Introduction
Acute myocardial infarction (AMI) is caused by acute
and persistent ischemia, as well as hypoxia of the coronary artery
(1). The mortality of AMI in China
has increased to 1 million per year in the past two decades based
on an analysis of hospital data from 1990 to 2010 (2). Therefore, it is necessary to explore
the regulatory mechanism underlying AMI development for the
improvement of therapeutic strategies.
Long non-coding RNAs (lncRNAs) are a form of long
RNAs (>200 nucleotides) with no translation ability but an
ability to affect gene expression at the transcriptional level
(3). A number of studies have
demonstrated that lncRNAs are associated with the development of
AMI. For example, lncRNA metastasis associated lung adenocarcinoma
transcript 1 facilitated AMI development via the
miR-320/phosphatase and tensin homolog axis (4). In addition, another study revealed
that hypoxia stimulated the expression of Testis-Specific
Transcript, Y-Linked 15 (TTTY-15), and TTTY-15 knockdown refrained
hypoxia-induced injury (5).
Moreover, it has been reported that maternally expressed 3 (MEG3)
is associated with AMI development (6,7);
however, the regulatory mechanism underlying MEG3 in AMI is not
completely understood.
MicroRNAs (miRNAs/miRs), a type of small RNAs (~22
nucleotides) without translation capacity, negatively regulate
target genes expression by mediating mRNA degradation or repressing
mRNA translation (8). A number of
miRNAs have been identified to be involved in AMI development. For
example, Chen et al (9)
demonstrated that miR-133 improved cardiac injury in AMI. In
addition, another study indicated that miR-145 downregulation in
plasma was related to AMI (10) and
miR-325-3p was also reported to participate in the development of
AMI (11).
Transient receptor potential cation channel
subfamily V member 4 (TRPV4) is a gene locus on human chromosome
12q24.11, which encodes a nonselective cation channel protein that
is important for systemic osmotic pressure (12). Dysregulation of TRPV4 has been
reported in AMI (13). However, the
regulatory mechanism underlying miR-325-3p and TRPV4 in AMI has not
been previously reported. Therefore, the present study investigated
the functions and mechanisms underlying MEG3 in hypoxia-induced
injury.
Materials and methods
Cell culture and treatment
The rat cardiomyocyte cell line (H9c2) was purchased
from Procell Life Science & Technology Co., Ltd. and cultured
in DMEM (Beijing Solarbio Science & Technology Co., Ltd.)
supplemented with 10% FBS (Genetimes Technology, Inc.) in an
incubator at 37°C. H9c2 cells were divided into two groups: i)
Normoxia group (cultured with 95% air and 5% CO2); and
ii) hypoxia group (cultured with 94.5% N2, 5%
CO2 and 0.5% O2).
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA from H9c2 cells was extracted using
TriQuick Reagent (Beijing Solarbio Science & Technology Co.,
Ltd.). Reverse transcription for lncRNAs and mRNAs was performed
using AMV Reverse Transcriptase (Beijing Solarbio Science &
Technology Co., Ltd.), whereas reverse transcription for miRNA was
performed using the TaqMan miRNA kit (Applied Biosystems; Thermo
Fisher Scientific, Inc.). Total RNA (1 µg), 1 µl Oligo (dT), 0.5 µl
dNTPs, 2 µl 5 X reaction buffer, 1 µl AMV Reverse Transcriptase and
4.5 DEPC H2O was mixed and followed by reverse
transcription (42°C for 60 min). Subsequently, qPCR was performed
using SYBR® Premix Ex Taq II (Takara Biotechnology Co.,
Ltd.). The following parameters were used for qPCR: 1 cycle at 98°C
for 3 min, followed by 40 cycles for 15 sec at 94°C, for 30 sec at
60°C, and for 1 min at 72°C. The expression of mRNA and miRNA were
quantified using the 2−ΔΔCq method (14) and normalized to the internal
reference genes β-actin and U6, respectively. The following primers
(synthesized by BGI Group) were used for qPCR: HIF1α forward,
5′-CTGACCCTGCACTCAATCAAG-3′ and reverse,
5′-TGGGACTATTAGGCTCAGGTG-3′; MEG3 forward,
5′-GGCAGGATCTGGCATAGAGG-3′ and reverse,
5′-CGAGTCAGGAAGCAGTGGGTT-3′; miR-325-3p forward,
5′-GCCAGCACCTTCACAAAGTAGC and reverse, 5′-CCATGCTAGACAACAGCTCTG-3′;
TRPV4 forward, 5′-CCCGAGAGAACACCAAGTTTG-3′ and reverse,
5′-GACCGTCATTGTTAAGCACAGTCT-3′; β-actin forward,
5′-CGTTGACATCCGTAAAGACC-3′ and reverse, 5′-TAGAGCCACCAATCCACACA-3′;
and U6 forward, 5′-CTCGCTTCGGCAGCACA-3′ and reverse,
5′-AACGCTTCACGAATTTGCGT-3′.
Western blotting
Total protein was extracted from H9c2 cells using a
protein extraction kit (Beyotime Institute of Biotechnology). Total
protein was quantified using a bicinchoninic protein assay kit
(Beyotime Institute of Biotechnology). The protein samples (35 µg
per lane) were separated via 10% SDS-PAGE and transferred onto PVDF
membranes (EMD Millipore). The membranes were blocked in 5% skimmed
milk for 4 h at 37°C, and incubated for 12 h at 4°C with the
following primary antibodies (all purchased from Abcam): HIF1α
(cat. no. ab51608, 1:400), Bcl-2 (cat. no. ab196495, 1:1,000), Bax
(cat. no. ab104156, 1:1,000), cleaved caspase-3 (cat. no. ab2302,
1:1,000), TRPV4 (cat. no. ab39260, 1:1,000) and β-actin (cat. no.
ab8227, 1:1,000). Subsequently, the membranes were incubated with a
secondary antibody goat anti-rabbit IgG H&L (HRP) (cat. no.
ab205718, 1:20,000; Abcam) for 2 h at 37°C. Protein bands were
visualized using an ECL kit (Beyotime Institute of Biotechnology).
β-actin was used as the loading control. Quantification of band
intensities by densitometry was performed using the ImageJ software
version (V1.8.0._172; National Institutes of Health).
Cell transfection
The small interfering siRNA targeted against MEG3
(si-MEG3; 5′-GCTTCTCGAGGCCTGTCTATT-3′), siRNA targeted against
TRPV4 (si-TRPV4; 5′-CGUGUCCUUCUACAUCAATT-3′), negative control (NC)
siRNA (si-NC), miR-325-3p mimic (miR-325-3p), mimic NC (miR-NC),
miR-325-3p inhibitor (anti-miR-325-3p) and inhibitor NC
(anti-miR-NC) were purchased from Shanghai GeneChem Co., Ltd. The
fragment of MEG3 was inserted into the pcDNA vector to construct
the MEG3 overexpression plasmid (MEG3). The blank load pcDNA vector
was used as the negative control. Cells (4×105
cells/well) were transfected with siRNA (10 nM at final
concentration), mimic (25 nM at final concentration), inhibitor (50
nM at final concentration), overexpression plasmid (4 µg) or equal
amounts of NCs using Lipofectamine® 2000 (Invitrogen;
Thermo Fisher Scientific, Inc.). Cells were replaced with fresh
medium 6 h after transfection before the hypoxia group received
hypoxia treatment for 24 h after transfection for subsequent
experiments.
MTT assay
An MTT assay (Beijing Solarbio Science &
Technology Co., Ltd.) was performed to detect H9c2 cell viability.
H9c2 cells were seeded (5×103 cells/well) into a 96-well
plate and incubated at 37°C for 24 h. Then, 24 h after
transfection, cells were incubated for 0, 24, 48 or 72 h in hypoxic
conditions. Subsequently, cells were incubated with MTT for 4 h at
37°C. DMSO was added to dissolve the formazan. The absorbance of
each well was measured at a wavelength of 490 nm using a microplate
reader.
Flow cytometry analysis of cell
apoptosis
The Annexin V/propidium iodide cell apoptosis
analysis kit (Wuhan Servicebio Technology Co., Ltd.) was utilized
to assess H9c2 cell apoptosis (early + late apoptosis). Briefly,
H9c2 cells (1×106) were incubated in hypoxic conditions
for 24 h and then resuspended in binding buffer. Subsequently,
cells were incubated with Annexin V for 10 min at 37°C in darkness
and then stained with PI for 5 min at 37°C in the dark. H9c2 cell
apoptosis was assessed using an ACEA NovoCyte flow cytometer
(2060R, ACEA; Agilent Technologies, Inc.) with ACEA NovoExpress
software version 1.2.1 (ACEA; Agilent Technologies, Inc.).
Lactate dehydrogenase (LDH)
determination assay
The LDH Activity Detection kit (cat. no. BC0685;
Beijing Solarbio Science & Technology Co., Ltd.) was used to
detect LDH release in H9c2 cells according to the manufacturer's
instructions.
Dual-Luciferase reporter assay
The interaction between miR-325-3p and MEG3 or TRPV4
was predicted using LncBase Predicted (version 2; carolina.imis.athena-innovation.gr/diana_tools/web/index.php?r=lncbasev2%2Findex-predicted)
or Targets can Human 7.1 (www.targetscan.org) online databases, respectively.
The wild-type (WT) and mutant (MUT) sequences of MEG3 or the
3′-untranslated regions (3′-UTRs) of TRPV4 were inserted into the
pmirGLO vector (Hunan YouBio Medical Device Co., Ltd.) to construct
WT-MEG3, MUT-MEG3, WT-TRPV4-3′UTR and MUT-TRPV4-3′UTR luciferase
reports, respectively. H9c2 cells (4×105 cells/well)
were co-transfected with WT-MEG3 (2 µg), MUT-MEG3 (2 µg),
WT-TRPV4-3′UTR (2 µg) or MUT-TRPV4-3′UTR (2 µg) and miR-325-3p (25
nM at final concentration) or miR-NC (25 nM at final concentration)
using Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.). Cells were replaced with fresh medium 6 h after
transfection before the hypoxia group received hypoxia treatment 24
h after transfection for subsequent experiments. Luciferase
activities were detected 48 h after transfection using the
Dual-Lucy Assay kit (Beijing Solarbio Science & Technology Co.,
Ltd.). Renilla luciferase activities were normalized as
control.
Statistical analysis
Quantitative data from three independent repeats are
presented as the mean ± SD. Comparisons between two groups or among
multiple groups were analyzed using the unpaired Student's t-test
or one-way ANOVA followed by Tukey's post hoc test, respectively.
Statistical analyses were performed using GraphPad Prism software
(version 7; GraphPad Software, Inc.). P<0.05 was considered to
indicate a statistically significant difference.
Results
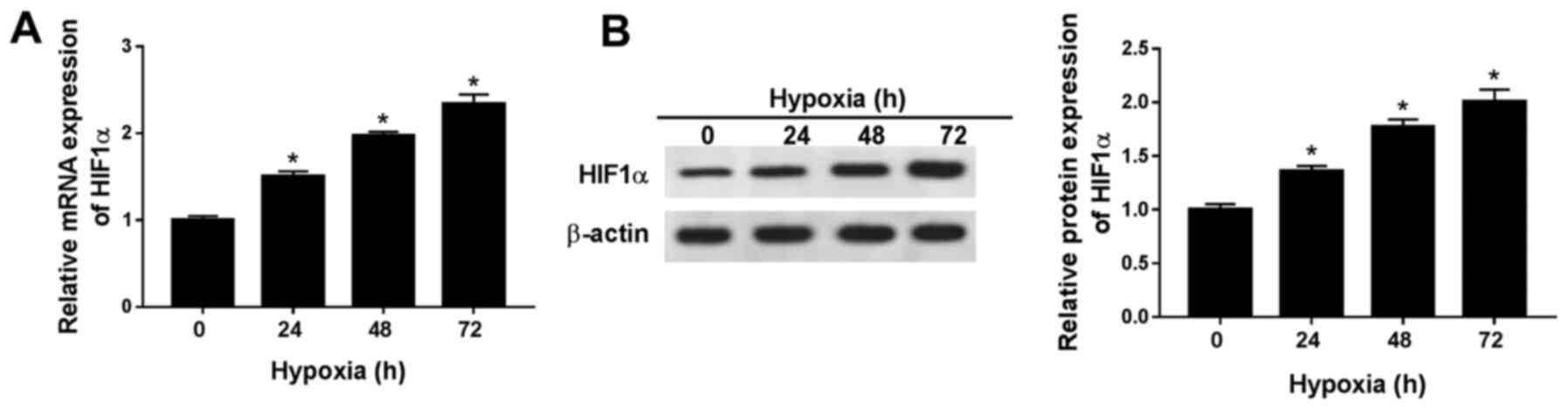
HIF1α expression levels are
significantly increased in hypoxic H9c2 cells
To establish a hypoxic rat cell model, H9c2 cells
were cultured in 0.5% O2 conditions. HIF1α is an
indicator of hypoxia (15), thus
the expression levels of HIF1α were detected in H9c2 cells after
hypoxia treatment for 0, 24, 48 or 72 h. The mRNA and protein
expression levels of HIF1α were significantly increased in H9c2
cells after hypoxia treatment for 24, 48 and 72 h compared with
normoxic H9c2 cells (hypoxia treatment for 0 h; Fig. 1A and B). The results indicated that
the hypoxia-induced cardiomyocyte model was successfully
established. As hypoxia treatment over a long duration may result
in irreversible damage to H9c2 cells, hypoxia treatment for 24 h
was selected for subsequent experiments.
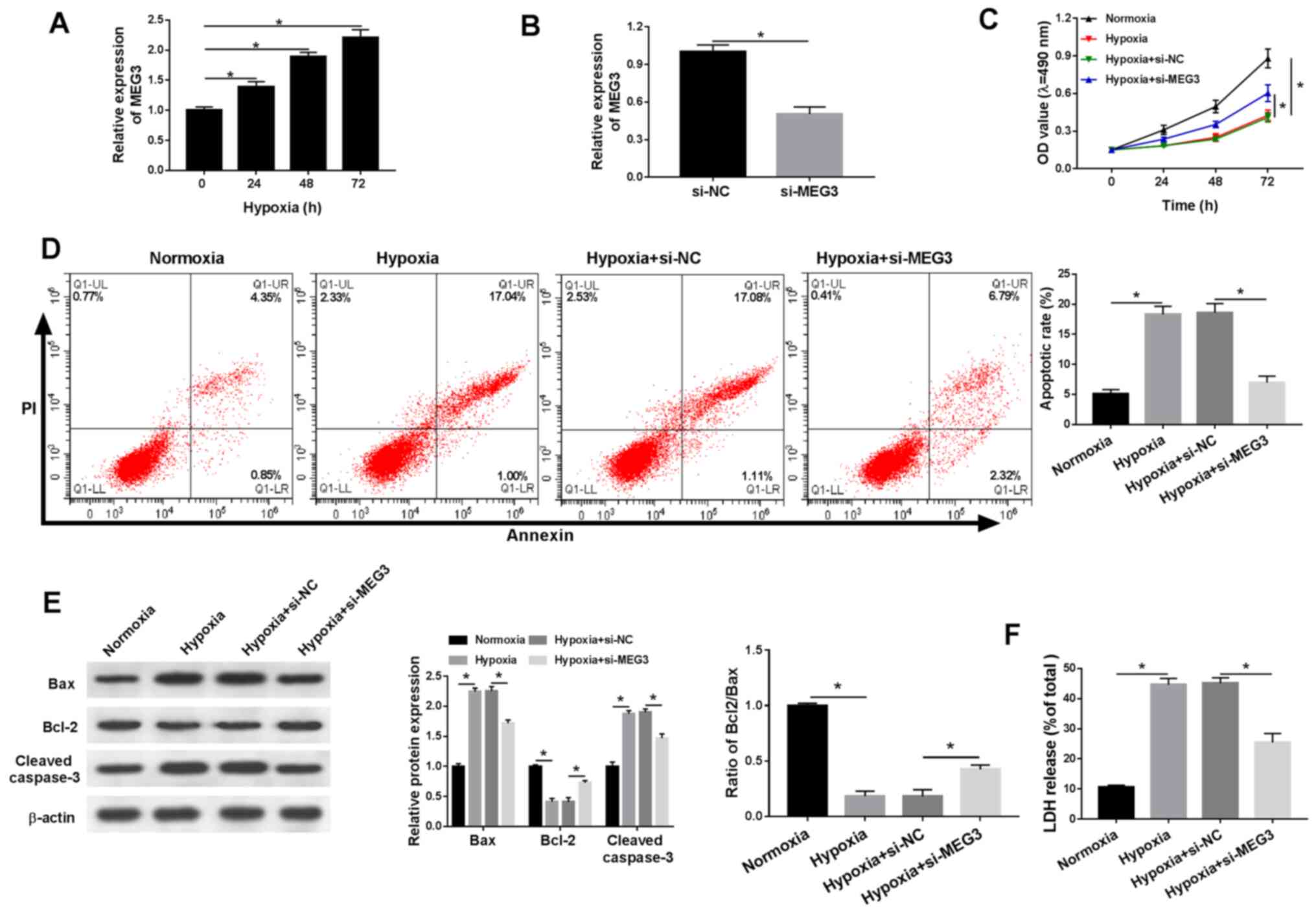
MEG3 knockdown mitigates
hypoxia-induced injury in H9c2 cells
The present study demonstrated that lncRNA MEG3
expression was significantly increased in a time-dependent manner
under hypoxic conditions compared with normoxic H9c2 cells
(Fig. 2A). In addition, the
expression levels of MEG3 were significantly lower in
hypoxia-treated H9c2 cells transfected with si-MEG3 compared with
the si-NC group, indicating the transfection efficiency of si-MEG3
(Fig. 2B). Additionally, compared
with normoxic H9c2 cells, cell viability was significantly reduced
under hypoxic conditions, which was inhibited by transfection with
si-MEG3 (Fig. 2C). Furthermore,
hypoxia treatment significantly induced H9c2 cell apoptosis
compared with normoxic H9c2 cells, and si-MEG3 attenuated
hypoxia-induced apoptosis compared with si-NC (Fig. 2D). Also, compared with normoxic H9c2
cells, the protein expression levels of proapoptotic factors (Bax
and cleaved casapase-3) were significantly increased under hypoxic
conditions compared with normoxic conditions, but partly reversed
by si-MEG compared with si-NC. By contrast, compared with normoxic
H9c2 cells, the expression levels of the antiapoptotic marker Bcl-2
were significantly decreased in hypoxic H9c2 cells, resulting in a
decreased Bcl2/Bax ratio and MEG3 knockdown partially reversed
hypoxia-induced alterations to the Bcl2/Bax ratio compared with
si-NC (Fig. 2E). LDH is related to
cardiomyocytes injury (16), thus
the release of LDH under hypoxic conditions was also monitored.
Compared with normoxic H9c2 cells, LDH release was significantly
increased in hypoxia-treated H9c2 cells, but significantly reduced
in hypoxia-treated H9c2 cells transfected with si-MEG3 compared
with H9c2 cells transfected with si-NC (Fig. 2F). The results suggested that MEG3
knockdown alleviated hypoxia-induced injury in H9c2 cells.
miR-325-3p negatively interacts with
MEG3 in hypoxic H9c2 cells
To elucidate the mechanism underlying MEG3 in
hypoxic cardiomyocytes, LncBase Predicted was used to identify the
putative target of MEG3. Based on the bioinformatics analysis,
miR-325-3p was identified as a potential target of lncRNA MEG3, and
the difference in expression of miR-325-3p was the most significant
among the candidate miRNAs in si-MEG treated cells (data not
shown). As shown in Fig. 3A,
miR-325-3p displayed complementary sequences with MEG3. The
Dual-Luciferase reporter assay results demonstrated that miR-325-3p
significantly decreased the luciferase activity of WT-MEG3 in
hypoxia-treated H9c2 cells compared with the miR-NC group, whereas
the luciferase activity of MUT-MEG3 was not significantly altered
by transfection with miR-325-3p compared with miR-NC (Fig. 3B). In addition, the expression
levels of miR-325-3p were significantly downregulated in
hypoxia-treated H9c2 cells compared with normoxic H9c2 cells
(Fig. 3C). Subsequently, MEG3 was
overexpressed in hypoxia-induced H9c2 cells by transfection with
MEG3. The RT-qPCR results confirmed the transfection efficiency of
MEG3 (Fig. 3D). In addition, the
expression levels of miR-325-3p were significantly increased in
hypoxia-treated H9c2 cells transfected with si-MEG3 compared with
hypoxia-treated H9c2 cells transfected with si-NC. By contrast,
miR-325-3p expression levels were significantly decreased in the
MEG3 group compared with the vector group in hypoxia-treated H9c2
cells (Fig. 3E). The results
suggested that MEG3 reduced the expression levels of miR-325-3p in
hypoxia-treated H9c2 cells.
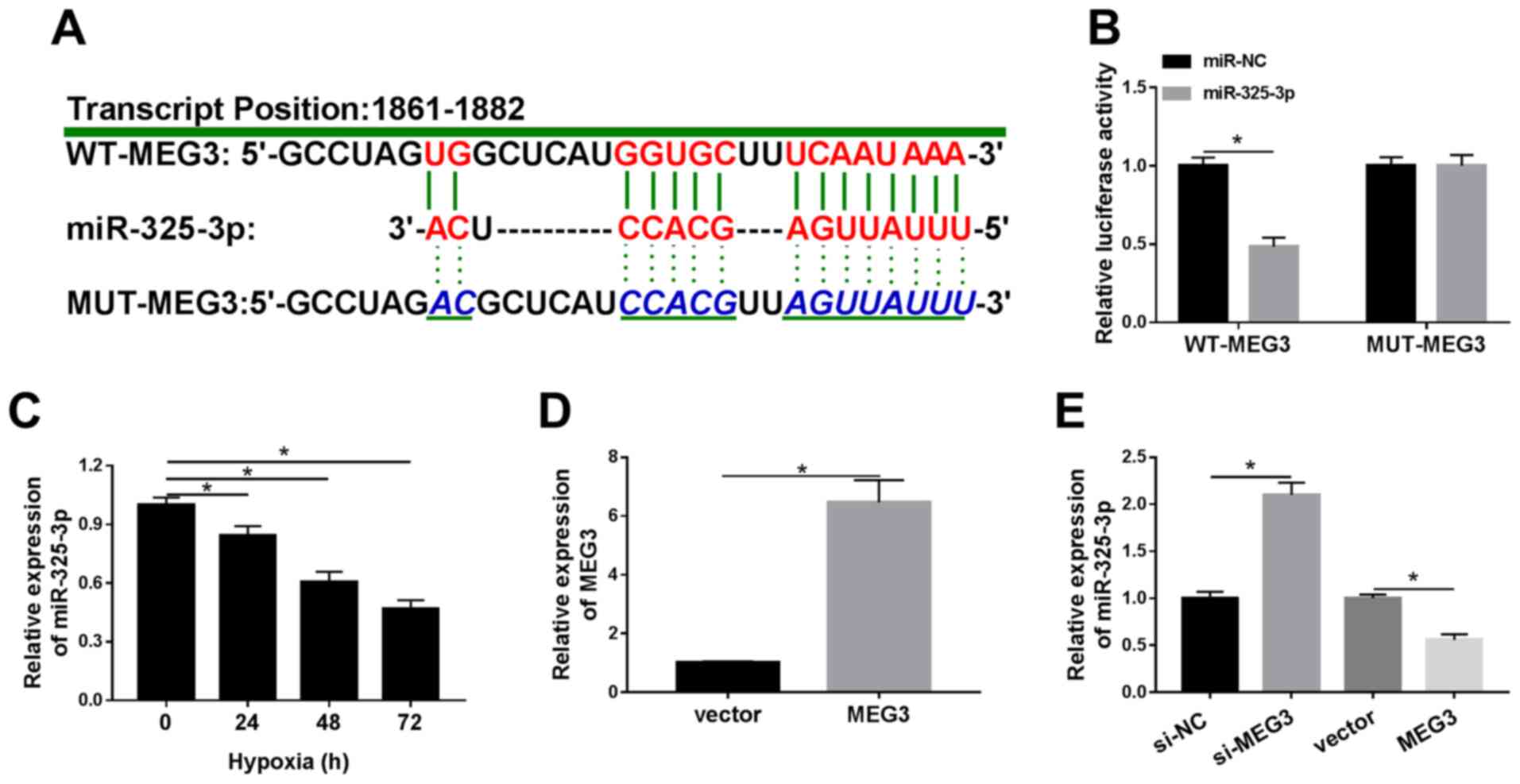 | Figure 3.miR-325-3p negatively interacts with
MEG3 in hypoxia-treated H9c2 cells. (A) The complementary binding
sites between MEG3 and miR-325-3p, as well as the mutant sequences
of MEG3. (B) The luciferase activities of WT-MEG3 and MUT-MEG3 in
hypoxia-treated H9c2 cells transfected with miR-325-3p or miR-NC.
(C) miR-325-3p expression levels in H9c2 cells after hypoxia
treatment for 24, 48 or 72 h. (D) MEG3 expression levels in
hypoxia-treated H9c2 cells transfected with vector or MEG3. (E)
miR-325-3p expression levels in hypoxia-treated H9c2 cells
transfected with si-NC, si-MEG3, vector or MEG3. n=3. *P<0.05.
miR, microRNA; MEG3, maternally expressed 3; WT, wild-type; MUT,
mutant; NC, negative control; si, small interfering RNA. |
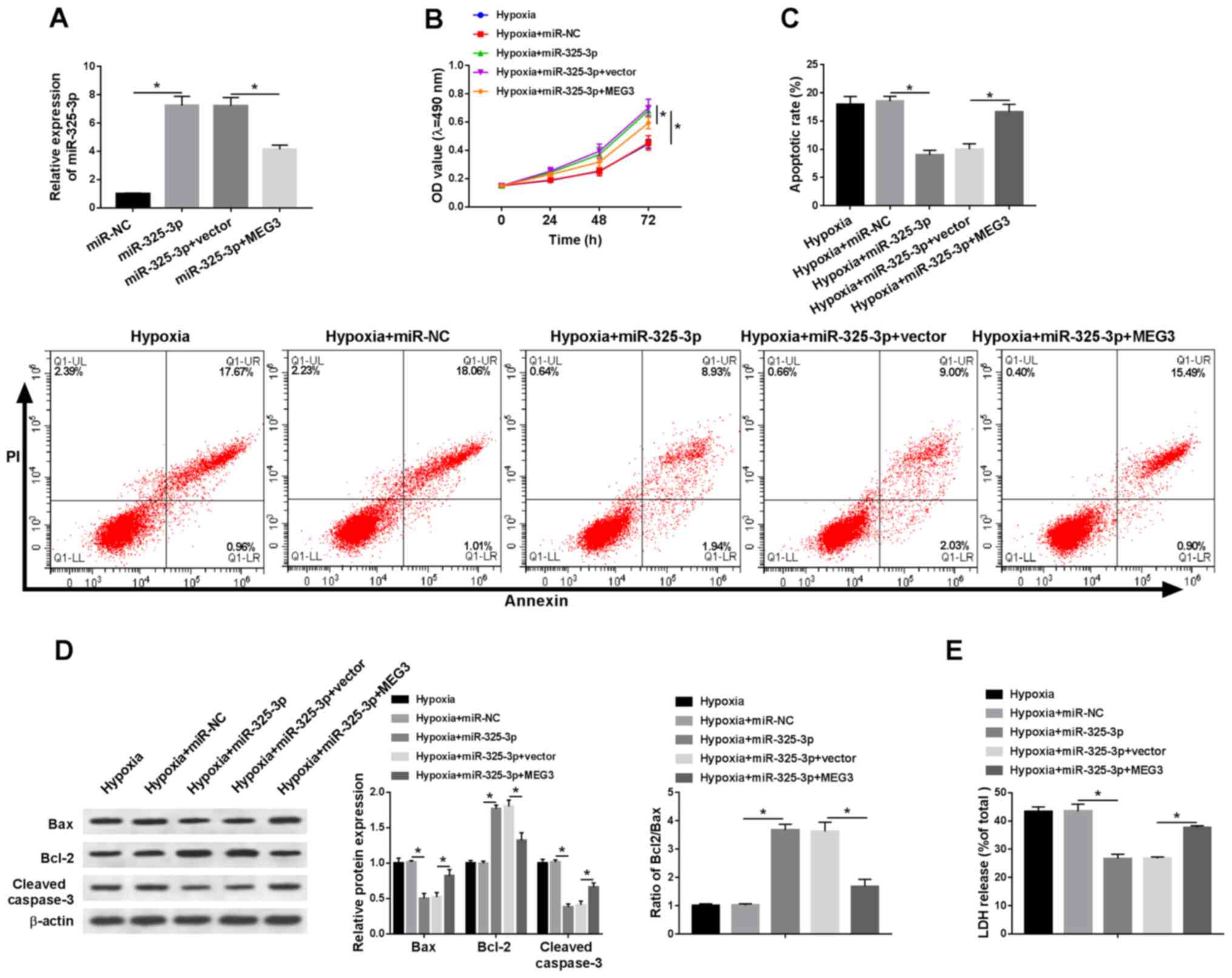
miR-325-3p overexpression improves
hypoxia-induced injury in H9c2 cells by downregulating MEG3
To explore whether MEG3-associated hypoxia injury
was mediated by miR-325-3p, rescue experiments were performed. The
expression levels of miR-352-3p were detected in hypoxia-treated
H9c2 cells. Compared with the miR-NC group, the expression levels
of miR-325-3p were significantly elevated in miR-325-3p-transfected
H9c2 cells under hypoxic conditions, which was partially inhibited
by MEG3 compared with the vector group (Fig. 4A). In addition, MEG3 reduced
miR-325-3p-mediated effects on cell viability in H9c2 cells under
hypoxic conditions (Fig. 4B).
Additionally, the rate of apoptosis and LDH release were
significantly decreased in hypoxia-treated H9c2 cells transfected
with miR-325-3p compared with the hypoxia + miR-NC group, which was
partially recovered in hypoxia-treated H9c2 cells co-transfected
with miR-325-3p and MEG3 compared with the hypoxia + miR-325-3p +
vector group (Fig. 4C and E). In
addition, MEG3 reversed miR-325-5p-mediated downregulation of Bax
and cleaved caspase-3 protein expression levels, and attenuated
miR-325-3p-mediated upregulation of Bcl-2 protein expression levels
in hypoxia-treated H9c2 cells (Fig.
4D). The results suggested that MEG3 aggravated hypoxia-induced
injury in H9c2 cells by regulating miR-325-3p.
TRPV4 is a candidate target of
miR-325-3p in hypoxia-treated H9c2 cells
To further investigate the mechanism underlying
miR-325-3p in hypoxia-treated cardiomyocytes, the potential
candidate was identified using TargetScan. As shown in Fig. 5A, the 3′UTR of TRPV4 displayed a
complementary base pairing with miR-325-3p. In addition, the
luciferase activity of the WT-TRPV4-3′UTR reporter was
significantly decreased in miR-325-3p-transfected H9c2 cells under
hypoxia conditions compared with the miR-NC group, whereas
miR-325-3p displayed no significant effect on the luciferase
activity of MUT-TRPV4-3′UTR compared with miR-NC (Fig. 5B). Additionally, the mRNA and
protein expression levels of TRPV4 were significantly increased by
hypoxia treatment in a time-dependent manner compared with normoxic
H9c2 cells (Fig. 5C and D). In
addition, the expression levels of miR-325-3p were significantly
reduced in H9c2 cells transfected with anti-miR-325-3p compared
with the anti-miR-NC group (Fig.
5E). Moreover, the mRNA and protein expression levels of TRPV4
were significantly reduced in the miR-325-3p group compared with
the miR-NC group, but significantly upregulated in the
anti-miR-325-3p group compared with the anti-miR-NC group (Fig. 5F and G). The results indicated that
TRPV4 was negatively regulated by miR-325-3p in hypoxia-treated
H9c2 cells.
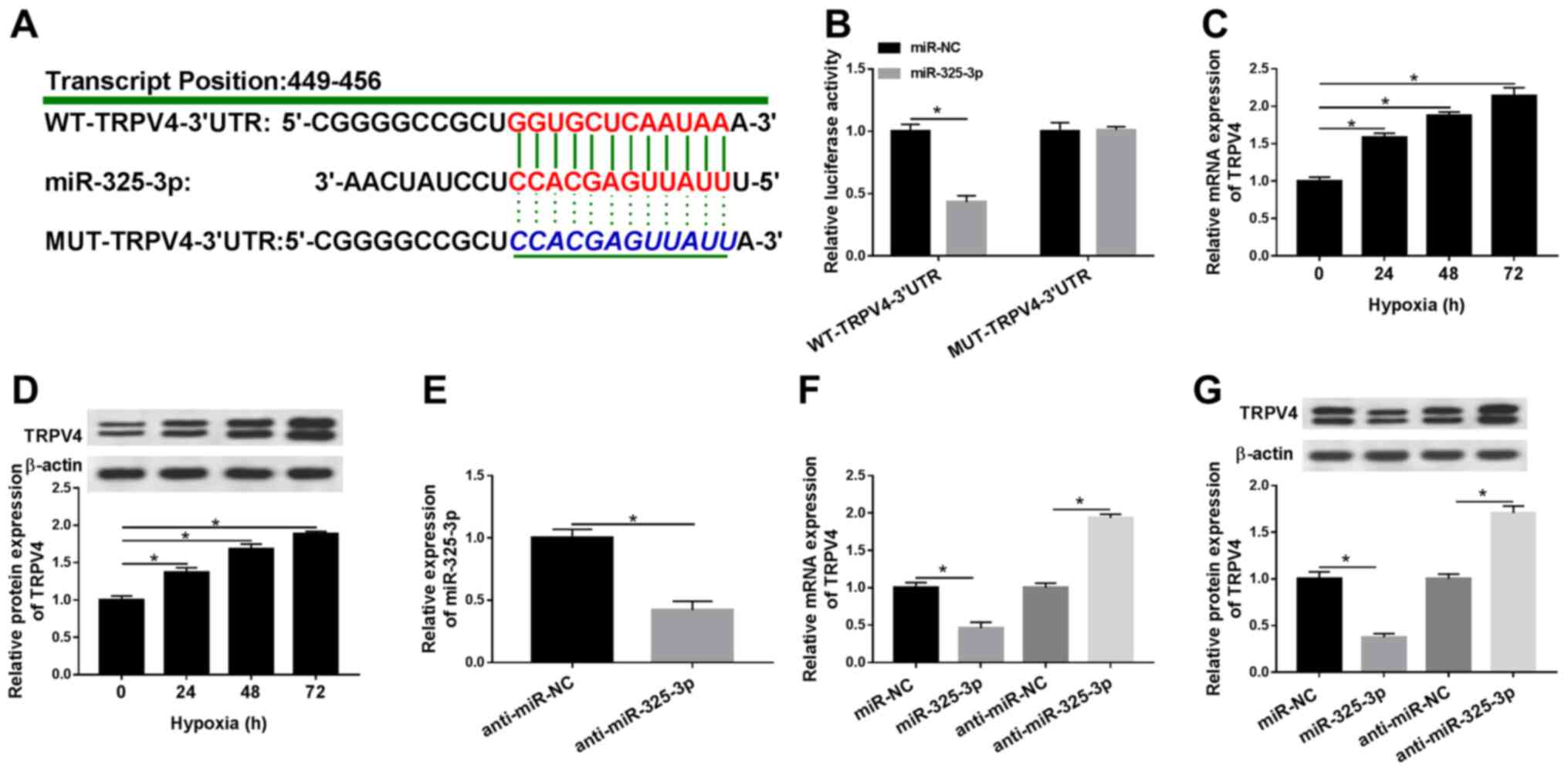 | Figure 5.TRPV4 is a candidate target for
miR-325-3p in hypoxia-treated H9c2 cells. (A) The complementary
binding sites between TRPV4 and miR-325-3p, as well as the mutant
sequences of TRPV4. (B) The luciferase activities of WT-TRPV4-3′UTR
and MUT-TRPV4-3′UTR in hypoxia-treated H9c2 cells transfected with
miR-325-3p or miR-NC. TRPV4 (C) mRNA and (D) protein expression
levels in H9c2 cells after hypoxia treatment for 24, 48 or 72 h.
(E) miR-325-3p expression levels in hypoxia-treated H9c2 cells
transfected with anti-miR-NC or anti-miR-325-3p. TRPV4 (F) mRNA and
(G) protein expression levels in hypoxia-treated H9c2 cells
transfected with miR-NC, miR-325-3p, anti-miR-NC or
anti-miR-325-3p. n=3. *P<0.05. TRPV4, transient receptor
potential cation channel subfamily V member 4; miR, microRNA; WT,
wild-type; UTR, untranslated region; MUT, mutant; NC, negative
control. |
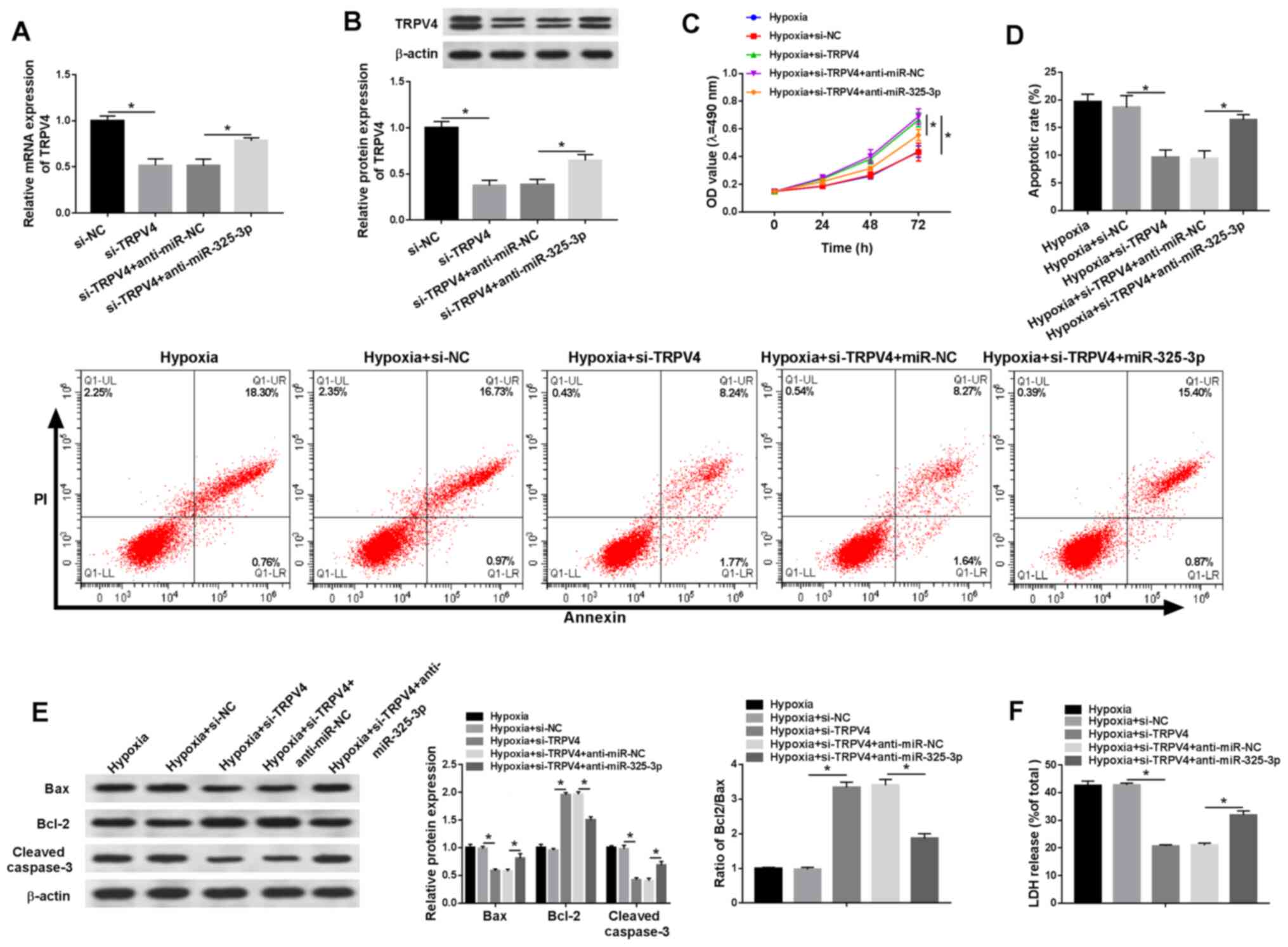
TRPV4 knockdown reduces
hypoxia-induced injury in H9c2 cells via miR-325-3p
To investigate whether the effects of miR-325-3p on
hypoxia-treated cardiomyocytes were mediated by TRPV4, si-TRPV4 and
anti-miR-325-3p were co-transfected into hypoxia-treated H9c2
cells. The mRNA and protein expression levels of TRPV4 were
significantly reduced in the si-TRPV4 group compared with the si-NC
group, which was partially reversed by anti-miR-325-3p compared
with anti-miR-NC (Fig. 6A and B).
In addition, co-transfection with anti-miR-325-3p reduced
si-TRPV4-mediated effects on cell viability in hypoxia-treated H9c2
cells (Fig. 6C). Additionally, the
rate of apoptosis and LDH release were significantly downregulated
in si-TRPV4-transfected H9c2 cells under hypoxic conditions
compared with the si-NC group, but partially reversed in
hypoxia-treated H9c2 cells co-transfected with si-TRPV4 and
anti-miR-325-3p compared with the hypoxia + si-TRPV4 + anti-miR-NC
group (Fig. 6D and F). Furthermore,
the western blotting results demonstrated that anti-miR-325-3p
reversed si-TRPV4-induced downregulation of Bax and cleaved
caspase-3 protein expression levels, and attenuated
si-TRPV4-mediated upregulation of Bcl-2 protein expression levels
in hypoxia-treated H9c2 cells (Fig.
6E). The results indicated that downregulation of TRPV4
mitigated hypoxia injury in H9c2 cells by regulating
miR-325-3p.
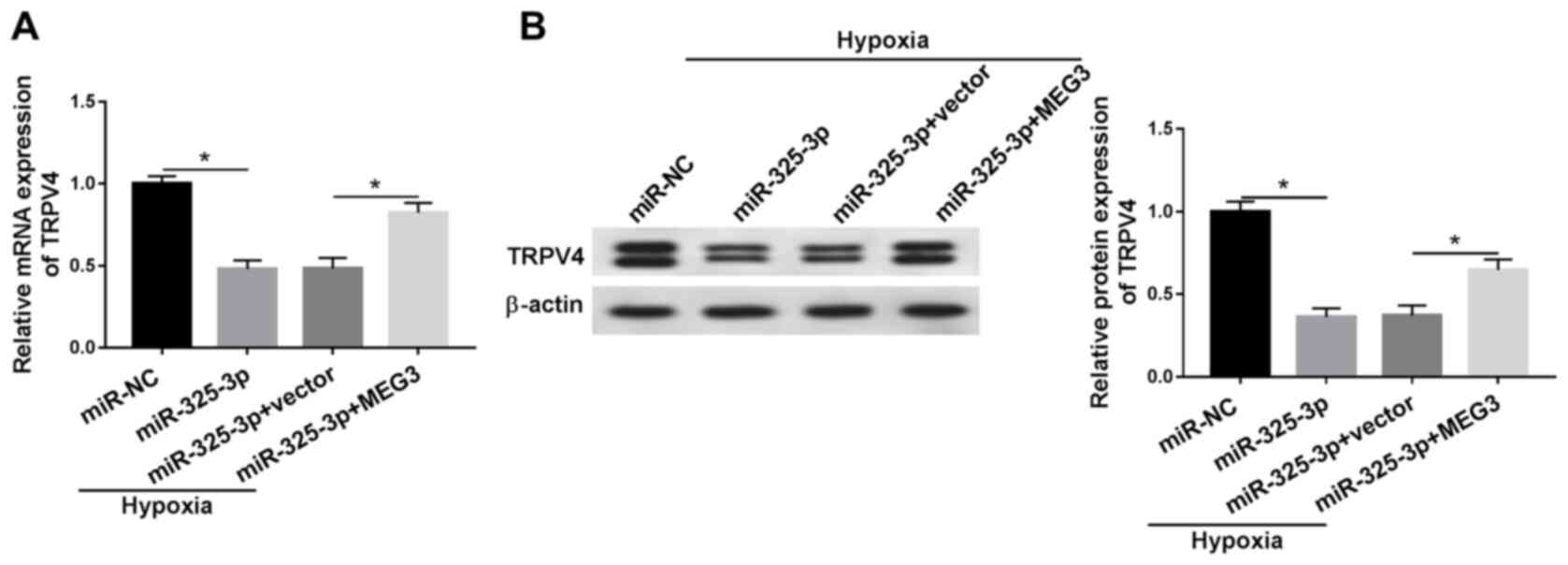
MEG3 upregulates TRPV4 expression in
hypoxia-treated H9c2 cells by downregulating miR-325-3p
To assess the relationship among MEG3, miR-325-3p
and TRPV4 in hypoxia-induced cardiomyocytes, MEG3 rescue
experiments were performed. Compared with the miR-NC group, the
mRNA and protein expression levels of TRPV4 were significantly
downregulated in miR-325-3p-transfected hypoxic H9c2 cells, but
partly rescued in hypoxic H9c2 cells co-transfected with miR-325-3p
and MEG3 compared with the hypoxia + miR-325-3p + vector group
(Fig. 7A and B). The results
demonstrated that MEG3 increased TRPV4 expression in
hypoxia-treated H9c2 cells via miR-325-3p. In addition, the changes
of MEG3, miR-325-3p and TRPV4 expression displayed no significant
effect on the expression levels of HIF1α in H9c2 cells (data not
shown).
Discussion
Acute myocardial infarction, a primary threat to
human health, causes cardiac cell death (17). A previous study revealed that
lncRNAs serve important roles, including regulating vessel growth
and function, controlling the contractile phenotype of smooth
muscle cells, hypertrophy, mitochondrial function and apoptosis in
cardiac diseases (18). The results
of the present study demonstrated that MEG3 regulated TRPV4
expression to reduce hypoxia-induced injury via miR-325-3p.
Accumulating evidence revealed that MEG3 is
dysregulated in numerous complex diseases. For instance, a report
on Alzheimer's disease demonstrated that MEG3 improved neuronal
damage via the PI3K/AKT signaling pathway (19). Another study on non-alcoholic fatty
liver disease reported that MEG3 was upregulated in high-content
hydrogen water (HHW)-treated mice in contrast to the mice treated
with deionized water, and MEG3 improved non-alcoholic fatty liver
disease under HHW conditions via the miR-136/nuclear factor,
erythroid 2 like 2 axis (20). The
aforementioned results indicated that MEG3 serves different roles
in different types of diseases. In the present study, MEG3
expression was significantly increased under hypoxic conditions
compared with normoxic conditions. Compared with the si-NC group,
MEG3 knockdown decreased the inhibitory effect of hypoxia on cell
viability, and partially reversed the promoting effects of hypoxia
on apoptosis and LDH release in H9c2 cells. The roles of MEG3
identified in the present study were consistent with previous
studies (6,7), which indicated that MEG3 aggravated
hypoxia-induced injury in AMI.
Previous studies demonstrated that miR-325-3p was
implicated in cellular processes in different types of diseases.
For example, Zhang et al (21) reported that miR-325-3p decreases
aquaporin 5 expression to confine hepatitis B virus replication in
hepatocellular carcinoma. Another study investigating
hypoxic-ischemic brain damage indicated that miR-325-3p aggravates
rat neonatal hypoxia-ischemia brain injury by modulating
aralkylamine N-acetyltransferase (22). The results of the present
demonstrated that miR-325-3p was sponged by MEG3 in hypoxia-treated
H9c2 cells. Moreover, miR-325-3p expression levels were
significantly decreased under hypoxic conditions compared with
normoxic conditions. MEG3 regulated hypoxia-mediated effects on
viability, apoptosis and LDH release in H9c2 cells via miR-325-3p.
Similar results were reported in a previous study that also
demonstrated that MEG3 facilitated hypoxia-stimulated injury by
sponging miR-325-3p in myocardial infarction mouse model (11).
Accumulating evidence has indicated that the
aberrant expression of TRPV4 was related to the development of
heart diseases. For example, Wu et al (23) demonstrated that TRPV4 was
significantly upregulated in the hypoxia/reoxygenation (H/R) model
compared with a normal groups cultured in normoxic conditions, and
accelerated cell injury in myocardial ischemia/reperfusion. Another
study revealed that TRPV4 decreased cardiomyocyte stability in H/R
conditions (24). In the present
study, TRPV4 was negatively regulated by miR-325-3p, and the level
of TRPV4 was significantly elevated in hypoxia-treated H9c2 cells
compared with normoxic H9c2 cells. In addition, TRPV4 knockdown
promoted cell viability, and reduced apoptosis and LDH release in
hypoxia-treated H9c2 cells compared with si-NC. The aforementioned
results of TRPV4 were consistent with a previous study that
dysregulation of TRPV4 occurs in AMI (13). In addition, MEG3 modulated TRPV4
expression under hypoxic conditions by sponging miR-325-3p. The
results suggested that MEG3 may regulate TRPV4 to aggravate
hypoxia-induced injury in AMI by sponging miR-325-3p. Although the
roles of MEG3, miR-325-3p and TRPV4 have been previously reported,
the present study suggested the regulatory network of the
MEG3/miR-325-3p/TRPV4 axis in hypoxia-induced injury, which may aid
with understanding the mechanisms underlying AMI development.
The present study had a number of limitations.
First, only a rat cardiomyocyte cell line H9c2 was used. Although
the H9c2 cell line is commonly used for the study of
hypoxia-induced injury, two or more cardiomyocytes cell lines
should be used to further verify the results of the present study.
Second, the present study did not perform in vivo
experiments. Therefore, future studies should employ other suitable
cell lines and an established AMI mouse model.
In conclusion, the present study demonstrated that
MEG3 and TRPV4 expression levels were significantly increased, and
miR-325-3p expression levels were significantly decreased in
hypoxic H9c2 cells compared with normoxic H9c2 cells. MEG3
knockdown relieved hypoxia-stimulated injury by downregulating
TRPV4 expression via miR-325-3p. The results of the present study
may aid with identifying the mechanism underlying AMI.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analysed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YZ designed the study. YZ, XGL, DZ, XYL and JD
performed the experiments. XGL and DZ analyzed the data. YZ and XGL
wrote the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare they have no competing
interests.
References
|
1
|
Anderson JL and Morrow DA: Acute
myocardial infarction. N Engl J Med. 376:2053–2064. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Li J, Li X, Wang Q, Hu S, Wang Y, Masoudi
FA, Spertus JA, Krumholz HM and Jiang L; China PEACE Collaborative
Group, : ST-segment elevation myocardial infarction in China from
2001 to 2011 (The China PEACE-retrospective acute myocardial
infarction study): A retrospective analysis of hospital data.
Lancet. 385:441–451. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Mathieu EL, Belhocine M, Dao L, Puthier D
and Spicuglia S: Functions of lncRNA in development and diseases.
Med Sci (Paris). 30:790–796. 2014.(In French). View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hu H, Wu J, Li D, Zhou J, Yu H and Ma L:
Knockdown of lncRNA MALAT1 attenuates acute myocardial infarction
through miR-320-Pten axis. Biomed Pharmacother. 106:738–746. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Huang S, Tao W, Guo Z, Cao J and Huang X:
Suppression of long noncoding RNA TTTY15 attenuates hypoxia-induced
cardiomyocytes injury by targeting miR-455-5p. Gene. 701:1–8. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wu H, Zhao ZA, Liu J, Hao K, Yu Y, Han X,
Li J, Wang Y, Lei W, Dong N, et al: Long noncoding RNA Meg3
regulates cardiomyocyte apoptosis in myocardial infarction. Gene
Ther. 25:511–523. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gong L, Xu H, Chang H, Tong Y, Zhang T and
Guo G: Knockdown of long non-coding RNA MEG3 protects H9c2 cells
from hypoxia-induced injury by targeting microRNA-183. J Cell
Biochem. 119:1429–1440. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Huang Y, Shen XJ, Zou Q, Wang SP, Tang SM
and Zhang GZ: Biological functions of microRNAs: A review. J
Physiol Biochem. 67:129–139. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chen Y, Zhao Y, Chen W, Xie L, Zhao ZA,
Yang J, Chen Y, Lei W and Shen Z: MicroRNA-133 overexpression
promotes the therapeutic efficacy of mesenchymal stem cells on
acute myocardial infarction. Stem Cell Res Ther. 8:2682017.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhang M, Cheng YJ, Sara JD, Liu LJ, Liu
LP, Zhao X and Gao H: Circulating microRNA-145 is associated with
acute myocardial infarction and heart failure. Chin Med J (Engl).
130:51–56. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang DY, Wang BJ, Ma M, Yu K, Zhang Q and
Zhang XW: MicroRNA-325-3p protects the heart after myocardial
infarction by inhibiting RIPK3 and programmed necrosis in mice. BMC
Mol Biol. 20:172019. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Grace MS, Bonvini SJ, Belvisi MG and
McIntyre P: Modulation of the TRPV4 ion channel as a therapeutic
target for disease. Pharmacol Ther. 177:9–22. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Rath G, Saliez J, Behets G, Romero-Perez
M, Leon-Gomez E, Bouzin C, Vriens J, Nilius B, Feron O and Dessy C:
Vascular hypoxic preconditioning relies on TRPV4-dependent calcium
influx and proper intercellular gap junctions communication.
Arterioscler Thromb Vasc Biol. 32:2241–2249. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kietzmann T, Samoylenko A, Roth U and
Jungermann K: Hypoxia-inducible factor-1 and hypoxia response
elements mediate the induction of plasminogen activator inhibitor-1
gene expression by insulin in primary rat hepatocytes. Blood.
101:907–914. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yu B and Wang W: Cardioprotective effects
of morroniside in rats following acute myocardial infarction.
Inflammation. 41:432–436. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Qiu H, Liu JY, Wei D, Li N, Yamoah EN,
Hammock BD and Chiamvimonvat N: Cardiac-generated prostanoids
mediate cardiac myocyte apoptosis after myocardial ischaemia.
Cardiovasc Res. 95:336–345. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Uchida S and Dimmeler S: Long noncoding
RNAs in cardiovascular diseases. Circ Res. 116:737–750. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yi J, Chen B, Yao X, Lei Y, Ou F and Huang
F: Upregulation of the lncRNA MEG3 improves cognitive impairment,
alleviates neuronal damage, and inhibits activation of astrocytes
in hippocampus tissues in Alzheimer's disease through inactivating
the PI3K/Akt signaling pathway. J Cell Biochem. 120:18053–18065.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang X and Wang J: High-content hydrogen
water-induced downregulation of miR-136 alleviates non-alcoholic
fatty liver disease by regulating Nrf2 via targeting MEG3. Biol
Chem. 399:397–406. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhang Z, Han Y, Sun G, Liu X, Jia X and Yu
X: MicroRNA-325-3p inhibits cell proliferation and induces
apoptosis in hepatitis B virus-related hepatocellular carcinoma by
down-regulation of aquaporin 5. Cell Mol Biol Lett. 24:132019.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yang Y, Sun B, Huang J, Xu L, Pan J, Fang
C, Li M, Li G, Tao Y, Yang X, et al: Up-regulation of miR-325-3p
suppresses pineal aralkylamine N-acetyltransferase (Aanat) after
neonatal hypoxia-ischemia brain injury in rats. Brain Res.
1668:28–35. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wu QF, Qian C, Zhao N, Dong Q, Li J, Wang
BB, Chen L, Yu L, Han B, Du YM and Liao YH: Activation of transient
receptor potential vanilloid 4 involves in hypoxia/reoxygenation
injury in cardiomyocytes. Cell Death Dis. 8:e28282017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Gorbunov AS, Maslov LN, Jaggi AS, Singh N,
De Petrocellis L, Boshchenko AA, Roohbakhsh A, Bezuglov VV and
Oeltgen PR: Physiological and pathological role of TRPV1, TRPV2 and
TRPV4 channels in heart. Curr Cardiol Rev. 15:244–251. 2019.
View Article : Google Scholar : PubMed/NCBI
|