Introduction
The gene sequence of scavenger receptor class B type
I (SR-BI) was first determined by Calvo and Vega (1) in 1993 and named CLA-1 (now
SCARB1). SR-BI shares close sequence identity with cluster
determinant 36 and lysosomal integral membrane protein 2 (LIMP-2).
Acton et al (2) proposed
that SR-BI was a high-affinity high-density lipoprotein (HDL)
physiological membrane protein receptor that mediated the selective
uptake of cholesterol esters (CEs) in HDL particles via
hepatocytes. Subsequent research has focused on the transcriptional
regulation, structure and function of SR-BI. SR-BI is highly
conserved across different species, and is widely expressed in
different tissues, including liver, gonad, adrenal gland,
intestinal tract, macrophages, endothelial cells (ECs), platelets,
placenta, adipocytes and smooth muscle cells (3). SR-BI is a member of the scavenger
receptor superfamily and can recognize a variety of ligands,
including HDL, natural and modified low-density lipoprotein (LDL),
very-low-density lipoprotein (VLDL), anion-containing
phospholipids, lipoprotein a (Lpa), phosphatidylserine (PS) and
apoptotic cells (4). SR-B1 has
also been demonstrated to exhibit a high affinity for native HDL
(4,5).
The main physiological function of SR-BI is binding
with HDL to mediate the selective uptake and reverse transport of
cholesterol (3). When circulating
LDL particles bind to the LDL receptor (LDLR), cholesterol is
delivered to cells via endocytosis of the holo-particle (3). After endocytosis, CEs carried in LDL
are hydrolysed by acid lipase in the lysosome, releasing free
cholesterol (FC) that is transported to the endoplasmic reticulum,
where it is either re-esterified to CEs and stored in lipid
droplets in the cytoplasm, or transported to the plasma membrane
(3). In contrast, the mechanism by
which CEs in HDL particles enter the cell is completely different
from internalisation of the CE-rich lipoprotein particles, which is
mediated by LDLR. After HDL particles bind to SR-BI on the cell
surface, CEs, FC and triglycerides are selectively delivered to the
inside of the cell through hydrophobic channels formed by the
extracellular domain of the SR-BI molecule, while the whole
lipoprotein particle remains outside the cell (2,6).
This process results in the selective uptake of cholesterol. In
steroidogenic cells, CEs that enter the cell in this way are
hydrolysed by hormone-sensitive lipases before being used (7).
HDL is an anti-atherosclerotic lipoprotein, possibly
due to its role in reverse cholesterol transport (RCT) (6). A meta-analysis indicated that a
decrease in plasma HDL was associated with increased occurrence of
atherosclerosis and cardiovascular diseases (8). Excess cholesterol in macrophages or
foam cells in atherosclerotic lesions can be transferred to HDL
particles through unidirectional outflow mediated by ATP-binding
cassette A1 and ATP-binding cassette G1 (9), or flow to mature HDL particles via
passive diffusion through hydrophobic channels formed by the
extracellular domain of SR-B1 molecules (9). Both mechanisms decrease the storage
of CEs in cells and reduce the incidence of atherosclerosis and the
risk of cardiovascular diseases (9). Mature HDL particles circulate through
the blood to the liver, where they interact with SR-BI on the
hepatocyte's membrane (10). This
reverse transport of cholesterol to the liver results in the
synthesis of bile acids for secretion into the lumen of the
intestine and the ensuing excretion of excess cholesterol from the
body (10). Interactions between
HDL and SR-BI on the hepatocyte membrane and the selective uptake
of CEs are crucial RCT steps, which serve a critical role in
cardiovascular protection (11).
Structure of SR-BI
The human SCARB1 gene encoding the SR-BI
protein is located at 12q24 on chromosome 12, is ~87 kb in length
and comprises 13 exons and 12 introns (3). SR-B1 has a predicted molecular weight
of 57 kDa and contains multiple glycosylation sites (3). However, due to post-translational
modification, it migrates at ~82 kDa when assessed using SDS-PAGE
(12).
The human SCARB1 mRNA yields five protein
variants through alternative splicing (3). Isoform 1 is the main gene product,
consisting of 509 amino acid (aa) residues from exons 1–12. This
was the first isoform identified and named SR-BI (3). Isoform 2 comprises 409 aa residues
and has the same C-terminus as isoform 1 but lacks aa residues
43–142. Isoform 3 is the longest variant with 552 aa residues.
Isoform 4 (also called SR-BIII) is a polypeptide comprised of 474
aa, with the same C-terminus as isoform 1, but a different
N-terminus (aa 1–42). Isoform 5 (also called SR-BII) comprises 506
aa residues and has a unique C-terminus (aa 468–552).
SR-B1 has two short intracellular domains at the
N-terminus (aa 1–11) and C-terminus (aa 462–509), two transmembrane
domains (aa 12–32 and 441–461) and a large extracellular domain
containing multiple N-linked glycosylation sites (3). The C-terminal cytoplasmic domain is
essential for SR-BI signal transduction and contains kinase
phosphorylation sites and protein binding sites, as well as a
serine that can be phosphorylated by the kinase SIK-1, which
increases the efficiency of CEs uptake mediated by SR-BI (3).
A common C-terminal sequence (aa 468–552) in isomers
1, 2 and 4 includes the VLQEAKL motif that is required for binding
to the PDZ domain-containing protein PDZK1, which is essential for
maintaining the stability and function of SR-BI (13). The other C-terminal sequence (aa
487–494), IQAYSESL, is a motif required to bind the guanine
nucleotide exchange factor dedicator of cytokinesis 4 (DOCK4),
which is essential for the uptake and transendocytosis of LDL in
ECs (14). Determination of the
crystal structure of LIMP-2, a homologue of SR-B1, provided a basic
framework for the structure of this protein family (15), which revealed that the main
extracellular domain of SR-BI contained an antiparallel β-barrel
core and multiple short α-helical structures that facilitated the
formation of a hydrophobic channel and promoted cholesterol
transport.
SR-BI in hepatocytes
SR-BI on the hepatocyte membrane mediates the
selective uptake of CEs in plasma HDL particles and the RCT
process. SR-B1 is also known to regulate lipoprotein homeostasis,
inhibit the development of atherosclerosis and reduce the incidence
of cardiovascular diseases (16,17).
Van Eck et al (18) established SR-BI knockout
(SR-BI−/−) mice and fed them a western-type diet for 20
weeks. Compared with wild-type mice, cholesterol levels in the
liver and the expression of key regulatory factors related to
cholesterol homeostasis [3-hydroxy-3-methyl-glutaryl-coenzyme A
(HMG-CoA) reductase, LDLR and cholesterol 7α-hydroxylase] were not
affected, but a large amount of cholesterol-rich HDL accumulated in
the circulatory system along with abnormally large HDL particles,
which indicated impaired cholesterol delivery to the liver.
SR-BI-deficient mice displayed an ~40% reduction in biliary
cholesterol content, lipid deposition in the aortic wall and the
increased occurrence of atherosclerosis, which indicated that the
RCT process was damaged and the cholesterol clearance capacity was
impaired (18). Apolipoprotein E
(ApoE) single-knockout mice did not usually suffer early
atherosclerosis, severe cardiac dysfunction, cardiac infarction or
premature death when fed a standard diet. However, SR-BI and ApoE
double gene knockout mice experienced severe hypercholesterolemia
after ingesting a standard diet, and atherosclerotic lesions
appeared in the early stages (4–5 weeks), resulting in coronary
artery occlusion, myocardial infarction and cardiac dysfunction,
with most mice dying at 5–8 weeks of age (19,20).
Moreover, after a period of high-fat and high-cholesterol
consumption, SR-BI and LDLR double gene knockout mice exhibited
higher cholesterol expression levels, more severe atherosclerotic
lesions and a higher incidence of myocardial infarction and
mortality, compared with LDLR knockout (LDLR−/−) mice in
the control group (21). These
studies indicated that SR-BI serves an important role in the
prevention of atherosclerosis and the reduction of cardiovascular
diseases, partly due to selective cholesterol uptake and RCT
mediated by SR-BI on the hepatocyte membrane.
SR-BI on hepatocytes is beneficial to cardiovascular
fitness through the regulation of HDL cholesterol metabolism
(22). Liver-specific SR-BI
deficiency induced atherosclerotic lesions, while overexpression of
SR-BI in hepatocytes decreased atherosclerosis (22). After transient overexpression of
SR-BI in hepatocytes, LDLR−/− mice fed with a high-fat
and high-cholesterol diet displayed a marked reduction in early and
advanced atherosclerotic lesions, and also experienced
significantly decreased HDL cholesterol but unchanged or only
modestly reduced non-HDL cholesterol levels (23). Huby et al (24), produced a hypomorphic allele
(hypomSR-BI), an SR-BI conditional knockout mouse model, by
targeting the SR-BI gene through loxP site insertion, resulting in
a decrease in SR-BI expression. After consuming an atherogenic diet
for 11 weeks, plasma total cholesterol (TC) levels were increased
2-fold compared with the control group, and hypomSR-BI-KO (liver)
and SR-BI−/− mice developed severe hypercholesterolemia
due to the accumulation of VLDL-sized particles in the circulation
(24). The degree of
atherosclerosis was more severe in hypomSR-BI mice, hypomSR-BI-KO
(liver) mice and SR-BI−/− mice compared with the control
mice, by 2.5-fold, 32-fold and 48-fold, respectively. There was no
difference in plasma lipid expression levels between hypomSR-BI-KO
(liver) mice and SR-BI−/− mice. However, compared with
SR-BI−/− mice, hypomSR-BI-KO (liver) mice had fewer
atherosclerotic lesions (24). The
aforementioned studies indicated that SR-BI expressed in
hepatocytes serves a critical role in anti-atherosclerosis, and
SR-BI expressed in other tissues or cells also perform
anti-atherosclerotic functions to a limited extent.
In addition to the selective uptake of CEs from HDL,
SR-BI expressed in the liver can also clear VLDL and Lpa. Van Eck
et al (25) demonstrated
that at 20 mins after the intravenous injection of (125)I-β-VLDL,
the liver uptake ability of SR-BI−/− mice was decreased
by 44% compared with that of wild-type mice, while VLDL in plasma
was significantly increased (45.9±8.8 vs. 6.1±2.3 µg/ml). A study
by Yang et al (26)
demonstrated that SR-BI acted as a receptor for atherosclerotic
particles containing Lpa and mediated the selective uptake of
lipids in Lpa. SR-BI−/− mice presented with the
significantly reduced clearance rate of Lpa in the plasma, while
SR-BI overexpression in the liver could significantly increase the
clearance rate. Humans carry a number of different SCARB1
gene mutations that reduce the ability of the liver to uptake CEs
in HDL and Lpa, resulting in significantly elevated plasma HDL and
Lpa expression levels (27),
although this is accompanied by higher HDL levels and the risk of
cardiovascular disease is significantly increased. The
aforementioned studies in mice and humans indicated that in
addition to mediating the reverse transport of cholesterol in
surrounding tissues to liver cells via HDL particles, generating
and excreting bile, liver SR-BI also had an anti-atherogenic role
via regulating the homeostasis of lipoproteins in the blood.
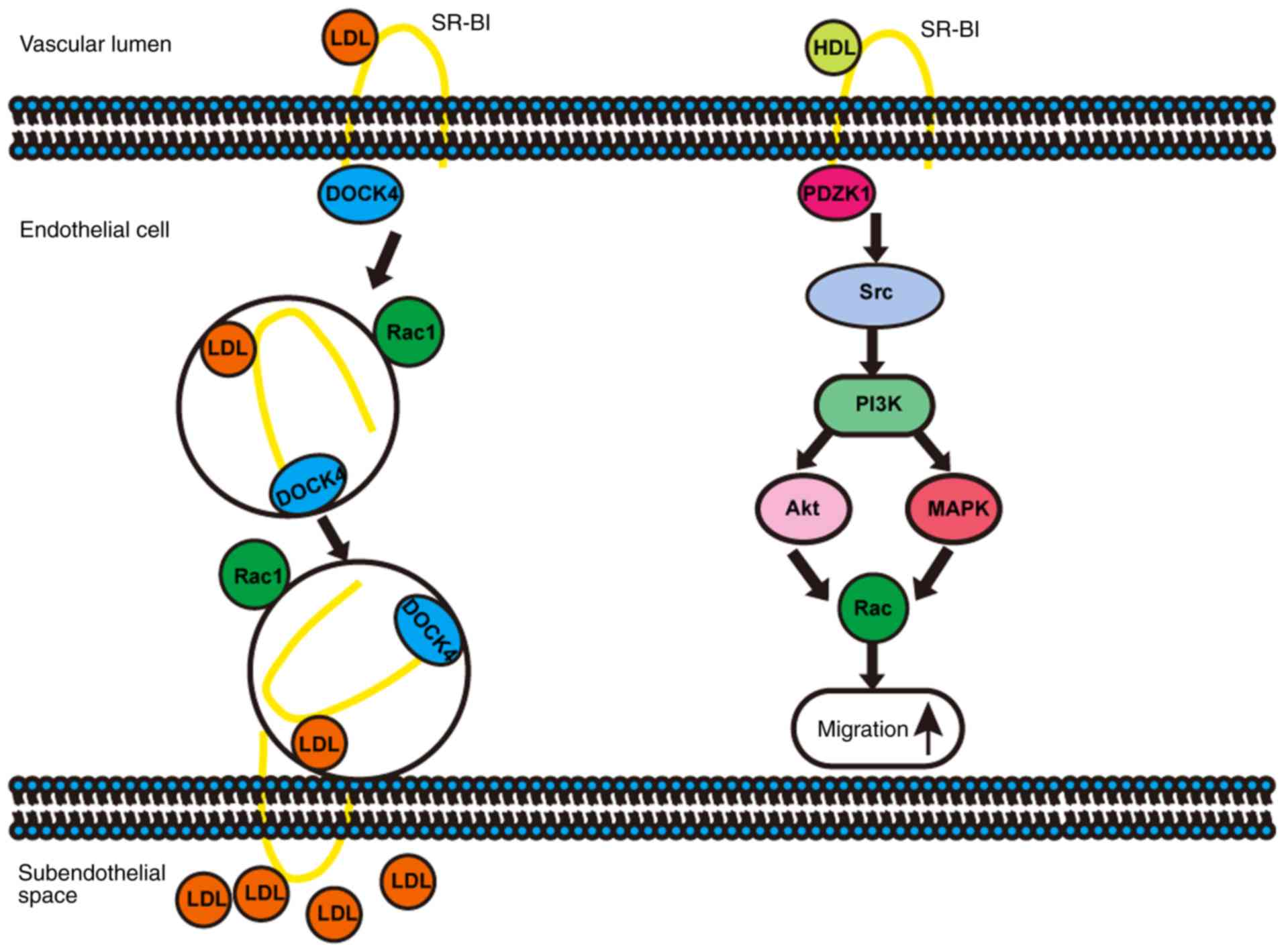
SR-BI in endothelial cells
SR-BI in ECs not only binds with HDL to activate
downstream signalling pathways (28) or transport HDL to the subintimal
region via transcytosis (29) to
exert an anti-atherosclerotic effect, but also mediates the
delivery of LDL into the artery wall via transcytosis (14,30),
thereby promoting atherosclerosis (Fig. 1).
SR-BI regulates the function of ECs under the
stimulation of HDL via activating Src kinases, phosphatidylinositol
3-kinase (PI3K), protein kinase B/mitogen-activated protein kinases
and the Rac GTP enzyme, resulting in changes in the actin
cytoskeleton, promotion of EC migration (Fig. 1), enhancement of EC repair and
regeneration and improvement of endothelial monolayer integrity
in vivo (31,32). The HDL/SR-BI interaction induced
the phosphorylation of endothelial nitric oxide synthase and the
synthesis of nitric oxide via phosphorylation of Src kinases,
protected ECs from apoptosis and promoted the growth and migration
of ECs (32). Moreover,
phosphorylation of Src kinases induced by HDL/SR-BI required the
assistance of the adaptor protein PDZK1 (3,33).
In the aforementioned study, compared with the control group,
EC-specific SR-BI-overexpressing C57BL/6N mice and
ApoE−/− mice fed a high-fat and high-cholesterol diet
displayed significantly decreased plasma levels of TC and FC, while
HDL was remarkably increased, and the severity of atherosclerosis
was decreased. However, for C57BL/6N mice and ApoE−/−
mice fed a normal chow diet, EC-specific SR-BI overexpression had
no significant effect on plasma lipids and lipoproteins (34), suggesting that SR-BI expressed on
ECs may serve an important role in the regulation of lipid and
lipoprotein homeostasis and preventing atherosclerosis.
SR-BI on ECs also serve a role in transcytosis,
transporting plasma lipoproteins to the endometrium (29,30).
Rohrer et al (29)
indicated that SR-BI could mediate the transport of HDL in the
circulation to the intima of vessels, receive cholesterol in foam
cells or macrophages and reduce cholesterol deposition beneath the
arterial endothelium, thereby exerting an anti-atherosclerotic
effect. The diameter of translocated HDL particles was decreased,
but protein moieties remained intact (29). A study by Fung et al
(35) demonstrated that SR-BI on
the brain microvascular endothelium could mediate HDL transcytosis,
and this process did not depend on the adaptor protein PDZK1.
Armstrong et al (30)
revealed that SR-BI mediated the transcytosis of LDL in ECs.
Overexpression of SR-BI enhanced the endocytosis of LDL in ECs, but
the effect was significantly reduced when an excess of HDL (a
high-affinity ligand for SR-BI) was administered or the expression
of SR-BI was depleted by small interfering RNA-mediated silencing
(30).
A recent study by Huang et al (14) demonstrated that SR-BI mediated LDL
transcytosis associated with ECs via DOCK4 to promote the
development of atherosclerosis (Fig.
1). Transcytosis of LDL particles through ECs required direct
binding to SR-BI expressed on the apical membranes of ECs, and an 8
aa (IQAYSESL) cytoplasmic motif in C-terminus of SR-BI that
recruited DOCK4 (36), which
promoted the internalisation of SR-BI and transcytosis of LDL
across EC monolayers by coupling LDL and SR-BI via Rho GTPases
Ras-associated C3 botulinum toxin substrate 1 (Rac1) activation
(14). Selective deletion of SR-BI
in ECs in male and female wild-type, ApoE−/− and
LDLR−/− mice had the protective effect against
atherosclerosis but it did not alter circulating levels of TC, HDL
or the lipoprotein profile (14).
The aforementioned results indicated that the role of SR-BI
expressed on ECs in atherosclerosis is complex, and diverse results
from different studies might be related to the metabolic state of
the body and the cholesterol load, which suggested that accurate
individualised treatment in different populations could be achieved
when formulating anti-atherosclerosis strategies targeting
SR-BI.
SR-BI in macrophages
The SR-BI protein was also expressed on macrophages,
such as Kupffer cells in the liver (37) and foam cells in atherosclerotic
plaques (38). SR-BI expressed on
macrophages has an important role in selective cholesterol uptake
(39), cholesterol efflux
(40,41), HDL-induced anti-inflammatory
responses (41,42) and the inhibition of atherosclerotic
progression (43).
Tao et al (43) demonstrated that macrophage SR-BI
bound to PS and induced Src phosphorylation, which led to the
activation of the downstream molecules PI3K and Rac1, phagocytosis
and elimination of apoptotic cells in atherosclerotic lesions,
thereby reducing necrosis, inhibiting inflammatory responses and
exerting an anti-atherosclerotic effect. Moreover, SR-BI also
mediated HDL-induced anti-inflammatory effects by inhibiting the
activation of NF-κB and the production of tumor necrosis factor α
(42).
Although SR-BI was expressed in many bone
marrow-derived cells (BMDCs), including B and T lymphocytes,
lymphocyte SR-BI did not affect the development of atherosclerosis
(44). In order to investigate the
role of macrophage SR-BI in atherosclerosis, Van Eck et al
(45) used bone marrow
transplantation technology to specifically regulate the expression
of SR-BI in leukocytes of LDLR−/− mice, and indicated
that macrophage SR-BI had a unique dual role in the process of
atherosclerosis. After feeding LDLR−/− mice with a
western-type diet for 9 or 12 weeks, the presence of SR-BI in BMDCs
reduced the development of atherosclerosis. However, after only 4
weeks in LDLR−/− mice fed a western diet and wild-type
mice fed a high-cholesterol and high-cholate diet, expression of
SR-BI in BMDCs promoted the development of small fat streak lesions
(45). In addition, SR-BI-null and
apoE-hypomorphic (SR-BI−/−/ApoE+/−) mice
transplanted with bone marrow derived from
SR-BI+/+/ApoE+/− mice and fed a high-fat
high-cholesterol and cholate-containing diet for 4 weeks revealed a
significant reduction in the degree of coronary atherosclerosis and
the incidence of myocardial infarction compared with mice
transplanted with bone marrow derived from SR-BI−/− mice
(46).
SR-BI expressed by macrophages has also been
revealed to mediate cholesterol efflux from macrophages into mature
HDL particles (40), which is an
important stage in the process of anti-atherosclerosis. Compared
with the wild-type group, the rate of cholesterol efflux from
macrophages isolated from SR-BI−/− mice to mature HDL
particles was reduced by 20%. In addition, Badeau et al
(47) indicated that SR-BI
inhibitors significantly reduced cholesterol efflux from THP-1
macrophages. A previous clinical study has demonstrated that
cholesterol outflow to HDL in macrophages from human carriers of
the SCARB1 gene P297S mutation was significantly reduced
compared with control subjects (48). In conclusion, macrophage SR-BI
exerted an anti-atherosclerotic effect by eliminating apoptotic
cells, inducing anti-inflammatory responses and promoting
cholesterol efflux.
SR-BI in platelets
Platelets are closely associated with the
development of atherosclerosis (49), and their activation and aggregation
can generate thrombus, accelerate the progression of
atherosclerotic lesions and subsequently occlude arteries, which
results in reduced blood/oxygen supply to organs or tissues,
ultimately leading to the occurrence of cardiovascular events, such
as myocardial infarction and stroke (49,50).
Therefore, inhibiting the abnormal aggregation and activation of
platelets could reduce the occurrence of cardiovascular events
(50). Valiyaveettil et al
(51) found that the SR-BI protein
was present in both resting and activated platelets, and that
oxidised HDL particles could bind to and interact with SR-BI to
inhibit platelet activation and aggregation, thereby reducing the
risk of thrombosis. In addition, Brodde et al (52) demonstrated that natural
HDL3 (the main component of HDL) bound to SR-BI on
platelets and activated the signal pathways, which inhibited
platelet activation. Furthermore, SR-BI−/− mice were
more likely to suffer deep vein thrombosis (53), and the time required for arterial
occlusion was shorter in a model of FeCl3-induced
arterial thrombosis (54). The
increase of circulating platelet and bone marrow megakaryocyte
numbers was impaired in SR-BI−/− mice when challenged
with thrombopoietin. However, proplatelets were normally produced
by megakaryocytes from normolipidemic bone marrow-specific
SR-BI−/− mice (55).
Furthermore, studies in humans indicated that carriers of the P297S
mutation of the SCARB1 gene had significantly increased FC
levels in platelets and impaired platelet function (48).
Conclusion
In conclusion, SR-BI is a multi-ligand membrane
receptor that has a high affinity for HDL and is believed to be the
physiological receptor for HDL. Expression of SR-BI in different
tissues and cell types exhibit different roles in the process of
atherosclerosis. SR-BI in hepatocytes and platelets have been
demonstrated to inhibit the development of atherosclerotic lesions
and thrombosis, and act as a protective factor for cardiovascular
diseases (16,17). However, SR-BI expressed in vascular
ECs has been indicated to promote the deposition of LDL under the
endothelium, aggravating atherosclerosis and representing a risk
factor for cardiovascular diseases. The effects of macrophage SR-BI
on the pathogenesis of atherosclerosis and cardiovascular diseases
are complex and depend on the metabolic and inflammatory status at
different stages of atherosclerosis. Therefore, the role of SR-BI
in the cardiovascular system remains ambiguous, and all the
previously reported cardioprotective and atherosclerotic effects of
SR-BI must be considered when designing therapeutic agents to treat
atherosclerosis and cardiovascular diseases.
Acknowledgements
The authors would like to thank their colleagues, Dr
Yunfei Chen and Dr Chunling Xue (Medical Science Research Center,
Chinese Academy of Medical Sciences and Peking Union Medical
College) for their helpful support and editing of this
manuscript.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant no. 81970417) and the
Fundamental Research Funds for the Central Universities (grant no.
3332019028).
Availability of data and materials
Not applicable.
Authors' contributions
BTM and JJ wrote the manuscript. XBW, RZ, SN, LN, XD
and CWL reviewed and edited the manuscript. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
SR-BI
|
scavenger receptor class B type I
|
|
HDL
|
high-density lipoprotein
|
|
EC
|
endothelial cell
|
|
LIMP-2
|
lysosomal integral membrane protein
2
|
|
CE
|
cholesterol ester
|
|
LDL
|
low-density lipoprotein
|
|
VLDL
|
very-low-density lipoprotein
|
|
Lpa
|
lipoprotein a
|
|
PS
|
phosphatidylserine
|
|
LDLR
|
LDL receptor
|
|
FC
|
free cholesterol
|
|
RCT
|
reverse cholesterol transport
|
|
DOCK4
|
dedicator of cytokinesis 4
|
|
ApoE
|
apolipoprotein E
|
|
TC
|
total cholesterol
|
|
BMDC
|
bone marrow-derived cell
|
References
|
1
|
Calvo D and Vega MA: Identification,
primary structure, and distribution of CLA-1, a novel member of the
CD36/LIMPII gene family. J Biol Chem. 268:2599–18935. 1993.
|
|
2
|
Acton S, Rigotti A, Landschulz KT, Xu S,
Hobbs HH and Krieger M: Identification of scavenger receptor SR-BI
as a high density lipoprotein receptor. Science. 271:518–520. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Shen WJ, Azhar S and Kraemer FB: SR-B1: A
unique multifunctional receptor for cholesterol influx and efflux.
Annu Rev Physiol. 80:95–116. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Shen WJ, Asthana S, Kraemer FB and Azhar
S: Scavenger receptor B type 1: Expression, molecular regulation,
and cholesterol transport function. J Lipid Res. 59:1114–1131.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Shen WJ, Hu J, Hu Z, Kraemer FB and Azhar
S: Scavenger receptor class B type I (SR-BI): A versatile receptor
with multiple functions and actions. Metabolism. 63:875–886. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hoekstra M: SR-BI as target in
atherosclerosis and cardiovascular disease-A comprehensive
appraisal of the cellular functions of SR-BI in physiology and
disease. Atherosclerosis. 258:153–161. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kraemer FB, Shen WJ, Harada K, Patel S,
Osuga J, Ishibashi S and Azhar S: Hormone-sensitive lipase is
required for high-density lipoprotein cholesteryl ester-supported
adrenal steroidogenesis. Mol Endocrinol. 18:549–557. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Prospective Studies Collaboration,
Lewington S, Whitlock G, Clarke R, Sherliker P, Emberson J, Halsey
J, Qizilbash N, Peto R and Collins R: Blood cholesterol and
vascular mortality by age, sex, and blood pressure: A meta-analysis
of individual data from 61 prospective studies with 55,000 vascular
deaths. Lancet. 370:1829–1839. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Phillips MC: Molecular mechanisms of
cellular cholesterol efflux. J Biol Chem. 289:24020–24029. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ji Y, Wang N, Ramakrishnan R, Sehayek E,
Huszar D, Breslow JL and Tall AR: Hepatic scavenger receptor BI
promotes rapid clearance of high density lipoprotein free
cholesterol and its transport into bile. J Biol Chem.
274:33398–33402. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Brundert M, Ewert A, Heeren J, Behrendt B,
Ramakrishnan R, Greten H, Merkel M and Rinninger F: Scavenger
receptor class B type I mediates the selective uptake of
high-density lipoprotein-associated cholesteryl ester by the liver
in mice. Arterioscler Thromb Vasc Biol. 25:143–148. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Acton SL, Scherer PE, Lodish HF and
Krieger M: Expression cloning of SR-BI, a CD36-related class B
scavenger receptor. J Biol Chem. 269:21003–21009. 1994.PubMed/NCBI
|
|
13
|
Trigatti BL: SR-B1 and PDZK1: Partners in
HDL regulation. Curr Opin Lipidol. 28:201–208. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Huang L, Chambliss KL, Gao X, Yuhanna IS,
Behling-Kelly E, Bergaya S, Ahmed M, Michaely P, Luby-Phelps K,
Darehshouri A, et al: SR-B1 drives endothelial cell LDL
transcytosis via DOCK4 to promote atherosclerosis. Nature.
569:565–569. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Neculai D, Schwake M, Ravichandran M,
Zunke F, Collins RF, Peters J, Neculai M, Plumb J, Loppnau P,
Pizarro JC, et al: Structure of LIMP-2 provides functional insights
with implications for SR-BI and CD36. Nature. 504:172–176. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Linton MF, Tao H, Linton EF and Yancey PG:
SR-BI: A multifunctional receptor in cholesterol homeostasis and
atherosclerosis. Trends Endocrinol Metab. 28:461–472. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Sahebi R, Hassanian SM, Ghayour-Mobarhan
M, Farrokhi E, Rezayi M, Samadi S, Bahramian S, Ferns GA and Avan
A: Scavenger receptor Class B type I as a potential risk
stratification biomarker and therapeutic target in cardiovascular
disease. J Cell Physiol. 234:16925–16932. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Van Eck M, Twisk J, Hoekstra M, Van Rij
BT, Van der Lans CA, Bos IS, Kruijt JK, Kuipers F and Van Berkel
TJ: Differential effects of scavenger receptor BI deficiency on
lipid metabolism in cells of the arterial wall and in the liver. J
Biol Chem. 278:23699–23705. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Braun A, Trigatti BL, Post MJ, Sato K,
Simons M, Edelberg JM, Rosenberg RD, Schrenzel M and Krieger M:
Loss of SR-BI expression leads to the early onset of occlusive
atherosclerotic coronary artery disease, spontaneous myocardial
infarctions, severe cardiac dysfunction, and premature death in
apolipoprotein E-deficient mice. Circ Res. 90:270–276. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Braun A, Zhang S, Miettinen HE, Ebrahim S,
Holm TM, Vasile E, Post MJ, Yoerger DM, Picard MH, Krieger JL, et
al: Probucol prevents early coronary heart disease and death in the
high-density lipoprotein receptor SR-BI/apolipoprotein E double
knockout mouse. Proc Natl Acad Sci USA. 100:7283–7288. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Fuller M, Dadoo O, Serkis V, Abutouk D,
MacDonald M, Dhingani N, Macri J, Igdoura SA and Trigatti BL: The
effects of diet on occlusive coronary artery atherosclerosis and
myocardial infarction in scavenger receptor class B, type
1/low-density lipoprotein receptor double knockout mice.
Arterioscler Thromb Vasc Biol. 34:2394–2403. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Leiva A, Verdejo H, Benitez ML, Martinez
A, Busso D and Rigotti A: Mechanisms regulating hepatic SR-BI
expression and their impact on HDL metabolism. Atherosclerosis.
217:299–307. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kozarsky KF, Donahee MH, Glick JM, Krieger
M and Rader DJ: Gene transfer and hepatic overexpression of the HDL
receptor SR-BI reduces atherosclerosis in the cholesterol-fed LDL
receptor-deficient mouse. Arterioscler Thromb Vasc Biol.
20:721–727. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Huby T, Doucet C, Dachet C, Ouzilleau B,
Ueda Y, Afzal V, Rubin E, Chapman MJ and Lesnik P: Knockdown
expression and hepatic deficiency reveal an atheroprotective role
for SR-BI in liver and peripheral tissues. J Clin Invest.
116:2767–2776. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Van Eck M, Hoekstra M, Out R, Bos IS,
Kruijt JK, Hildebrand RB and Van Berkel TJ: Scavenger receptor BI
facilitates the metabolism of VLDL lipoproteins in vivo. J Lipid
Res. 49:136–146. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yang XP, Amar MJ, Vaisman B, Bocharov AV,
Vishnyakova TG, Freeman LA, Kurlander RJ, Patterson AP, Becker LC
and Remaley AT: Scavenger receptor-BI is a receptor for
lipoprotein(a). J Lipid Res. 54:2450–2457. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yang X, Sethi A, Yanek LR, Knapper C,
Nordestgaard BG, Tybjærg-Hansen A, Becker DM, Mathias RA, Remaley
AT and Becker LC: SCARB1 gene variants are associated with the
phenotype of combined high high-density lipoprotein cholesterol and
high lipoprotein (a). Circ Cardiovasc Genet. 9:408–418. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Saddar S, Mineo C and Shaul PW: Signaling
by the high-affinity HDL receptor scavenger receptor B type I.
Arterioscler Thromb Vasc Biol. 30:144–150. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Rohrer L, Ohnsorg PM, Lehner M, Landolt F,
Rinninger F and von Eckardstein A: High-density lipoprotein
transport through aortic endothelial cells involves scavenger
receptor BI and ATP-binding cassette transporter G1. Circ Res.
104:1142–1150. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Armstrong SM, Sugiyama MG, Fung KY, Gao Y,
Wang C, Levy AS, Azizi P, Roufaiel M, Zhu SN, Neculai D, et al: A
novel assay uncovers an unexpected role for SR-BI in LDL
transcytosis. Cardiovasc Res. 108:268–277. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Seetharam D, Mineo C, Gormley AK, Gibson
LL, Vongpatanasin W, Chambliss KL, Hahner LD, Cummings ML, Kitchens
RL, Marcel YL, et al: High-density lipoprotein promotes endothelial
cell migration and reendothelialization via scavenger receptor-B
type I. Circ Res. 98:63–72. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Mineo C and Shaul PW: Role of high-density
lipoprotein and scavenger receptor B type I in the promotion of
endothelial repair. Trends Cardiovasc Med. 17:156–161. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhu W, Saddar S, Seetharam D, Chambliss
KL, Longoria C, Silver DL, Yuhanna IS, Shaul PW and Mineo C: The
scavenger receptor class B type I adaptor protein PDZK1 maintains
endothelial monolayer integrity. Circ Res. 102:480–487. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Vaisman BL, Vishnyakova TG, Freeman LA,
Amar MJ, Demosky SJ, Liu C, Stonik JA, Sampson ML, Pryor M,
Bocharov AV, et al: Endothelial expression of scavenger receptor
class B, type I protects against development of atherosclerosis in
mice. Biomed Res Int. 2015:6071202015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Fung KY, Wang C, Nyegaard S, Heit B, Fairn
GD and Lee WL: SR-BI mediated transcytosis of HDL in brain
microvascular endothelial cells is independent of caveolin,
clathrin, and PDZK1. Front Physiol. 8:8412017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Gadea G and Blangy A: Dock-family exchange
factors in cell migration and disease. Eur J Cell Biol. 93:466–477.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Fluiter K, van der Westhuijzen DR and van
Berkel TJ: In vivo regulation of scavenger receptor BI and the
selective uptake of high density lipoprotein cholesteryl esters in
rat liver parenchymal and kupffer cells. J Biol Chem.
273:8434–8438. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Yancey PG, de la Llera-Moya M, Swarnakar
S, Monzo P, Klein SM, Connelly MA, Johnson WJ, Williams DL and
Rothblat GH: High density lipoprotein phospholipid composition is a
major determinant of the bi-directional flux and net movement of
cellular free cholesterol mediated by scavenger receptor BI. J Biol
Chem. 275:36596–36604. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Matveev S, van der Westhuyzen DR and Smart
EJ: Co-expression of scavenger receptor-BI and caveolin-1 is
associated with enhanced selective cholesteryl ester uptake in
THP-1 macrophages. J Lipid Res. 40:1647–1654. 1999.PubMed/NCBI
|
|
40
|
Ji Y, Jian B, Wang N, Sun Y, Moya ML,
Phillips MC, Rothblat GH, Swaney JB and Tall AR: Scavenger receptor
BI promotes high density lipoprotein-mediated cellular cholesterol
efflux. J Biol Chem. 272:20982–20985. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Mineo C: Lipoprotein receptor signalling
in atherosclerosis. Cardiovasc Res. 116:1254–1274. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Song GJ, Kim SM, Park KH, Kim J, Choi I
and Cho KH: SR-BI mediates high density lipoprotein (HDL)-induced
anti-inflammatory effect in macrophages. Biochem Biophys Res
Commun. 457:112–118. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Tao H, Yancey PG, Babaev VR, Blakemore JL,
Zhang Y, Ding L, Fazio S and Linton MF: Macrophage SR-BI mediates
efferocytosis via Src/PI3K/Rac1 signaling and reduces
atherosclerotic lesion necrosis. J Lipid Res. 56:1449–1460. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Karackattu SL, Picard MH and Krieger M:
Lymphocytes are not required for the rapid onset of coronary heart
disease in scavenger receptor class B type I/apolipoprotein E
double knockout mice. Arterioscler Thromb Vasc Biol. 25:803–808.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Van Eck M, Bos IS, Hildebrand RB, Van Rij
BT and Van Berkel TJ: Dual role for scavenger receptor class B,
type I on bone marrow-derived cells in atherosclerotic lesion
development. Am J Pathol. 165:785–794. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Pei Y, Chen X, Aboutouk D, Fuller MT,
Dadoo O, Yu P, White EJ, Igdoura SA and Trigatti BL: SR-BI in bone
marrow derived cells protects mice from diet induced coronary
artery atherosclerosis and myocardial infarction. PLoS One.
8:e724922013. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Badeau RM, Metso J, Wähälä K, Tikkanen MJ
and Jauhiainen M: Human macrophage cholesterol efflux potential is
enhanced by HDL-associated 17beta-estradiol fatty acyl esters. J
Steroid Biochem Mol Biol. 116:44–49. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Vergeer M, Korporaal SJ, Franssen R, Meurs
I, Out R, Hovingh GK, Hoekstra M, Sierts JA, Dallinga-Thie GM,
Motazacker MM, et al: Genetic variant of the scavenger receptor BI
in humans. N Engl J Med. 364:136–145. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Lindemann S, Krämer B, Daub K, Stellos K
and Gawaz M: Molecular pathways used by platelets to initiate and
accelerate atherogenesis. Curr Opin Lipidol. 18:566–573. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Olas B: Biochemistry of blood platelet
activation and the beneficial role of plant oils in cardiovascular
diseases. Adv Clin Chem. 95:219–243. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Valiyaveettil M, Kar N, Ashraf MZ, Byzova
TV, Febbraio M and Podrez EA: Oxidized high-density lipoprotein
inhibits platelet activation and aggregation via scavenger receptor
BI. Blood. 111:1962–1971. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Brodde MF, Korporaal SJ, Herminghaus G,
Fobker M, Van Berkel TJ, Tietge UJ, Robenek H, Van Eck M, Kehrel BE
and Nofer JR: Native high-density lipoproteins inhibit platelet
activation via scavenger receptor BI: Role of negatively charged
phospholipids. Atherosclerosis. 215:374–382. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Brill A, Yesilaltay A, De Meyer SF,
Kisucka J, Fuchs TA, Kocher O, Krieger M and Wagner DD:
Extrahepatic high-density lipoprotein receptor SR-BI and apoA-I
protect against deep vein thrombosis in mice. Arterioscler Thromb
Vasc Biol. 32:1841–1847. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Korporaal SJ, Meurs I, Hauer AD,
Hildebrand RB, Hoekstra M, Cate HT, Praticò D, Akkerman JW, Van
Berkel TJ, Kuiper J and Van Eck M: Deletion of the high-density
lipoprotein receptor scavenger receptor BI in mice modulates
thrombosis susceptibility and indirectly affects platelet function
by elevation of plasma free cholesterol. Arterioscler Thromb Vasc
Biol. 31:34–42. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Ouweneel AB, Hoekstra M, van der Wel EJ,
Schaftenaar FH, Snip OSC, Hassan J, Korporaal SJA and Van Eck M:
Hypercholesterolemia impairs megakaryopoiesis and platelet
production in scavenger receptor BI knockout mice. Atherosclerosis.
282:176–182. 2019. View Article : Google Scholar : PubMed/NCBI
|