with
5.5 mM glucose and CFs in the HG group were cultured with 40 mM
glucose with or without 5 µM R568 or 3 µM Calhex231. (A and B)
CaSR, α-SMA, Col-I, Col-III, MMP2 and MMP9 expression was
determined by western blotting. (C) Concentration of Col-I and
Col-III in the CFs supernatant determined by ELISA. *P<0.05 vs.
control; #P<0.05 vs. HG. n≥3. CaSR, calcium sensing
receptor; CFs, cardiac fibroblasts; HG, high glucose; α-SMA,
α-smooth muscle actin; Col-I, collagen I; Col-III, collagen III;
MMP2, matrix metalloproteinase 2; MMP9, matrix metalloproteinase
9. |
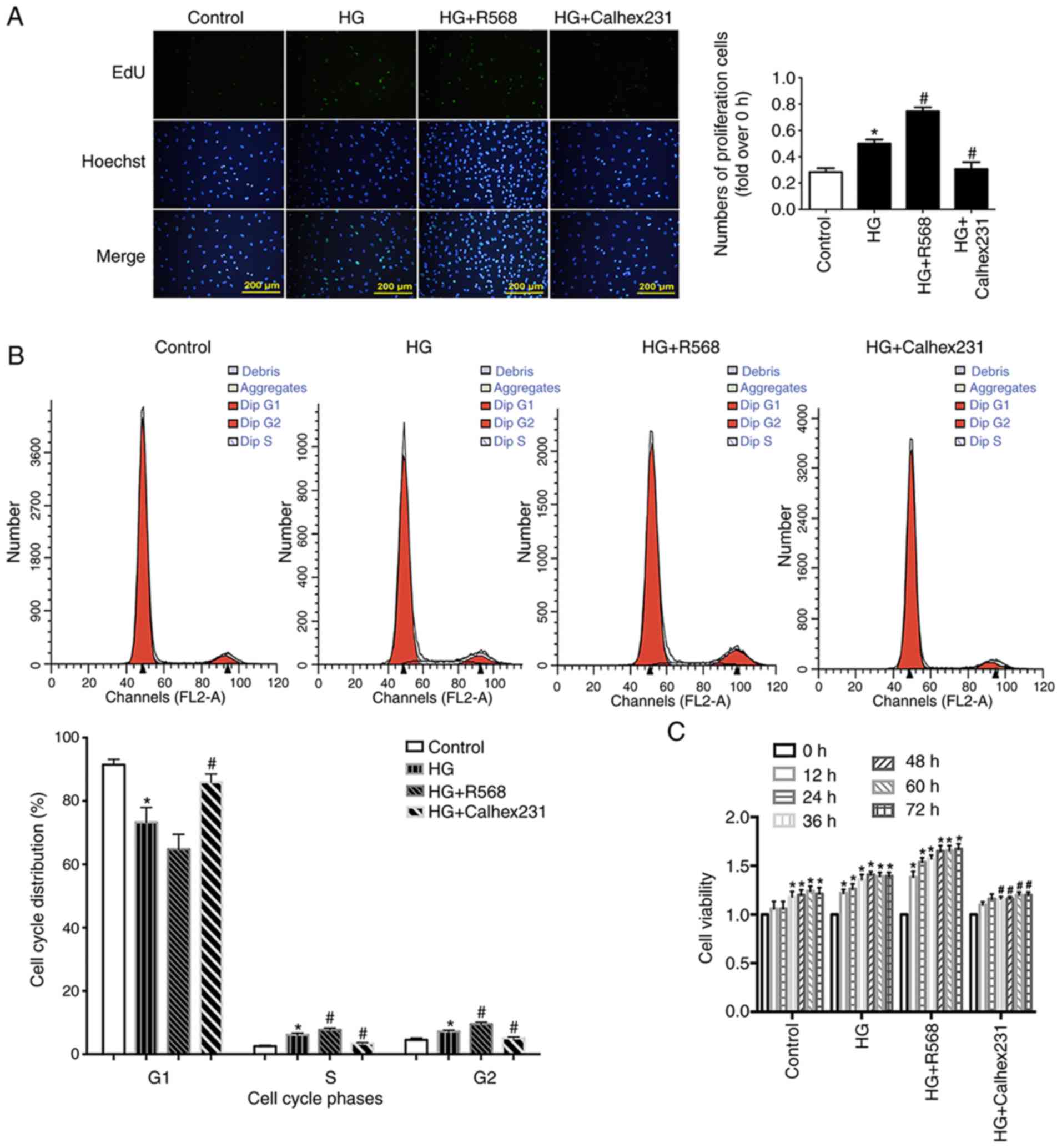
To detect cell proliferation, EdU assay was
performed at the 24 h time point. Cell proliferation was
significantly higher in the HG and HG + R568 groups compared with
the control group. However, cell proliferation was significantly
lower in the HG + Calhex231 group compared with the HG group
(Fig. 2A).
Cell proliferation is closely related to the cell
cycle, including the transition from the G1 phase to the
S and G2 phases (27).
Compared with the control group, the results from cell cycle
analysis demonstrated that the S and G2 phase
distribution of CFs was significantly increased in the HG and HG +
R568 groups, and that the G1 phase distribution was
significantly decreased in the HG group (Fig. 2B). In addition, CF viability at
different time points (0, 12, 24, 36, 48, 60 and 72 h) was detected
with the CCK-8 assay. CF viability in the HG and HG + R568 groups
was significantly increased compared with the control group.
However, compared with the HG group, cell viability was
significantly lower in the HG + Calhex231 group at each time point
(Fig. 2C).
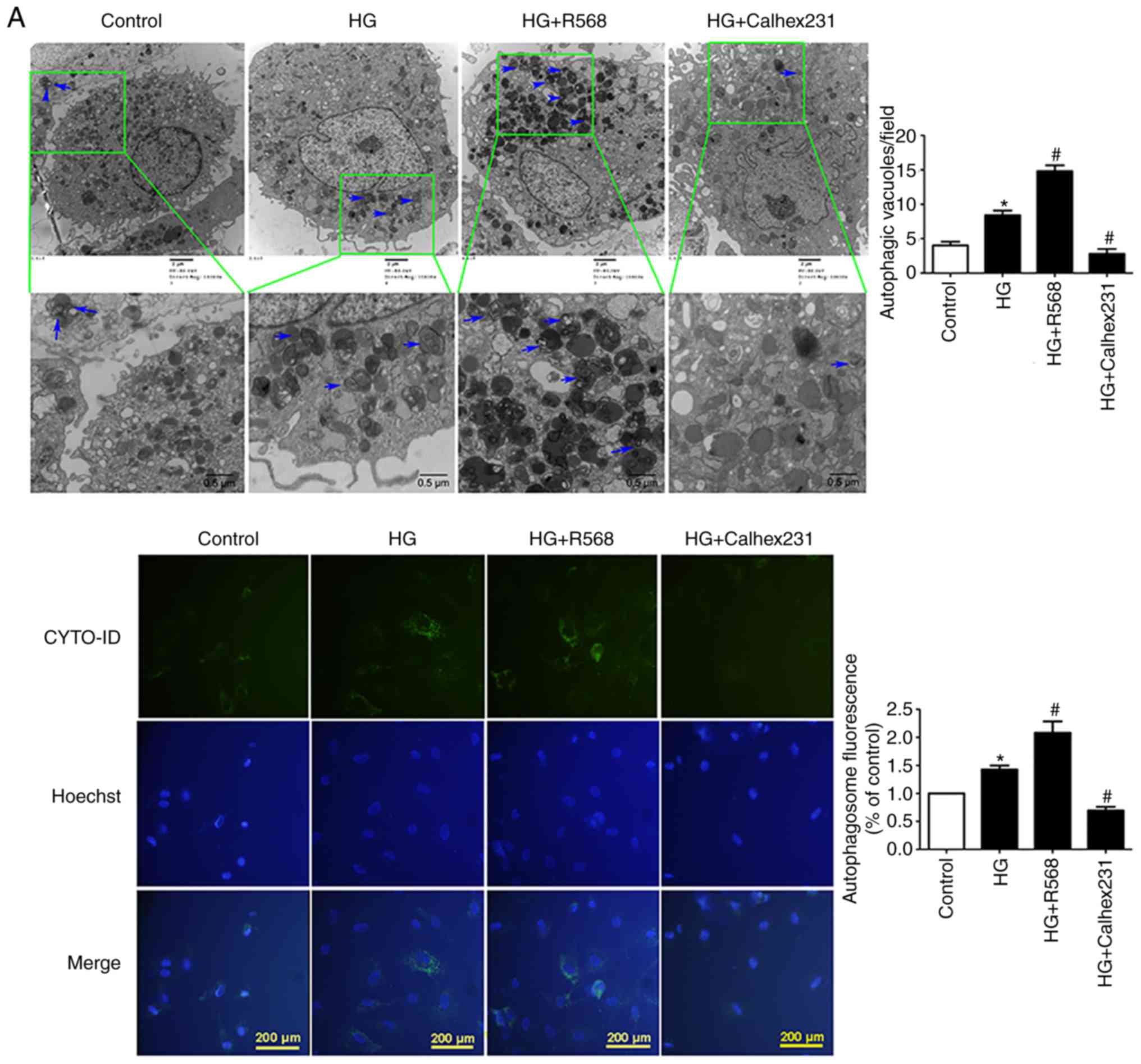
CaSR activation enhances HG-induced
autophagy
CFs were treated with HG, R568 or Calhex231 for 48
h. A significant increase in the number of autophagosomes was
observed in the HG and HG + R568 groups via TEM and the CYTO-ID
autophagy detection kit; however, Calhex231 significantly decreased
the number of autophagosomes (Fig.
3A). The results from western blotting demonstrated that p62
protein expression was significantly downregulated, whereas Beclin1
and LC3-II expression was significantly upregulated in the HG and
HG + R568 groups. Compared with the HG group, opposite results were
observed in the Calhex231 group (Fig.
3B).
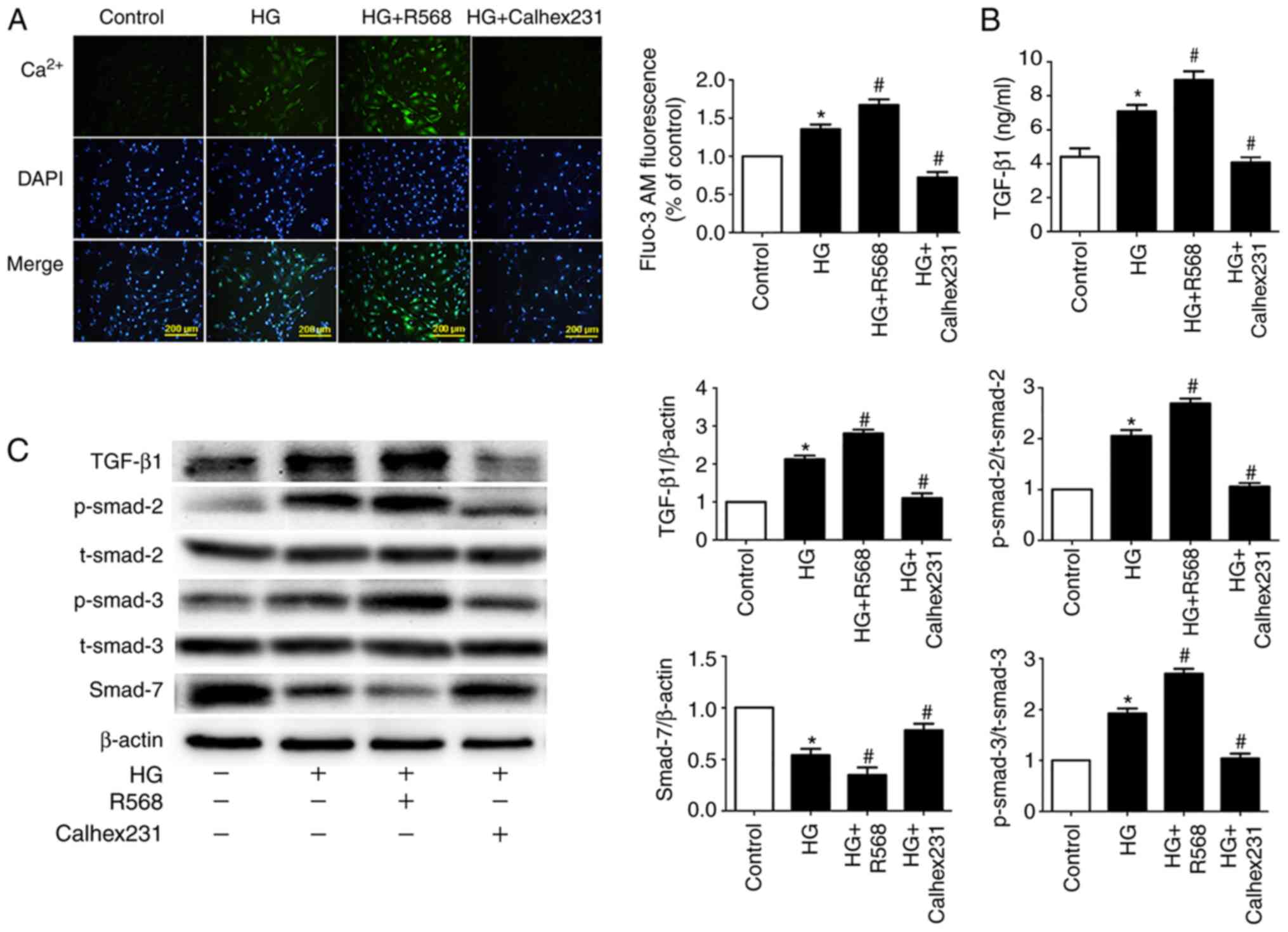
CaSR regulates intracellular
Ca2+ and the TGF-β1/Smads signaling
pathway
To further investigate the underlying mechanism of
myocardial fibrosis, Fluo-3/AM fluorescent dyes were used to detect
the fluorescence intensity of cytosolic Ca2+. Compared
with the control group, the results indicated that fluorescence
intensity was higher in the HG and HG + R568 groups. Compared with
the HG group, fluorescence intensity was lower in the HG +
Calhex231 group (Fig. 4A). To
detect CF TGF-β1 secretion, the ELISA assay was
performed. The concentration of TGF-β1 was significantly
higher in the HG and HG + R568 groups compared with the control
group. Conversely, the concentration of TGF-β1 was
significantly lower in the HG + Calhex231 group compared with the
HG group (Fig. 4B).
Activation of the TGF-β1/Smads signaling
pathway can lead to myocardial fibrosis (28); therefore, western blotting was
performed to detect the expression of primary proteins. The
expression of TGF-β1 and p-Smad2/3 proteins was
significantly increased, whereas Smad7 expression was significantly
decreased in the HG and HG + R568 groups compared with the control
group. As expected, the HG + Calhex231 group displayed the opposite
results compared with the HG group (Fig. 4C).
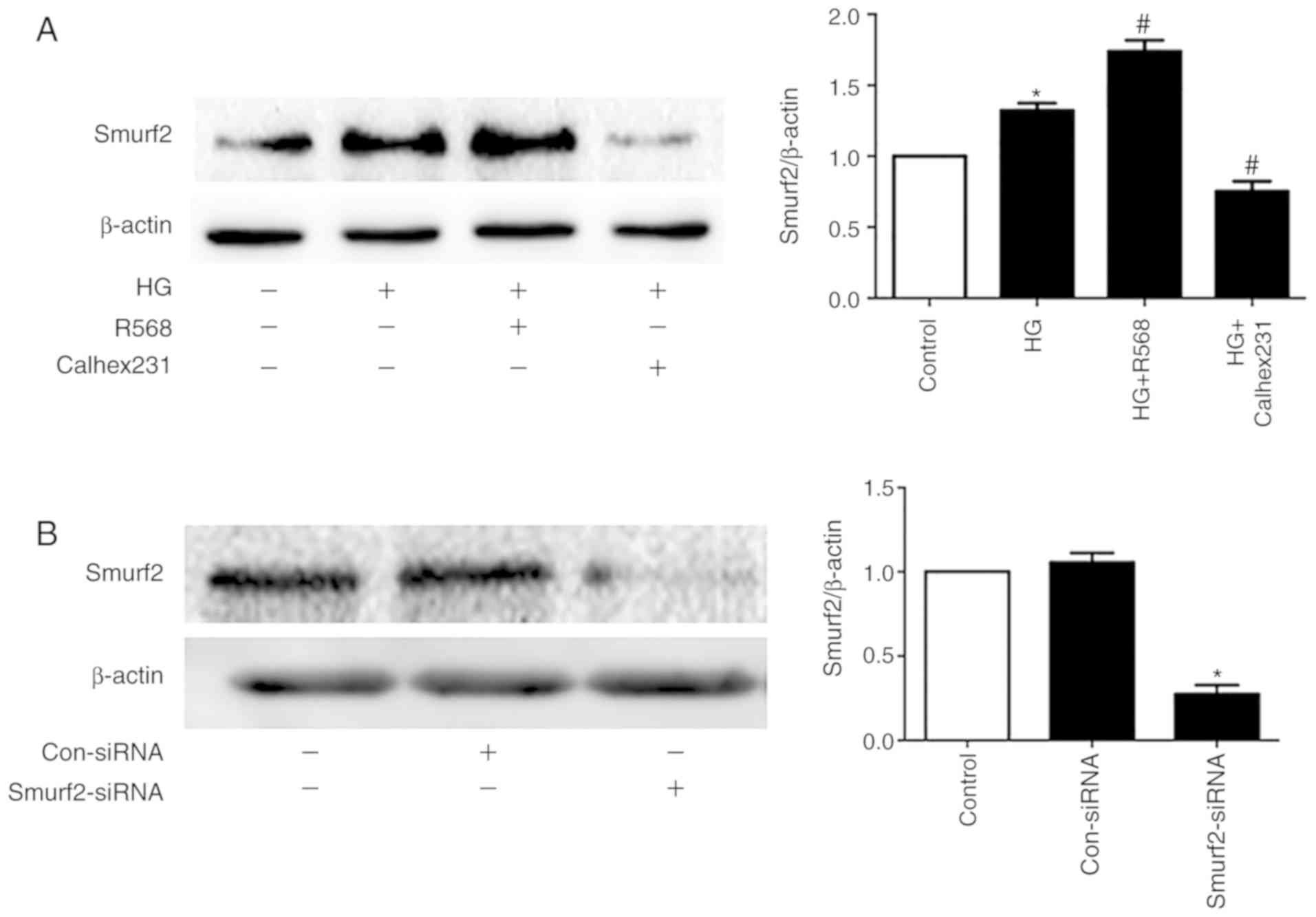
Smurf2-ubiquitin proteasome pathway
and autophagy are involved in HG-induced myocardial fibrosis
The results from western blotting demonstrated that
Smurf2 expression was significantly increased in the HG and R568
groups compared with the control group, but Calhex231 displayed the
opposite effect compared with the HG group, suggesting CaSR
activation increased Smurf2 expression (Fig. 5A). Furthermore, Smurf2 expression
was significantly inhibited following Smurf2-siRNA transfection
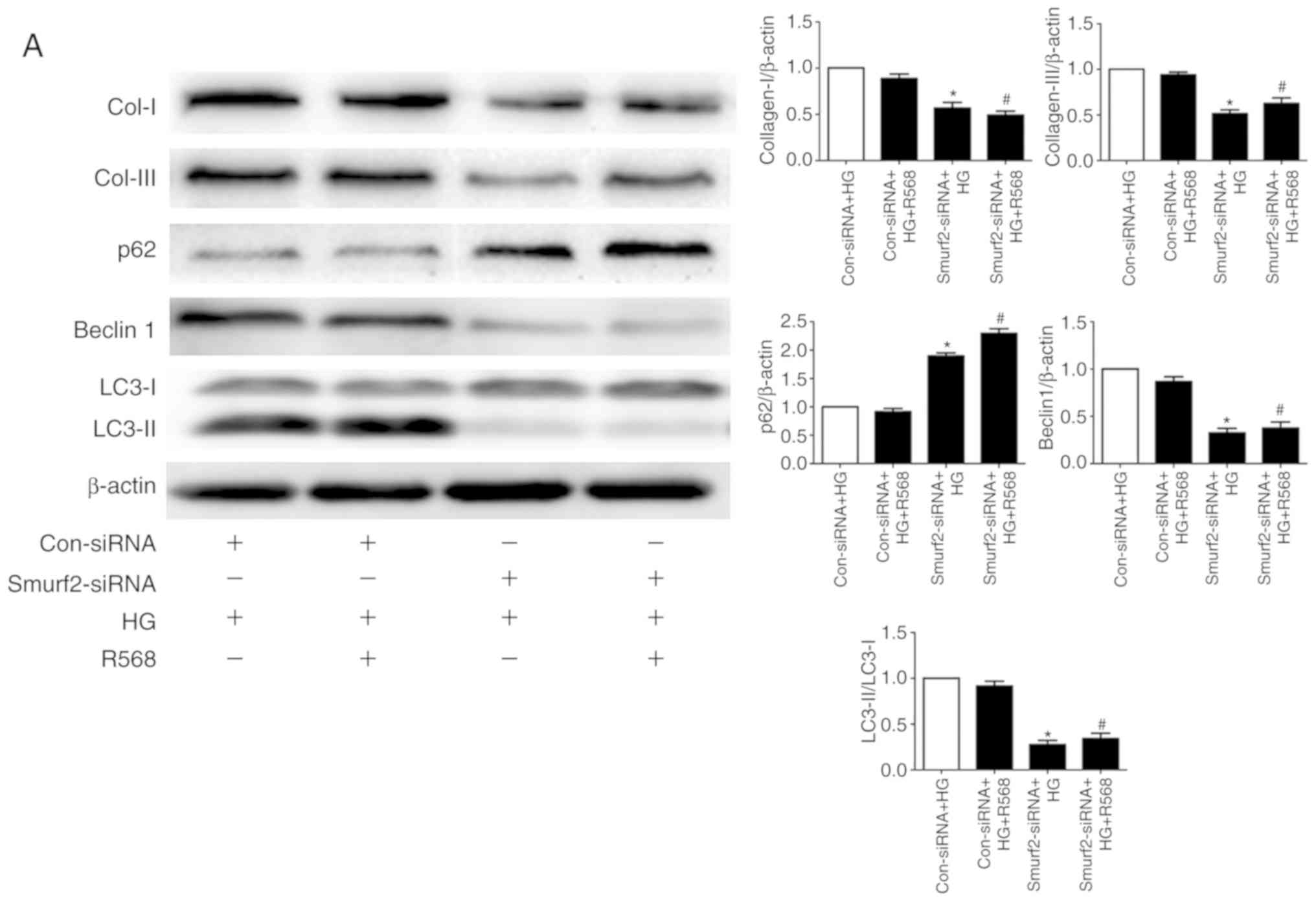
compared with transfection with control siRNA (Fig. 5B). Although CFs were treated with
HG and R658, the detection of ubiquitination levels demonstrated
that the expression levels of SnoN and Smad7 were significantly
suppressed by Smurf2-siRNA. The results also suggested that
Smurf2-siRNA decreased the enhancing effect of HG and R658
(Fig. 5C). Furthermore, compared
with the Con-siRNA + HG and Con-siRNA + HG + R568 groups,
Smurf2-siRNA significantly decreased the expression levels of
Col-I/III, Beclin1 and LC3-II, while p62 expression was
significantly increased in transfected CFs of Smurf2-siRNA + HG and
Smurf2-siRNA + HG + R568 groups (Fig.
6A).
Discussion
Diabetes mellitus is a metabolic syndrome
characterized by hyperglycemia that is caused by insufficient
insulin secretion or resistance, which can eventually lead to DCM
(29). Myocardial fibrosis is a
process of cardiac remodeling and inflammatory gradual
infiltration, which causes numerous conditions, including DCM,
myocardial infarct (30).
According to previous data, CFs serve a key role in the
pathological process of myocardial fibrosis (31) by maintaining homeostasis and
remodelling the extracellular matrix (32). Previous studies have confirmed that
durative hyperglycemia can induce CF proliferation, boost
myofibroblast trans-differentiation and increase the secretion of
extracellular matrix proteins; however, the underlying mechanisms
remain unclear (33–35).
Collagen secretion (Col-I/III) and increased
extracellular matrix accumulation that occurs during myocardial
fibrosis can lead to the proliferation and activation of CFs
(36). A previous study indicated
that CaSR is expressed in CFs (37); however, no study has investigated
the relationship between CaSR expression and myocardial fibrosis in
DCM. R568 is a positive allosteric modulator (calcimimetics) and
can activate CaSR expression to increase intracellular
Ca2+concentration. Calhex231 is an inhibitor of the CaSR
via negative allosteric modulation and can reduce intracellular
calcium (38,39). In the present study, when CFs were
treated with HG, the protein expression of CaSR, α-SMA, Col-I,
Col-III, MMP2 and MMP9 was significantly increased. EdU staining,
CCK-8 assays and the cell cycle analysis indicated that HG enhanced
CF proliferation and promoted the secretion of Col-I/III into the
supernatant. In addition, the CaSR agonist (R568) and the CaSR
inhibitor (Calhex231) promoted and inhibited HG-induced
alterations, respectively. The results indicated that CaSR was
closely associated with HG-induced myocardial fibrosis in DCM.
Autophagy is a highly conservative catabolic process
that is associated with several cardiac pathologies (40). Autophagy can also stabilize and
maintain the metabolic process of intracellular environmental
balance. In injured cells, damaged substances and organelles are
phagocytosed by autophagosomes of spherical double layer membranes,
and then transported to lysosomes via specific mechanisms to
degradation and reuse (41).
Previous studies have confirmed that autophagy participates in the
process of fibrosis (42). The
present study investigated therefore the role of CaSR in CF
autophagy. In CFs treated with HG and R568, a large number of
autophagosomes were observed, and the expression of Beclin1 and
LC3-II was increased, whereas p62 expression was decreased. These
results suggested that CaSR activation induced fibroblast
proliferation and phenotype conversion, which may be associated
with the autophagy pathway.
The TGF-β1/Smads signaling pathway is the
primary route of tissue fibrosis, which occurs during autophagy.
p62, LC3-II and Beclin1 are the key proteins associated with
autophagy (28). Smad7 can degrade
Smad2/3, preventing nuclear translocation and restraining
activation of the TGF-β1/Smads signaling pathway
(43). SnoN is a member of the SKI
proto-oncogene family, which can inhibit Smad2/3 and Smad4 to form
a combined complex, resulting in fibrosis inhibition (44). The present study further indicated
that HG and CaSR agonists remarkably enhanced the expression of
TGF-β1 and p-Smad2/3, and degraded Smad7. However, CaSR
antagonists had the opposite effect.
As a primary secondary messenger, intracellular
Ca2+ concentrations regulate numerous physiological
functions such as electrophysiology of cardiac myocytes, the
contraction of smooth muscle cells and immune cell response
(45–47). Previous studies have indicated that
CaSR could increase intracellular calcium via the G
protein-phospholipase C-inositol triphosphate signaling pathway
(20,48). However, why the activation of CaSR
can enhance the effects of HG on myocardial fibrosis has not been
reported. It was speculated that increased intracellular calcium
could induce Smurf2 expression, which could degrade SnoN and Smad7
proteins via the ubiquitin proteasome signaling pathway (49,50).
This hypothesis was investigated in the present study by detecting
the ubiquitination level of SnoN and Smad7. Smurf2-siRNA
significantly reduced the ubiquitination level of SNoN and Smad7,
which prevented the stimulating effects of HG and R568. Col-I/III
are essential components of the extracellular matrix; therefore,
Col-I/III content increases can account for myocardial fibrosis.
The present study detected the relationship between autophagy and
alterations to Col-I/III expression in CFs. Smurf2-siRNA
significantly decreased the expression of Col-I/III, Beclin1 and
LC3-II in CFs. However, the expression of p62 was significantly
increased. Taken together, these results indicated that
downregulation of autophagy could inhibit collagen secretion by
CFs.
In summary, it has been hypothesized that the
continuous stimulation of hyperglycemia could upregulate CaSR
expression in CFs to increase intracellular Ca2+
(51) and activate
Smurf2-ubiquitin proteasome and autophagy. The present study
demonstrated that proliferative and activated CFs promoted collagen
deposition, which may be a cause of myocardial fibrosis (Fig. 6B), but this requires further
investigation. However, the present study only investigated the
functional role of CaSR in HG-treated CFs. To identify the
underlying mechanisms of CaSR-mediated autophagy in HG-induced
cardiac fibrosis, further investigation is required in vivo,
which may help the discovery of novel strategies for the prevention
and treatment of DCM.
Acknowledgements
Not applicable.
Funding
This research was supported by the Research Projects
of Basic Scientific Research Business Expenses in Higher
Institutions of Heilongjiang Province (grant no. 2018-KYYWF
MY-0070).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
HY and ZW conceived and supervised the study. HY,
JX, YZ and LL designed experiments. QW, YY, BZ and YL performed
experiments. XX and HY analyzed data. HY wrote the manuscript. All
authors reviewed the results and approved the final version of the
manuscript.
Ethics approval and consent to
participate
The animal raising and handling procedures were
performed in accordance with the Guide for the Care and Use of
Laboratory Animals and approved by the Mudanjiang Medical
University Medical Science Ethics Committee (Mudanjiang,
China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
MF
|
myocardial fibrosis
|
|
CaSR
|
calcium sensing receptor
|
|
DCM
|
diabetic cardiomyopathy
|
|
MMP2/9
|
matrix metalloproteinase 2/9
|
|
α-SMA
|
α-smooth muscle actin
|
|
TGF-β1
|
transforming growth
factor-β1
|
|
CFs
|
cardiac fibroblasts
|
|
SnoN
|
ski-related novel gene
|
|
Smurf2
|
smad ubiquitin regulatory factor 2
|
References
|
1
|
Sun L, Yu M, Zhou T, Zhang S, He G, Wang G
and Gang X: Current advances in the study of diabetic
cardiomyopathy: From clinicopathological features to molecular
therapeutics (Review). Mol Med Rep. 20:2051–2062. 2019.PubMed/NCBI
|
|
2
|
Xue H, Tao Y, Deng Y, Yu J, Sun Y and
Jiang G: Metformin accelerates wound healing in type 2 diabetic
db/db mice. Mol Med Rep. 16:8691–8698. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Preshaw PM, Alba AL, Herrera D, Jepsen S,
Konstantinidis A, Makrilakis K and Taylor R: Periodontitis and
diabetes: A two-way relationship. Diabetologia. 55:21–31. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Westermeier F, Riquelme JA, Pavez M,
Garrido V, Díaz A, Verdejo HE, Castro PF, García L and Lavandero S:
New molecular insights of insulin in diabetic cardiomyopathy. Front
Physiol. 7:1252016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Fowlkes V, Clark J, Fix C, Law BA, Morales
MO, Qiao X, Ako-Asare K, Goldsmith JG, Carver W, Murray DB and
Goldsmith EC: Type II diabetes promotes a myofibroblast phenotype
in cardiac fibroblasts. Life Sci. 92:669–676. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Cavalera M, Wang J and Frangogiannis NG:
Obesity, metabolic dysfunction, and cardiac fibrosis:
Pathophysiological pathways, molecular mechanisms, and therapeutic
opportunities. Transl Res. 164:323–335. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hutchinson KR, Lord CK, West TA and
Stewart JA Jr: Cardiac fibroblast-dependent extracellular matrix
accumulation is associated with diastolic stiffness in type 2
diabetes. PLoS One. 8:e720802013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang Y, Gao P, Wei C, Li H, Zhang L, Zhao
Y, Wu B, Tian Y, Zhang W, Wu L, et al: Calcium sensing receptor
protects high glucose-induced energy metabolism disorder via
blocking gp78-ubiquitin proteasome pathway. Cell Death Dis.
8:e27992017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chen Z, Miller CL, Brown EM and Yang JJ:
The calcium sensing receptor: From calcium sensing to signaling.
Sci China Life Sci. 58:14–27. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tennakoon S, Aggarwal A and Kállay E: The
calcium-sensing receptor and the hallmarks of cancer. Biochim
Biophys Acta. 1863:1398–1407. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Tharmalingam S and Hampson DR: The
calcium-sensing receptor and integrins in cellular differentiation
and migration. Front Physiol. 7:1902016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hendy GN and Lucie C: Calcium-sensing
receptor gene: Regulation of expression. Front Physiol. 7:3942016.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Goru SK, Pandey A and Gaikwad AB: E3
ubiquitin ligases as novel targets for inflammatory diseases.
Pharmacol Res. 106:1–9. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Bizet AA, Trankhanh N, Saksena A, Liu K,
Buschmann MD and Philip A: CD109-mediated degradation of TGF-β
receptors and inhibition of TGF-β responses involve regulation of
SMAD7 and Smurf2 localization and function. J Cell Biochem.
113:238–246. 2011. View Article : Google Scholar
|
|
15
|
Yuan H, Guo SX and Zhang JM: Effect of
autophagy in traumatic brain injury. Chin J Pathophysiol.
27:1652–1656. 2011.
|
|
16
|
Levine B and Kroemer G: Autophagy in the
pathogenesis of disease. Cell. 132:27–42. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ghavami S, Cunnington RH, Gupta S, Yeganeh
B, Filomeno KL, Freed DH, Chen S, Klonisch T, Halayko AJ, Ambrose
E, et al: Autophagy is a regulator of TGF-β1-induced fibrogenesis
in primary human atrial myofibroblasts. Cell Death Dis.
6:e16962015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wu J, Tian Z, Sun Y, Lu C, Liu N, Gao Z,
Zhang L, Dong S, Yang F, Zhong X, et al: Exogenous H2S
facilitating ubiquitin aggregates clearance via autophagy
attenuates type 2 diabetes-induced cardiomyopathy. Cell Death Dis.
8:e29922017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Peng X, Li HX, Shao HJ, Li GW, Sun J, Xi
YH, Li HZ, Wang XY, Wang LN, Bai SZ, et al: Involvement of
calcium-sensing receptors in hypoxia-induced vascular remodeling
and pulmonary hypertension by promoting phenotypic modulation of
small pulmonary arteries. Mol Cell Biochem. 396:87–98. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Xu C, Zhang W, Jiang C, Sun Y and Wang R:
Involvement of calcium sensing receptor in myocardial
ischemia/reperfusion injury and apoptosis. J Mol Cell Cardiol.
42:S80–S81. 2007. View Article : Google Scholar
|
|
21
|
Pan YL, Han ZY, He SF, Yang W, Cheng J,
Zhang Y and Chen ZW: miR133b5p contributes to hypoxic
preconditioningmediated cardioprotection by inhibiting the
activation of caspase8 and caspase-3 in cardiomyocytes. Mol Med
Rep. 17:7097–7104. 2018.PubMed/NCBI
|
|
22
|
Liu Z, Hua J, Cai W, Zhan Q, Lai W, Zeng
Q, Ren H and Xu D: N-terminal truncated peroxisome
proliferator-activated receptor-γ coactivator-1α alleviates
phenylephrine-induced mitochondrial dysfunction and decreases lipid
droplet accumulation in neonatal rat cardiomyocytes. Mol Med Rep.
18:2142–2152. 2018.PubMed/NCBI
|
|
23
|
Liu S, Chen S, Li M, Zhang B, Shen P, Liu
P, Zheng D, Chen Y and Jiang J: Autophagy activation attenuates
angiotensin II-induced cardiac fibrosis. Arch Biochem Biophys.
590:37–47. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Tran QG, Yoon HR, Cho K, Lee SJ, Crespo
JL, Ramanan R and Kim HS: Dynamic interactions between
autophagosomes and lipid droplets in chlamydomonas reinhardtii.
Cells. 8:9922019. View Article : Google Scholar
|
|
25
|
Sun Z, Wang Y, Ji S, Wang K and Zhao Y:
Computer-aided analysis with Image J for quantitatively assessing
psoriatic lesion area. Skin Res Technol. 21:437–443. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Mo Y, Lou Y, Zhang A, Zhang J, Zhu C,
Zheng B, Li D, Zhang M, Jin W, Zhang L and Wang J: PICK1 deficiency
induces autophagy dysfunction via lysosomal impairment and
amplifies sepsis-induced acute lung injury. Mediators Inflamm.
2018:67573682018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Estévez-García IO, Cordoba-Gonzalez V,
Lara-Padilla E, Fuentes-Toledo A, Falfán-Valencia R,
Campos-Rodríguez R and Abarca-Rojano E: Glucose and glutamine
metabolism control by APC and SCF during the G1-to-S
phase transition of the cell cycle. J Physiol Biochem. 70:569–581.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wu L, Zhang Q, Mo W, Feng J, Li S, Li J,
Liu T, Xu S, Wang W, Lu X, et al: Quercetin prevents hepatic
fibrosis by inhibiting hepatic stellate cell activation and
reducing autophagy via the TGF-β1/Smads and PI3K/Akt pathways. Sci
Rep. 7:92892017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Groop L and Pociot F: Genetics of
diabetes-are we missing the genes or the disease? Mol Cell
Endocrinol. 382:726–739. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Adeghate E: Molecular and cellular basis
of the aetiology and management of diabetic cardiomyopathy: A short
review. Mol Cell Biochem. 261:187–191. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lam S, Verhagen NAM, Strutz F, van der
Pijl JW, Daha MR and van Kooten C: Glucose-induced fibronectin and
collagen type III expression in renal fibroblasts can occur
independent of TGF-beta1. Kidney Int. 63:878–888. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kehlet SN, Willumsen N, Armbrecht G,
Dietzel R, Brix S, Henriksen K and Karsdal MA: Age-related collagen
turnover of the interstitial matrix and basement membrane:
Implications of age- and sex-dependent remodeling of the
extracellular matrix. PLoS One. 13:e01944582018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Russo I and Frangogiannis NG:
Diabetes-associated cardiac fibrosis: Cellular effectors, molecular
mechanisms and therapeutic opportunities. J Mol Cell Cardiol.
90:84–93. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Loboda A, Sobczak M, Jozkowicz A and Dulak
J: TGF-β1/Smads and miR-21 in renal fibrosis and inflammation.
Mediators Inflamm. 2016:83192832016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yao M, Wang X, Wang X, Zhang T, Chi Y and
Gao F: The Notch pathway mediates the angiotensin II-induced
synthesis of extracellular matrix components in podocytes. Int J
Mol Med. 36:294–300. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Yasuda J, Fukui K, Okada M and Yamawaki H:
T3 peptide, a fragment of tumstatin, stimulates proliferation and
migration of cardiac fibroblasts through activation of Akt
signaling pathway. Naunyn Schmiedebergs Arch Pharmacol.
390:1135–1144. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhang X, Zhang T, Wu J, Yu X, Zheng D,
Yang F, Li T, Wang L, Zhao Y, Dong S, et al: Calcium sensing
receptor promotes cardiac fibroblast proliferation and
extracellular matrix secretion. Cell Physiol Biochem. 33:5572014.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Nakamura A, Hotsubo T, Kobayashi K,
Mochizuki H, Ishizu K and Tajima T: Loss-of-function and
gain-of-function mutations of calcium-sensing receptor: Functional
analysis and the effect of allosteric modulators NPS R-568 and NPS
2143. J Clin Endocrinol Metab. 98:E1692–E1701. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Petrel C, Kessler A, Dauban P, Dodd RH,
Rognan D and Ruat M: Positive and negative allosteric modulators of
the Ca2+-sensing receptor interact within overlapping
but not identical binding sites in the transmembrane domain. J Biol
Chem. 279:18990–18997. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Gottlieb RA and Mentzer RM Jr: Autophagy:
An affair of the heart. Heart Fail Rev. 18:575–584. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Chi J, Wang L, Zhang X, Fu Y, Liu Y, Chen
W, Liu W, Shi Z and Yin X: Activation of calcium-sensing
receptor-mediated autophagy in AngiotensinII-induced cardiac
fibrosis in vitro. Biochem Biophys Res Commun. 497:571–576. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Liu L, Wang C, Sun D, Jiang S, Li H, Zhang
W, Zhao Y, Xi Y, Shi S, Lu F, et al: Calhex231 ameliorates cardiac
hypertrophy by inhibiting cellular autophagy in vivo and in vitro.
Cell Physiol Biochem. 36:1597–1612. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Yao LH: Diverse roles of TGF-β/Smads in
renal fibrosis and inflammation. Int J Biol Sci. 7:1056–1067. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Deheuninck J and Luo K: Ski and SnoN,
potent negative regulators of TGF-beta signaling. Cell Res.
14:65–70. 2004.
|
|
45
|
Smith GL and Eisner DA: Calcium buffering
in the heart in health and disease. Circulation. 139:2358–2371.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Iino M: Spatiotemporal dynamics of
Ca2+ signaling and its physiological roles. Proc Jpn
Acad Ser B Phys Biol Sci. 86:244–256. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Baba Y and Kurosaki T: Physiological
function and molecular basis of STIM1-mediated calcium entry in
immune cells. Immunol Rev. 231:174–188. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Zhang WH, Fu SB, Lu FH, Wu B, Gong DM, Pan
ZW, Lv YJ, Zhao YJ, Li QF, Wang R, et al: Involvement of
calcium-sensing receptor in ischemia/reperfusion-induced apoptosis
in rat cardiomyocytes. Biochem Biophys Res Commun. 347:872–881.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Lönn P, Vanlandewijck M, Raja E, Kowanetz
M, Watanabe Y, Kowanetz K, Vasilaki E, Heldin CH and Moustakas A:
Transcriptional induction of salt-inducible kinase 1 by
transforming growth factor β leads to negative regulation of type I
receptor signaling in cooperation with the Smurf2 ubiquitin ligase.
J Biol Chem. 287:12867–12878. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Cai Y, Shen XZ, Zhou CH and Wang JY:
Abnormal expression of Smurf2 during the process of rat liver
fibrosis. Chin J Dig Dis. 7:237–245. 2010. View Article : Google Scholar
|
|
51
|
Duran J, Troncoso M, Lagos D, Ramos S,
Marin G and Estrada M: GDF11 modulates Ca2+-dependent
Smad2/3 signaling to prevent cardiomyocyte hypertrophy. Int J Mol
Sci. 19:15082018. View Article : Google Scholar
|