Hepatitis C virus (HCV) is an infectious disease of
global concern. The World Health Organization suggests that there
are >71 million individuals infected with HCV worldwide and
~475,000 deaths are caused by HCV infections annually (1,2). The
prevalence and incidence of HCV infections are the highest in low-
and middle-income countries (3,4). The
HCV can cause chronic liver disease, which is long-lasting and can
progress to fibrosis, cirrhosis and hepatocellular carcinoma (HCC)
if it is not treated in a timely manner (5–7). The
dormant period of the HCV is long, sometimes up to 20–30 years, and
the clinical symptoms are not obvious, thus they are easy to ignore
once infected (8,9).
During the early course of HCV infection, the
activation and proliferation of hepatic stellate cells (HSCs) is
the central link in the development of liver fibrosis (10,11).
When the HCV infection causes damage to the liver, cytokines and
reactive oxygen species released by the tissues activate HSCs to
develop into myofibroblasts (MFBs), which undergo considerable
proliferation, and secrete collagen and metalloproteinase
inhibitors (12–14), the secretion of which notably
increases extracellular matrix deposition and decreases its
degradation, respectively (15).
The excessive deposition of the extracellular matrix results in the
destruction of the structure of the liver, eventually leading to
the development of liver fibrosis (16).
At present, several serological makers and viral
antigens in the serum, such as HCV antibody (Ab), HCV core antigen
(cAg) and HCV-RNA, are routinely assessed for to diagnose the
status of the HCV infection and liver disease associated with the
HCV (17–19). Nevertheless, given the complexity
and variability in HCV infections, the levels of serological
markers do not sufficiently reflect the status of HCV infection and
disease progression (20,21). Therefore, it is necessary to
identify additional indices that may be used to differentiate
varying degrees of HCV progression. The changes in the function and
status of HSCs was discovered to be closely associated with the
course of HCV infection (22).
Therefore, in the present review, the functional changes of
important cell populations, such as HSCs, during alterations to
liver immune function following the infection with HCV are
described, with the aim of highlighting avenues for the
identification of complementary laboratory indices that may be used
for the diagnosis of HCV infection.
As early as the 19th century, German scientist von
Kuffer first discovered the presence of stellate cells in the
hepatic sinus space (23). In
1996, international standardization stipulated that such stellate
cells were named HSCs (24). In
addition to the known functions of storing lipid droplets and
participating in fibrosis, HSCs also possess several other
important functions, such as their role in mediating the immune
response of the liver (25–27).
Previous studies have reported that HSCs were heterogeneous and
plastic in the liver, and different subsets of HSC phenotypes
exist, which all have various functions (28–30).
In the physiological liver, HSCs exist in a quiescent
non-proliferative state, termed quiescent (q)HSCs (31). qHSCs were discovered to serve a
role in the storage and transport of retinoids (vitamin A
compounds), and the quantity of vitamin A lipid droplets in the
cytoplasm of qHSCs in the liver may account for up to 45–72% of the
total content in the human body (32). Nervous system markers, such as
glial fibril acidic protein and neurotensin, nerve growth factor
receptor and desmin were also discovered to be expressed in qHSCs
(33,34). qHSCs can also secrete extracellular
matrix protein and protein substances, such as laminin,
polysaccharide protein and type IV collagen, which are required for
the formation of the basement membrane (35–37).
Following chronic liver damage as a result of
alcoholic steatohepatitis, non-alcoholic fatty liver disease,
hepatitis B or HCV infection, or cholestatic liver injury, several
external stimuli and cell types converge upon HSCs to promote their
activation and development into MFB cells (38–40).
The other cell types involved in the activation of HSCs include
liver macrophages, hepatic sinusoidal endothelial cells, natural
killer (NK) cells, B cells and hepatocytes (41). These cells secrete various
components, including oxidative stress products, cytokines and
apoptotic bodies, amongst others, to activate the HSCs (42–44).
MFBs possess a potent ability to contract and migrate through the
upregulation of fibrosis markers, such as type I collagen, α-smooth
muscle actin (SMA), matrix metalloproteinases and tissue inhibitor
of matrix metalloproteinases (45–47).
α-SMA is present in vascular smooth muscle cells and fibroblasts,
and is a widely recognized marker of HSC activation (48,49).
In addition, previous studies have identified that cytokine
receptor-like factor 1, secreted phosphoprotein 1, lysyl oxidase,
lysyl oxidase-like 2 and IL-17 receptor A are also recognized
markers associated with the activated phenotype of qHSCs, and are
upregulated following activation (50–54).
HSCs also secrete chemokines to recruit cells and
regulate the local immune microenvironment (55). Current studies have reported that
HSCs expressed chemokine receptors, such as C-C chemokine receptor
type (CCR)5, CCR7 and C-X-C motif chemokine receptor (CXCR)3, and
secreted the chemokines C-C motif chemokine (CCL)5, CCL3, CCL2,
C-X-C motif chemokine ligand (CXCL)10, CXCL9 and CXCL8 (56,57).
Chronic HCV infection can cause liver damage and
result in a range of mild to more severe diseases, such as chronic
hepatitis C (CHC), fibrosis and cirrhosis (67). The activation of HSCs was
identified as an important signal in the control of extracellular
matrix synthesis and degradation in HCV-induced liver fibrosis
(68). At the cellular level, the
activatory properties on HSCs are considered to be associated with
the amino domain of HCV core protein (69). Furthermore, HCV infection was
discovered to stimulate the innate immune response, and changes in
the functions of HSCs were also affected by other important immune
cells, such as NK cells, natural killer T (NKT) cells and
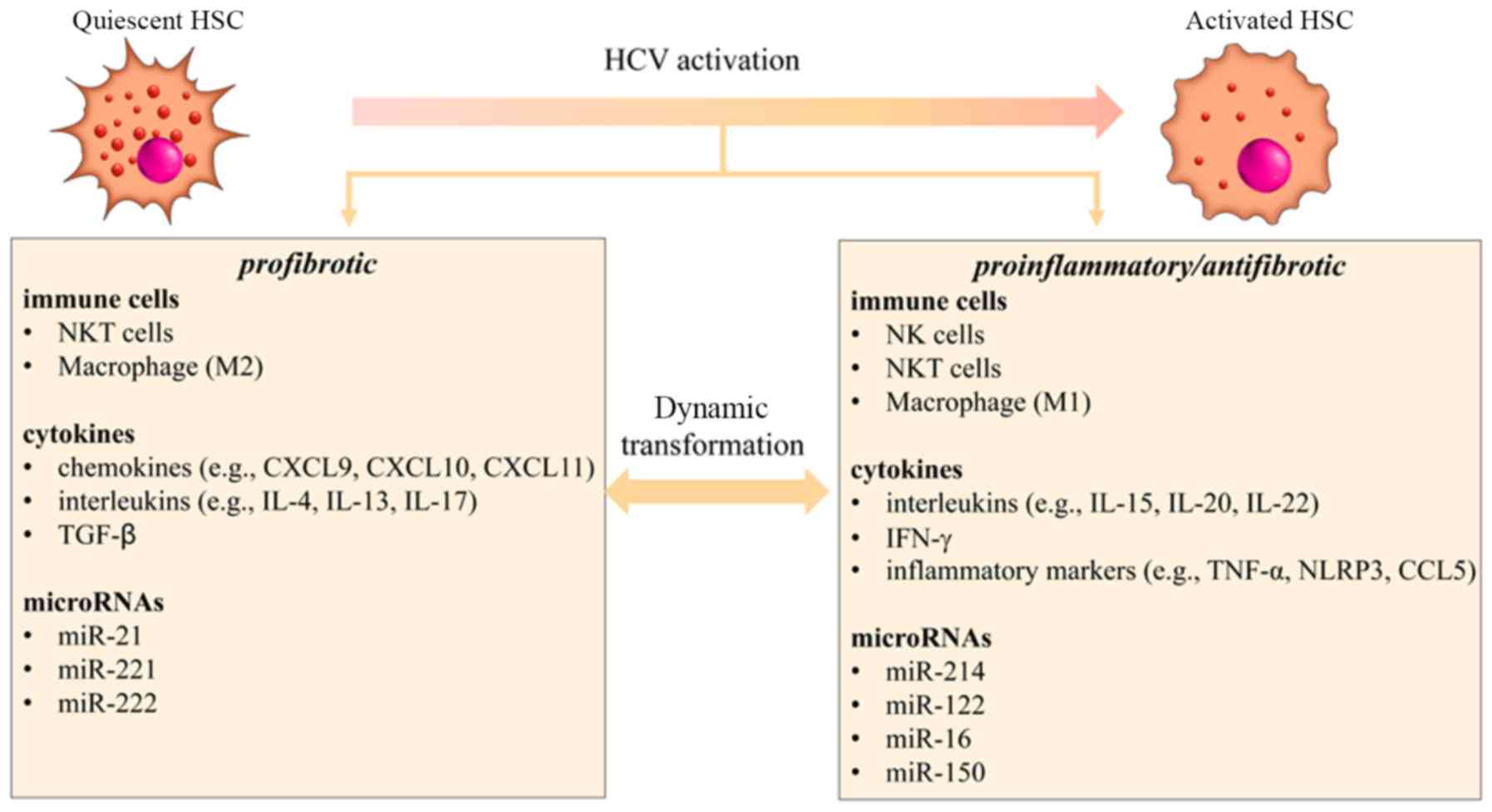
macrophages (70,71). At the molecular level, during HCV
infection, two types of cytokines that are closely associated with
the activation of HSCs were discovered to serve different roles;
one type of chemokine primarily promoted fibrosis, such as CXCL9,
CXCL10, CXCL11, IL-4, IL-13, IL-17 and TGF-β, while the other type
of cytokine contributed to the inflammatory response, such as IL-5,
IL-20, IL-22, IFN-γ, TNF-α and CCL5 (72). At the mRNA level, previous studies
have illustrated that several different microRNAs (miRNAs/miRs)
were abnormally expressed in HCV-induced liver fibrosis, such as
miR-16, miR-21, miR-122, miR-150, miR-214 and miR-221, where they
were involved in the activation of HSCs (Fig. 1) (73–75).
HSCs can express several HCV co-receptors that
interact with the HCV proteins to promote liver fibrosis (76). For example, the binding of the HCV
E2 protein and CD81 on the surface of HSCs may result in an
increase in the fibrogenic effects occur to HSCs (77,78).
In addition, the expression of HCV core and NS3-NS5 proteins was
suggested to promote HSC proliferation and induce the secretion of
proinflammatory cytokines in HSCs, such as IL-8 and
monocyte-chemotactic protein-1 (79,80).
Exosomes are small extracellular vesicles that are
secreted by the majority of cells through the endocytic pathway;
however, there is no direct contact between the different cells.
Exosomes carry different biomolecules and are therefore an
important vehicle for intracellular and intercellular communication
(81). Notably, previous studies
have confirmed the existence of exosome-mediated communications
between HCV-infected hepatocytes and HSCs (82,83).
Exosomes secreted from HCV-infected hepatocytes (HCV-exo) were
discovered to possess the potential to activate HSCs (84). A high expression of miR-19a in
exosomes was observed from HCV-exo, which in turn enhances fibrosis
marker genes and activates the STAT3-mediated TGF-β signaling
pathway (85). In a previous study
on the activation and function of NK cells in the pathogenesis of
HCV infection, hepatocytes were reported to produce a host of
cytokines, including IFN-α/β, which activated NK cells and enhanced
NK cell-mediated cell cytotoxicity (86). NK cells generally display
antifibrotic properties, including the inhibition of liver fibrosis
by selectively expressing death receptor ligands for the receptors
on activated HSCs and by producing the antifibrotic cytokine IFN-γ
(87). In addition, NKT cells were
revealed to perform similar antifibrotic functions as NK cells by
killing HSCs and producing IFN-γ (88). However, NKT cells also produce
profibrotic cytokines to promote liver fibrogenesis; for example,
the secretion of IL-4 and IL-13 from NKT cells were significantly
increased in patients with HCV with cirrhosis (89). Macrophages are divided into two
phenotypically and functionally distinct subsets, classically and
alternatively (M2) activated macrophages (90). M2 macrophages are considered to
possess anti-inflammatory and profibrotic effects by producing
profibrotic cytokines (CCL3, CCL5, TGF-β and TNF-α) (91). Clinically, it has also been
reported that the expression levels of CCL5 are upregulated in the
serum and liver of patients with HCV (56).
IL-15 and its high-affinity receptor IL-15 receptor
(R)α are widely expressed in immune cells and liver cells (107,108). NK cells, NKT cells and
CD8+ T cells generate IL-15 to maintain system
homeostasis and promote liver regeneration (109). A previous study confirmed that
IL-15 and IFN-γ exhibited protective effects against HCV via the
ERK signaling pathway in vitro (110). Jiao et al (108) reported that increased fibrosis
was observed in IL-15Rα knockout (KO) mice. Furthermore, the study
demonstrated that collagen production was increased in HSCs
isolated from IL-15RαKO mice. Therefore, IL-15 and IL-15Rα may
serve a protective role in the development of liver fibrosis by
regulating the expression levels of fibrotic molecules and collagen
in HSCs and maintaining the balance of NK cells in vivo.
IL-17 was discovered to be important in the
development of liver fibrosis in mice (111). It was reported that IL-17 was
strongly associated with an improved prognosis in patients with CHC
(112). Thus, IL-17 may be a
therapeutic target for the treatment of fibrosis. IL-17 regulates
fibrosis through two separate mechanisms. First, IL-17 stimulates
macrophages to express the inflammatory cytokines IL-6, IL-1β and
TNF-α, as well as the major fibrogenic cytokine, TGF-β1 (113). Additionally, IL-17 directly
stimulates HSCs to express type I collagen and promotes their
activation, and MFBs were discovered to be formed through the STAT3
signaling pathway during the fibrosis of HSCs (114–116).
IL-20, a proinflammatory cytokine in the IL-10
cytokine family, reportedly activates qHSCs and upregulates TGF-β
expression levels (117,118). In a mouse model of cell injury
induced by CCL4, the use of antibodies to neutralize IL-20 or IL-20
receptors inhibited HSC activation and liver fibrosis, and
downregulated TGF-β production (119).
IL-22 is also a member of the IL-10 cytokine family
that activates the STAT3 signaling pathway in hepatocytes, which
has been illustrated to promote the development of HCC (120). IL-22 simultaneously expresses
IL-10 receptor 2 and IL-22 receptor 1, both of which were
identified to induce the senescence of HSCs, thereby improving
liver fibrosis (121). Previous
studies have revealed that the increase in IL-22 in the liver of
mice reduced the expression of fibrosis-associated genes, and
accelerated the recovery of the liver damage caused by fibrosis by
increasing the number of senescent HSCs and decreasing the
expression levels of α-SMA (122–124).
miRNAs are small non-coding RNAs of ~22 nucleotides
in length that regulate post-transcriptional gene expression by
altering mRNA degradation (125).
The abnormal expression of different miRNAs, such as miR-122,
miR-126, miR-129, miR-199a and miR-155, in HCV-induced liver
fibrosis and HCC has been previously reported (126). Activated HSCs were discovered to
express a low number of miRNAs (n=259), of which 47 were
downregulated and 212 were upregulated upon activation (127). Clinical data also revealed that
miR-21 expression levels were associated with the viral load,
fibrosis and serum liver transaminase levels (128). It was also identified that
miR-221/222 expression levels were upregulated in the human liver,
and the upregulation was dependent on the progression of fibrosis
(129). In addition, the
increased expression levels of miR-221/222 have also been confirmed
in a mouse model of liver fibrosis (130). By contrast, antifibrotic miRNAs
include miR-19 (85), miR-214
(131), miR-16 (132), miR-122 (133) and miR-150 (134), amongst others. For example,
connective tissue growth factor 2 (CCN2) was discovered to drive
fibrogenesis in HSCs (135),
while in the fibrotic or steatotic liver, the upregulation of CCN2
was associated with the mutual downregulation of miR-214 (131). The expression levels of miR-122
were also discovered to be negatively correlated with fibrosis,
liver transaminase levels and patient age in another study
(134).
At present, the primary markers used to detect the
presence of the HCV in the laboratory include HCV-Ab, HCV-cAg,
HCV-RNA, and the presence of an HCV genotype and subtype (136,137). For the initial diagnosis, the
most commonly used methods for detection are ELISAs and
chemiluminescence immunoassays (CLIAs) for analyzing the presence
of HCV-Ab in the blood, as they are relatively easy to perform and
provide results quickly (138,139). However, the HCV-Ab test often
provides false-positive results in patients with a chronic
infectious disease (140–142). For example, it was reported that
in 477 individuals with an anti-HCV response analyzed using a
recombinant immunoblot assay (RIBA), 105 (22%) were confirmed as
false positives (143). Thus, if
a sample is reactive in the primary screening test, further tests
are required to confirm this result. The additional tests usually
used are RIBAs or nucleic acid amplification assays (NATs)
(2,144). NATs are more specific than ELISAs
or CLIAs and have a higher detection accuracy, but a shorter time
window for detection (145). In
addition, NATs require specific laboratory equipment and trained
personnel; thus, it is difficult to perform this assay in
conventional laboratories (146).
Liver biopsies to measure liver fibrosis have been
almost completely replaced by noninvasive methods, including the
detection of biochemical markers, such as alanine aminotransferase
(ALT) and aspartate aminotransferase (AST), as well as scoring
systems, such as the AST to platelet ratio index (APRI) score and
fibrosis (FIB)-4 score (147,148). The scoring systems of APRI and
FIB 4 are generally cheap and simple to use for the evaluation of
liver fibrosis; however, FIB-4 and APRI have been shown to have a
considerably higher rate of false-negatives or false-positives in
the detection of both fibrosis and cirrhosis (20). Thus, the results should be further
confirmed using more accurate tests.
Aberrant lymphocyte proliferation is a primary
characteristic of CHC, with evidence of focal and bridging necrosis
and lobular degeneration in the portal area (149). These lesions can be observed by
pathological examination, such as hematoxylin and eosin staining
and immunohistochemical staining (150). However, the pathogenesis and
mechanism underlying the formation of liver lesions in the process
of CHC infection has not been fully determined; therefore, it is
difficult to accurately diagnose HCV infection through pathological
methods (151). Other diagnostic
methods, such as image-based examinations, including abdominal
ultrasound examination, CT, magnetic resonance (MR) imaging or MR
scans, can be used for the partial screening of liver diseases
(152); however, these methods
are not without their own problems. For example, regarding
specificity, it is difficult to determine whether the presence of
lesions was caused by the HCV infection or not and it is hard to
accurately detect the course of HCV (153). A breakdown of the markers,
methods and their advantages and disadvantages are described in
Table I.
Despite numerous studies investigating the role of
HSCs in different models of liver fibrosis caused by various types
of disease, there remains a lack of research into the changes in
the characteristics and functions of HSCs following HCV infection.
At present, routine laboratory serological examinations,
pathological examinations and other methods are used for the
diagnosis of HCV infection; however, each technique has its
limitations. For example, considering that HCV infection is a
dynamic process, the serum indicators are unstable and patients
with varying degrees of HCV infection and courses have differing
serum marker levels. Thus, a new test is required to supplement or
replace preliminary screening, particularly in patients who are
diagnosed as positive, to assess the extent and status of HCV
infection. HSCs are a vital immune cell population activated during
HCV infection. Following an HCV infection, the expression of
specific molecular markers and chemokines or the secretion of
cytokines associated with HSCs are synchronously altered. At
present, to the best of our knowledge, there remains a lack of
studies investigating the alterations to HSC-related indicators. If
the expression of one or several of the markers are discovered to
be consistently altered during the course of HCV infection, they
may serve as a suitable marker to assess the stage of HCV infection
and they may also highlight novel avenues for understanding and
eventually treating an HCV infection.
Not applicable.
The present work was supported by the People's
Liberation Army Major Project (grant no. ANJ13J001).
Not applicable.
WW reviewed the functional characteristics of the
HSCs and the role of HSCs in the process of HCV infection; XZF
wrote the introduction and the conclusion. XLH and XZF corrected
the grammar of the manuscript; JFL reviewed the indicators for the
detection of CHC infection section; and JMY edited the figure and
table in the manuscript. All authors read and approved the final
manuscript.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
|
1
|
Wiktor S: How feasible is the global
elimination of HCV infection. Lancet. 393:1265–1267. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Spearman CW, Dusheiko GM, Hellard M and
Sonderup M: Hepatitis C. Lancet. 394:1451–1466. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Li Y, Zhao L, Geng N, Zhu W, Liu H and Bai
H: Prevalence and characteristics of hepatitis C virus infection in
Shenyang City, Northeast China, and prediction of HCV RNA
positivity according to serum anti-HCV level: Retrospective review
of hospital data. Virol J. 17:362020. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lee MH, Yang HI, Yuan Y, L'Italien G and
Chen CJ: Epidemiology and natural history of hepatitis C virus
infection. World J Gastroenterol. 20:9270–9280. 2014.PubMed/NCBI
|
|
5
|
Martinello M, Hajarizadeh B, Grebely J,
Dore GJ and Matthews GV: Management of acute HCV infection in the
era of direct-acting antiviral therapy. Nat Rev Gastroenterol
Hepatol. 15:412–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Toyoda H, Kumada T, Tada T, Mizuno K, Sone
Y, Akita T, Tanaka J and Johnson PJ: The impact of HCV eradication
by direct-acting antivirals on the transition of precancerous
hepatic nodules to HCC: A prospective observational study. Liver
Int. 39:448–454. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Shiffman ML and Benhamou Y: Cure of HCV
related liver disease. Liver Int. 35 (Suppl 1):S71–S77. 2015.
View Article : Google Scholar
|
|
8
|
Owusu Sekyere S, Schlevogt B, Mettke F,
Kabbani M, Deterding K, Wirth TC, Vogel A, Manns MP, Falk CS,
Cornberg M and Wedemeyer H: HCC immune surveillance and antiviral
therapy of hepatitis C virus infection. Liver Cancer. 8:41–65.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lin MV, King LY and Chung RT: Hepatitis C
virus-associated cancer. Annu Rev Pathol. 10:345–370. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang Y, Li J, Wang X, Sang M and Ho W:
Hepatic stellate cells, liver innate immunity, and hepatitis C
virus. J Gastroenterol Hepatol. 28 (Suppl 1):S112–S115. 2013.
View Article : Google Scholar
|
|
11
|
Kocabayoglu P, Lade A, Lee YA, Dragomir
AC, Sun X, Fiel MI, Thung S, Aloman C, Soriano P, Hoshida Y and
Friedman SL: β-PDGF receptor expressed by hepatic stellate cells
regulates fibrosis in murine liver injury, but not carcinogenesis.
J Hepatol. 63:141–147. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Cheng JC, Tseng CP, Liao MH, Peng CY, Yu
JS, Chuang PH, Huang JT and Chen JJW: Activation of hepatic
stellate cells by the ubiquitin C-terminal hydrolase 1 protein
secreted from hepatitis C virus-infected hepatocytes. Sci Rep.
7:44482017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Gieseler RK, Marquitan G, Schlattjan M,
Sowa JP, Bechmann LP, Timm J, Roggendorf M, Gerken G, Friedman SL
and Canbay A: Hepatocyte apoptotic bodies encasing nonstructural
HCV proteins amplify hepatic stellate cell activation: Implications
for chronic hepatitis C. J Viral Hepat. 18:760–767. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Saeed A, Baloch K, Brown RJ, Wallis R,
Chen L, Dexter L, McClure CP, Shakesheff K and Thomson BJ: Mannan
binding lectin-associated serine protease 1 is induced by hepatitis
C virus infection and activates human hepatic stellate cells. Clin
Exp Immunol. 174:265–273. 2013.PubMed/NCBI
|
|
15
|
El-Ahwany E, Nagy F, Zoheiry M, Shemis M,
Nosseir M, Taleb HA, El Ghannam M, Atta R and Zada S: Circulating
miRNAs as predictor markers for activation of hepatic stellate
cells and progression of HCV-induced liver fibrosis. Electron
Physician. 8:1804–1810. 2016. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Munsterman ID, Kendall TJ, Khelil N, Popa
M, Lomme R, Drenth JPH and Tjwa ETTL: Extracellular matrix
components indicate remodelling activity in different fibrosis
stages of human non-alcoholic fatty liver disease. Histopathology.
73:612–621. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Warkad SD, Nimse SB, Song KS and Kim T:
HCV detection, discrimination and genotyping technologies. Sensors
(Basel). 18:34232018. View Article : Google Scholar
|
|
18
|
Vanhommerig JW, van de Laar TJ, Koot M,
van Rooijen MS, Schinkel J, Speksnijder AG, Prins M, de Vries HJ
and Bruisten SM: Evaluation of a hepatitis C virus (HCV) antigen
assay for routine HCV screening among men who have sex with men
infected with HIV. J Virol Methods. 213:147–150. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Laperche S, Le Marrec N, Girault A,
Bouchardeau F, Servant-Delmas A, Maniez-Montreuil M, Gallian P,
Levayer T, Morel P and Simon N: Simultaneous detection of hepatitis
C virus (HCV) core antigen and anti-HCV antibodies improves the
early detection of HCV infection. J Clin Microbiol. 43:3877–3883.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Mazzola G, Adamoli L, Calvaruso V,
Macaluso FS, Colletti P, Mazzola S, Cervo A, Trizzino M, Di Lorenzo
F, Iaria C, et al: Suboptimal performance of APRI and FIB-4 in
ruling out significant fibrosis and confirming cirrhosis in HIV/HCV
co-infected and HCV mono-infected patients. Infection. 47:409–415.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sakiani S, Koh C and Heller T:
Understanding the presence of false-positive antibodies in acute
hepatitis. J Infect Dis. 210:1886–1889. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chida T, Ito M, Nakashima K, Kanegae Y,
Aoshima T, Takabayashi S, Kawata K, Nakagawa Y, Yamamoto M, Shimano
H, et al: Critical role of CREBH-mediated induction of transforming
growth factor β 2 by hepatitis C virus infection in fibrogenic
responses in hepatic stellate cells. Hepatology. 66:1430–1443.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wake K: ‘Sternzellen’ in the liver:
Perisinuosoidal cells with special reference to storage of vitamin
A. Am J Anat. 132:429–462. 1971. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
No authors listed, . Hepatic stellate cell
nomenclature. Hepatology. 23:1931996.PubMed/NCBI
|
|
25
|
Zhao W, Zhang L, Yin Z, Su W, Ren G, Zhou
C, You J, Fan J and Wang X: Activated hepatic stellate cells
promote hepatocellular carcinoma development in immunocompetent
mice. Int J Cancer. 129:2651–2661. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Higashi T, Friedman SL and Hoshida Y:
Hepatic stellate cells as key target in liver fibrosis. Adv Drug
Deliv Rev. 121:27–42. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhao W, Zhang L, Xu Y, Zhang Z, Ren G,
Tang K, Kuang P, Zhao B, Yin Z and Wang X: Hepatic stellate cells
promote tumor progression by enhancement of immunosuppressive cells
in an orthotopic liver tumor mouse model. Lab Invest. 94:182–191.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhou CL, Kong DL, Liu JF, Lu ZK, Guo HF,
Wang W, Qiu JF, Liu XJ and Wang Y: MHC II−, but not MHC
II+, hepatic stellate cells contribute to liver fibrosis
of mice in infection with schistosoma japonicum. Biochim Biophys
Acta Mol Basis Dis. 1863:1848–1857. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Najar M, Fayyad-Kazan H, Faour WH, El
Taghdouini A, Raicevic G, van Grunsven LA, Najimi M, Sokal E and
Lagneaux L: Immuno-biological comparison of hepatic stellate cells
in a reverted and activated state. Biomed Pharmacother. 98:52–62.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Bansal MB: Hepatic stellate cells:
Fibrogenic, regenerative or both? Heterogeneity and context are
key. Hepatol Int. 10:902–908. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lee UE and Friedman SL: Mechanisms of
hepatic fibrogenesis. Best Pract Res Clin Gastroenterol.
25:195–206. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Li H, Lan J, Han C, Guo K, Wang G, Hu J,
Gong J, Luo X and Cao Z: Brg1 promotes liver fibrosis via
activation of hepatic stellate cells. Exp Cell Res. 364:191–197.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Senoo H, Mezaki Y and Fujiwara M: The
stellate cell system (vitamin A-storing cell system). Anat Sci Int.
92:387–455. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Senoo H, Kojima N and Sato M: Vitamin
A-storing cells (stellate cells). Vitam Horm. 75:131–159. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Bi Y, Mukhopadhyay D, Drinane M, Ji B, Li
X, Cao S and Shah VH: Endocytosis of collagen by hepatic stellate
cells regulates extracellular matrix dynamics. Am J Physiol Cell
Physiol. 307:C622–C633. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Chen Y, Ou Y, Dong J, Yang G, Zeng Z, Liu
Y, Liu B, Li W, He X and Lan T: Osteopontin promotes collagen I
synthesis in hepatic stellate cells by miRNA-129-5p inhibition. Exp
Cell Res. 362:343–348. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wang C, Yang S, Huang J, Chen S, Li Y and
Li Q: Activation of corticotropin releasing factor receptors up
regulates collagen production by hepatic stellate cells via
promoting p300 expression. Biol Chem. 397:437–444. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Testino G, Leone S, Fagoonee S and
Pellicano R: Alcoholic liver fibrosis: Detection and treatment.
Minerva Med. 109:457–471. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Malagnino V, Bottero J, Miailhes P,
Lascoux-Combe C, Girard PM, Zoulim F, Lacombe K and Boyd A:
Hepatitis B virus genotype G and liver fibrosis progression in
chronic hepatitis B and human immunodeficiency virus coinfection. J
Med Virol. 91:630–641. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Chung SI, Moon H, Ju HL, Cho KJ, Kim DY,
Han KH, Eun JW, Nam SW, Ribback S, Dombrowski F, et al: Hepatic
expression of sonic hedgehog induces liver fibrosis and promotes
hepatocarcinogenesis in a transgenic mouse model. J Hepatol.
64:618–627. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Tsuchida T and Friedman SL: Mechanisms of
hepatic stellate cell activation. Nat Rev Gastroenterol Hepatol.
14:397–411. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Pradere JP, Kluwe J, De Minicis S, Jiao
JJ, Gwak GY, Dapito DH, Jang MK, Guenther ND, Mederacke I, Friedman
R, et al: Hepatic macrophages but not dendritic cells contribute to
liver fibrosis by promoting the survival of activated hepatic
stellate cells in mice. Hepatology. 58:1461–1473. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Jin H, Jia Y, Yao Z, Huang J, Hao M, Yao
S, Lian N, Zhang F, Zhang C, Chen X, et al: Hepatic stellate cell
interferes with NK cell regulation of fibrogenesis via curcumin
induced senescence of hepatic stellate cell. Cell Signal. 33:79–85.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Li X, Su Y, Hua X, Xie C, Liu J, Huang Y,
Zhou L, Zhang M, Li X and Gao Z: Levels of hepatic Th17 cells and
regulatory T cells upregulated by hepatic stellate cells in
advanced HBV-related liver fibrosis. J Transl Med. 15:752017.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Corpechot C, Barbu V, Wendum D, Kinnman N,
Rey C, Poupon R, Housset C and Rosmorduc O: Hypoxia-induced VEGF
and collagen I expressions are associated with angiogenesis and
fibrogenesis in experimental cirrhosis. Hepatology. 35:1010–1021.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Hong IH, Park SJ, Goo MJ, Lee HR, Park JK,
Ki MR, Kim SH, Lee EM, Kim AY and Jeong KS: JNK1 and JNK2 regulate
α-SMA in hepatic stellate cells during CCl4-induced fibrosis in the
rat liver. Pathol Int. 63:483–491. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Giannandrea M and Parks WC: Diverse
functions of matrix metalloproteinases during fibrosis. Dis Model
Mech. 7:193–203. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Holm Nielsen S, Willumsen N, Leeming DJ,
Daniels SJ, Brix S, Karsdal MA, Genovese F and Nielsen MJ:
Serological assessment of activated fibroblasts by alpha-smooth
muscle actin (α-SMA): A noninvasive biomarker of activated
fibroblasts in lung disorders. Transl Oncol. 12:368–374. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Elzamly S, Agina HA, Elbalshy AE,
Abuhashim M, Saad E and Abd Elmageed ZY: Integration of VEGF and
α-SMA expression improves the prediction accuracy of fibrosis in
chronic hepatitis C liver biopsy. Appl Immunohistochem Mol Morphol.
25:261–270. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Stefanovic L and Stefanovic B: Role of
cytokine receptor-like factor 1 in hepatic stellate cells and
fibrosis. World J Hepatol. 4:356–364. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Latoche JD, Ufelle AC, Fazzi F, Ganguly K,
Leikauf GD and Fattman CL: Secreted phosphoprotein 1 and
sex-specific differences in silica-induced pulmonary fibrosis in
mice. Environ Health Perspect. 124:1199–1207. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Kumar P, Smith T, Raeman R, Chopyk DM,
Brink H, Liu Y, Sulchek T and Anania FA: Periostin promotes liver
fibrogenesis by activating lysyl oxidase in hepatic stellate cells.
J Biol Chem. 293:12781–12792. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Dongiovanni P, Meroni M, Baselli GA,
Bassani GA, Rametta R, Pietrelli A, Maggioni M, Facciotti F, Trunzo
V, Badiali S, et al: Insulin resistance promotes lysyl oxidase like
2 induction and fibrosis accumulation in non-alcoholic fatty liver
disease. Clin Sci (Lond). 131:1301–1315. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Zhang SC, Zheng YH, Yu PP, Min TH, Yu FX,
Ye C, Xie YK and Zhang QY: Lentiviral vector-mediated
down-regulation of IL-17A receptor in hepatic stellate cells
results in decreased secretion of IL-6. World J Gastroenterol.
18:3696–3704. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Ehling J and Tacke F: Role of chemokine
pathways in hepatobiliary cancer. Cancer Lett. 379:173–183. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Kim BM, Abdelfattah AM, Vasan R, Fuchs BC
and Choi MY: Hepatic stellate cells secrete Ccl5 to induce
hepatocyte steatosis. Sci Rep. 8:74992018. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Li Z, Zhang Q, Zhang Q, Xu M, Qu Y, Cai X
and Lu L: CXCL6 promotes human hepatocyte proliferation through the
CXCR1-NFkB pathway and inhibits collagen I secretion by hepatic
stellate cells. Biochem Cell Biol. 94:229–235. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Puche JE, Saiman Y and Friedman SL:
Hepatic stellate cells and liver fibrosis. Compr Physiol.
3:1473–1492. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Ma PF, Gao CC, Yi J, Zhao JL, Liang SQ,
Zhao Y, Ye YC, Bai J, Zheng QJ, Dou KF, et al: Cytotherapy with
M1-polarized macrophages ameliorates liver fibrosis by modulating
immune microenvironment in mice. J Hepatol. 67:770–779. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Sasaki R, Devhare PB, Steele R, Ray R and
Ray RB: Hepatitis C virus-induced CCL5 secretion from macrophages
activates hepatic stellate cells. Hepatology. 66:746–757. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Höchst B, Schildberg FA, Sauerborn P,
Gäbel YA, Gevensleben H, Goltz D, Heukamp LC, Türler A, Ballmaier
M, Gieseke F, et al: Activated human hepatic stellate cells induce
myeloid derived suppressor cells from peripheral blood monocytes in
a CD44-dependent fashion. J Hepatol. 59:528–535. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Wang B, Trippler M, Pei R, Lu M, Broering
R, Gerken G and Schlaak JF: Toll-like receptor activated human and
murine hepatic stellate cells are potent regulators of hepatitis C
virus replication. J Hepatol. 51:1037–1045. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Jeong WI, Park O, Suh YG, Byun JS, Park
SY, Choi E, Kim JK, Ko H, Wang H, Miller AM and Gao B: Suppression
of innate immunity (natural killer cell/interferon-γ) in the
advanced stages of liver fibrosis in mice. Hepatology.
53:1342–1351. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Radaeva S, Wang L, Radaev S, Jeong WI,
Park O and Gao B: Retinoic acid signaling sensitizes hepatic
stellate cells to NK cell killing via upregulation of NK cell
activating ligand RAE1. Am J Physiol Gastrointest Liver Physiol.
293:G809–G816. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Langhans B, Alwan AW, Krämer B, Glässner
A, Lutz P, Strassburg CP, Nattermann J and Spengler U: Regulatory
CD4+T cells modulate the interaction between NK cells
and hepatic stellate cells by acting on either cell type. J
Hepatol. 62:398–404. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Li Y, Lu L, Qian S, Fung JJ and Lin F:
Hepatic stellate cells directly inhibit B cells via programmed
death-ligand 1. J Immunol. 196:1617–1625. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Bedossa P and Paradis V: Approaches for
treatment of liver fibrosis in chronic hepatitis C. Clin Liver Dis.
7:195–210. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Ignat SR, Dinescu S, Hermenean A and
Costache M: Cellular interplay as a consequence of inflammatory
signals leading to liver fibrosis development. Cells. 9:4612020.
View Article : Google Scholar
|
|
69
|
Shahin K, Hosseini SY, Jamali H, Karimi
MH, Azarpira N and Zeraatian M: The enhancing impact of amino
termini of hepatitis C virus core protein on activation of hepatic
stellate cells. Gastroenterol Hepatol Bed Bench. 13:57–63.
2020.PubMed/NCBI
|
|
70
|
Wang L, Wang Y and Quan J: Exosomes
derived from natural killer cells inhibit hepatic stellate cell
activation and liver fibrosis. Hum Cell. 33:582–589. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Cai B, Dongiovanni P, Corey KE, Wang X,
Shmarakov IO, Zheng Z, Kasikara C, Davra V, Meroni M, Chung RT, et
al: Macrophage MerTK promotes liver fibrosis in nonalcoholic
steatohepatitis. Cell Metab. 31:406–421. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Tsukamoto H: Cytokine regulation of
hepatic stellate cells in liver fibrosis. Alcohol Clin Exp Res.
23:911–916. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Wang S, Li M, Zhao X, Wang H, Zhu J, Wang
C, Zhou M, Dong H and Zhou R: Upregulation of KSRP by miR-27b
attenuates schistosomiasis-induced hepatic fibrosis by targeting
TGF-β1. FASEB J. 34:4120–4133. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Huang JL, Fu YP, Gan W, Liu G, Zhou PY,
Zhou C, Sun BY, Guan RY, Zhou J, Fan J, et al: Hepatic stellate
cells promote the progression of hepatocellular carcinoma through
microRNA-1246-RORα-Wnt/β-Catenin axis. Cancer Lett. 476:140–151.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Winkler I, Bitter C, Winkler S, Weichenhan
D, Thavamani A, Hengstler JG, Borkham-Kamphorst E, Kohlbacher O,
Plass C, Geffers R, et al: Identification of Pparγ-modulated miRNA
hubs that target the fibrotic tumor microenvironment. Proc Natl
Acad Sci USA. 117:454–463. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Dawood RM, El-Meguid MA, Ibrahim MK, Bader
El Din NG, Barakat A, El-Wakeel K, Alla MDAA, Wu GY and El Awady
MK: Dysregulation of fibrosis related genes in HCV induced liver
disease. Gene. 664:58–69. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Chouteau P, Defer N, Florimond A,
Caldéraro J, Higgs M, Gaudin A, Mérour E, Dhumeaux D, Lerat H and
Pawlotsky JM: Hepatitis C virus (HCV) protein expression enhances
hepatic fibrosis in HCV transgenic mice exposed to a fibrogenic
agent. J Hepatol. 57:499–507. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Mazzocca A, Sciammetta SC, Carloni V,
Cosmi L, Annunziato F, Harada T, Abrignani S and Pinzani M: Binding
of hepatitis C virus envelope protein E2 to CD81 up-regulates
matrix metalloproteinase-2 in human hepatic stellate cells. J Biol
Chem. 280:11329–11339. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Bataller R, Paik YH, Lindquist JN,
Lemasters JJ and Brenner DA: Hepatitis C virus core and
nonstructural proteins induce fibrogenic effects in hepatic
stellate cells. Gastroenterology. 126:529–540. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Coenen M, Nischalke HD, Krämer B, Langhans
B, Glässner A, Schulte D, Körner C, Sauerbruch T, Nattermann J and
Spengler U: Hepatitis C virus core protein induces fibrogenic
actions of hepatic stellate cells via toll-like receptor 2. Lab
Invest. 91:1375–1382. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Charrier A, Chen R, Chen L, Kemper S,
Hattori T, Takigawa M and Brigstock DR: Exosomes mediate
intercellular transfer of pro-fibrogenic connective tissue growth
factor (CCN2) between hepatic stellate cells, the principal
fibrotic cells in the liver. Surgery. 156:548–555. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Li M, Jiang M, Meng J and Tao L: Exosomes:
Carriers of pro-fibrotic signals and therapeutic targets in
fibrosis. Curr Pharm Des. 25:4496–4509. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Kim JH, Lee CH and Lee SW: Exosomal
transmission of MicroRNA from HCV replicating cells stimulates
transdifferentiation in hepatic stellate cells. Mol Ther Nucleic
Acids. 14:483–497. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Khatun M and Ray RB: Mechanisms underlying
hepatitis C virus-associated hepatic fibrosis. Cells. 8:12492019.
View Article : Google Scholar
|
|
85
|
Devhare PB, Sasaki R, Shrivastava S, Di
Bisceglie AM, Ray R and Ray RB: Exosome-mediated intercellular
communication between hepatitis C virus-infected hepatocytes and
hepatic stellate cells. J Virol. 91:e02225–e02216. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Njiomegnie GF, Read SA, Fewings N, George
J, McKay F and Ahlenstiel G: Immunomodulation of the natural killer
cell phenotype and response during HCV infection. J Clin Med.
9:10302020. View Article : Google Scholar
|
|
87
|
Glässner A, Eisenhardt M, Krämer B, Körner
C, Coenen M, Sauerbruch T, Spengler U and Nattermann J: NK cells
from HCV-infected patients effectively induce apoptosis of
activated primary human hepatic stellate cells in a TRAIL-, FasL-
and NKG2D-dependent manner. Lab Invest. 92:967–977. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Wang H and Yin S: Natural killer T cells
in liver injury, inflammation and cancer. Expert Rev Gastroenterol
Hepatol. 9:1077–1085. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
de Lalla C, Galli G, Aldrighetti L, Romeo
R, Mariani M, Monno A, Nuti S, Colombo M, Callea F, Porcelli SA, et
al: Production of profibrotic cytokines by invariant NKT cells
characterizes cirrhosis progression in chronic viral hepatitis. J
Immunol. 173:1417–1425. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Locati M, Curtale G and Mantovani A:
Diversity, mechanisms, and significance of macrophage plasticity.
Annu Rev Pathol. 15:123–147. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Suzuki K, Meguro K, Nakagomi D and
Nakajima H: Roles of alternatively activated M2 macrophages in
allergic contact dermatitis. Allergol Int. 66:392–397. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Brass A and Brenndörfer ED: The role of
chemokines in hepatitis C virus-mediated liver disease. Int J Mol
Sci. 15:4747–4779. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Wasmuth HE and Weiskirchen R: Pathogenesis
of liver fibrosis: Modulation of stellate cells by chemokines. Z
Gastroenterol. 48:38–45. 2010.(In German). View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Liang YJ, Luo J, Lu Q, Zhou Y, Wu HW,
Zheng D, Ren YY, Sun KY, Wang Y and Zhang ZS: Gene profile of
chemokines on hepatic stellate cells of schistosome-infected mice
and antifibrotic roles of CXCL9/10 on liver non-parenchymal cells.
PLoS One. 7:e424902012. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Marra F and Tacke F: Roles for chemokines
in liver disease. Gastroenterology. 147:577–594. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Tan HX, Gong WZ, Zhou K, Xiao ZG, Hou FT,
Huang T, Zhang L, Dong HY, Zhang WL, Liu Y and Huang ZC:
CXCR4/TGF-β1 mediated hepatic stellate cells differentiation into
carcinoma-associated fibroblasts and promoted liver metastasis of
colon cancer. Cancer Biol Ther. 21:258–268. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Ferrari SM, Fallahi P, Ruffilli I, Elia G,
Ragusa F, Paparo SR, Patrizio A, Mazzi V, Colaci M, Giuggioli D, et
al: Immunomodulation of CXCL10 secretion by hepatitis C virus:
Could CXCL10 Be a prognostic marker of chronic hepatitis C? J
Immunol Res. 2019:58789602019. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Pineda-Tenor D, Berenguer J, Jiménez-Sousa
MA, Guzmán-Fulgencio M, Aldámiz-Echevarria T, Carrero A,
García-Álvarez M, Diez C, Tejerina F, Briz V and Resino S: CXCL9,
CXCL10 and CXCL11 polymorphisms are associated with sustained
virologic response in HIV/HCV-coinfected patients. J Clin Virol.
61:423–429. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Liu Y, Chen L, Zou Z, Zhu B, Hu Z, Zeng P,
Wu L and Xiong J: Hepatitis C virus infection induces elevation of
CXCL10 in human brain microvascular endothelial cells. J Med Virol.
88:1596–1603. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Zeremski M, Dimova R, Astemborski J,
Thomas DL and Talal AH: CXCL9 and CXCL10 chemokines as predictors
of liver fibrosis in a cohort of primarily African-American
injection drug users with chronic hepatitis C. J Infect Dis.
204:832–836. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Sahin H, Borkham-Kamphorst E, Kuppe C,
Zaldivar MM, Grouls C, Al-samman M, Nellen A, Schmitz P, Heinrichs
D, Berres ML, et al: Chemokine Cxcl9 attenuates liver
fibrosis-associated angiogenesis in mice. Hepatology. 55:1610–1619.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Joshi D, Carey I, Foxton M, Al-Freah M,
Bruce M, Heaton N, Quaglia A, O'Grady J, Aluvihare V and Agarwal K:
CXCL10 levels identify individuals with rapid fibrosis at 12 months
post-transplant for hepatitis C virus and predict treatment
response. Clin Transplant. 28:569–578. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Gorin JB, Malone DFG, Strunz B, Carlsson
T, Aleman S, Björkström NK, Falconer K and Sandberg JK: Plasma
FABP4 is associated with liver disease recovery during
treatment-induced clearance of chronic HCV infection. Sci Rep.
10:20812020. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Lu Y, Lin LY, Tan JG, Deng HP, Li XH,
Zhang Z, Li Y, Zhou Z, Xu X, Xie X and Mei SJ: A correlation study
between gene polymorphism of Th cell expressed chemokine receptor
CXCR3 and its ligand levels with HCV infection prognosis. Eur Rev
Med Pharmacol Sci. 21:1290–1295. 2017.PubMed/NCBI
|
|
105
|
Berres ML, Asmacher S, Lehmann J, Jansen
C, Görtzen J, Klein S, Meyer C, Strunk HM, Fimmers R, Tacke F, et
al: CXCL9 is a prognostic marker in patients with liver cirrhosis
receiving transjugular intrahepatic portosystemic shunt. J Hepatol.
62:332–339. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Chalin A, Lefevre B, Devisme C, Barget N,
Amiot L and Samson M: Circulating levels of CXCL11 and CXCL12 are
biomarkers of cirrhosis in patients with chronic hepatitis C
infection. Cytokine. 117:72–78. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Patidar M, Yadav N and Dalai SK:
Interleukin 15: A key cytokine for immunotherapy. Cytokine Growth
Factor Rev. 31:49–59. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Jiao J, Ooka K, Fey H, Fiel MI, Rahmman
AH, Kojima K, Hoshida Y, Chen X, de Paula T, Vetter D, et al:
Interleukin-15 receptor α on hepatic stellate cells regulates
hepatic fibrogenesis in mice. J Hepatol. 65:344–353. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Golden-Mason L, Kelly AM, Doherty DG,
Traynor O, McEntee G, Kelly J, Hegarty JE and O'Farrelly C: Hepatic
interleuklin 15 (IL-15) expression: Implications for local NK/NKT
cell homeostasis and development. Clin Exp Immunol. 138:94–101.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Vahedi F, Lee AJ, Collins SE, Chew MV,
Lusty E, Chen B, Dubey A, Richards CD, Feld JJ, Russell RS, et al:
IL-15 and IFN-γ signal through the ERK pathway to inhibit HCV
replication, independent of type I IFN signaling. Cytokine.
124:1544392019. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Isailovic N, Daigo K, Mantovani A and
Selmi C: Interleukin-17 and innate immunity in infections and
chronic inflammation. J Autoimmun. 60:1–11. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Abou El-Khier NT, Elhammady D, Arafa MM,
Shahin D, Eladl E, Abousamra NK, Sharaf-Eldeen O, Shaker G and
Esmael ME: Th17 and IL-17 as predictors of hepatic inflammation in
patients with chronic hepatitis C virus infection and treated with
direct antiviral therapy. Egypt J Immunol. 25:61–74.
2018.PubMed/NCBI
|
|
113
|
Gu Y, Hu X, Liu C, Qv X and Xu C:
Interleukin (IL)-17 promotes macrophages to produce IL-8, IL-6 and
tumour necrosis factor-alpha in aplastic anaemia. Br J Haematol.
142:109–114. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Zhang Y, Huang D, Gao W, Yan J, Zhou W,
Hou X, Liu M, Ren C, Wang S and Shen J: Lack of IL-17 signaling
decreases liver fibrosis in murine schistosomiasis japonica. Int
Immunol. 27:317–325. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Amara S, Lopez K, Banan B, Brown SK,
Whalen M, Myles E, Ivy MT, Johnson T, Schey KL and Tiriveedhi V:
Synergistic effect of pro-inflammatory TNFα and IL-17 in periostin
mediated collagen deposition: Potential role in liver fibrosis. Mol
Immunol. 64:26–35. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Elkhawaga AA, Hosni A, Zaky DZ, Kamel AA,
Mohamed NA, Abozaid MA and El-Masry MA: Association of treg and
TH17 cytokines with HCV pathogenesis and liver pathology. Egypt J
Immunol. 26:55–63. 2019.PubMed/NCBI
|
|
117
|
Zhang LJ, Yu JP, Li D, Huang YH, Chen ZX
and Wang XZ: Effects of cytokines on carbon tetrachloride-induced
hepatic fibrogenesis in rats. World J Gastroenterol. 10:77–81.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Niess JH and Francés R: Editorial: The
IL-20 cytokines and related family members in immunity and
diseases. Front Immunol. 10:19762019. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Chiu YS, Wei CC, Lin YJ, Hsu YH and Chang
MS: IL-20 and IL-20R1 antibodies protect against liver fibrosis.
Hepatology. 60:1003–1014. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Rutz S, Eidenschenk C and Ouyang W: IL-22,
not simply a Th17 cytokine. Immunol Rev. 252:116–132. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Kronenberger B, Rudloff I, Bachmann M,
Brunner F, Kapper L, Filmann N, Waidmann O, Herrmann E,
Pfeilschifter J, Zeuzem S, et al: Interleukin-22 predicts severity
and death in advanced liver cirrhosis: A prospective cohort study.
BMC Med. 10:1022012. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Sertorio M, Hou X, Carmo RF, Dessein H,
Cabantous S, Abdelwahed M, Romano A, Albuquerque F, Vasconcelos L,
Carmo T, et al: IL-22 and IL-22 binding protein (IL-22BP) regulate
fibrosis and cirrhosis in hepatitis C virus and schistosome
infections. Hepatology. 61:1321–1331. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Kong X, Feng D, Wang H, Hong F, Bertola A,
Wang FS and Gao B: Interleukin-22 induces hepatic stellate cell
senescence and restricts liver fibrosis in mice. Hepatology.
56:1150–1159. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Lu DH, Guo XY, Qin SY, Luo W, Huang XL,
Chen M, Wang JX, Ma SJ, Yang XW and Jiang HX: Interleukin-22
ameliorates liver fibrogenesis by attenuating hepatic stellate cell
activation and downregulating the levels of inflammatory cytokines.
World J Gastroenterol. 21:1531–1545. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Zhao Z, Lin CY and Cheng K: siRNA- and
miRNA-based therapeutics for liver fibrosis. Transl Res. 214:17–29.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Shaker OG and Senousy MA: Serum microRNAs
as predictors for liver fibrosis staging in hepatitis C
virus-associated chronic liver disease patients. J Viral Hepat.
24:636–644. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Coll M, El Taghdouini A, Perea L,
Mannaerts I, Vila-Casadesús M, Blaya D, Rodrigo-Torres D, Affò S,
Morales-Ibanez O, Graupera I, et al: Integrative miRNA and gene
expression profiling analysis of human quiescent hepatic stellate
cells. Sci Rep. 5:115492015. View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Marquez RT, Bandyopadhyay S, Wendlandt EB,
Keck K, Hoffer BA, Icardi MS, Christensen RN, Schmidt WN and
McCaffrey AP: Correlation between microRNA expression levels and
clinical parameters associated with chronic hepatitis C viral
infection in humans. Lab Invest. 90:1727–1736. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Appourchaux K, Dokmak S, Resche-Rigon M,
Treton X, Lapalus M, Gattolliat CH, Porchet E, Martinot-Peignoux M,
Boyer N, Vidaud M, et al: MicroRNA-based diagnostic tools for
advanced fibrosis and cirrhosis in patients with chronic hepatitis
B and C. Sci Rep. 6:349352016. View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Ogawa T, Enomoto M, Fujii H, Sekiya Y,
Yoshizato K, Ikeda K and Kawada N: MicroRNA-221/222 upregulation
indicates the activation of stellate cells and the progression of
liver fibrosis. Gut. 61:1600–1609. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Chen L, Charrier A, Zhou Y, Chen R, Yu B,
Agarwal K, Tsukamoto H, Lee LJ, Paulaitis ME and Brigstock DR:
Epigenetic regulation of connective tissue growth factor by
MicroRNA-214 delivery in exosomes from mouse or human hepatic
stellate cells. Hepatology. 59:1118–1129. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Guo CJ, Pan Q, Li DG, Sun H and Liu BW:
miR-15b and miR-16 are implicated in activation of the rat hepatic
stellate cell: An essential role for apoptosis. J Hepatol.
50:766–778. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
133
|
Li J, Ghazwani M, Zhang Y, Lu J, Li J, Fan
J, Gandhi CR and Li S: miR-122 regulates collagen production via
targeting hepatic stellate cells and suppressing P4HA1 expression.
J Hepatol. 58:522–528. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Zheng J, Lin Z, Dong P, Lu Z, Gao S, Chen
X, Wu C and Yu F: Activation of hepatic stellate cells is
suppressed by microRNA-150. Int J Mol Med. 32:17–24. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
135
|
Leask A: CCN2/decorin interactions: A
novel approach to combating fibrosis? J Cell Commun Signal.
5:249–250. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
136
|
Yue ZH, Xia CS and Wang H: Performance
evaluation of the mindray anti-HCV assay for the detection of
hepatitis C virus infection. J Clin Lab Anal. 32:e226002018.
View Article : Google Scholar : PubMed/NCBI
|
|
137
|
Wasitthankasem R, Vongpunsawad S, Siripon
N, Suya C, Chulothok P, Chaiear K, Rujirojindakul P, Kanjana S,
Theamboonlers A, Tangkijvanich P and Poovorawan Y: Genotypic
distribution of hepatitis C virus in Thailand and Southeast Asia.
PLoS One. 10:e01267642015. View Article : Google Scholar : PubMed/NCBI
|
|
138
|
He J, Xiu B, Wang G, Chen K, Feng X, Song
X, Zhu C, Ling S and Zhang H: Double-antigen sandwich ELISA for the
detection of anti-hepatitis C virus antibodies. J Virol Methods.
171:163–168. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
139
|
Kim S, Kim JH, Yoon S, Park YH and Kim HS:
Clinical performance evaluation of four automated chemiluminescence
immunoassays for hepatitis C virus antibody detection. J Clin
Microbiol. 46:3919–3923. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
140
|
Heinrichs A, Antoine M, Steensels D,
Montesinos I and Delforge ML: HCV false positive immunoassays in
patients with LVAD: A potential trap! J Clin Virol. 78:44–46. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
141
|
Ali A and Lal A: False positivity of
serological tests for hepatitis C virus. J Ayub Med Coll
Abbottabad. 22:43–45. 2010.PubMed/NCBI
|
|
142
|
Vo MT, Bruhn R, Kaidarova Z, Custer BS,
Murphy EL and Bloch EM: A retrospective analysis of false-positive
infectious screening results in blood donors. Transfusion.
56:457–465. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
143
|
Moorman AC, Drobenuic J and Kamili S:
Prevalence of false-positive hepatitis C antibody results, national
health and nutrition examination study (NHANES) 2007–2012. J Clin
Virol. 89:1–4. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
144
|
Scott JD and Gretch DR: Molecular
diagnostics of hepatitis C virus infection: A systematic review.
JAMA. 297:724–732. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
145
|
Safic Stanic H, Babic I, Maslovic M, Dogic
V, Bingulac-Popovic J, Miletic M, Jurakovic-Loncar N, Vuk T,
Strauss-Patko M and Jukic I: Three-year experience in NAT screening
of blood donors for transfusion transmitted viruses in croatia.
Transfus Med Hemother. 44:415–420. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
146
|
Bustin SA: Quantification of mRNA using
real-time reverse transcription PCR (RT-PCR): Trends and problems.
J Mol Endocrinol. 29:23–39. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
147
|
Papadopoulos N, Vasileiadi S, Papavdi M,
Sveroni E, Antonakaki P, Dellaporta E, Koutli E, Michalea S,
Manolakopoulos S, Koskinas J and Deutsch M: Liver fibrosis staging
with combination of APRI and FIB-4 scoring systems in chronic
hepatitis C as an alternative to transient elastography. Ann
Gastroenterol. 32:498–503. 2019.PubMed/NCBI
|
|
148
|
Kim WR, Berg T, Asselah T, Flisiak R, Fung
S, Gordon SC, Janssen HL, Lampertico P, Lau D, Bornstein JD, et al:
Evaluation of APRI and FIB-4 scoring systems for non-invasive
assessment of hepatic fibrosis in chronic hepatitis B patients. J
Hepatol. 64:773–780. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
149
|
Sagnelli C, Uberti-Foppa C, Pasquale G, De
Pascalis S, Coppola N, Albarello L, Doglioni C, Lazzarin A and
Sagnelli E: Factors influencing liver fibrosis and
necroinflammation in HIV/HCV coinfection and HCV monoinfection.
Infection. 41:959–967. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
150
|
Bedossa P and Patel K: Biopsy and
noninvasive methods to assess progression of nonalcoholic fatty
liver disease. Gastroenterology. 150:1811–1822. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
151
|
Francque SM, De Pauw FF, Van den Steen GH,
Van Marck EA, Pelckmans PA and Michielsen PP: Biopsy of focal liver
lesions: Guidelines, comparison of techniques and cost-analysis.
Acta Gastroenterol Belg. 66:160–165. 2003.PubMed/NCBI
|
|
152
|
Childers RE and Ahn J: Diagnosis of
alcoholic liver disease: Key foundations and new developments. Clin
Liver Dis. 20:457–471. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
153
|
Colli A, Fraquelli M, Andreoletti M,
Marino B, Zuccoli E and Conte D: Severe liver fibrosis or
cirrhosis: Accuracy of US for detection-analysis of 300 cases.
Radiology. 227:89–94. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
154
|
Shiha G, Seif S, Eldesoky A, Elbasiony M,
Soliman R, Metwally A, Zalata K and Mikhail N: A simple bedside
blood test (Fibrofast; FIB-5) is superior to FIB-4 index for the
differentiation between non-significant and significant fibrosis in
patients with chronic hepatitis C. Hepatol Int. 11:286–291. 2017.
View Article : Google Scholar : PubMed/NCBI
|