Introduction
Inflammatory bowel disease is a chronic, repetitive
and non-specific gastrointestinal inflammatory disease comprising
two conditions: Ulcerative colitis (UC) and Crohn's disease
(1). As a chronic intestinal
disease, UC produces inflammatory reactions and immune response at
the colonic mucosa (2),
accompanied by weight loss, diarrhea and bloody stools (3). Long-term severe colitis is known to
cause colorectal cancer (4).
The pathogenesis of UC involves tissue damage in the
colon and deterioration of the digestive and absorptive functions
(5). Inflammation of the colon is
due to an imbalance between the expression of pro-inflammatory and
anti-inflammatory factors, and the migration of neutrophils to the
damaged site, leading to the accumulation of inflammatory factors
(6). Interleukin (IL)-6, closely
associated with the differentiation and maturation of B cells,
promotes the function of pro-inflammatory cells and inhibits the
regulation of immune-related cells including T cells (7), both of which increase the levels of
pro-inflammatory factors including IL-1 and IL-17 (8). Therefore, blocking the expression of
pro-inflammatory cytokines may be a method for the potential
treatment of UC (9). NF-κB
inhibitor α is involved in the regulation of the development of
early inflammation (10). The
inflammatory response leads to the aggregation of peroxides and the
imbalance of oxidation and antioxidation in the colon, particularly
the over-accumulation of reactive oxygen species (ROS), causes
oxidative damage (11).
Antioxidant enzymes, including superoxide dismutase (SOD), are the
first line of defense against oxidative stress, helping to remove
peroxides from the body, but generally tend to promote oxidation in
UC (12). During this process, the
nuclear factor erythroid-2-related factor 2 (Nrf2) recognizes the
transcription-enhancing sequence associated with antioxidation and
repairs bodily damage by enhancing the expression of antioxidant
enzymes (13).
Dextran sulfate sodium (DSS) affects the normal
replication of DNA in cells, causes the aggregation of inflammatory
factors and destruction of the intestinal microenvironment via the
overproduction of ROS and eventually leads to the onset of UC
(14). DSS was used to induce the
occurrence of UC in the present study. Sulfasalazine (SASP), which
reduces the infiltration of inflammatory monocytes, has been used
to treat UC in clinics (15) and
was applied as a positive control agent in this study.
Gloeostereum incarnatum (GI) is an edible and
medicinal fungus, commonly cultivated in north-central China and
Hokkaido, Japan (16). The
anti-inflammatory activities of GI extracts were found to reduce
nitric oxide (NO) production in lipopolysaccharide (LPS)-induced
RAW264.7 cells and delay melanin synthesis in B16/F10 cells
(17). In our previous study, GI
was confirmed to show immunomodulatory function in a
cyclophosphamide monohydrate-induced mouse model by increasing
immune-related factors including immunoglobulin (Ig) (7). Due to its anti-microbial activities,
GI shows beneficial effects on gastrointestinal diseases including
gastric ulcer and enteritis (18).
However, thus far, there has been no systematic research on the
protective effect of GI against UC.
In the present study, based on the detection of the
main components of GI, its anti-inflammation and antioxidative
properties were successfully confirmed in C57BL/6 mice with UC
induced via oral administration of 3.5% DSS. The data provide
experimental evidence for the applicability of GI for protection
against UC.
Materials and methods
Detection of GI components
GI was purchased from TengHui Agriculture,
identified by Professor Yu Li (Jilin Agricultural University,
China) and pulverized by a pulverizer (XL-06A, Guangzhou Xulang
Machinery Equipment Co., Ltd.).
Detection of main components
In GI powder, the contents of total sugar, reducing
sugar, mannitol, total ash, total flavonoids, total triterpenoids,
crude fat and crude fiber were detected using phenol-sulfuric acid
assay (19), 3,5-dinitrosalicylic
acid colorimetry (20), iodometry
(21), regression analysis
(22), the alumina colorimetric
method (23), vanillin-glacial
acetic acid-perchloric acid colorimetry (24), petroleum ether extraction (25) and the Ankom filter bag method
(26), respectively.
Detection of minerals
Inductively coupled plasma optical emission
spectrometry was used to detect the contents of mercury (Hg), lead
(Pb), selenium (Se), arsenic (As), cadmium (Cd), zinc (Zn), iron
(Fe), manganese (Mn), chromium (Cr), calcium (Ca), copper (Cu),
sodium (Na) and potassium (K) in GI (27).
Detection of fatty acids
An oil sample of GI was extracted using a chloroform
and methanol solution (2:1) and treated by alkaline hydrolysis to
prepare the corresponding fatty acid methyl esters. Fatty acids
were detected using gas chromatography-mass spectrometry (QP2010,
Shimadzu Corporation) based on retention times (28).
Detection of amino acids
The protein in GI was hydrolyzed to single amino
acid residues by hydrochloric acid hydrolysis and the contents of
different amino acids were analyzed using an automatic amino acid
analyzer (L-8900, Hitachi, Ltd.) (29).
Establishment of UC model mice and
agent administration
A total of 48 male C57BL/6 mice [22–25 g; 8–10 weeks
old; SPF grade, SCXK (LIAO) 2015-001], purchased from Liaoning
Changsheng Biotechnology Co., Ltd., were housed in an exhaust
ventilation cage system with a temperature of 22±2°C and humidity
of 50±10%, with ad libitum access to food and water and a
12-h light/dark cycle. All animal experiments were approved by the
Experimental Animal Ethics Committee of Jilin University [approval
no. SYXK(JI)2014-0013]. The study was conducted using the
Laboratory Guidelines for Animal Care (30,31)
and the Guide for the Care and Use of Laboratory Animals (eighth
edition) (32).
After 7 days of adaptive feeding, all mice with the
exception of the control mice drank freely available clean water
containing 3.5% DSS (S14048, Shanghai Yuanye Biological Technology
Co., Ltd.) every day for 7 days. From the 8th to the 28th day, the
mice drank 3.5% DSS on 1 day out of every 3 days. At the 7th day,
the mice were randomly divided into 6 groups: The control group
(n=8) that orally received double distilled (D.D.) water, the model
group (n=8) that orally received D.D. water, the positive control
group (n=8) that orally received 0.6 g/kg of SASP dissolved in D.D.
water and the GI-treated groups that orally received 1.0 g/kg
(n=8), 2.0 g/kg (n=8) and 4.0 g/kg (n=8) of GI suspended in D.D.
water for the remaining 21 days. The doses of GI and SASP were
selected based on previous studies (7,33)
and our preliminary experiments (data not shown). On the 28th day,
blood samples were collected from the orbital venous plexus of the
mice and the mice were then euthanized by intraperitoneal injection
of 100 mg/kg sodium pentobarbital (Xiya Reagent Co., Ltd.);
mortality was characterized by cessation of heartbeat (34). Tissues including the colon, liver,
spleen and kidney of each mouse were collected for further
detection. The length of the colon and the organ index of the
liver, spleen and kidney were calculated. The following formula was
used to calculate the organ index (35): Organ index (%)=mean organ
weight/mean body weight ×100.
During the entire experimental period, the activity,
physiological status and body weight were monitored daily. The
status of the UC mice was scored using the disease activity index
(DAI) according to the degree of weight loss, fecal viscosity and
bleeding (36).
Detection of biochemical factors in
the serum and colon of UC mice
Colon tissue was homogenized in D.D. water contained
1.0% 50-mM phenylmethanesulfonyl fluoride (PMSF; Sigma-Aldrich;
Merck KGaA) and 1.0% protease inhibitor cocktail (Sigma-Aldrich;
Merck KGaA) using a homogenizer [S-18KS, Leopard scientific
instruments (Beijing) Co., Ltd.]. After centrifuging the homogenate
at 1,000 × g for 10 min at 4°C, the supernatant was collected, and
the protein content was determined using a BCA protein assay kit
(EMD Millipore). The levels of IL-1β (E20180501A), IL-2
(E20180501A), IL-6 (E20180501A), IL-12 (E20180501A), tumor necrosis
factor (TNF)-α (E20180501A), TNF-β (E20180501A), interferon (IFN)-α
(E20180501A) and IFN-γ (E20180501A) in the colon and IgA
(E20180401A), IgM (E20180401A), IgG (E20170601A), SOD (E20180401A),
catalase (CAT) (E20180401A), ROS (E20180401A) and NO (E20180401A)
in the colon and serum were measured using ELISA kits purchased
from Shanghai Yuanye Biological Technology Co., Ltd.
Histopathological examination of
organs in the UC mice
The tissues were fixed in 4% paraformaldehyde at
37°C for 24 h and dehydrated in an ascending series of ethanol,
embedded in paraffin and then cut into 5–8-µm-thick sections by a
microtome (Leica Microsystems GmbH). The samples were dewaxed with
xylene at 37°C and rehydrated in a descending series of ethanol.
The samples were stained with 0.45% hematoxylin for 10 min and 0.5%
eosin for 3 min, both at 37°C, and then observed and images
captured using a light microscope (magnification, ×400; Olympus
Corporation).
Western blot analysis of colon
tissue
The proteins were extracted from colon tissue as
detailed in our previous study (37). Total protein was quantified using a
bicinchoninic acid protein assay kit (EMD Millipore) and 40 µg
protein/well was separated by 12% SDS-PAGE and transferred to
polyvinylidene fluoride membranes (EMD Millipore) using the same
method as that used in our previous study (37). The membranes were blocked with 5%
BSA (Sigma-Aldrich; Merck KGaA) at 4°C for 4 h and then incubated
overnight with primary antibodies against nuclear factor erythroid
2-related factor 2 (Nrf2; 1:2,000; cat. no. ab89443), CAT (1:2,000;
cat. no. ab16731), heme oxygenase-1 (HO-1; 1:2,000; cat. no.
ab137749), SOD-1 (1:2,000; cat. no. ab16831), SOD-2 (1:5,000; cat.
no. ab13533), phosphorylated (p)-NF-κB (1:2,000; cat. no. ab86299),
total (T)-NF-κB (1:1,000; cat. no. ab7970), p-inhibitor of NF-κB
kinase α+β (IKKα+β; 1:1,000; cat. no. ab195907), T-IKKα+β (1:1,000;
cat. no. ab178870), p-inhibitor of NF-κB (IκBα; 1:500; cat. no.
ab12135), T-IκBα (1:1,000; cat. no. ab32518) and
glyceraldehyde-3-phosphate dehydrogenase (GAPDH; 1:1,000; cat. no.
ab8245; all from Abcam) at 4°C. The antibodies were diluted
according to the ratio given in the manufacturer's protocols.
Following washing with a Tris-buffered saline solution containing
0.05% Tween 20, the membranes were incubated with goat anti-rabbit
(1:5,000; cat. no. IH-0011) or goat anti-mouse secondary antibody
(1:5,000; cat. no. IH-0031; both Beijing Dingguo Changsheng
Biotechnology Co Ltd.) at 4°C for 4 h. Electrochemiluminescence
detection kits (EMD Millipore) and a Gel Imaging System (UVP, LLC)
were used to detect changes in the protein content. The optical
densities of bands were measured using ImageJ 1.48u software
(National Institutes of Health).
Statistical analysis
Data are expressed as the mean ± standard error of
the mean. One-way analysis of variance (ANOVA) and subsequent
post-hoc multiple comparisons (Tukey's test) were performed using
SPSS 16.0 software (SPSS, Inc.). P<0.05 was considered to
indicate a statistically significant difference.
Results
Evaluation of GI components
GI contained 16.6% of total sugar, 16.8% of crude
fiber, 2.1% of mannitol, 13.9% of total ash, 6.3% of reducing
sugar, 0.394% of total flavonoids, 5.328×10−3% of total
triterpenes and 2.8% of crude fat (Table I). The presence of 35 fatty acids
was detected, with linoleic acid (1.626%), oleic acid (0.236%) and
hexadecenoic acid (0.229%) being the most abundant; by contrast,
the fatty acids caprylic acid, undecanoic acid, translinoleic acid
and docosadienoic acid were not detected (Table I). GI contained 17 amino acids and
11 minerals, with no harmful elements (Table I).
 | Table I.Composition of Gloeostereum
incarnatum. |
Table I.
Composition of Gloeostereum
incarnatum.
| A, Main
components |
|---|
|
|---|
| Compounds | Contents, % |
|---|
| Total sugar | 16.6 |
| Total ash | 13.9 |
| Total triterpenes
(×10−3) |
5.328 |
| Crude fiber | 16.8 |
| Reducing sugar | 6.3 |
| Crude fat | 2.8 |
| Mannitol | 2.1 |
| Total
flavonoids |
0.394 |
|
| B, Fatty
acids |
|
|
Compounds | Contents,
% |
|
| Caprylic acid
(C8:0) | ND |
| Capric acid
(C10:0) | ND |
| Undecanoic acid
(C11:0) | ND |
| Lauric acid
(C12:0) | 0.015 |
| Tridecanoic acid
(C13:0) | ND |
| Myristic acid
(C14:0) | 0.003 |
| Myristoleic acid
(C14:1) | ND |
| Pentadecanoic acid
(C15:0) | 0.028 |
| Pentadecenoic acid
(C15:1) | ND |
| Hexadecanoic acid
(C16:0) | 0.229 |
| Palmitoleic acid
(C16:1) | 0.021 |
| Heptadecanoic acid
(C17:0) | 0.005 |
| Margaroleic acid
(C17:1) | 0.004 |
| Stearic acid
(C18:0) | 0.014 |
| Elaidic acid
(C18:1n9t) | ND |
| Oleic acid
(C18:1n9c) | 0.236 |
| Translinoleic acid
(C18:2n6t) | ND |
| Linolic acid
(C18:2n6c) | 1.626 |
| α-Linolenic acid
(C18:3n3) | ND |
| γ-Linolenic acid
(C18:3n6) | ND |
| Arachidic acid
(C20:0) | ND |
| Cis-11-Eicosenoic
acid C20:1n9) | 0.067 |
| Docosanoic acid
(C22:0) | ND |
| Erucic acid
(C22:1n9) | ND |
| Docosadienoic acid
(C22:2n6) | ND |
| Docosahexaenoic
acid (C22:6n3) | ND |
| Tricosanoic acid
(C23:0) | 0.005 |
| Tetracosanoic acid
(C24:0) | 0.008 |
| Nervonic acid
(C24:1n9) | 0.016 |
|
Dihomo-gamma-linolenic acid (C20:3n6) | ND |
| Arachidonic acid
(C20:4n6) | ND |
| Eicosapentaenoic
acid (C20:5n3) | ND |
| Heneicosanoic acid
(C21:0) | ND |
| Eicosadienoic acid
(C20:2) | 0.005 |
| Eicosatrienoic acid
(C20:3n3) | ND |
|
| C, Amino
acids |
|
|
Compounds | Contents,
% |
|
| Aspartic acid | 0.728 |
| L-Threonine | 0.405 |
| Serine | 0.471 |
| Glutamic acid | 1.012 |
| Glycine | 0.400 |
| Alanine | 0.463 |
| Cystine | 0.266 |
| Valine | 0.406 |
| Methionine | 0.096 |
| Isoleucine | 0.343 |
| Leucine | 0.616 |
| Tyrosine | 0.248 |
| Phenylalanine | 0.335 |
| Lysine | 0.221 |
| Histidine | 0.188 |
| Arginine | 0.510 |
| Proline | 0.377 |
|
| D,
Minerals |
|
|
Compounds | Contents,
mg/kg |
|
| Mercury | ND |
| Lead | 0.11493 |
| Selenium | 0.03450 |
| Arsenic | 0.12343 |
| Cadmium | ND |
| Zinc | 44.64 |
| Iron | 105.9 |
| Manganese | 6.135 |
| Chromium | 3.15357 |
| Calcium | 601.5 |
| Copper | 9.230 |
| Sodium | 345.8 |
| Potassium | 48,605 |
Protective effect of GI against
UC
In the model mice with UC, DSS free-drinking caused
marked body weight loss (P<0.05; Table II) and the worsening of the DAI
(P<0.001; Fig. 1A), both of
which were reversed after 21-day GI administration (P<0.05;
Table II and Fig. 1A). Compared with the model mice, GI
treatment in the UC mice prevented the increase of the spleen and
liver indexes (P<0.05; Table
II) and increased the kidney indexes (P<0.05; Table II); however, SASP demonstrated no
significant effects on the liver indexes (Table II). Inflammatory cell infiltration
and ulceration of the colon can lead to the shortening of the
colon, which was noted in the model mice (Fig. 1B). SASP and GI notably prevented
this pathological reduction of the colon length (Fig. 1B). In the colons of the model mice
with UC, a large number of exfoliated lymphocytes were identified;
meanwhile, the goblet cells were reduced and the crypt cells were
damaged, which were all significantly alleviated following SASP and
GI treatment (Fig. 1C and D).
Furthermore, inflammatory cell infiltration in the spleen (Fig. 1E), interstitial edema in the liver
(Fig. 1F) and eosinophilic renal
tubular epithelial cells in the kidney (Fig. 1G) were all observed in the
DSS-only-treated mice (the model mice). These pathological
alterations were all alleviated following SASP and GI
administration (Fig. 1E-G).
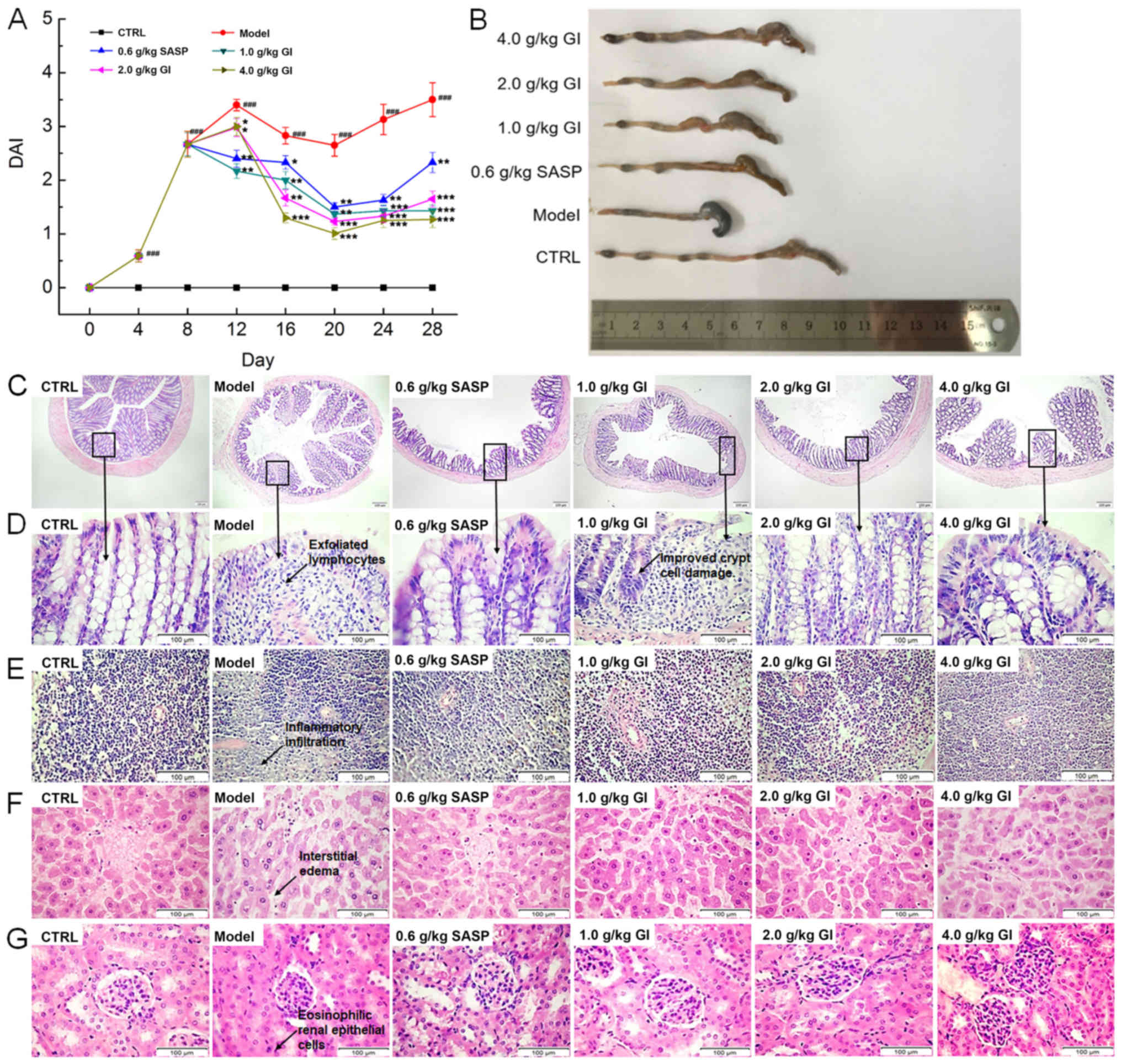 | Figure 1.GI regulates the physiology and
pathology of UC mice. The mice were continuously exposed to DSS
(3.5% DSS dissolved in D.D. water) for 28 days, SASP (0.6 g/kg of
SASP dissolved in D.D. water) and GI (1.0, 2.0 and 4.0 g/kg of GI
suspended in D.D. water) were administered from the 7th day. (A) GI
reduced the DAI index of UC mice and (B) ameliorated the shortening
of colon length. Hematoxylin and eosin staining of (C) colon (scale
bar, 100 µm; magnification, ×40), (D) colon, (E) spleen, (F) liver
and (G) kidney tissues (scale bar, 100 µm; magnification, ×400)
from C57BL/6 mice. ###P<0.001 vs. control mice;
*P<0.05, **P<0.01 and ***P<0.001 vs. DSS-induced UC mice.
GI, Gloeostereum incarnatum; UC, ulcerative colitis; DSS,
dextran sulfate sodium; D.D., double distilled; SASP,
sulfasalazine; DAI, disease activity index. |
| Table II.Effect of GI on body weight and organ
indexes. |
Table II.
Effect of GI on body weight and organ
indexes.
|
|
| 3.5% dextran
sulfate sodium |
|---|
|
|
|
|
|---|
|
|
|
|
| Gl, g/kg |
|---|
|
|
|
|
|
|
|---|
| Variable | Control | Model | Sulfasalazine, 0.6
g/kg | 1.0 | 2.0 | 4.0 |
|---|
| Body weight, g |
|
|
|
|
|
|
| Day
1 | 22.8±0.2 | 22.6±0.4 | 22.3±0.4 | 22.5±0.3 | 22.6±0.4 | 22.6±0.3 |
| Day
7 | 22.4±0.3 |
20.3±0.3a | 20.2±0.3 | 20.5±0.4 | 20.3±0.3 | 20.5±0.3 |
| Day 14 | 24.4±0.3 |
20.6±0.8b | 21.2±0.5 | 21.4±0.7 | 21.6±0.3 | 21.2±0.7 |
| Day
21 | 25.4±0.4 |
20.8±0.8b | 22.1±0.4 | 22.3±0.5 |
22.9±0.3d | 22.0±0.5 |
| Day
28 | 24.3±0.4 |
21.2±0.5b | 22.1±0.3 |
22.7±0.5d |
23.4±0.3d | 21.5±0.3 |
| Organ index, % |
|
|
|
|
|
|
|
Spleen | 0.25±0.05 |
0.56±0.10c |
0.45±0.05d | 0.49±0.14 |
0.44±0.10d |
0.38±0.11e |
|
Liver | 3.72±0.31 |
4.34±0.24a | 4.32±0.13 |
3.95±0.44d |
3.79±0.40d |
3.58±0.27e |
|
Kidney | 1.16±0.05 |
1.03±0.05a |
1.13±0.06d | 1.10±0.10 |
1.11±0.05d | 1.01±0.07 |
Anti-inflammatory effects of GI
Inflammation of the mucosal surface of the colon
leads to the development of UC (38). The free-drinking of DSS in the
present study increased the levels of pro-inflammatory factors
including IL-1β, IL-2, IL-6, IL-12, TNF-α, TNF-β, IFN-α and IFN-γ
(P<0.05; Table III). GI and
SASP demonstrated strong anti-inflammatory effects, as evidenced by
the regulation of inflammatory cytokines (P<0.05; Table III). Compared with the model
mice, 21-day GI administration in the UC mice resulted in >19.3%
(P<0.05), >31.7% (P<0.05), >31.5% (P<0.05),
>30.8% (P<0.05), >23.4% (P<0.05), >30.2%
(P<0.05), >28.1% (P<0.05) and >33.5% (P<0.05)
reduction in IL-1β, IL-2, IL-6, IL-12, TNF-α, TNF-β, IFN-α and
IFN-γ levels, respectively, in the colon (Table III).
| Table III.Effect of GI on inflammatory factors
of colon tissues of mice with ulcerative colitis. |
Table III.
Effect of GI on inflammatory factors
of colon tissues of mice with ulcerative colitis.
|
|
| 3.5% dextran
sulfate sodium |
|---|
|
|
|
|
|---|
|
|
|
|
| GI, g/kg |
|---|
|
|
|
|
|
|
|---|
| Inflammatory
factors | Control | Model | SASP, 0.6 g/kg | 1.0 | 2.0 | 4.0 |
|---|
| IL-1β,
pg/mgprot | 45.7±3.9 |
60.0±3.5a |
44.1±3.1d |
48.4±1.4c |
42.2±3.8c |
39.7±3.7d |
| IL-2,
pg/mgprot | 158.1±9.4 |
219.1±14.6a |
153.8±11.9c |
130.2±11.0d |
149.5±16.4c |
128.4±10.1d |
| IL-6,
pg/mgprot | 61.6±3.7 |
83.7±6.3a |
54.5±8.2c |
57.3±1.4c | 62.1±6.8 |
50.0±4.8d |
| IL-12,
pg/mgprot | 37.3±2.6 |
57.2±6.3a |
31.9±3.9d |
30.5±2.0d |
39.6±2.0c |
27.8±2.1d |
| TNF-α,
pg/mgprot | 407.5±21.9 |
489±22.2a | 429.0±28.9 |
374.5±27.2c |
358.0±43.1c |
337.5±14.6d |
| TNF-β,
pg/mgprot | 78.6±7.8 |
119.8±11.5b |
62.9±7.7d |
70.1±4.1d |
83.6±10.9c |
63.3±4.4d |
| IFN-α,
pg/mgprot | 19.7±1.6 |
28.8±2.6b |
16.3±1.9d |
20.7±1.2c |
20.2±1.6c |
17.2±1.8d |
| IFN-γ,
pg/mgprot | 72.4±6.6 |
126.6±14.8b |
72.9±9.5d |
75.3±5.4d |
84.2±5.5c |
69.8±7.0c |
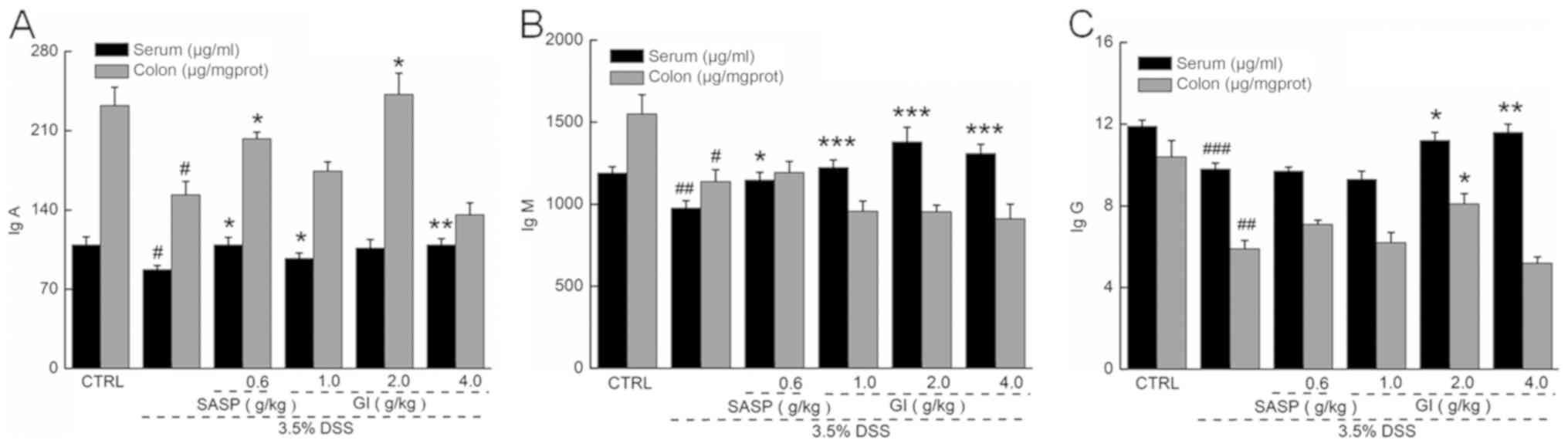
The immune response is involved in the pathogenesis
of UC (39). Compared with the
control mice, the UC mice (the model group) demonstrated lower
levels of IgA, IgG and IgM in the colon and serum (P<0.05;
Fig. 2). GI enhanced the levels of
IgA (P<0.05) (Fig. 2A) and IgG
(P<0.05; Fig. 2C) in the colon
and serum and the levels of IgM (P<0.001) (Fig. 2B) in the serum of the UC mice, but
did not enhance the levels of IgM in the colon (Fig. 2B).
Effect of GI on oxidative factors
Oxidative stress destroys the cellular
macromolecules of the colon, which is the key to the pathogenesis
of UC (40). Compared with the
control mice, the UC mice demonstrated lower levels of SOD and CAT
and higher levels of ROS and NO in the serum and colon tissues
(P<0.05; Table IV). In the
serum, GI resulted in >14.8% (P<0.01) and >17.4%
(P<0.05) increase in SOD and CAT levels and >12.9%
(P<0.05) reduction in NO levels, but no effect on ROS levels
(Table IV). In the colon tissues,
GI failed to affect CAT levels, but resulted in >36.3%
(P<0.05) increase in SOD levels and >25.8% (P<0.05) and
>45.2% (P<0.05) reductions in ROS and NO levels respectively
(Table IV). SASP significantly
prevented the pathological alterations in the levels of anti- and
pro-oxidative factors in the colon tissues (P<0.05; Table IV).
| Table IV.Effect of GI on oxidative factors in
serum and colon tissues of mice with ulcerative colitis. |
Table IV.
Effect of GI on oxidative factors in
serum and colon tissues of mice with ulcerative colitis.
| A, Serum |
|---|
|
|---|
|
|
| DSS |
|---|
|
|
|
|
|---|
|
|
|
|
| GI, g/kg |
|---|
| Oxidative
factors | CTRL | Model | SASP, 0.6 g/kg | 1.0 | 2.0 | 4.0 |
|---|
| SOD, U/ml | 99.7±3.3 |
81.7±2.6c |
94.8±3.1e | 83.9±3.1 |
93.8±2.2e |
98.2±1.9f |
| CAT, U/ml | 26.8±0.9 |
21.9±1.1b |
28.1±1.6e | 24.7±1.7 |
29.5±1.7e |
25.7±0.5d |
| ROS, U/ml | 177.1±3.1 | 174.2±3.9 | 165.6±2.8 | 167.3±5.7 | 172.4±2.1 | 179.9±0.7 |
| NO, µmol/l | 14.0±0.6 |
17.8±0.8b |
14.3±0.6d |
15.3±0.8d |
15.5±0.8d |
15.1±0.4d |
|
| B,
Colon |
|
|
|
| DSS |
|
|
|
|
|
|
|
|
| GI,
g/kg |
|
|
|
|
|
|
| Oxidative
factors | CTRL | Model | SASP, 0.6
g/kg | 1.0 | 2.0 | 4.0 |
|
| SOD, U/mgprot | 152.0±9.6 |
104.5±9.1b |
145.0±11.2d | 113.8±3.7 | 119.7±8.6 |
142.4±4.3d |
| CAT, U/mgprot | 31.3±2.2 |
21.4±2.1a |
27.8±0.9d | 22.2±0.6 | 25.3±1.9 | 22.2±1.4 |
| ROS, U/mgprot | 310.7±23.9 |
418.2±33.7a |
308.5±22.7d |
292.2±19.3d |
310.3±13.2d |
284.1±13.0d |
| NO, µmol/gprot | 13.8±1.1 |
21.9±2.8a |
11.5±1.9e |
11.6±0.9d |
12.0±1.7d |
8.0±0.5e |
GI regulates the Nrf2/NF-κB signaling
in the colon of UC mice
Compared with the model mice, the UC mice
administered with GI demonstrated lower expression levels, in the
colon, of Nrf2 and its downstream proteins, including CAT, HO-1,
SOD-1 and SOD-2, and higher phosphorylated activities of NF-κB,
IKKα/β and IκBα, all of which were prevented by GI administration
(P<0.05; Fig. 3).
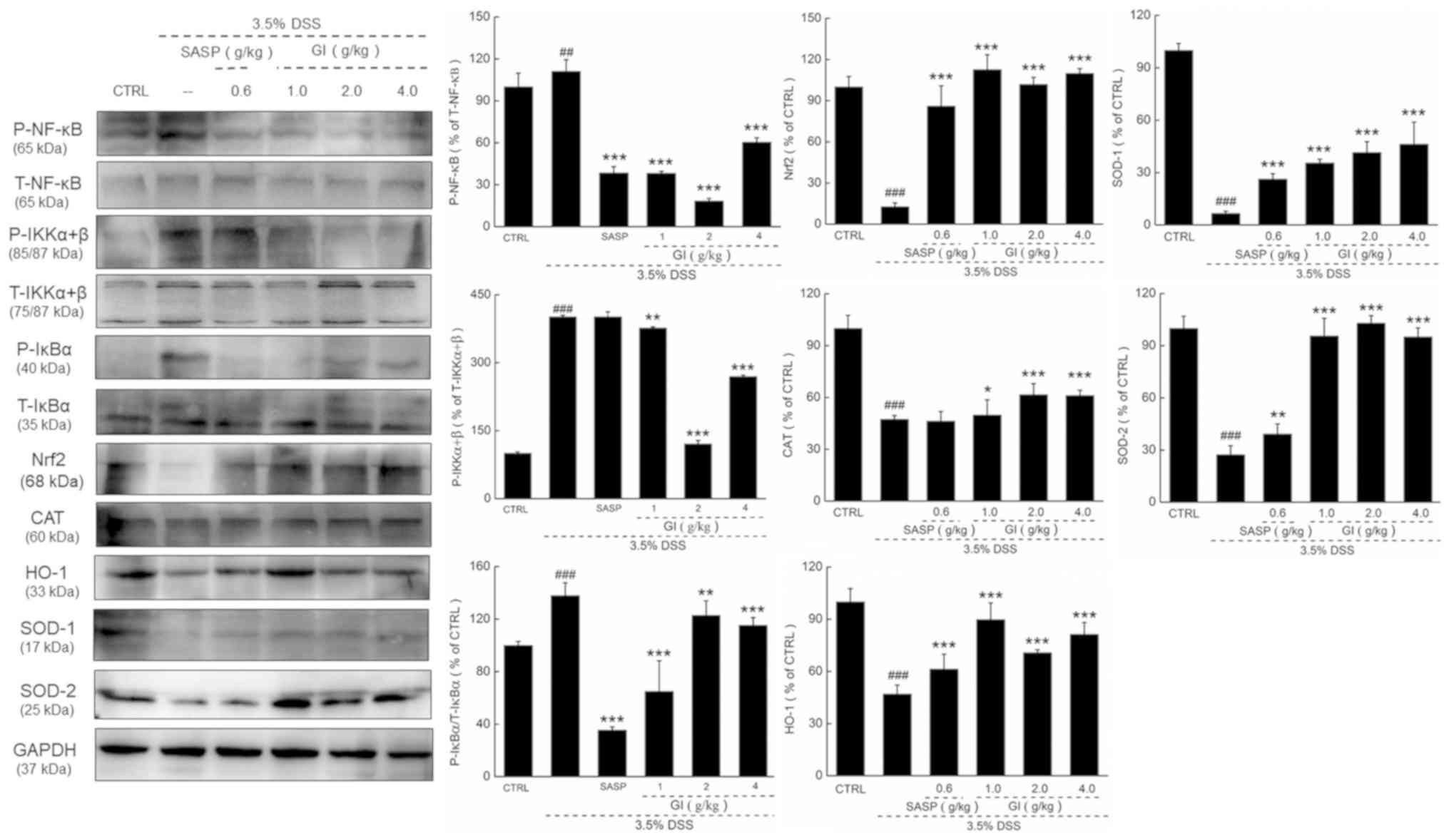 | Figure 3.GI demonstrates antioxidant and
anti-inflammatory functions via modulation the activation of
Nrf2/NF-κB signaling. GI enhanced the expression levels of Nrf2,
CAT, HO-1, SOD-1 and SOD-2, and reduced the phosphorylation level
of NF-κB, IKKα+β and IκBα. The quantitative expression of each
protein was normalized using GAPDH. ##P<0.01 and
###P<0.001 vs. control mice; *P<0.05, **P<0.01
and ***P<0.001 vs. DSS-induced UC mice. GI, Gloeostereum
incarnatum; SASP, sulfasalazine; P-, phosphorylated; T-, total;
Nrf2, nuclear factor erythroid 2-related factor 2; CAT, catalase;
HO-1, heme oxygenase-1; SOD, superoxide dismutase; IKK, inhibitor
of NF-κB kinase; IκB, inhibitor of NF-κB; DSS, dextran sulfate
sodium; CTRL, control. |
Discussion
The commonly used anti-colitis drugs mainly include
non-steroidal drugs, which exhibit side effects including
intestinal ulcers (41,42). Recently, edible mushrooms have
attracted researchers' attention due to their diverse
pharmacological effects and minimal adverse effects. For example,
Ganoderma lucidum alleviates intestinal damage induced by
indomethacin in mice (43) and
Hericium erinaceus improves testicular damage via its
antioxidative effects and regulation of the GPR41/43 receptor in UC
rats and Caco-2 cells (44). Based
on the immunomodulatory and anti-inflammatory effects of GI
reported previously (7), the
present study successfully confirmed its protective effect against
UC in C57BL/6 mice.
GI is a medicinal and edible fungus with complex
ingredients and multiple nutritional value components. Its natural
and crude character not only supports its low toxicity with various
pharmacological efficacy, but also helps to explain its non-dose
dependent effects during the experiments in the present study,
which may show anti-UC effects via multiple targets. Indeed, the
non-dose dependent manner can be noted in the research associated
with Traditional Chinese Medicine (37). GI contains 6.3% crude fiber, which
is beneficial for intestinal peristalsis and eases constipation
(45). Mixed fiber can markedly
regulate blood sugar levels, in addition to inflammatory factors,
and exhibits a beneficial regulatory effect on the intestinal
microbiota (46). GI contains
0.394% flavonoids, which exert antioxidant effects by regulating
the body's oxygen free radical levels. In an LPS-damaged mice
model, a flavonoid-rich fraction of Ocimum gratissimum
leaves regulated oxidative stress and inflammation in the liver and
brain by reducing the levels of TNF-α and malondialdehyde (47). The flavonoid apigenin can reduce
the elevation of pro-inflammatory cytokines in the colon, reduce
the density of eosinophils and transform M1 pro-inflammatory
macrophages to the M2 anti-inflammatory phenotype in diet-induced
obese mice (48). GI contains
5.328%x10−3 triterpenes, which trigger the
anti-inflammatory response and reduce the expression of
pro-inflammatory cytokines in a streptozotocin-induced diabetes
model (49). Thus, GI shows a good
nutritional foundation for its antioxidation and anti-inflammation
activities.
UC is a process driven by T helper type 2
(Th-2)-like T-cells combined with the infiltration of lymphocytes
and macrophages (50). DSS-induced
damage to the colon tissue in mice causes the activation of
macrophages, destruction of the composition of the lamina propria
cells and aggregation of inflammatory factors, which eventually
leads to imbalance in Th-1/2 cells (51). Cluster of differentiation 1a acts
as an inflammatory mediator of UC, inducing T cell activation to
elicit an immune response in the body (52). As a central factor in immune
mediation, the elevated levels of secretory IgA in the colon help
to prevent intestinal damage and restore mucosal barrier function
(53). IgG exerts important
protective and regulatory effects in the placental barrier
(54) and IgM serves an important
role in its own immune regulation (55). In the present study, GI increased
the levels of Ig in the UC mice to stimulate an immune
response.
Furthermore, TGF-β serves a role in pro-inflammatory
mediator production during the development of UC, which can
regulate the levels of ILs, TNFs and IFNs (56). During the development of UC,
signaling of the receptor-interacting protein kinase 3 can
upregulate the expression of repair-associated cytokines, including
cyclooxygenase 2 and IL-22 (57).
As a pro-inflammatory factor, abnormal release of TNF-α recruits
more macrophages and neutrophils, which in turn increases
inflammatory damage and intestinal permeability (58). In the morphological observations of
the current study, reduced goblet cells, crypt abscesses and
inflammatory cell infiltration were noted in the colon tissue
following DSS administration; these were alleviated following GI
administration. The overexpression of IL-6 may aggravate the
inflammatory cell infiltration at the injured site and recruit more
pro-inflammatory factors (8),
which further activate the inducible nitric oxide synthase and
cyclooxygenase-2 pathways, in turn increasing neutrophil
aggregation (59). In the present
study, the regulatory activity of GI on inflammatory factors may
have been involved in its protective effect against UC in C57BL/6
mice.
As an upstream regulatory protein, NF-κB can also be
activated by increased levels of pro-inflammatory factors (60). The IKKα/β complex is an aggregate
composed of a catalytic subunit (61) and is essential for the initiation
of the NF-κB pathway (62). In
general, IκBα forms a dimer with NF-κB in the cytoplasm. When the
inflammatory factor stimulates the IKKα/β complex, IKKα/β
phosphorylates IκBα, thereby separating it from NF-κB. NF-κB is
thus activated and enters the nucleus to induce the expression of
relevant genes to reduce the expression of pro-inflammatory
cytokines (63). In the present
study, GI markedly reduced the phosphorylated activation of NF-κB
by suppressing the phosphorylated IKKα/β and IκBα.
Inflammatory cells in the colon produce ROS and the
accumulation of ROS damages proteins and nucleic acids, leading to
oxidative stress (64). SOD and
CAT are enzymatic antioxidants that catalyze the decomposition of
oxides or peroxides to avoid oxidative damage (65). A previous study indicates that SOD
reduces inflammation by enhancing the body's antioxidant function
(66). SOD decomposes superoxide
into hydrogen peroxide (H2O2) and
H2O2 can be further decomposed into
H2O by CAT to exert antioxidant effects (67). The abnormal levels of anti- and
pro-oxidation factors caused by DSS in the present study were all
restored by GI treatment. Nrf2 is an important starting element in
antioxidant systems (68) that can
counteract the damaging effect of peroxide by activating the
peroxiredoxin-1 gene (69).
Under peroxidative conditions, Nrf2 dissociates from the dimer and
transfers to the nucleus, binding to the antioxidant responsive
element (70). Binding to this
promoter activates the expression of a cytoprotective enzyme,
including HO-1, CAT, or SOD (71).
Nrf2 upregulates the GST-A4 gene expression via the
mitogen-activated protein kinase pathway to protect against UC
(69). Nrf2 and NF-κB are
inseparable in the process of anti-inflammation and antioxidation.
The activation of Nrf2 attenuates inflammatory signaling by
inhibiting NF-κB entry into the nucleus, whereas NF-κB directly
inhibits the Nrf2 pathway at the RNA level (72). The data from the present study
suggested that Nrf2/NF-κB signaling is involved in GI-mediated
protection against UC; however, more experiments are required to
investigate the synergistic process between Nrf2 and NF-κB
signaling.
There remains a limitation the present study.
Although the constituents contained in GI, based on the present
data, were detected, the active ingredients with the anti-UC effect
were not isolated and purified; this will be further
investigated.
In summary, GI was identified to alleviate the
physiological and pathological state of DSS-induced UC in mice via
its antioxidant and anti-inflammatory functions, which may be
associated with its modulation of the activation of Nrf2/NF-κB
signaling.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National Key
Research & Development Program of China (grant no.
2018YFE0107800), the Science and Technology Development Program of
Jilin Province in China (grant no. 20170623092-TC), the Science and
Technology Bureau of Changchun in China (grant no. 15SS11) and the
Special Projects of the Cooperation between Jilin University and
Jilin Province of China (grant no. SXGJSF2017-1).
Availability of data and materials
All data generated and analyzed during this study
are included in this published article.
Authors' contributions
DW and YL contributed to the conceptual design of
the research. XiaL, XinL, YoZ, YaZ, SL and NZ performed the
experiments. DW, YL, XiaL and XinL analyzed the data and wrote the
manuscript. DW and YL helped perform the analysis with constructive
discussions. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The experimental animal protocol was approved by the
Animal Ethics Committee of Jilin University [approval no. SYXK (JI)
2014-0013]. The present study was carried out under relevant
guidelines and regulations.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Yue B, Ren YJ, Zhang JJ, Luo XP, Yu ZL,
Ren GY, Sun AN, Deng C, Wang ZT and Dou W: Anti-inflammatory
effects of fargesin on chemically induced inflammatory bowel
disease in mice. Molecules. 23:34182018. View Article : Google Scholar
|
|
2
|
Cawthorpe D and Davidson M: Temporal
comorbidity of mental disorder and ulcerative colitis. Perm J.
19:52–57. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Vargas Robles H, Citalán Madrid AF, García
Ponce A, Silva Olivares A, Shibayama M, Betanzos A, Del Valle
Mondragón L, Nava P and Schnoor M: Experimental colitis is
attenuated by cardioprotective diet supplementation that reduces
oxidative stress, inflammation, and mucosal damage. Oxid Med Cell
Longev. 2016:84732422016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bopanna S, Ananthakrishnan AN, Kedia S,
Yajnik V and Ahuja V: Risk of colorectal cancer in Asian patients
with ulcerative colitis: A systematic review and meta-analysis.
Lancet Gastroenterol Hepatol. 2:269–276. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Tatiya-Aphiradee N, Chatuphonprasert W and
Jarukamjorn K: Immune response and inflammatory pathway of
ulcerative colitis. J Basic Clin Physiol Pharmacol. 30:1–10. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chaussade S and Denizot Y: Mediators of
inflammation and hemorrhagic rectocolitis. Ann Gastroenterol
Hepatol (Paris). 27:117–121. 1991.(In French). PubMed/NCBI
|
|
7
|
Wang D, Li Q, Qu Y, Wang M, Li L, Liu Y
and Li Y: The investigation of immunomodulatory activities of
Gloeostereum incaratum polysaccharides in
cyclophosphamide-induced immunosuppression mice. Exp Ther Med.
15:3633–3638. 2018.PubMed/NCBI
|
|
8
|
Zhang H and Chen W: Interleukin 6
inhibition by triptolide prevents inflammation in a mouse model of
ulcerative colitis. Exp Ther Med. 14:2271–2276. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Jeon YD, Bang KS, Shin MK, Lee JH, Chang
YN and Jin JS: Regulatory effects of Glycyrrhizae radix extract on
DSS-induced ulcerative colitis. BMC Complement Altern Med.
16:4592016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ai XY, Qin Y, Liu HJ, Cui ZH, Li M, Yang
JH, Zhong WL, Liu YR, Chen S, Sun T, et al: Apigenin inhibits
colonic inflammation and tumorigenesis by suppressing STAT3-NF-κB
signaling. Oncotarget. 8:100216–100226. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang Z, Li S, Cao Y, Tian X, Zeng R, Liao
DF and Cao D: Oxidative stress and carbonyl lesions in ulcerative
colitis and associated colorectal cancer. Oxid Med Cell Longev.
2016:98752982016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Jin Y, Yang J, Lin L, Lin Y and Zheng C:
The attenuation of Scutellariae radix extract on oxidative stress
for colon injury in lipopolysaccharide-induced RAW264.7 cell and
2,4,6-trinitrobenzene sulfonic acid-induced ulcerative colitis
rats. Pharmacogn Mag. 12:153–159. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Bellezza I, Giambanco I, Minelli A and
Donato R: Nrf2-Keap1 signaling in oxidative and reductive stress.
Biochim Biophys Acta Mol Cell Res. 1865:721–733. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhang ZL, Fan HY, Yang MY, Zhang ZK and
Liu K: Therapeutic effect of a hydroxynaphthoquinone fraction on
dextran sulfate sodium-induced ulcerative colitis. World J
Gastroenterol. 20:15310–15318. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Shin MR, Kim KJ, Kim SH, Kim SJ, Seo BI,
An HJ and Roh SS: Comparative evaluation between sulfasalazine
alone and in combination with herbal medicine on DSS-induced
ulcerative colitis mice. BioMed Res Int. 2017:67426522017.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Asai R, Mitsuhashi S, Shigetomi K,
Miyamoto T and Ubukata M: Absolute configurations of (−)-hirsutanol
A and (−)-hirsutanol C produced by Gloeostereum incarnatum.
J Antibiot (Tokyo). 64:693–696. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Soo LT: Free radical scavenging,
anti-inflammatory and melannin synthesis inhibitory activities of
Gloeostereum incarnatum. J Mushrooms. 12:107–116. 2014.
View Article : Google Scholar
|
|
18
|
Bunbamrung N, Intaraudom C, Dramae A,
Boonyuen N, Veeranondha S, Rachtawee P and Pittayakhajonwut P:
Antimicrobial activity of illudalane and alliacane sesquiterpenes
from the mushroom Gloeostereum incarnatum BCC41461.
Phytochem Lett. 20:274–281. 2017. View Article : Google Scholar
|
|
19
|
Rover MR, Johnston PA, Lamsal BP and Brown
RC: Total water-soluble sugars quantification in bio-oil using the
phenol-sulfuric acid assay. J Anal Appl Pyrolysis. 104:194–201.
2013. View Article : Google Scholar
|
|
20
|
Lindsay H: A colorimetric estimation of
reducing sugars in potatoes with 3,5-dinitrosalicylic acid. Potato
Res. 16:176–179. 1973. View Article : Google Scholar
|
|
21
|
Devapriya F, Sajith P, Ranganathan R and
Shanmugam J: Prevalence of biofilm and beta-lactamase producing
Staphylococcus in nasal and throat isolates from healthy
volunteers: A medical alert. Nepal J Med Sci. 3:79–83. 2014.
View Article : Google Scholar
|
|
22
|
Popek S: Application of regression
analysis as a method to determine total ash content in some
selected nectar honeys. Nahrung. 47:36–38. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bahloul N, Bellili S, Aazza S, Chérif A,
Faleiro ML, Antunes MD, Miguel MG and Mnif W: Aqueous extracts from
tunisian diplotaxis: Phenol content, antioxidant and
anti-acetylcholinesterase activities, and impact of exposure to
simulated gastrointestinal fluids. Antioxidants. 5:122016.
View Article : Google Scholar
|
|
24
|
Li JJ, Hu XQ, Zhang XF, Liu JJ and Cao LS:
Study on variation of main ingredients from spores and fruiting
bodies of Ganoderma lucidum. Zhongguo Zhong Yao Za Zhi.
39:4246–4251. 2014.(In Chinese). PubMed/NCBI
|
|
25
|
Dong QF, Wang JL, Zhang SF, Wang Z, Zhang
CX, Gao H, Zhang HM and Zhang L: Antifungal activity of crude
extracts and fat-soluble constituents of Holotrichia diomphalia
larvae. Bioresour Technol. 99:8521–8523. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Marichal MJ, Trujillo AI, Cadenazzi M and
Arias G: Fiber analysis: Evaluation of screen printing fabric
filters bags by three statistical approaches. Anim Feed Sci
Technol. 169:79–85. 2011. View Article : Google Scholar
|
|
27
|
da Costa WKOC, da Silva CS, Figueiredo
JFD, Nóbrega JA and Paim APS: Direct analysis of deodorants for
determination of metals by inductively coupled plasma optical
emission spectrometry. J Pharm Biomed Anal. 155:247–252. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Utami PI, Rahayu WS, Nugraha I and Rochana
AN: Fatty acid analysis of lipid extracted from rats by gas
chromatography-mass spectrometry method. 2ND Annual applied science
and engineering conference (AASEC 2017). IOP Conf Series Mater Sci
Eng. 2882018.
|
|
29
|
Paquin CS and Paquin R: Integration,
identification and concentration measurement of amino acids in
plant samples by means of an automatic amino acid analyzer linked
to a mini-computer. J Chromatogr A. 156:79–85. 1978. View Article : Google Scholar
|
|
30
|
Couto M and Cates C: Laboratory guidelines
for animal care. Methods Mol Biol. 1920:407–430. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
China sets lab animal guidelines. Science.
351:1372–1373. 2016.
|
|
32
|
Committee for the Update of the Guide for
the Care and Use of Laboratory Animals: Guide for the Care and Use
of Laboratory Animals: Eighth Edition. The National Academies
Press; Washington, DC: 2011
|
|
33
|
Gan HT, Chen YQ and Ouyang Q:
Sulfasalazine inhibits activation of nuclear factor-kappaB in
patients with ulcerative colitis. J Gastroenterol Hepatol.
20:1016–1024. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Cao Z, Dai W, Zhang R, Chen L, Yang X, Hu
L, Chiang LY and Liu W: Opening of the adenosine
triphosphate-sensitive potassium channel attenuates morphine
tolerance by inhibiting JNK and astrocyte activation in the spinal
cord. Clin J Pain. 32:617–623. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Taba MY, Mohammadi S, Jalali M, Beheshti F
and Attari SS: Effects of different doses of curcumin on testicular
histopathology, apoptosis, and reproductive organs weight index in
mice D-galactose-induced aging model. Comp Clin Pathol.
28:997–1002. 2019. View Article : Google Scholar
|
|
36
|
Wang R, Wang L, Luo Y, Wang D, Du R, Du J
and Wang Y: Maggot protein ameliorates dextran sulphate
sodium-induced ulcerative colitis in mice. Biosci Rep.
38:BSR201817992018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Meng B, Zhang Y, Wang Z, Ding Q, Song J
and Wang D: Hepatoprotective effects of Morchella esculenta
against alcohol-induced acute liver injury in the C57BL/6 mouse
related to Nrf-2 and NF-κB signaling. Oxid Med Cell Longev.
6029876:20192019.
|
|
38
|
Sarlos P, Kovesdi E, Magyari L, Banfai Z,
Szabo A, Javorhazy A and Melegh B: Genetic update on inflammatory
factors in ulcerative colitis: Review of the current literature.
World J Gastrointest Pathophysiol. 5:304–321. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Gong Y, Niu W, Tang Y, Zhang Q, Liu S, Liu
X, Wang X and Xu Y: Aggravated mucosal and immune damage in a mouse
model of ulcerative colitis with stress. Exp Ther Med.
17:2341–2348. 2019.PubMed/NCBI
|
|
40
|
Yan H, Wang H, Zhang X, Li X and Yu J:
Ascorbic acid ameliorates oxidative stress and inflammation in
dextran sulfate sodium-induced ulcerative colitis in mice. Int J
Clin Exp Med. 8:20245–20253. 2015.PubMed/NCBI
|
|
41
|
Stadnicki A and Frysz-Naglak D:
Non-steroidal anti-inflammatory drugs and intestinal side effects.
Wiad Lek. 60:286–290. 2007.(In Polish). PubMed/NCBI
|
|
42
|
Lychkova AÉ and Puzikov AM: Non-steroidal
anti-inflammatory drugs in the correction of experimental
ulcerative colitis. Eksp Klin Gastroenterol. 7:59–63. 2014.(In
Russian).
|
|
43
|
Nagai K, Ueno Y, Tanaka S, Hayashi R,
Shinagawa K and Chayama K: Polysaccharides derived from
Ganoderma lucidum fungus mycelia ameliorate
indomethacin-induced small intestinal injury via induction of
GM-CSF from macrophages. Cell Immunol. 320:20–28. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Wang D, Zhang Y, Yang S, Zhao D and Wang
M: A polysaccharide from cultured mycelium of Hericium
erinaceus relieves ulcerative colitis by counteracting
oxidative stress and improving mitochondrial function. Int J Biol
Macromol. 125:572–579. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Mehmood MH, Aziz N, Ghayur MN and Gilani
AH: Pharmacological basis for the medicinal use of Psyllium husk
(Ispaghula) in constipation and diarrhea. Dig Dis Sci.
56:1460–1471. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Zhai X, Lin D, Zhao Y, Li W and Yang X:
Effects of dietary fiber supplementation on fatty acid metabolism
and intestinal microbiota diversity in C57BL/6J mice fed with a
high-fat diet. J Agric Food Chem. 66:12706–12718. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Ajayi AM, Ben-Azu B, Onasanwo SA, Adeoluwa
O, Eduviere A and Ademowo OG: Flavonoid-rich fraction of Ocimum
gratissimum attenuates lipopolysaccharide-induced sickness
behavior, inflammatory and oxidative stress in mice. Drug Res
(Stuttg). 69:151–158. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Gentile D, Fornai M, Colucci R, Pellegrini
C, Tirotta E, Benvenuti L, Segnani C, Ippolito C, Duranti E, Virdis
A, et al: The flavonoid compound apigenin prevents colonic
inflammation and motor dysfunctions associated with high fat
diet-induced obesity. PLoS One. 13:e01955022018. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Wu YS and Chen SN: Extracted triterpenes
from Antrodia cinnamomea reduce the inflammation to promote
the wound healing via the STZ inducing hyperglycemia-diabetes mice
model. Front Pharmacol. 7:1542016. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Majewska-Szczepanik M, Góralska M,
Marcińska K, Zemelka-Wiącek M, Strzępa A, Dorożyńska I and
Szczepanik M: Epicutaneous immunization with protein antigen TNP-Ig
alleviates TNBS-induced colitis in mice. Pharmacol Rep.
64:1497–1504. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Queen D, Hedayat AA, Magro C and Geskin
LJ: An unusual cause of bilateral orbital swelling: Immunoglobulin
G4-related orbital disease arising in a patient with ulcerative
colitis. JAAD Case Rep. 5:634–638. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Al-Amodi O, Jodeleit H, Beigel F, Wolf E,
Siebeck M and Gropp R: CD1a-expressing monocytes as mediators of
inflammation in ulcerative colitis. Inflamm Bowel Dis.
24:1225–1236. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Cui Y, Zhu C, Ming Z, Cao J, Yan Y, Zhao
P, Pang G, Deng Z, Yao Y and Chen Q: Molecular mechanisms by which
casein glycomacropeptide maintains internal homeostasis in mice
with experimental ulcerative colitis. PLoS One. 12:e01810752017.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Ignat'eva NV, Ziganshina MM, Shilova NV,
Khasbiullina NR, Bovin NV, Tyutyunnik VL and Sukhikh GT: Isolation
of IgG associated with human placenta. Bull Exp Biol Med.
167:120–122. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Kubagawa H, Honjo K, Ohkura N, Sakaguchi
S, Radbruch A, Melchers F and Jani PK: Functional roles of the IgM
Fc receptor in the immune system. Front Immunol. 10:9452019.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Binabaj MM, Asgharzadeh F, Avan A, Rahmani
F, Soleimani A, Parizadeh MR, Ferns GA, Ryzhikov M, Khazaei M and
Hassanian SM: EW-7197 prevents ulcerative colitis-associated
fibrosis and inflammation. J Cell Physiol. 234:11654–11661. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Xu YL, Tang HL, Zhu SY, Peng HR, Qi ZT and
Wang W: RIP3 deficiency exacerbates inflammation in dextran sodium
sulfate-induced ulcerative colitis mice model. Cell Biochem Funct.
35:156–163. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Liang J, Liang J, Hao H, Lin H, Wang P, Wu
Y, Jiang X, Fu C, Li Q, Ding P, et al: The extracts of Morinda
officinalis and its hairy roots attenuate dextran sodium
sulfate-induced chronic ulcerative colitis in mice by regulating
inflammation and lymphocyte apoptosis. Front Immunol. 8:9052017.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Yang Y, He J, Suo Y, Lv L, Wang J, Huo C,
Zheng Z, Wang Z, Li J, Sun W and Zhang Y: Anti-inflammatory effect
of taurocholate on TNBS-induced ulcerative colitis in mice. Biomed
Pharmacother. 81:424–430. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Gupta RA, Motiwala MN, Dumore NG, Danao KR
and Ganjare AB: Effect of piperine on inhibition of FFA induced
TLR4 mediated inflammation and amelioration of acetic acid induced
ulcerative colitis in mice. J Ethnopharmacol. 164:239–246. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Koppe C, Verheugd P, Gautheron J,
Reisinger F, Kreggenwinkel K, Roderburg C, Quagliata L, Terracciano
L, Gassler N, Tolba RH, et al: IκB kinaseα/β control biliary
homeostasis and hepatocarcinogenesis in mice by phosphorylating the
cell-death mediator receptor-interacting protein kinase 1.
Hepatology. 64:1217–1231. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Li Y, Li J, Li B, Qin H, Peng X, Zhao Y
and Chen Y: Anthocyanin suppresses CoCrMo particle-induced
osteolysis by inhibiting IKKα/β mediated NF-κB signaling in a mouse
calvarial model. Mol Immunol. 85:27–34. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Reale C, Iervolino A, Scudiero I,
Ferravante A, D'Andrea LE, Mazzone P, Zotti T, Leonardi A, Roberto
L, Zannini M, et al: NF-κB essential modulator (NEMO) is critical
for thyroid function. J Biol Chem. 291:5765–5773. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Nikkhah-Bodaghi M, Maleki I, Agah S and
Hekmatdoost A: Zingiber officinale and oxidative stress in
patients with ulcerative colitis: A randomized, placebo-controlled,
clinical trial. Complement Ther Med. 43:1–6. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Meurer MC, Mees M, Mariano LNB, Boeing T,
Somensi LB, Mariott M, da Silva RCMVAF, Dos Santos AC, Longo B,
Santos França TC, et al: Hydroalcoholic extract of Tagetes
erecta L. flowers, rich in the carotenoid lutein, attenuates
inflammatory cytokine secretion and improves the oxidative stress
in an animal model of ulcerative colitis. Nutr Res. 66:95–106.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Kang JE, Kim HD, Park SY, Pan JG, Kim JH
and Yum DY: Dietary supplementation with a Bacillus superoxide
dismutase protects against γ-radiation-induced oxidative stress and
ameliorates dextran sulphate sodium-induced ulcerative colitis in
mice. J Crohn's Colitis. 12:860–869. 2018. View Article : Google Scholar
|
|
67
|
Rana SV, Sharma S, Prasad KK, Sinha SK and
Singh K: Role of oxidative stress & antioxidant defence in
ulcerative colitis patients from north India. Indian J Med Res.
139:568–571. 2014.PubMed/NCBI
|
|
68
|
Almeer RS, Mahmoud SM, Amin HK and Abdel
Moneim AE: Ziziphus spina-christi fruit extract suppresses
oxidative stress and p38 MAPK expression in ulcerative colitis in
rats via induction of Nrf2 and HO-1 expression. Food Chem Toxicol.
115:49–62. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Sabzevary-Ghahfarokhi M, Shohan M, Shirzad
H, Rahimian G, Soltani A, Ghatreh-Samani M, Deris F, Bagheri N,
Shafigh M and Tahmasbi K: The regulatory role of Nrf2 in
antioxidants phase2 enzymes and IL-17A expression in patients with
ulcerative colitis. Pathol Res Pract. 214:1149–1155. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Liu D, Huo X, Gao L, Zhang J, Ni H and Cao
L: NF-κB and Nrf2 pathways contribute to the protective effect of
Licochalcone A on dextran sulphate sodium-induced ulcerative
colitis in mice. Biomed Pharmacother. 102:922–929. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Li J, Wang H, Zheng Z, Luo L, Wang P, Liu
K, Namani A, Jiang Z, Wang XJ and Tang X: Mkp-1 cross-talks with
Nrf2/Ho-1 pathway protecting against intestinal inflammation. Free
Radic Biol Med. 124:541–549. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Saber S, Khalil RM, Abdo WS, Nassif D and
El-Ahwany E: Olmesartan ameliorates chemically-induced ulcerative
colitis in rats via modulating NFκB and Nrf-2/HO-1 signaling
crosstalk. Toxicol Appl Pharmacol. 364:120–132. 2019. View Article : Google Scholar : PubMed/NCBI
|