Introduction
Ulcerative colitis (UC) is a chronic and recurrent
autoimmune disease, which continues to exert multidimensional
burdens on patients and health care systems (1). Pathogenesis of UC is associated with
intestinal microbiota, immune response dysregulation, gene variants
and environmental risk factors, such as smoking (2). Despite advancements in the treatment
of this chronic condition, these clinical symptoms frequently
reoccur, along with persistent side effects, such as opportunistic
infections (3). Thus, novel
effective therapeutic strategies, such as biological therapy, are
required to improve the prognosis of patients with UC.
Intestinal microbiota play a key role in the
pathogenesis of UC (4,5). A previous study reported that the
levels of Roseburia intestinalis (R.I) significantly
decrease in patients with IBD, and alleviate colitis pathology by
maintaining the Treg/Th17 balance and intestinal epithelial barrier
in murine colitis models (6–8).
Flagellins are important structural components of bacterial
flagellum and are reported to have beneficial effects on the
intestinal epithelial barrier (9,10).
However, the role of R.I flagellin in UC and its underlying
molecular mechanism remain unknown.
Pyroptosis is a novel type of proinflammatory
programmed cell death, which differs from necrosis and apoptosis,
and is closely associated with activation of the nucleotide-binding
oligomerization segment-like receptor family 3 (NLRP3) inflammasome
(11,12). Following stimulation, NLRP3
combines with the adaptor protein, apoptosis-associated speck-like
protein (ASC), which recruits caspase-1 to form the inflammasome
complex (13–15). Subsequently, proximity-induced
self-cleavage converts caspase-1 into cleaved caspase-1, which in
turn cleaves pro-interleukin (IL)-1β, an inactive precursor of
pro-inflammatory cytokine IL-1β, into biologically active IL-1β
(16). Cleaved caspase-1 also
cleaves Gasdermin D to yield the N-terminal fragment (GSDMD-N),
which forms membrane pores to trigger the release of IL-1β
(17,18). Increasing evidence suggests that
activation of the NLRP3 inflammasome and pyroptosis are involved in
the inflammatory process of colitis (19,20).
However, the role of R.I flagellin on the activation of the
NLRP3 inflammasome and pyroptosis in UC remain unknown.
In the present study, the effects of R.I
flagellin on dextran sulfate sodium (DSS)- induced colitis and
inflammasome activation were investigated both in vivo and
in vitro. In addition, the role of R.I flagellin on
inflammasome-triggered pyroptosis was analyzed via western blotting
and Transmission electron microscopy (TEM). Moreover, a microRNA
(miRNA/miR) inhibitor was used to study the role of miR-223-3p on
the signaling mechanisms of R.I flagellin. The present
findings suggested that R.I flagellin may be used as a novel
probiotic product for the treatment of UC.
Materials and methods
Mice
Male C57BL/6J mice (age, 6 weeks; weight, 17–18 g;
n=18) were purchased from the Animal Center at Xiangya School of
Medicine (Changsha, China) and housed in specific pathogen-free
conditions at 22–26°C, under a 12/12 h light/dark cycle with 40–70%
humidity and ad libitum access to food and water. Animal
health and behavior were monitored daily. The following humane
endpoints were established for euthanasia: i) The inability to
access food or water for ~24 h, ii) the inability to remain upright
for ~24 h and iii) a 20% decrease in optimal body weight. All
experimental protocols were approved by the Ethics Committee of
Medical Research, Third Xiangya Hospital of Central South
University (approval no. 2018-S092), and were performed in
accordance with the National Institutes of Health Guidelines for
the Care and Use of Laboratory Animals.
Experimental design
Mice were randomly divided into three groups
(n=6/group) as follows: Control group, DSS group and DSS +
R.I flagellin group. Acute colitis was induced by
administrating 3% (w/v) DSS (molecular weight, 36,000-50,000; MP
Biomedicals, Inc.) in drinking water for 7 days. Based on a
previous study, R.I flagellin was administered once a day
via intraperitoneal injection, at a dose of 50 mg/kg in 150 µl
normal saline to the wild-type (WT) mice with DSS-induced colitis
from day 0 to 7 (21). The control
and DSS groups received 150 µl normal saline. On day 7, ~0.4 ml of
peripheral blood was collected from the eyes of the mice, under
anesthesia. Subsequently, all mice were sacrificed by cervical
dislocation and mortality was confirmed by cardiac arrest,
respiratory arrest and complete loss of nerve reflex. Then, the
peritoneum was opened and colon and spleen tissues were collected.
Colon tissues were quickly used for western blotting or reverse
transcription-quantitative (RT-q)PCR. In addition, colon tissues
were fixed in 4% phosphate-buffered formaldehyde solution at 4°C
for 24 h, embedded in paraffin and then used for histopathological
analysis and immunofluorescence.
Disease activity index (DAI) and
histological scorings of colitis
Mice were weighed daily and visually inspected for
diarrhea and rectal bleeding to assess the DAI. The DAI score was
calculated as the mean of three parameters: Weight loss, 0–4 (0, no
loss; 1, 1–5% loss; 2, 6–10% loss; 3, 11–20% loss; 4, >20%
weight loss); stool consistency, 0–4 (0, normal; 2, loose stool; 4,
diarrhea) and stool bleeding, 0–4 (0, no blood; 2, presence; 4,
gross blood). For hematoxylin and eosin detection, colon tissue
samples were fixed in PBS containing 10% neutral-buffered formalin
(Shanghai Rongbai biological technology Co., Ltd.) at 4°C for 24 h,
embedded in paraffin, cut into 5-µm thick sections and stained with
hematoxylin for 30 sec and eosin for 2 min (both at room
temperature). A light microscope (magnification, ×100; DP72;
Olympus Corporation) was used to observe the colonic
histopathology. Histopathological changes (the changes of colonic
mucosal ulceration, glandular defects and lymphocyte infiltration)
were evaluated using a previously established scoring system
(22).
Immunofluorescence
Colonic tissue sections were incubated with rabbit
anti-NLRP3 primary antibody (1:200; Servicebio, Inc.; cat. no.
GB11300) overnight at 4°C. Sections were subsequently washed with
0.01 M PBS three times, prior to incubation with Cy3 conjugated
donkey anti-rabbit secondary antibody (1:200; Servicebio, Inc.;
cat. no. GB21403; http://www.servicebio.cn/goodsdetail?id=255) for 1 h
at room temperature. Sections were re-washed with 0.01 M PBS,
stained with 2.5 µg/ml DAPI for 20 min at room temperature, and
observed under a confocal laser scanning microscope (Olympus
Corporation) at ×100 magnification.
Cell culture and reagents
THP-1 cells were purchased from the American Type
Culture Collection and maintained in RPMI-1640 medium (Gibco;
Thermo Fisher Scientific, Inc.) supplemented with 10% FBS (Gibco;
Thermo Fisher Scientific, Inc.), at 37°C in a humidified incubator
of 5% CO2. Cells were differentiated into macrophages
via incubation with 150 ng/ml phosphomolybdic acid (Sigma-Aldrich;
Merck KGaA) for 36 h at 37°C. The following concentrations of
agonists or stimuli were used: 1 µg/ml LPS (Sigma-Aldrich; Merck
KGaA), 5 mM ATP (Sigma-Aldrich; Merck KGaA) and 200 ng/ml flagellin
(Detai Biologics Co., Ltd.).
RT-qPCR
Total RNA from colon, peritoneal macrophages or
THP-1 cells was extracted using TRIzol reagent (Invitrogen; Thermo
Fisher Scientific, Inc.), according to the manufacturer's protocol
and reverse-transcribed into cDNA using the Reverse Transcription
kit (Thermo Fisher Scientific, Inc.). The temperature protocol for
miRNA reverse transcription was: 42°C for 60 min, followed by 70°C
for 5 min. qPCR was subsequently performed using the SYBR Green
qPCR Master mix (Vazyme Biotech Co., Ltd.). miRNA expression levels
were assessed using the Mir-X miRNA First-Strand Synthesis and
Mir-X miRNA qRT-PCR TB Green kits (Takara Biotechnology Co., Ltd.).
The temperature protocol for miRNA reverse transcription was: 37°C
for 60 min, followed by 85°C for 5 min and stored at 4°C. The
primer sequences used for qPCR are presented in Table SI. The following thermocycling
conditions were used for qPCR: Initial denaturation: 95°C for 30
sec; followed by 40 cycles at 95°C for 5 sec and 60°C for 30 sec;
Dissociation Curve: 95°C for 60 sec, 55°C for 30 sec and 95°C for
60 sec. Relative gene expression levels were calculated using the
2−ΔΔCq method (23).
mRNA expression was normalized to GAPDH, while miRNA expression was
normalized to the small nuclear RNA, U6.
Prediction of miRNAs that can bind to
NLRP3
The database of miRDB (http://www.mirdb.org; version, 5.0) and TargetScan
(http://www.targetscan.org; version, 7.2)
was used to identify miRNAs which can bind to 3′-untranslated
region (UTR) of NLRP3. Predictions of these two databases
identified miR-5197-3p, miR-223-3p, miR-589-3p and miR-1305 as top
miRNA candidates. The sequences of miRNA were obtained from
TargetScan.
Enzyme-linked immunosorbent assay
(ELISA)
The expression levels of IL-1β, IL-18, TNF-α and
IL-6 in mouse serum were measured via ELISA (IL-1β, cat. no.
PMLB00C; IL-18, cat. no, 7625; TNF-α, cat. no, PMTA00B; IL-6, cat.
no, PM6000B; R&D Systems, Inc.), according to the
manufacturer's protocol. All experiments were performed twice and
repeated three times.
Western blotting
Total protein was extracted from colonic tissue
samples and THP-1 cells by homogenization in RIPA Lysis Buffer
(cat. no. CW2333; CoWin Biosciences), as previously described
(6). Protein concentration was
determined using a bicinchoninic acid assay (Thermo Fisher
Scientific, Inc.). Equal amounts of proteins (40 µg) were
electrophoresed on 12% SDS-PAGE gels and subsequently transferred
onto polyvinylidene difluoride membranes (Merck KGaA). Membranes
were blocked with 5% BSA (Wuhan Goodbio Technology Co., Ltd.) at
room temperature for 1 h, and then incubated with primary
antibodies against: NLRP3 (cat. no. ab214185; Abcam), ASC (cat. no.
13833; Cell Signaling Technology, Inc.), cleaved caspase-1 (cat.
no. 4199; Cell Signaling Technology, Inc.), GSDMD-N (cat. no.
93709; Cell Signaling Technology, Inc.) and GAPDH (cat. no.
10494-1-AP; ProteinTech Group, Inc.) overnight at 4°C (all
1:1,000). Following the primary incubation, membranes were
incubated with HRP-conjugated Affinipure Goat Anti-Rabbit/Mouse IgG
secondary antibody for 1 h at room temperature (1:5,000; cat. nos.
SA00001-2 and SA00001-2; ProteinTech Group, Inc.). Protein bands
were visualized using the ECL kit (Merck KGaA). Densitometry data
were analyzed using ImageJ (version 1.52V; National Institutes of
Health).
TEM
THP-1 cells were fixed with 2.5% glutaraldehyde in
4°C for 24 h, washed with PBS three times and subsequently fixed
with 1% osmium tetroxide in 4°C for 2 h. Following dehydration in a
descending ethanol series (30, 50, 70, 90 and 100%), samples were
embedded in Epon/Araldite mixture and subsequently cut into
50–60-nm-thick sections. Sections were stained with uranium acetate
and lead citrate (these were performed by the TEM assistants at the
Central South University), and observed under a transmission
electron microscope at ×20,000 magnification (cat. no. HT7700;
Hitachi, Ltd.).
Transfection
hsa-miR-223-3p mimic, hsa-miR-223-3p inhibitor,
Toll-like receptor 5 (TLR5) small interfering (si)RNA and scrambled
negative control (NC) were synthesized by Shanghai GenePharma Co.,
Ltd. The sequences are presented in Table SI. Transfection was performed
using Lipofectamine 3000 reagent (Invitrogen; Thermo Fisher
Scientific, Inc.). The concentration of miRNA mimics, inhibitor, NC
and si-TLR5 in these experiment was 100 nM. THP-1 cells were
collected for further experimentation, 48 h post-transfection.
Dual-luciferase reporter assay
The reporter plasmids, pmirGLO-NLRP3-3′-UTR,
pmirGLO-NLRP3-3′-UTR-MUT and pmirGLO-NC were purchased from
Shanghai GenePharma Co., Ltd. THP-1 cells were co-transfected with
the plasmids and hsa-miR-223-3p mimic/inhibitor using Lipofectamine
3000 reagent (Invitrogen; Thermo Fisher Scientific, Inc.).
Following incubation at 37°C for 24 h, luciferase activities were
detected using a Dual-Luciferase Reporter assay system (Promega
Corporation), and results were compared with Renilla
luciferase activity.
Statistical analysis
Statistical analysis was performed using GraphPad
Prism software (version 7.0; GraphPad Software Inc.). Each
experiment was repeated three times. Data are presented as the mean
± standard error of the mean. One-way ANOVA, followed by least
significant difference post-hoc test were performed to compare
differences between multiple groups. P<0.05 was considered to
indicate a statistically significant difference.
Results
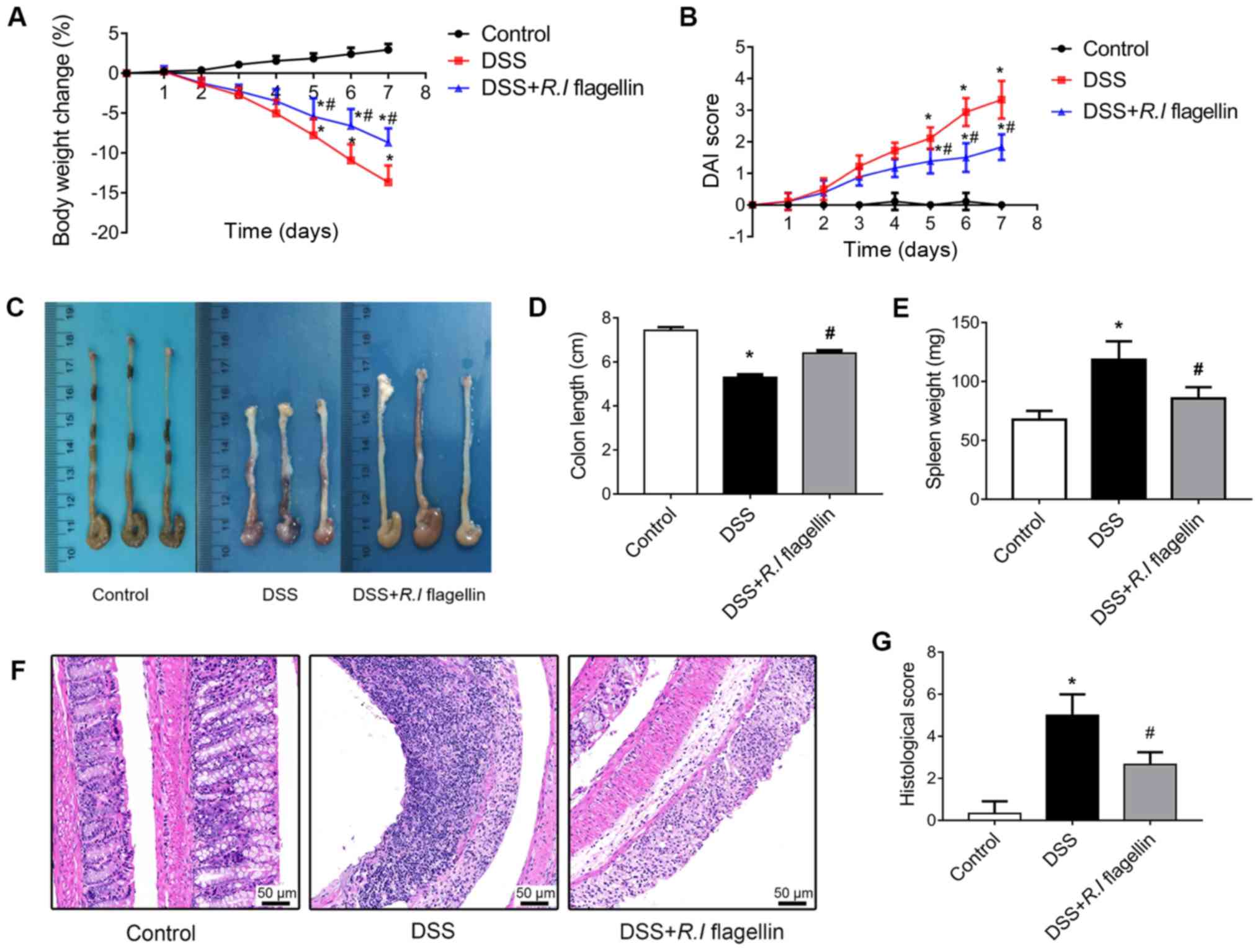
R.I flagellin attenuates inflammation
in a DSS-induced colitis model
In order to investigate the therapeutic effects of
R.I flagellin in vivo, a DSS-induced acute colitis
mouse model was established. DSS administration to 6-week-old mice
induced severe colitis, which was characterized by increased loss
of body weight and higher DAI score compared with the healthy
control mice. Notably, administration of R.I flagellin
significantly restored body weight (Fig. 1A) and DAI score (Fig. 1B) in DSS-treated mice. Furthermore,
the vital indicators of colitis, colonic shortening (Fig. 1C and D) and splenomegaly (Fig. 1E), were evidently mitigated
following administration of R.I flagellin. Consistent with
the remission of clinical signs, treatment with R.I
flagellin decreased mucosal ulceration, glandular defects and
lymphocyte infiltration in colonic tissues (Fig. 1F and G). Furthermore, treatment
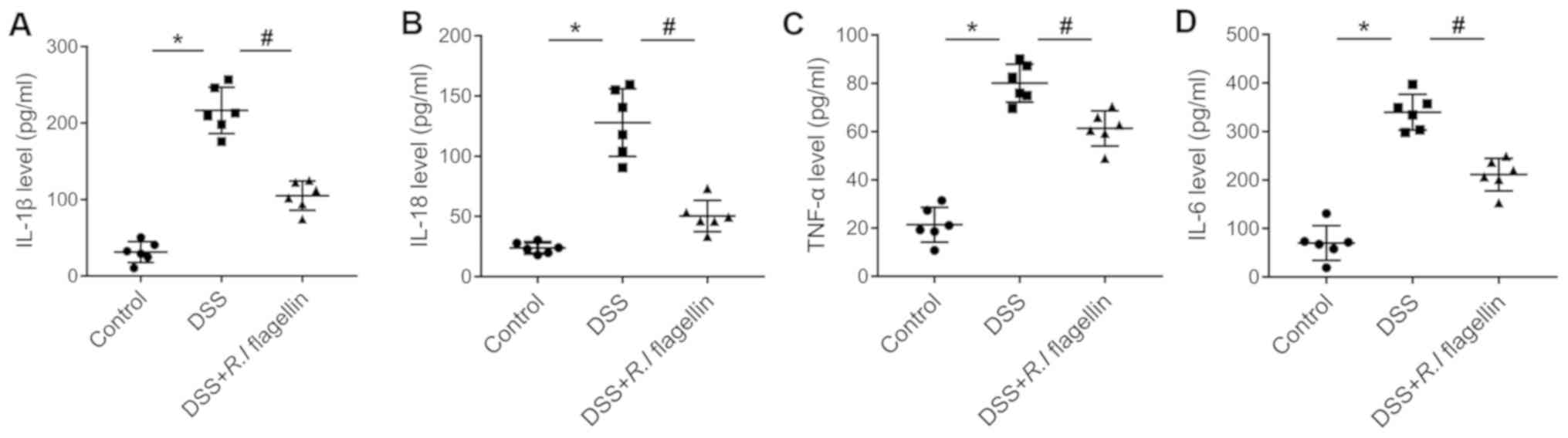
with R.I flagellin significantly inhibited DSS-induced
increased serum levels of IL-1β, IL-18, TNF-α and IL-6 (Fig. 2A-D). Collectively, these results
indicated that R.I flagellin is protective against
DSS-induced colitis.
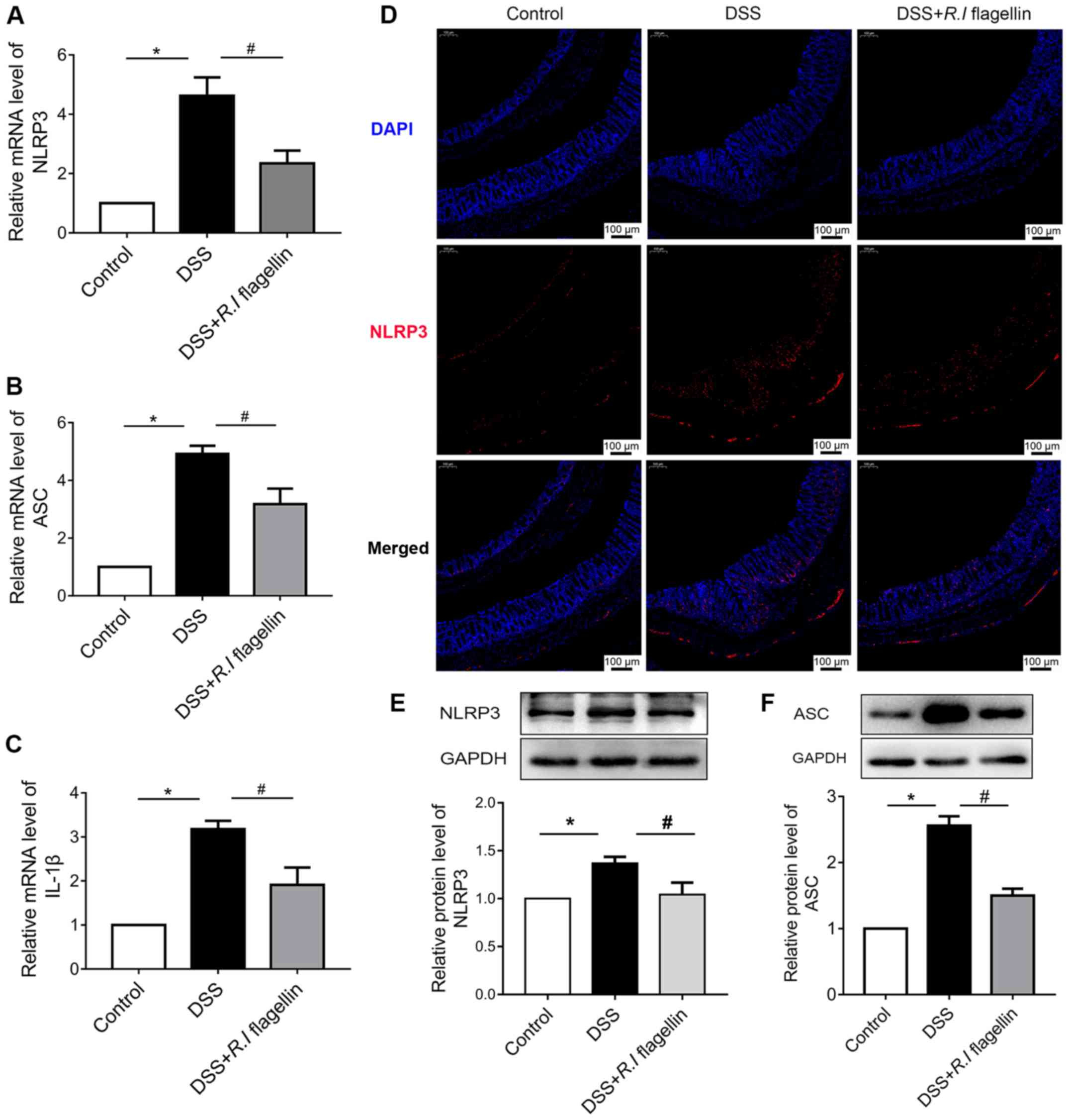
R.I flagellin decreases NLRP3
inflammasome activation in DSS-induced mice
RT-qPCR analysis was performed to assess the
expression levels of NLRP3, ASC and IL-1β in colonic tissues, to
determine whether R.I flagellin relieves inflammation by
regulating activation of the NLRP3 inflammasome. The results
demonstrated that the mRNA expression levels of NLRP3, ASC and
IL-1β were upregulated in the DSS group compared with the control
group, which was significantly reversed following treatment with
R.I flagellin (Fig. 3A-D).
Similarly, immunofluorescence staining and western blot analysis
indicated that NLRP3 distinctly increased in the DSS group, was
significantly counteracted following administration of R.I
flagellin (Fig. 3D and E).
Furthermore, western blot analysis demonstrated that ASC protein
expression significantly increased in the DSS group, was
significantly reversed following treatment with R.I
flagellin (Fig. 3F). ELISA
indicated that treatment with R.I flagellin significantly
inhibited DSS-induced increased serum levels of IL-1β (Fig. 2A); it has been shown that IL-1β is
associated with the NLRP3 inflammasome (24). Collectively, these results
indicated that R.I flagellin affected the inflammasome
activation of colon tissue.
R.I flagellin decreases NLRP3
inflammasome activation and pyroptosis in THP-1 macrophages
Macrophages are the gatekeepers of intestinal immune
homeostasis (25,26). The present study further
investigated the regulation of R.I flagellin on inflammasome
activation in THP-1 macrophages. Transcript and protein expression
levels of NLRP3 and ASC significantly increased following treatment
with LPS+ATP, the effects of which were significantly reversed
following treatment with R.I flagellin (Fig. 4A-E). Furthermore, IL-1β mRNA
expression and secretion significantly increased following
treatment with LPS+ATP compared with the control, the effects of
which were significantly counteracted following administration of
R.I flagellin (Fig. 4H and
I). Collectively, these results confirmed that R.I
flagellin can reverse the LPS+ATP- induced NLRP3 inflammasome
activation in THP-1 macrophages.
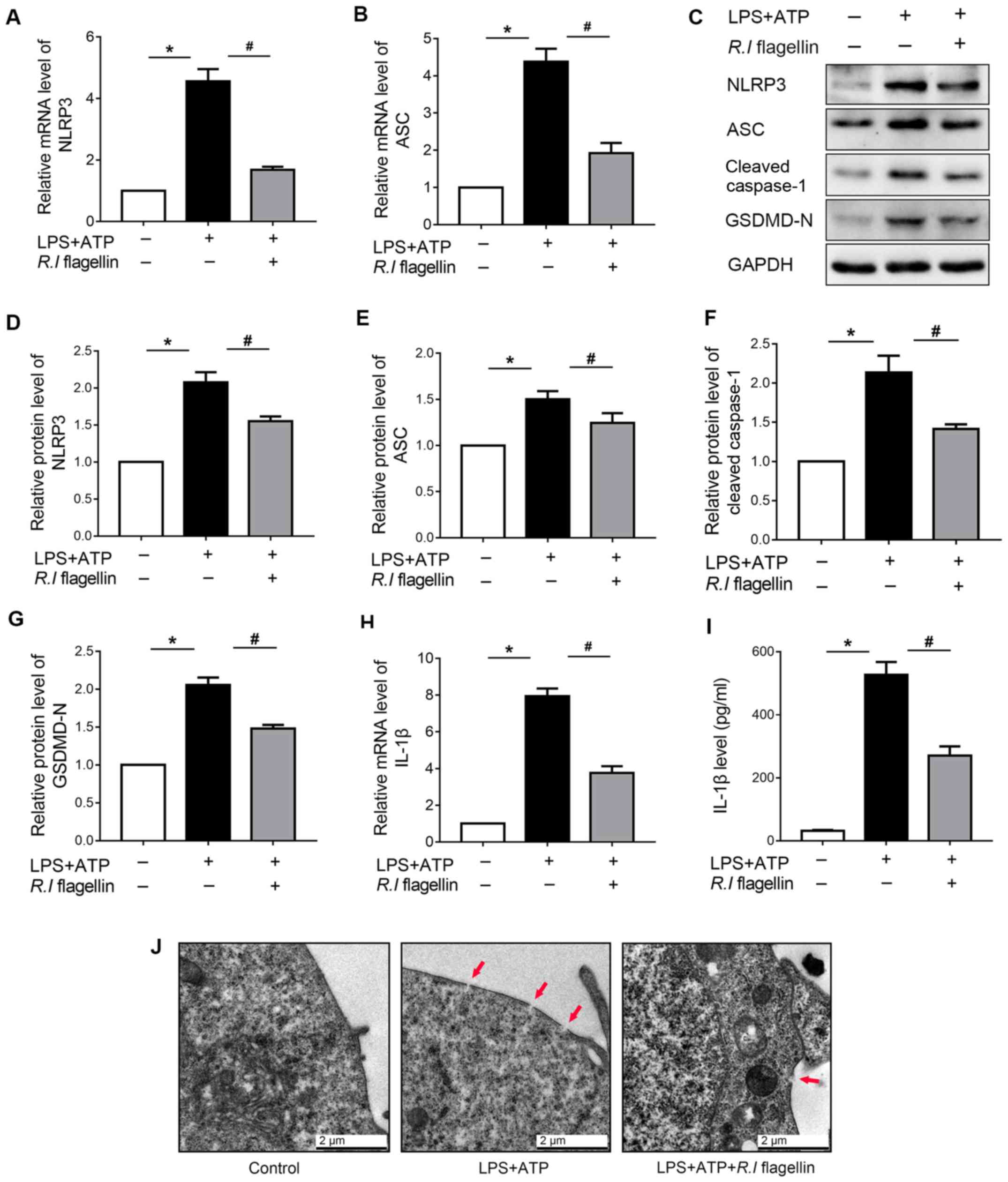 | Figure 4.R.I flagellin decreases NLRP3
inflammasome activation and pyroptosis in THP-1 macrophages.
RT-qPCR analysis was performed to determine mRNA expression levels
of (A) NLRP3 and (B) ASC. (C) Western blot analysis was performed
to determine the protein expression levels of (D) NLRP3, (E) ASC,
(F) cleaved caspase-1 and (G) GSDMD-N in THP-1 macrophages. (H)
RT-qPCR analysis was performed to determine IL-1β mRNA expression
in THP-1 macrophages. (I) IL-1β levels in the culture medium were
detected via the enzyme-linked immunosorbent assay. (J)
Representative electron micrographs of treated THP-1 macrophages
(magnification, ×20,000). Red arrowheads indicate membrane pores.
Data are presented as the mean ± standard error of the mean
(n=3/group). *P<0.05 vs. control group; #P<0.05
vs. LPS+ATP group. R.I, Roseburia intestinalis; NLRP3,
nucleotide-binding oligomerization segment-like receptor family 3;
RT-qPCR, reverse transcription-quantitative PCR; ASC,
apoptosis-associated speck-like protein; GSDMD-N, Gasdermin D
N-terminal fragment; IL, interleukin. |
The regulation of R.I flagellin in
inflammasome-triggered pyroptosis was also investigated. Expression
of the vital markers of pyroptosis, cleaved caspase-1 and GSDMD-N,
significantly increased following LPS+ATP treatment; however, the
expression levels significantly decreased following treatment with
R.I flagellin (Fig. 4C, F and
G). TEM analysis confirmed that GSDMD-N membrane pore decreased
following administration of R.I flagellin in LPS+ATP-induced
pyroptosis (Fig. 4J).
Collectively, these results indicated that R.I flagellin
alleviated LPS+ATP-induced NLRP3 inflammasome activation and
pyroptosis.
Silencing miR-223-3p reverses the
effect of R.I flagellin on NLRP3 inflammasome activation and
pyroptosis
In order to determine the underlying molecular
mechanism by which R.I flagellin mediates regulation of
NLRP3 inflammasome activation in human macrophages, the present
study identified microRNAs (miRNAs/miRs) that bind to NLRP3, using
the miRDB (http://www.mirdb.org) and TargetScan
(http://www.targetscan.org) databases.
THP-1 cells were treated with LPS+ATP and/or R.I flagellin
for 12 h. As presented in Fig. 5A,
treatment with LPS+ATP significantly downregulated the expression
levels of miR-223-3p and miR-589-3p compared with the control
group. However, only miR-223-3p exhibited a significant increase in
expression following treatment with R.I flagellin, compared
with the LPS+ATP group. The other miRNAs (miR-5197-3p and miR-1305)
were not markedly altered.
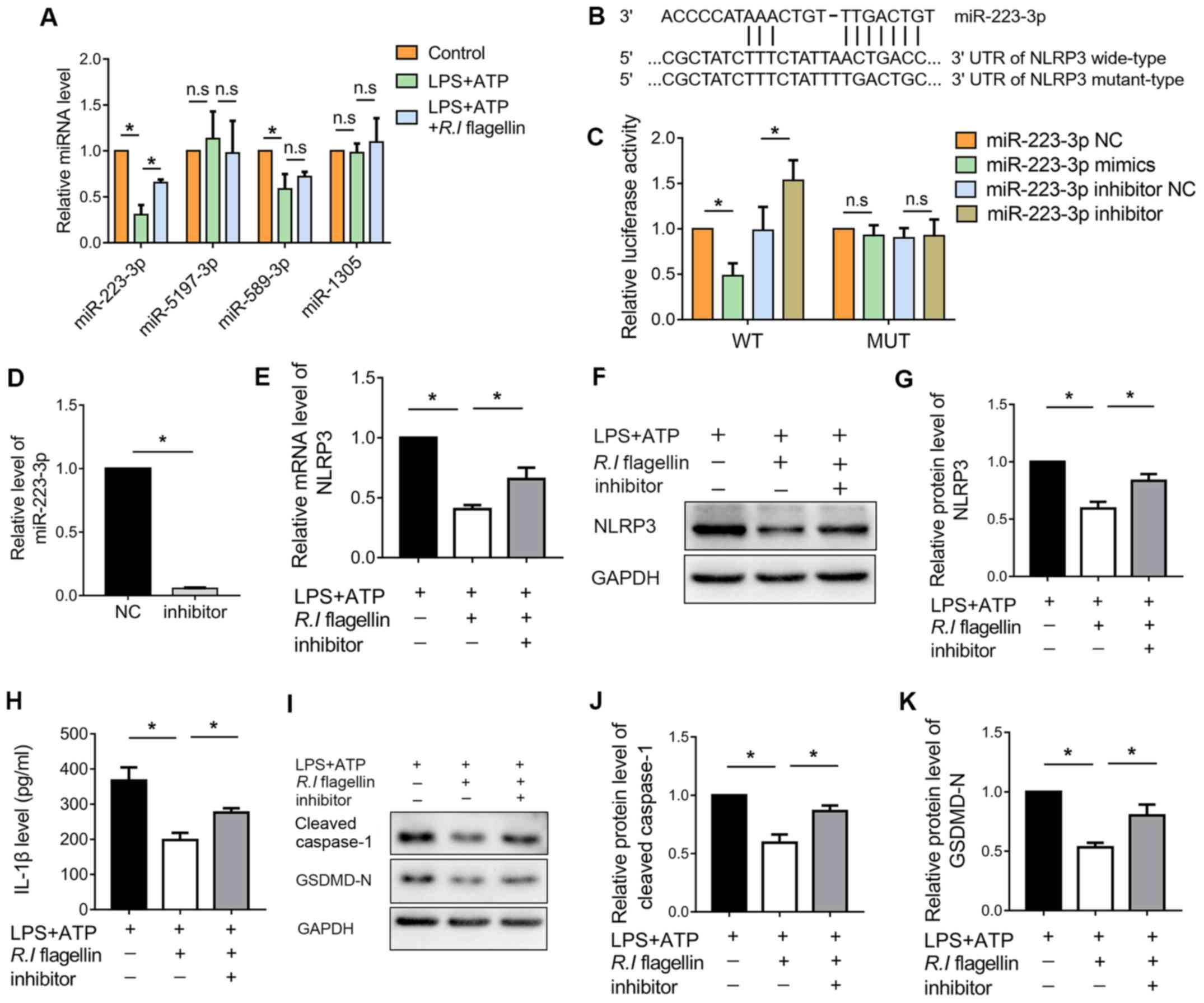 | Figure 5.Silencing miR-223-3p reverses the
effect of R.I flagellin on NLRP3 inflammasome activation and
pyroptosis in THP-1 macrophages. (A) RT-qPCR analysis was performed
to determine the expression levels of miR-223-3p, miR-5197-3p,
miR-589-3p and miR-1305. (B) Schematic diagram of the predicted
binding sequence of miR-223-3p in the 3′-UTR of NLRP3. (C)
Luciferase activity for the WT and MUT 3′-UTR of NLRP3 pGL3 vector
co-transfected with miR-223-3p NC, miR-223-3p mimics, miR-223-3p
inhibitor NC and miR-223-3p inhibitor. RT-qPCR analysis was
performed to determine the expression levels of (D) miR-223-3p and
(E) NLRP3. (F and G) Western blot analysis was performed to
determine NLRP3 protein expression. (H) IL-1β levels in the culture
medium were detected via the enzyme-linked immunosorbent assay. (I)
Western blot analysis was performed to determine the protein
expression levels of (J) cleaved caspase-1 and (K) GSDMD-N. Data
are presented as the mean ± standard error of the mean (n=3/group).
*P<0.05. miR/miRNA, microRNA; R.I, Roseburia
intestinalis; NLRP3, nucleotide-binding oligomerization
segment-like receptor family 3; RT-qPCR, reverse
transcription-quantitative PCR; IL, interleukin; GSDMD-N, Gasdermin
D N-terminal fragment; n.s, no significance; UTR, untranslated
region; NC, negative control; WT, wild-type; MUT, mutant. |
The binding sequence of miR-223-3p in the NLRP3
3′-UTR was predicted to determine if NLRP3 was a downstream target
of miR-223-3p in THP-1 cells (Fig.
5B). As presented in Fig. 5C,
the dual-luciferase reporter assay demonstrated that binding of
miR-223-3p mimic to the WT NLRP3 3′-UTR significantly decreased
luciferase activity, while binding of miR-223-3p inhibitor to the
WT NLRP3 3′-UTR significantly increased luciferase activity in
THP-1 cells, compared with the respective controls. However,
binding of miR-223-3p mimic or inhibitor to MUT NLRP3 3′-UTR had no
significant effect on luciferase activity.
In order to determine whether miR-223-3p was
involved in activation of the NLRP3 inflammasome, THP-1 cells were
treated with LPS+ATP and/or R.I flagellin for 12 h following
transfection with miR-223-3p inhibitor for 24 h. As anticipated,
transfection with miR-223-3p inhibitor significantly downregulated
miR-223-3p expression in THP-1 cells (Fig. 5D). The inhibitory effect of
R.I flagellin on NLRP3 was reversed by downregulating
miR-223-3p (Fig. 5E-G).
Furthermore, inhibiting miR-223-3p expression counteracted the
restraint of R.I flagellin on the release of IL-1β (Fig. 5H). Collectively, these results
indicated that miR-223-3p was involved in the regulation of
R.I flagellin on NLRP3 inflammasome activation in THP-1
cells.
The present study also investigated the effect of
miR-223-3p on R.I flagellin-regulated pyroptosis. The
results indicated that miR-223-3p knockdown reversed R.I
flagellin-mediated downregulation of cleaved caspase-1 and GSDMD-N
(Fig. 5I-K). Collectively, these
results indicated that R.I flagellin alleviated
inflammasome-induced pyroptosis by inhibiting miR-223-3p/NLRP3
signaling in macrophages.
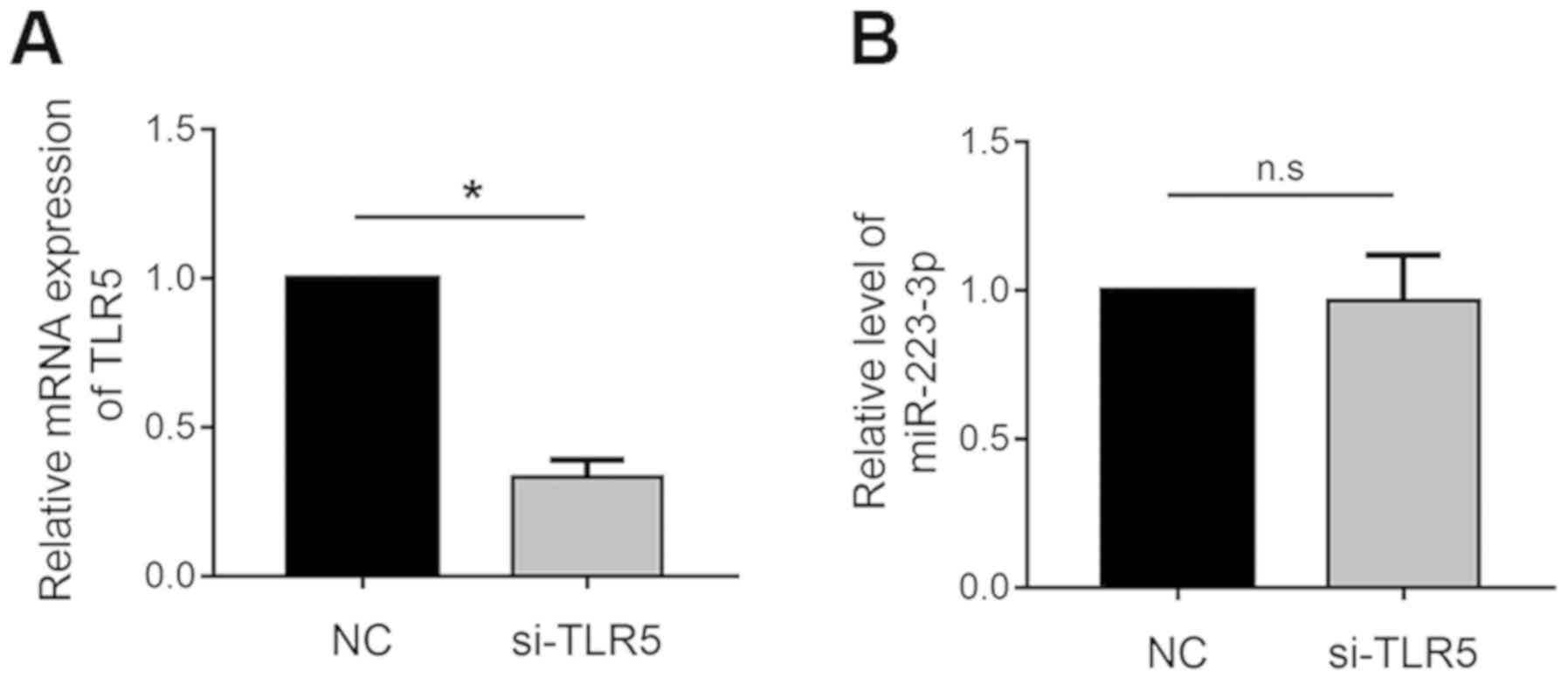
R.I flagellin may regulate miR-223-3p
expression through TLRs but not TLR5
TLR5 acts as a pattern recognition receptor and
responds to flagellin in order to modulate miRNA expression
(27,28). The present study aimed to
investigate whether the regulation of miR-223-3p expression by
R.I flagellin is mediated through the activity of TLR5. TLR5
gene expression was knocked down in THP-1 macrophages (Fig. 6A). However, transfection with
siRNA-TLR5 had no effect on the regulation of miR-223-3p expression
by R.I flagellin (Fig. 6B).
Collectively, these results indicated that R.I flagellin may
regulate miR-223-3p expression through TLRs but not TLR5
specifically.
Discussion
The present study demonstrated that: i) R.I
flagellin exhibited beneficial effects on colitis, ii) R.I
flagellin inhibited inflammasome activation, iii) R.I
flagellin was a negative regulator of inflammasome-triggered
pyroptosis and (4) miR-223-3p was
an upstream signal molecule of NLRP3 that plays a key role in the
anti-inflammatory mechanism of R.I flagellin. Collectively,
these results indicate a potential clinical application of
R.I flagellin to relieve intestinal inflammation.
The disorder of gut microbial communities has been
identified as the core defect that causes chronic intestinal
inflammation (4,5). Several studies have suggested that
some of the intestinal bacteria play important roles in relieving
intestinal inflammation and maintaining intestinal barrier
integrity (29–31). A previous study demonstrated that
R.I levels were significantly decreased in untreated
patients with IBD. Furthermore, R.I was reported to
alleviate colitis pathology by maintaining the Treg/Th17 balance in
an experimental colitis model (6).
However, the role of R.I flagellin, as the main structural
component of R.I, in relieving colitis remains unclear. The
results of the present study demonstrated that R.I flagellin
alleviated DSS-induced inflammation in a mouse model of colitis.
Furthermore, cytokines such as IL-1β, IL-18, TNF-α and IL-6 are
significantly elevated in UC, some of which have become important
therapeutic targets (32,33). The results of the present study
indicated that treatment with R.I flagellin significantly
inhibited these inflammatory cytokines, indicating that R.I
flagellin has a therapeutic effect on colitis. These results are
consistent with the anti-inflammatory effect of R.
intestinalis, suggesting that R.I-derived flagellin may
be the key component of R.I in inhibiting inflammation.
Contrary to studies that suggest that flagellin may
be an exogenous substance that triggers inflammation (34–36),
increasing evidence indicates that flagellins play a protective
role in inflammation and intestinal barrier functions (9,10,37,38).
For example, Salmonella typhimurium-derived flagellin can
induce the secretion of anti-inflammatory factors, such as IL-22,
from dendritic cells to protect intestinal epithelial barrier
(9). Thus, flagellins from
different sources may play different roles in regulating
inflammatory immunity. The results of the present study confirm the
role of flagellin as a biological agent for the treatment of
colitis.
Recent studies suggest that pyroptosis plays a
pivotal role in the occurrence and development of UC. Davis et
al (39) demonstrated that
intestinal epithelial cell pyroptosis is vital for the development
of epithelial barrier dysfunction and mucosal inflammation.
Furthermore, inhibition of pyroptosis may be a therapeutic
mechanism of action for mesalamine and corticosteroids (19). However, little is known about the
effect of probiotics and their components on pyroptosis. The
results of the present study verified that R.I flagellin can
alleviate NLRP3 inflammasome activation and inflammasome-triggered
pyroptosis.
miRNA-mediated regulation of several immune
processes, including pyroptosis have been extensively studied and
well established (40,41). The present study used miRDB and
TargetScan databases to identify miRNAs that have binding sites to
NLRP3, and demonstrated that the decreased miR-223-3p expression
was significantly altered following administration of R.I
flagellin. Furthermore, inhibiting miR-223-3p expression reversed
the effects of R.I flagellin on suppressing activation of
the NLRP3 inflammasome and downregulating expression of
pyroptosis-associated proteins. Collectively, these results
indicated that R.I flagellin alleviated inflammasome-induced
pyroptosis by inhibiting miR-223-3p/NLRP3 signaling.
Several studies have reported that TLR signaling can
modulate miRNA expression (27,42).
TLR5 plays an important role in innate immune responses to
flagellin (28,43). However, the results of the present
study demonstrated that transfection with si-TLR5 had no effect on
the regulation of miR-223-3p expression by R.I flagellin.
Previous studies have reported that flagellin is recognized by
other receptors, such as TLR2, NLRC4 and TLR11 (34,44,45).
Thus, it is suggested that R.I flagellin may regulate
miR-223-3p expression through TLRs but not TLR5. Prospective
studies will focus on investigating the association between other
recognition receptors and R.I flagellin through experiments
such as pull-down and mass spectrometry. Moreover, although this
study preliminarily investigated the inhibitory effect of flagellin
on colitis and its mechanism of inflammation suppression, its
practical role in patients with UC remains to be elucidated; this
is an important direction of follow-up research, and is crucial for
the clinical transformation application of R.I
flagellin.
In conclusion, the results of the present study
demonstrated that R.I flagellin relieved intestinal
inflammation by inhibiting inflammasome-induced pyroptosis through
miR-223-3p/NLRP3 signaling. Thus, R.I flagellin exhibits a
beneficial probiotic role in ameliorating UC.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was funded by the National Natural
Science Foundation of China (grant nos. 81670504 and 81970494), the
Fundamental Research Funds for the Central Universities of Central
South University (grant no. 502211903) and the Key Research and
Development Program of Hunan Province (grant no. 2019SK2041).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author upon reasonable
request.
Authors' contributions
XW, SP and WL performed the experiments. XM, MX, BT,
KN and TT analyzed the data, and XW prepared the manuscript. ZS
contributed to the data analysis and the revision of the
manuscript. XYW contributed to the conception of the study, and
guided the design and conduction of the study. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
All experimental protocols were approved by the
Ethics Committee of Medical Research, Third Xiangya Hospital of
Central South University (Changsha, China; approval no. 2018-S092),
and were performed in accordance with the National Institutes of
Health Guidelines for the Care and Use of Laboratory Animals.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Ungaro R, Mehandru S, Allen PB,
Peyrin-Biroulet L and Colombel JF: Ulcerative colitis. Lancet.
389:2695–1770. 2017. View Article : Google Scholar
|
|
2
|
Ananthakrishnan AN: Epidemiology and risk
factors for IBD. Nat Rev Gastroenterol Hepatol. 12:205–217. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Neurath MF: Current and emerging
therapeutic targets for IBD. Nat Rev Gastroenterol Hepatol.
14:269–278. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Marchesi JR, Adams DH, Fava F, Hermes GD,
Hirschfield GM, Hold G, Quraishi MN, Kinross J, Smidt H, Tuohy KM,
et al: The gut microbiota and host health: A new clinical frontier.
Gut. 65:330–339. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sartor RB and Wu GD: Roles for intestinal
bacteria, viruses, and fungi in pathogenesis of inflammatory bowel
diseases and therapeutic approaches. Gastroenterology.
152:327–339.e4. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Shen Z, Zhu C, Quan Y, Yang J, Yuan W,
Yang Z, Wu S, Luo W, Tan B and Wang X: Insights into Roseburia
intestinalis which alleviates experimental colitis pathology by
inducing anti-inflammatory responses. J Gastroenterol Hepatol.
33:1751–1760. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tan B, Luo W, Shen Z, Xiao M, Wu S, Meng
X, Wu X, Yang Z, Tian L and Wang X: Roseburia intestinalis
inhibits oncostatin M and maintains tight junction integrity in a
murine model of acute experimental colitis. Scand J Gastroenterol.
54:432–440. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhu C, Song K, Shen Z, Quan Y, Tan B, Luo
W, Wu S, Tang K, Yang Z and Wang X: Roseburia intestinalis
inhibits interleukin17 excretion and promotes regulatory T cells
differentiation in colitis. Mol Med Rep. 17:7567–7574.
2018.PubMed/NCBI
|
|
9
|
Kinnebrew MA, Buffie CG, Diehl GE,
Zenewicz LA, Leiner I, Hohl TM, Flavell RA, Littman DR and Pamer
EG: Interleukin 23 production by intestinal CD103(+)CD11b(+)
dendritic cells in response to bacterial flagellin enhances mucosal
innate immune defense. Immunity. 36:276–287. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhang B, Chassaing B, Shi Z, Uchiyama R,
Zhang Z, Denning TL, Crawford SE, Pruijssers AJ, Iskarpatyoti JA,
Estes MK, et al: Viral infection. Prevention and cure of rotavirus
infection via TLR5/NLRC4-mediated production of IL-22 and IL-18.
Science. 346:861–865. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bergsbaken T, Fink SL and Cookson BT:
Pyroptosis: Host cell death and inflammation. Nat Rev Microbiol.
7:99–109. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Shi J, Gao W and Shao F: Pyroptosis:
Gasdermin-mediated programmed necrotic cell death. Trends Biochem
Sci. 42:245–254. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lu A, Magupalli VG, Ruan J, Yin Q,
Atianand MK, Vos MR, Schröder GF, Fitzgerald KA, Wu H and Egelman
EH: Unified polymerization mechanism for the assembly of
ASC-dependent inflammasomes. Cell. 156:1193–1206. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wree A, Eguchi A, McGeough MD, Pena CA,
Johnson CD, Canbay A, Hoffman HM and Feldstein AE: NLRP3
inflammasome activation results in hepatocyte pyroptosis, liver
inflammation, and fibrosis in mice. Hepatology. 59:898–910. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Liu T, Zhou YT, Wang LQ, Li LY, Bao Q,
Tian S, Chen MX, Chen HX, Cui J and Li CW: NOD-like receptor
family, pyrin domain containing 3 (NLRP3) contributes to
inflammation, pyroptosis, and mucin production in human airway
epithelium on rhinovirus infection. J Allergy Clin Immunol.
144:777–787 e779. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Thornberry NA, Bull HG, Calaycay JR,
Chapman KT, Howard AD, Kostura MJ, Miller DK, Molineaux SM, Weidner
JR, Aunins J, et al: A novel heterodimeric cysteine protease is
required for interleukin-1 beta processing in monocytes. Nature.
356:768–774. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Evavold CL, Ruan J, Tan Y, Xia S, Wu H and
Kagan JC: The pore-forming protein gasdermin D regulates
interleukin-1 secretion from living macrophages. Immunity.
48:35–44.e6. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
He WT, Wan H, Hu L, Chen P, Wang X, Huang
Z, Yang ZH, Zhong CQ and Han J: Gasdermin D is an executor of
pyroptosis and required for interleukin-1β secretion. Cell Res.
25:1285–1298. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Davis EM, Zhang D, Glover SC, Stappenbeck
T, Wang SZ and Liu JJ: Inhibition of intestinal epithelial cell
pyroptosis and associated mucosal barrier defects is a potential
therapeutic mechanism of action for mesalamine in Ibd.
Gastroenterology. 156 (Suppl):S882019. View Article : Google Scholar
|
|
20
|
Ren TH, Zhou Y and Wu WY: Activation of
adenosine A3 receptor inhibits Nlrp3 inflammasome and pyroptosis of
colonic epithelial cells of patients with ulcerative colitis.
Gastroenterology. 156 (Suppl):S6272019. View Article : Google Scholar
|
|
21
|
Quan Y, Song K, Zhang Y, Zhu C, Shen Z, Wu
S, Luo W, Tan B, Yang Z and Wang X: Roseburia
intestinalis-derived flagellin is a negative regulator of
intestinal inflammation. Biochem Biophys Res Commun. 501:791–799.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wirtz S, Popp V, Kindermann M, Gerlach K,
Weigmann B, Fichtner-Feigl S and Neurath MF: Chemically induced
mouse models of acute and chronic intestinal inflammation. Nat
Protoc. 12:1295–1309. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Mangan MSJ, Olhava EJ, Roush WR, Seidel
HM, Glick GD and Latz E: Targeting the NLRP3 inflammasome in
inflammatory diseases. Nat Rev Drug Discov. 17:6882018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Koelink PJ, Bloemendaal FM, Li B, Westera
L, Vogels EWM, van Roest M, Gloudemans AK, van ′t Wout AB, Korf H,
Vermeire S, et al: Anti-TNF therapy in IBD exerts its therapeutic
effect through macrophage IL-10 signalling. Gut. 69:1053–1063.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Na YR, Stakenborg M, Seok SH and Matteoli
G: Macrophages in intestinal inflammation and resolution: A
potential therapeutic target in IBD. Nat Rev Gastroenterol Hepatol.
16:531–543. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
O'Neill LA, Sheedy FJ and McCoy CE:
MicroRNAs: The fine-tuners of Toll-like receptor signalling. Nat
Rev Immunol. 11:163–175. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yoon SI, Kurnasov O, Natarajan V, Hong M,
Gudkov AV, Osterman AL and Wilson IA: Structural basis of
TLR5-flagellin recognition and signaling. Science. 335:859–864.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Atarashi K, Tanoue T, Oshima K, Suda W,
Nagano Y, Nishikawa H, Fukuda S, Saito T, Narushima S, Hase K, et
al: Treg induction by a rationally selected mixture of Clostridia
strains from the human microbiota. Nature. 500:232–236. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Furusawa Y, Obata Y, Fukuda S, Endo TA,
Nakato G, Takahashi D, Nakanishi Y, Uetake C, Kato K, Kato T, et
al: Commensal microbe-derived butyrate induces the differentiation
of colonic regulatory T cells. Nature. 504:446–450. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Mardini HE and Grigorian AY: Probiotic mix
VSL#3 is effective adjunctive therapy for mild to moderately active
ulcerative colitis: A meta-analysis. Inflamm Bowel Dis.
20:1562–1567. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Haas SL, Abbatista M, Brade J, Singer MV
and Böcker U: Interleukin-18 serum levels in inflammatory bowel
diseases: Correlation with disease activity and inflammatory
markers. Swiss Med Wkly. 139:140–145. 2009.PubMed/NCBI
|
|
33
|
Monteleone G, Pallone F and Caprioli F:
Investigational cytokine-targeted therapies for ulcerative colitis.
Expert Opin Investig Drugs. 22:1123–1132. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Xie Y, Xu M, Xiao Y, Liu Z, Jiang C, Kuang
X, Wang C, Wu H, Peng J, Li C, et al: Treponema pallidum flagellin
FlaA2 induces IL-6 secretion in THP-1 cells via the Toll-like
receptor 2 signaling pathway. Mol Immunol. 81:42–51. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Nakamoto K, Watanabe M, Sada M, Inui T,
Nakamura M, Honda K, Wada H, Ishii H and Takizawa H: Pseudomonas
aeruginosa-derived flagellin stimulates IL-6 and IL-8 production in
human bronchial epithelial cells: A potential mechanism for
progression and exacerbation of COPD. Exp Lung Res. 45:255–266.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Whitehead GS, Hussain S, Fannin R, Trempus
CS, Innes CL, Schurman SH, Cook DN and Garantziotis S: TLR5
activation exacerbates airway inflammation in asthma. Lung.
198:289–298. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Crellin NK, Garcia RV, Hadisfar O, Allan
SE, Steiner TS and Levings MK: Human CD4+ T cells express TLR5 and
its ligand flagellin enhances the suppressive capacity and
expression of FOXP3 in CD4+CD25+ T regulatory cells. J Immunol.
175:8051–8059. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kim EH, Kim JH, Samivel R, Bae JS, Chung
YJ, Chung PS, Lee SE and Mo JH: Intralymphatic treatment of
flagellin-ovalbumin mixture reduced allergic inflammation in murine
model of allergic rhinitis. Allergy. 71:629–639. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Davis EM, Kaufmann Y, Goyne H, Claggett
BL, Jobin C and Liu JJ: Pyroptosis of intestinal epithelial cells
Is crucial to the development of mucosal barrier dysfunction and
intestinal inflammation. Gastroenterology. 152 (Suppl 1):S9672017.
View Article : Google Scholar
|
|
40
|
Kalla R, Ventham NT, Kennedy NA, Quintana
JF, Nimmo ER, Buck AH and Satsangi J: MicroRNAs: New players in
IBD. Gut. 64:504–517. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Lee S, Choi E, Cha MJ and Hwang KC:
Looking for pyroptosis-modulating miRNAs as a therapeutic target
for improving myocardium survival. Mediators Inflamm.
2015:2548712015. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Yang Y, Weng W, Peng J, Hong L, Yang L,
Toiyama Y, Gao R, Liu M, Yin M, Pan C, et al: Fusobacterium
nucleatum increases proliferation of colorectal cancer cells and
tumor development in mice by activating toll-like receptor 4
signaling to nuclear Factor-κB, and up-regulating expression of
MicroRNA-21. Gastroenterology. 152:851–866.e24. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Chassaing B, Ley RE and Gewirtz AT:
Intestinal epithelial cell toll-like receptor 5 regulates the
intestinal microbiota to prevent low-grade inflammation and
metabolic syndrome in mice. Gastroenterology. 147:1363–1377.e17.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Carvalho FA, Nalbantoglu I, Aitken JD,
Uchiyama R, Su Y, Doho GH, Vijay-Kumar M and Gewirtz AT: Cytosolic
flagellin receptor NLRC4 protects mice against mucosal and systemic
challenges. Mucosal Immunol. 5:288–298. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Hatai H, Lepelley A, Zeng W, Hayden MS and
Ghosh S: Toll-like receptor 11 (TLR11) interacts with flagellin and
profilin through disparate mechanisms. PLoS One. 11:e01489872016.
View Article : Google Scholar : PubMed/NCBI
|