Introduction
Exposure to radiation causes damage to healthy
organs. The side effects of radiotherapy, including recurrence,
secondary cancer and normal tissue injury, are among the clinical
problems faced by patients with cancer. Notably, partial or total
body exposure to 5–10 Gy irradiation in a single fraction is not
lethal but can lead to nephropathy (1). Accidental radiation exposure has been
suggested to induce chronic kidney disease (CKD) as a potential
late-onset effect; however, the underlying factors have yet to be
investigated in detail. Radiation-induced nephropathy is often
observed in patients with gastrointestinal or gynecological cancer,
lymphoma and sarcoma of the upper abdomen (2). A previous study on renal
failure-associated mortality in atomic bomb survivors suggested
that radiation-induced renal dysfunction was a contributory factor
to increased circulatory disease (3). Assessment of renal function should be
included in the long-term follow-up plan for patients with
substantial clinical or accidental radiation exposure to the kidney
region, since CKD has a significant role in human morbidity and
mortality. The precise mechanisms underlying pathogenesis and/or
mediators involved in radiation-induced nephropathy are currently
under investigation.
Generally, exposure to ionizing radiation (IR)
induces direct injury to normal tissues and promotes generation of
reactive oxygen species (ROS) that cause damage to macromolecules
(4). Formation of excessive ROS has
been implicated in the aging process and pathogenesis of various
human diseases (5). Cellular
senescence is a physiological process through which cells lose the
ability to divide and proliferate, contributing to various
physiological and pathological processes of biological aging.
Typically, cellular senescence encompasses irreversible growth
arrest, normal tissue repair and tumor suppression, and is induced
in response to IR (6,7). Senescent cells secrete proinflammatory
factors, a condition known as senescence-associated secretory
phenotype (SASP) (5). Several
molecules of SASP are associated with radiation-induced normal
tissue injury, including TNF-α, IL-1, IL-6, TGF-β, epidermal growth
factor and vascular endothelial growth factor (8). In particular, the inflammatory
cytokine TNF-α has been shown to be rapidly and consistently
expressed in irradiated and adjacent tissue, and to be involved in
the acute phase reaction (8).
Deficiency of TNF-α in a lung injury model has been reported to
effectively prevent symptoms of radiation pneumonitis (9). While TNF-α is implicated in radiation
mucositis, enteritis and dermatitis, its specific involvement in
radiation-induced kidney injury has not been fully elucidated
(8).
Klotho, an aging suppressor gene (10), is expressed in several tissues and
is inhibited by TNF-α (11). The
highest levels of Klotho are detected in the kidney and brain. The
protein is additionally expressed in parathyroid glands and the
heart, but with lower abundance (12). Emerging evidence has indicated that
deficiency of Klotho is an early biomarker for CKD and acute kidney
injury (10). Two existing forms of
Klotho with distinct functions have been identified to date;
specifically, a transmembrane and a soluble secreted form. The
extracellular domain of Klotho is cleaved by ADAM metallopeptidase
domain (ADAM)9/10/17 to generate a soluble secreted form (10). A previous study has revealed that
deficiency in transmembrane and soluble forms of Klotho in patients
with CKD contributed to the pathogenesis of secondary
hyperparathyroidism, vascular calcification, left ventricular
hypertrophy and worsening of kidney injury (13). Klotho expression and cleavage
processes by ADAM9/10/17 have been widely examined in a range of
pathophysiological conditions; however, the precise roles of these
molecules in CKD remain to be established (14,15).
The present study hypothesized that TNF-α may exert
an inhibitory effect on the expression of Klotho, which in turn may
promote radiation-induced aging of renal epithelial cells. The aim
of the present study was to demonstrate that IR induces senescence
in mouse internal medulla collection duct-3 (mIMCD-3) cells.
Therefore, the mRNA and protein expression levels of TNF-α and
Klotho were detected following exposure to radiation. In addition,
changes to the expression of the ectodomain shedding enzyme of
Klotho, ADAM9/10/17, were investigated. These data intended to
comprehensively suggest that radiation-induced renal dysfunction
may be associated with inhibition of Klotho activity and promotion
of cellular senescence. Finally, the present study aimed to provide
insight into the pathogenic mechanisms responsible for
radiation-induced kidney injury.
Materials and methods
Cell culture
The mIMCD-3 (CRL-2123™) cell line was obtained from
the American Type Culture Collection and cultured in Dulbecco's
modified Eagle's medium:F-12 medium (Gibco; Thermo Fisher
Scientific, Inc.) supplemented with 10% FBS (GenDEPOT), 100 U/ml
penicillin and 100 µg/ml streptomycin. Cells were grown in a
humidified atmosphere containing 95% air and 5% CO2 at
37°C.
In vitro and in vivo irradiation
For γ-irradiation, cells were exposed to a total of
6 Gy for 2 min (dose rate, 3 Gy/min) with a 137Cs γ-ray
source (Gamma-Service Medical GmbH). BALB/c and C57BL/6 mice were
purchased from Koatech Co., Ltd. Mice were housed at 22±2°C and
50±10% humidity under a 12-h light/dark cycle. Five animals were
housed per cage with free access to drinking water and standard
mouse chow. For irradiation, 8-week-old (male; weight, 20–25 g)
mice were placed in well-ventilated custom jigs, allowing for
precise delivery of radiation using the X-Rad 320 system (Precision
Inc.; 260 kV, 10 mA) to a total of 6 Gy for 3 min (dose rate, 2
Gy/min). Sham-irradiated mice were similarly placed in custom jigs
and positioned in the X-Rad 320 system without X-ray exposure. A
UNIDOSE® universal dosimeter (PTW-Freiburg) was used to
measure the dose rate (16). The
control and IR-treated mouse groups (n=3) were euthanized 1 and 3
days after irradiation by exposure to CO2 (30% flow rate
of gradual-fill CO2). Death was confirmed by monitoring
cardiac and respiratory arrest. Subsequently, kidneys were removed,
frozen in liquid nitrogen and stored at −70°C. For in vivo
studies, all animal experiments were conducted in accordance with
the Guidelines for the Use and Care of Laboratory Animals (16) and the study was approved by the
Institutional Animal Care and Use Committee (IACUC) of the Korea
Institute of Radiological and Medical Sciences (KIRAMS; IACUC
approval no. 2016-0065; Seoul, South Korea).
Cell counting and proliferation
assays
Cells were seeded onto 6-well plates
(2×104/well), incubated for 24 h, and subjected to 6 Gy
radiation. Cell numbers were counted 24, 48 and 72 h after
radiation and cells were suspended in fresh cell culture medium.
Viable cells were visualized using trypan blue exclusion (17) and counted with an Automated Cell
Counter (Bio-Rad Laboratories, Inc.). Cell proliferation was
assessed using the WST assay (Cyto X cell viability assay kit; LPS
Solution) according to the manufacturer's instructions.
Fluorescence-activated cell sorting
analysis
Cell cycle analysis was determined using a BD
Accuri™ C6 Plus flow cytometer (BD Biosciences). Briefly, cells
were seeded onto 6-well plates (2×104/well), incubated
for 24 h, and subjected to 6 Gy irradiation. After 72 h, cells were
harvested by centrifugation at 200 × g for 5 min at room
temperature, washed with PBS, fixed overnight at 4°C with ice-cold
75% ethanol, and incubated with RNase A (10 µg/ml) and propidium
iodide (PI; 50 µg/ml in 0.1% sodium citrate with 0.1% NP-40) for 15
min at room temperature in the dark. Cells in the G1, S and G2/M
phases were identified based on fluorescence intensity and cell
cycle distribution was analyzed using BD Accuri™ C6 Plus software
version 1.0.23.1 (BD Biosciences).
Western blotting and ELISA
Total cell lysates were prepared in RIPA lysis
buffer (GenDEPOT) containing a freshly added protease inhibitor
cocktail (GenDEPOT). The protein concentration was then measured
using the Bradford assay (Bio-Rad Laboratories, Inc.) Protein
extracts (15 µg) were then mixed with SDS sample buffer, boiled for
5 min and separated by SDS-PAGE on 12% (w/v) gels. Subsequently,
proteins were transferred to nitrocellulose membranes (EMD
Millipore). Blots were blocked in 5% BSA solution (BSA in 1X
TBS-0.1% Tween-20; GenDEPOT) at room temperature for 1 h. Blots
were incubated overnight at 4°C with primary antibodies (1:1,000
dilution) against heterochromatin protein 1γ (HP1γ; cat. no.
ab56978), ADAM9 (cat. no. ab186833), ADAM10 (cat. no. ab1997)
ADAM17 (cat. no. ab2051) (all Abcam), sirtuin 1 (SIRT1; cat. no.
sc-15404), β-actin (cat. no. sc-47778) (both Santa Cruz
Biotechnology, Inc.) and V5 (cat. no. R-960-25; Invitrogen; Thermo
Fisher Scientific, Inc.) followed by incubation with HRP-conjugated
goat anti-rabbit (cat. no. SA002-500) and goat anti-mouse (cat. no.
SA001-500) secondary antibodies (1:5,000 dilution; both GenDEPOT)
for 30 min at room temperature. Protein bands were detected with
Western Lightning Plus ECL (PerkinElmer) and semi-quantification of
protein expression was visualized using an Amersham Imager 600 (GE
Healthcare).
Klotho and TNF-α levels were measured in cell
lysates using the Mouse TNF-α ELISA Ready-Set-Go (cat. no. 88-7324;
Invitrogen; Thermo Fisher Scientific, Inc.) and Mouse Klotho ELISA
(cat. no. LS-F6578; LSBio) kits. Experiments were performed
according to the manufacturer's instructions without further
optimization and absorbance was read at 450 nm using the Multiskan
FC microplate photometer (Thermo Fisher Scientific, Inc.).
Immunocytochemistry
mIMCD-3 cells were seeded on 8-well cell culture
slides (4×103/well) (SPL Life Sciences), incubated for
24 h, and subjected to 6 Gy irradiation. After 72 h, slides were
rinsed with PBS and fixed in 4% paraformaldehyde overnight at 4°C,
followed by permeabilization in 0.1% Triton X-100 for 10 min at
room temperature and rinsing with PBS. Cells were blocked in 5% BSA
solution (BSA in 1X TBS-0.1% Tween-20) (GenDEPOT) at room
temperature for 1 h. Subsequently, cells were incubated overnight
at 4°C with primary antibodies (1:100 dilution) against HP1γ and
SIRT1, followed by incubation with the following secondary
antibodies (1:200 dilution): Goat anti-Rabbit IgG (H+L)
Cross-Adsorbed Secondary Antibody, Alexa Fluor 488 (cat. no.
A11008) and Goat anti-Mouse IgG (H+L) Cross-Adsorbed Secondary
Antibody, Alexa Fluor 488 (cat. no. A11001) (Invitrogen; Thermo
Fisher Scientific, Inc.) for 2 h at room temperature in the dark.
Specimens were then treated with 2 µg/ml PI (Sigma-Aldrich; Merck
KGaA) in PBS for 5 min at room temperature, mounted on glass
slides, and observed under a Zeiss LSM 710 confocal microscope
(Carl Zeiss AG).
Reverse transcription-PCR (RT-PCR) and
RT-quantitative (q)PCR
Total RNA was isolated from cells and tissues using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.) and quantified via formaldehyde-agarose gel electrophoresis.
Single-stranded cDNA was synthesized from RNA (4 µg) using 0.27 µg
oligo dT and amfiRivert reverse transcriptase (cat. no. R5600-200;
GenDEPOT) according to the manufacturer's instructions. The desired
cDNA fragments were amplified via RT-qPCR using the following
primers: Klotho, forward 5′-TGTGAATGAGGCTCTGAAAGC-3′, reverse
5′-GAGCGATCACTAAGTGAATACG-3′; TNF-α, forward
5′-CCACCACGCTCTTCTGTCTAC-3′, reverse 5′-AGGGTCTGGGCCATAGAACT-3′;
ADAM9, forward 5′-GGATATGGAGGAAGCGTGGA-3′, reverse
5′-GCAACAAGGGGGACGATTAG-3′; ADAM10, forward
5′-AGCAACATCTGGGGACAAAC-3′, reverse 5′-TGGCCAGATTCAACAAAACA-3′;
ADAM17, forward 5′-GTACGTCGATGCAGAGCAAA-3′, reverse
5′-GAAATCCCAAAATCGCTCAA-3′; and GAPDH, forward
5′-AAGGGCTCATGACCACAGTC-3′ and reverse 5′-TTCAGCTCTGGGATGACCTT-3′.
To determine mRNA expression, reactions were conducted in a total
volume of 20 µl containing 1 µl cDNA, 5X Hot FIREPol®
EvaGreen® qPCR Supermix (Solis BioDyne) and the relevant
primers. The Mic qPCR cycler for real-time PCR (Bio Molecular
Systems) was used to assess mRNA expression. Expression data were
normalized to the geometric mean of the housekeeping gene, GAPDH,
to control the variability in expression levels and analyzed using
the 2−ΔΔCq method (18).
Transfection
A total of 1 day prior to transfection, cells were
seeded onto 60-mm dishes (4×105/well). For
pcDNA3.1/V5-His-TOPO vector plasmid (cat. no. K480001, Invitrogen;
Thermo Fisher Scientific, Inc) and Klotho expression plasmid
(V5-klotho; cat. no. 17712; Addgene, Inc.) DNA transfection, 8 µg
DNA was added to 0.5 ml Opti-MEM (Gibco; Thermo Fisher Scientific,
Inc.), followed by the addition of 0.5 ml Opti-MEM containing 20 µl
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.). The mixture was incubated at room temperature
for 5 min and was then added to cells. After incubation for 6 h at
37°C, the medium was changed, followed by incubation of cells for
24 h and exposure to 6 Gy radiation. The empty pcDNA3.1/V5-His-TOPO
vector was used as the negative control.
Statistical analysis
Data are presented as the mean ± SD of at least
three experiments. Statistical comparisons among the groups were
analyzed with unpaired Student's t-test, or one-way ANOVA and
Tukey's post hoc test using SPSS software version 23 (IBM Corp.).
P<0.05 was considered to indicate a statistically significant
difference.
Results
Radiation induces senescence of kidney
cells
To determine whether IR induces cellular senescence
in mIMCD-3 cells, cell viability and senescence-associated markers,
HP1γ and SIRT1 (19), were
initially measured. Cell viability and the proportion of cells in
G1 phase of the cell cycle were significantly decreased in mIMCD-3
cells in response to irradiation (Fig.
1A-C). In addition, the expression levels of HP1γ and SIRT1
levels were significantly decreased in irradiated mIMCD-3 cells
(Fig. 1D-F), as determined via
western blotting. Immunofluorescence findings confirmed the
IR-induced decrease in the expression levels of HP1γ and SIRT1
(Fig. 1G and H). These results
indicated that irradiation may suppress viability and promote
senescence of kidney cells.
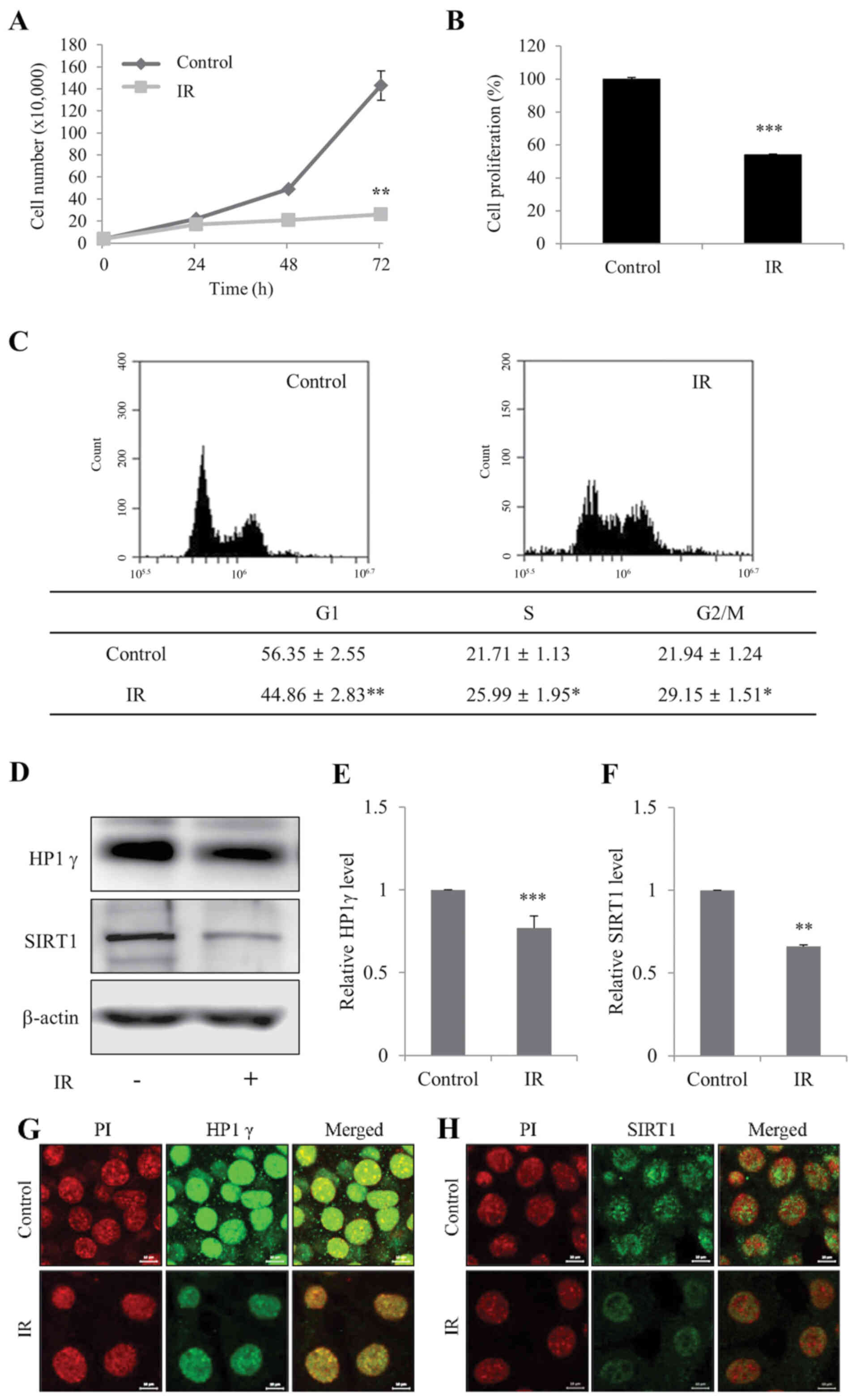 | Figure 1.Radiation induces senescence of
mIMCD-3 cells. (A) Control and IR-treated viable mIMCD-3 cells were
visualized using Trypan blue exclusion and counted. (B) Control and
IR-treated cell proliferation wase determined using the WST assay.
Data are presented as a percentage of control cell growth. (C)
Control and IR-treated mIMCD-3 cells were stained with PI and
subjected to flow cytometry. (D) Protein expression levels of HP1γ
and SIRT1 were determined via western blotting, with β-actin as the
loading control. Semi-quantification of (E) HP1γ and (F) SIRT1
protein expression levels in cell lysates relative to the control
group. Immunofluorescence analysis of (G) HP1γ and (H) SIRT1 using
a fluorescein isothiocyanate-conjugated secondary antibody and PI
nuclear staining followed by confocal microscopy (scale bar, 10
µm). mIMCD-3 cells were treated with or without 6 Gy radiation and
cultured for 72 h. Data are expressed as the mean ± SD (n=4),
*P<0.05, **P<0.01, ***P<0.001 vs. Control. mIMCD-3, mouse
inner medullary collecting duct-3; IR, ionizing radiation; HP1γ,
heterochromatin protein 1γ; SIRT1, sirtuin 1; PI, propidium
iodide. |
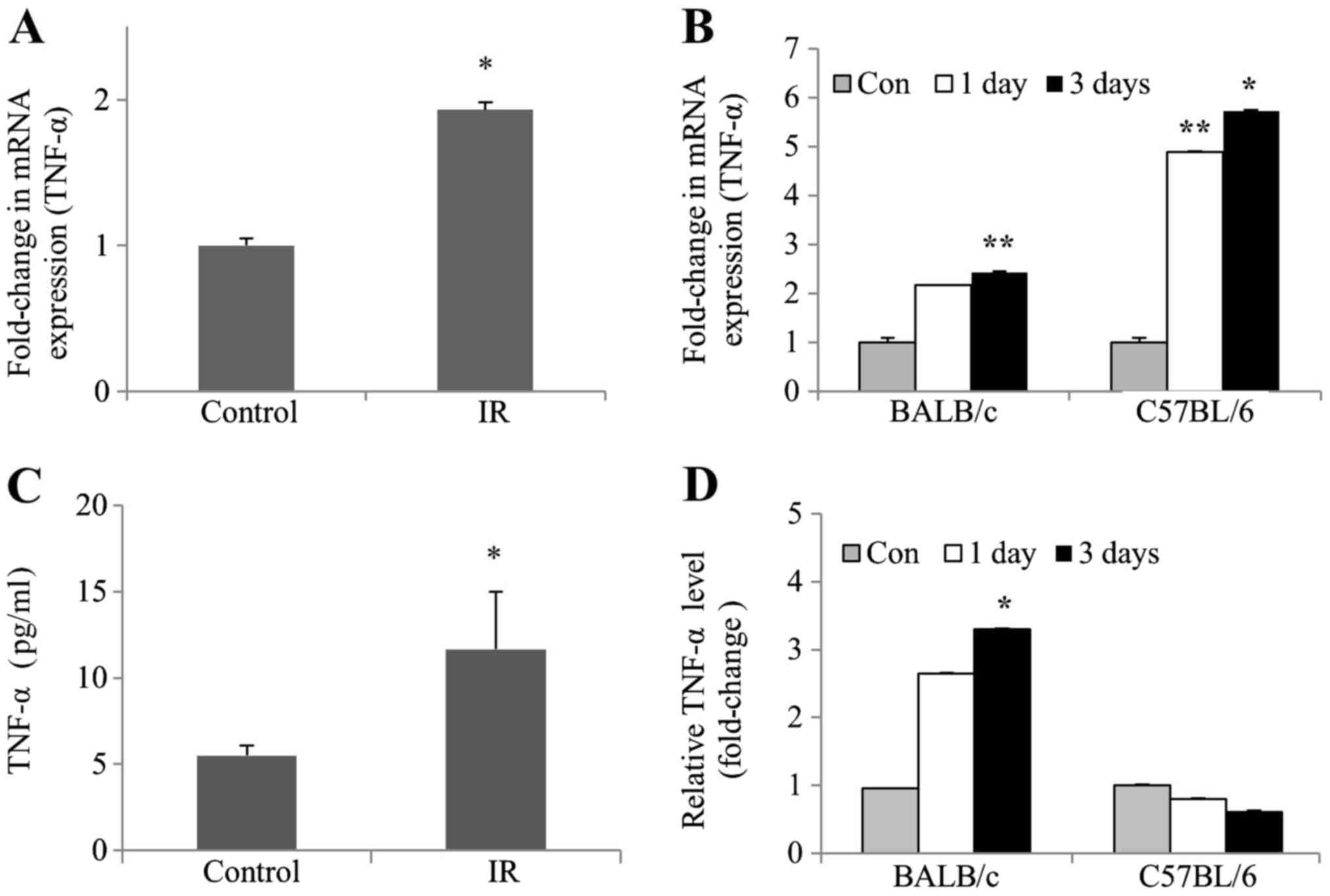
Radiation enhances TNF-α expression in
kidney cells and tissues of mice
To determine whether TNF-α levels were affected by
IR in vivo and in vitro, TNF-α mRNA expression and
protein levels were examined in irradiated mIMCD-3 cells, as well
as in the kidney tissues of BALB/c and C57BL/6 mice. RT-qPCR
experiments revealed a significant increase in the mRNA expression
levels of TNF-α, both in irradiated mIMCD-3 and C57BL/6 mice.
However, the mRNA expression levels of TNF-α in BALB/c mice were
only slightly increased (Fig. 2A and
B). Consistent with this finding, ELISA experiments revealed a
subsequent increase in TNF-α protein levels. Furthermore, TNF-α
protein levels were significantly increased in irradiated BALB/c
mice but only slightly decreased in irradiated C57BL mice (Fig. 2C and D). These collective data
clearly demonstrated that the protein and mRNA expression levels of
TNF-α were increased in irradiated kidney cells and BALB/c mice
tissues.
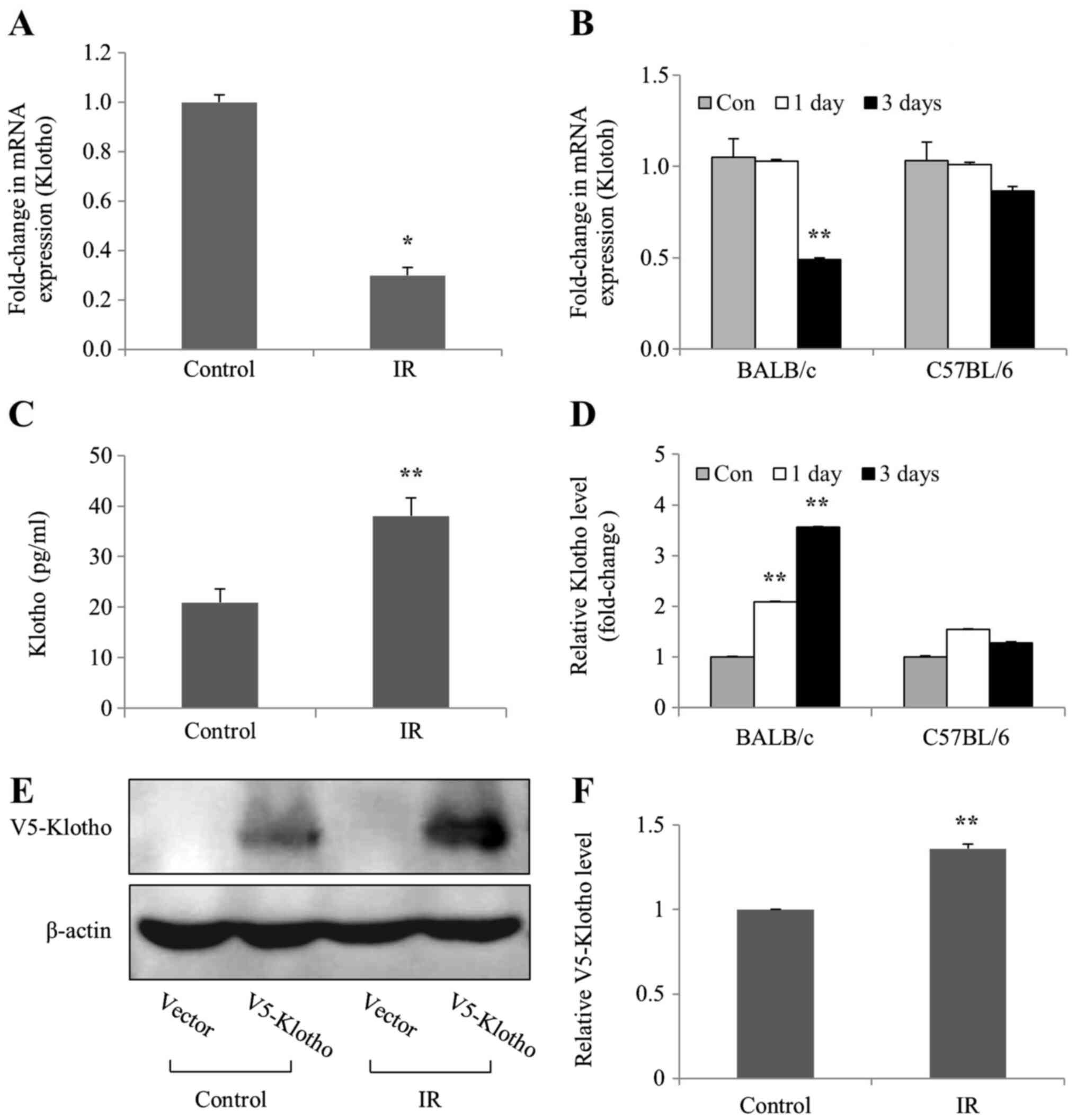
mRNA expression levels of Klotho are
decreased in irradiated kidney cells and tissues of mice
To investigate whether levels of the aging
suppressor Klotho were affected by IR in vivo and in
vitro, the mRNA expression and protein levels of Klotho were
examined in irradiated mIMCD-3 cells, and in the kidney tissues of
BALB/c and C57BL/6 mice. The mRNA expression levels of Klotho were
markedly decreased in irradiated mIMCD-3 cells, and in BALB/c mice
(Fig. 3A and B), as determined by
RT-qPCR. By contrast, ELISA detected considerably increased Klotho
protein levels in mIMCD-3 cells and BALB/c mice following
radiation; however, Klotho protein levels were slightly increased
on days 1 and 3 compared with the control in C57BL/6 mice (Fig. 3C and D). Transfection experiments
were performed to confirm the action site (membrane-anchored or
soluble secreted) of the Klotho protein expressed in the cells.
Consistent with ELISA findings, the results of western blot
analysis revealed increased levels of exogenous V5-tagged Klotho
(membrane-anchored) protein in irradiated mIMCD-3 cells (Fig. 3E and F). Notably, these findings
suggested that the mRNA expression levels of Klotho were decreased,
whereas the expression levels of the corresponding protein were
increased in irradiated mIMCD-3 cells and BALB/c mice.
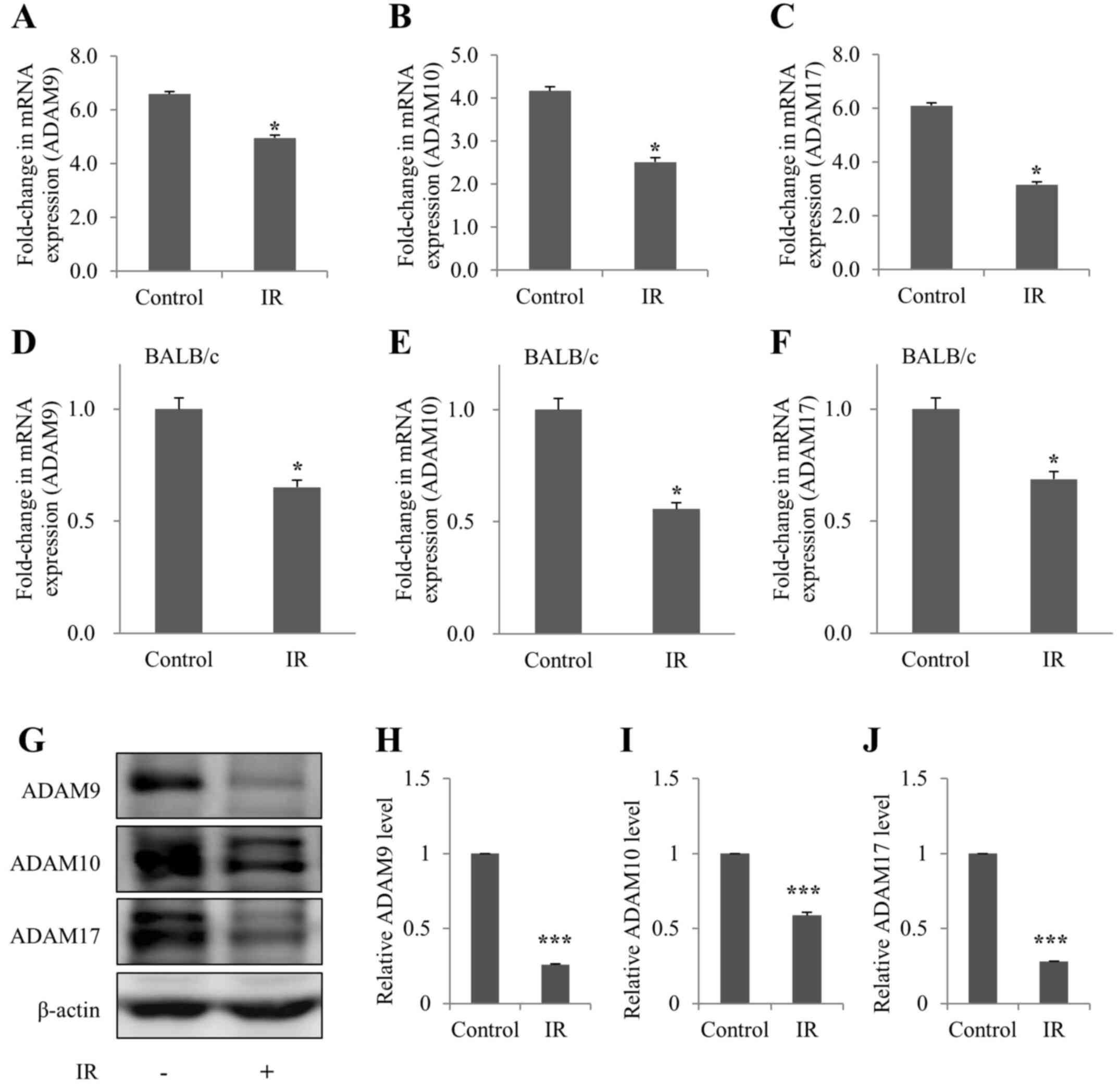
ADAM9/10/17 levels are decreased in
irradiated kidney cells and tissues of mice
The mRNA expression levels of ADAM9/10/17 were
subsequently detected in irradiated mIMCD-3 cells, and in the
kidney tissues of BALB/c mice, in order to determine whether
ADAM9/10/17, an ectodomain shedding enzyme of Klotho, was affected
by IR in vivo and in vitro. As determined by RT-qPCR,
the mRNA expression levels of ADAM9/10/17 were decreased in
irradiated mIMCD-3 cells and BALB/c mice (Fig. 4A-F). In addition, ADAM9/10/17
protein expression was decreased in irradiated mIMCD-3 cells
(Fig. 4G-J), as determined via
western blot analysis.
Discussion
There is growing awareness that radiation induces
injury in the normal tissue of organs that are late to respond to
radiation, such as the brain, kidneys and lungs, involving complex
and dynamic responses between multiple cell types that lead not
only to targeted cell death, but also to acute and chronic
alterations in cell function (20).
The pathogenesis of radiation-induced renal dysfunction is
multifaceted and is associated with the sequential interaction of
all cell types, including vascular, tubular and glomerular cells,
in the kidney. In particular, renal tubular cells and the
surrounding interstitium are notably radiosensitive (1,21–23).
Data from the present study revealed that irradiation induced
cellular senescence in mIMCD-3 mouse kidney cells. Among the genes
associated with cellular senescence, the mRNA expression levels of
the aging suppressor, Klotho (24),
were decreased and its negative regulator, TNF-α (11,25),
were increased in irradiated kidney cells and mouse kidney
tissues.
Cellular senescence is a common response of normal
cells to radiation, which contributes to tissue injury (26). A previous study demonstrated the
increased activity of senescence pathways in multiple kidney
diseases (27). In addition, a
reduction in the number of renal senescent cells via replacement
with non-senescent cells has been reported to decrease
aging-related injury (28). In the
present study, the expression levels of HP1γ, a commonly used
marker of senescence, were decreased in mIMCD-3 cells.
Additionally, irradiation induced downregulation of SIRT1 protein,
supporting the concept that radiation exposure may promote cellular
senescence of mIMCD-3 kidney cells.
Senescent cells secrete various proinflammatory
cytokines associated with normal tissue injury following
irradiation (8). In previous
studies, Klotho suppressed TNF-α-induced expression of adhesion
molecules in endothelial cells (24) and proinflammatory cytokines reduced
Klotho expression in the kidney (29), supporting the association of Klotho
with inflammation in kidney cells. Klotho is an aging suppressor
protein that is abundant in kidney tissue and Klotho-knockout mice
have been shown to develop a syndrome similar to premature aging
(30,31). The present study focused on
determining the relevance of radiation-induced cellular senescence
and the involvement of Klotho. Despite recent focus on the role of
Klotho in the kidney, its precise functions with regards to
pathophysiological implications remain to be established.
Klotho is a transmembrane protein whose
extracellular domain is cleaved by the ectodomain shedding enzyme,
ADAM9/10/17, to generate large amounts of soluble protein into the
blood, urine and cerebrospinal fluid. The Klotho protein exists as
both membrane-anchored and soluble secreted forms (30,32)
with potentially distinct functions. Membrane Klotho forms a
complex with fibroblast growth factor (FGF) receptors (FGFRs) and
functions as a coreceptor for FGF23, a bone-derived hormone that
controls mineral homeostasis. Soluble Klotho functions as an
endocrine factor with important roles in numerous processes,
including antisenescence, energy metabolism, inhibition of Wnt
signaling, antioxidation, and modulation of ion transport
independently of FGF23 and FGFRs (33–35).
In particular, the present study showed that while Klotho gene
transcription was downregulated, the corresponding translated
protein was consistently expressed after radiation exposure. In
addition, it was confirmed that the location of the expressed
Klotho protein was not truncated and was anchored on the cell
membrane with increased levels of the exogenous V5-tagged Klotho
(membrane-anchored) protein detected after mIMCD-3 cells radiation
exposure. ADAMs are membrane-anchored cell surface and secreted
proteins containing disintegrin and metalloproteinase domains that
are critical for regulation of cell phenotypes via effects on cell
adhesion, migration, proteolysis, and modulation of cell signaling
and biological responses (36). In
the present study, the expression of the ectodomain shedding enzyme
of Klotho, ADAM9/10/17, was reduced following exposure of the
kidney to radiation. However, BALB/c mice are sensitive to
radiation and C57BL/6 mice are resistant (37,38);
therefore, this study evaluated ADAMs expression in BALB/c mice.
Differences in radiation-induced genomic instability between these
mouse strains necessitate further study of the expression levels of
ADAMs that respond differently to radiation-sensitive or resistant
mouse strains. Notably, a decline in blood Klotho levels has
previously been detected in patients with aging-related disorders,
such as cardiovascular disease, cancer, hypertension and kidney
disease (39). The present findings
suggested that a reduction in Klotho gene expression, along with
disruption of physiological cleavage of Klotho via inhibition of
ADAM, may contribute to radiation-induced renal dysfunction via the
acceleration of senescence.
Commonly, damage of renal tubules is the ultimate
pathway leading to end-stage kidney disease. Accumulating studies
have suggested that the pathogenesis of radiation-induced
nephropathy is multifaceted and involves the unpredictable
interactions of all cell types of the kidney, as well as a number
of various inflammatory elements in the kidney (21,22,40,41).
Critically, some types of soluble Klotho may exist in the blood,
urine and other body fluids, where it performs a multitude of
functions in renal inflammatory response. However these pleiotropic
roles are poorly understood (8,41–43).
Moreover, the differences between BALB/c and C57BL/6 mice with
regards to changes in TNF-α and ADAM9/10/17 expression caused by
irradiation may be assessed in future research. The differences in
mouse strains with regards to inflammation and tubular cell
responses during damage may provide compelling potential targets
for additional studies of responsiveness to the development of
radiation-induced nephropathy (44–47).
In conclusion, to the best of our knowledge, the
present study is the first to indicate that radiation-induced renal
dysfunction may be associated with TNF-α-mediated inhibition of
Klotho gene expression, which could accelerate cellular senescence
of the kidney. Based on the current data, it may be suggested that
kidney dysfunction due to irradiation is caused by decreased
production of soluble Klotho via reduced expression of ADAM9/10/17.
These novel findings provide insights into the pathogenic
mechanisms underlying radiation-induced kidney injury, which should
facilitate the development of treatment strategies and support the
efficacy of Klotho as a biomarker. Therefore, Klotho may not only
be useful as a diagnostic and/or prognostic marker, but could
present a promising target to prevent and retard radiation-induced
kidney injury. Further investigation of the functional involvement
of Klotho-mediated renal tubular injury is imperative and will be
the main point of further study, along with overcoming limitations,
such as the use of a single cell line, a limited number of animals,
pathological mechanism studies, and experimental animal model
selection based on mouse strain difference. Therefore, future
studies are focusing on identifying the mechanisms of action of
ADAMs, and the various forms of Klotho that perform numerous
functions, in order to understand the mechanisms underlying
translational modification in irradiated kidney cells/tissues and
serum.
Acknowledgements
Not applicable.
Funding
This research was supported by a grant from the
Korea Institute of Radiological and Medical Sciences (KIRAMS),
funded by the Ministry of Science, ICT and Future Planning,
Republic of Korea (grant no. 50531-2020).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
EJK, ML and DYK designed the study, performed most
of the experiments, analyzed the data, and wrote the manuscript.
EJK and ML analyzed the data and provided advice on the project.
EJK supervised the project and provided financial support. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
The experimental protocol was conducted under the
Guidelines for the Use and Care of Laboratory Animals and the study
was approved by the IACUC of the Korea Institute of Radiological
and Medical Sciences (KIRAMS; IACUC approval no. 2016-0065; Seoul,
South Korea).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Cohen EP and Robbins ME: Radiation
nephropathy. Seminars in nephrology Elsevier. 486–499. 2003.
View Article : Google Scholar
|
|
2
|
Baradaran-Ghahfarokhi M: Radiation-induced
kidney injury. J Renal Inj Prev. 1:49–50. 2012.PubMed/NCBI
|
|
3
|
Little MP: A review of non-cancer effects,
especially circulatory and ocular diseases. Radiat Environ Biophys.
52:435–449. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wei J, Wang B, Wang H, Meng L, Zhao Q, Li
X, Xin Y and Jiang X: Radiation-induced normal tissue damage:
oxidative stress and epigenetic mechanisms. Oxid Med Cell Longev.
2019:30103422019. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Salminen A, Kauppinen A and Kaarniranta K:
Emerging role of NF-κB signaling in the induction of
senescence-associated secretory phenotype (SASP). Cell Signal.
24:835–845. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sturmlechner I, Durik M, Sieben CJ, Baker
DJ and van Deursen JM: Cellular senescence in renal ageing and
disease. Nat Rev Nephrol. 13:77–89. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Malaquin N, Tu V and Rodier F: Assessing
Functional Roles of the Senescence-Associated Secretory Phenotype
(SASP). Cellular Senescence. Springer; pp. 45–55. 2019, View Article : Google Scholar
|
|
8
|
Citrin DE and Mitchell JB: Mechanisms of
normal tissue injury from irradiation. Semin Radiat Oncol.
27:316–324. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hill RP, Zaidi A, Mahmood J and Jelveh S:
Investigations into the role of inflammation in normal tissue
response to irradiation. Radiother Oncol. 101:73–79. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kitagawa M, Sugiyama H, Morinaga H, Inoue
T, Takiue K, Ogawa A, Yamanari T, Kikumoto Y, Uchida HA, Kitamura
S, et al: A decreased level of serum soluble Klotho is an
independent biomarker associated with arterial stiffness in
patients with chronic kidney disease. PLoS One. 8:e566952013.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Thurston RD, Larmonier CB, Majewski PM,
Ramalingam R, Midura-Kiela M, Laubitz D, Vandewalle A, Besselsen
DG, Mühlbauer M, Jobin C, et al: Tumor necrosis factor and
interferon-γ down-regulate Klotho in mice with colitis.
Gastroenterology. 138:1384–1394, 1394.e1-2. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hu MC, Kuro-o M and Moe OW: Klotho and
kidney disease. J Nephrol. 23 (Suppl 16):S136–S144. 2010.PubMed/NCBI
|
|
13
|
Komaba H and Lanske B: Vitamin D and
Klotho in Chronic Kidney Disease. Vitamin D in Chronic Kidney
Disease. Springer Cham; pp. 179–194. 2016, View Article : Google Scholar
|
|
14
|
Erin N, Türker S, Elpek Ö and Yildirim B:
ADAM proteases involved in inflammation are differentially altered
in patients with gastritis or ulcer. Exp Ther Med. 15:1999–2005.
2018.PubMed/NCBI
|
|
15
|
van Loon EP, Pulskens WP, van der Hagen
EA, Lavrijsen M, Vervloet MG, van Goor H, Bindels RJ and Hoenderop
JG: Shedding of klotho by ADAMs in the kidney. Am J Physiol Renal
Physiol. 309:F359–F368. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kim EJ, Lee M, Kim DY, Kim KI and Yi JY:
Mechanisms of energy metabolism in skeletal muscle mitochondria
following radiation exposure. Cells. 8:9502019. View Article : Google Scholar
|
|
17
|
McRobb LS, McKay MJ, Gamble JR, Grace M,
Moutrie V, Santos ED, Lee VS, Zhao Z, Molloy MP and Stoodley MA:
Ionizing radiation reduces ADAM10 expression in brain microvascular
endothelial cells undergoing stress-induced senescence. Aging
(Albany NY). 9:1248–1268. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Slizhov P, Dolinina T, Pleskach N,
Vasilishina AA, Zherebtsov SV, Bulatnikova MA, Mikhelson VM and
Spivak IM: Markers of aging in cells of patients with cockayne
syndrome. General and individual differences. Cell Tissue Biol.
12:296–306. 2018. View Article : Google Scholar
|
|
20
|
Zhao W, Chuang EY, Mishra M, Awwad R,
Bisht K, Sun L, Nguyen P, Pennington JD, Wang TJ, Bradbury CM, et
al: Distinct effects of ionizing radiation on in vivo murine kidney
and brain normal tissue gene expression. Clin Cancer Res.
12:3823–3830. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Glatstein E, Fajardo LF and Brown JM:
Radiation injury in the mouse kidney - I. Sequential light
microscopic study. Int J Radiat Oncol Biol Phys. 2:933–943. 1977.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Williams MV: The cellular basis of renal
injury by radiation. Br J Cancer Suppl. 7:257–264. 1986.PubMed/NCBI
|
|
23
|
Jaggi JS, Seshan SV, McDevitt MR, Sgouros
G, Hyjek E and Scheinberg DA: Mitigation of radiation nephropathy
after internal alpha-particle irradiation of kidneys. Int J Radiat
Oncol Biol Phys. 64:1503–1512. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Maekawa Y, Ishikawa K, Yasuda O, Oguro R,
Hanasaki H, Kida I, Takemura Y, Ohishi M, Katsuya T and Rakugi H:
Klotho suppresses TNF-α-induced expression of adhesion molecules in
the endothelium and attenuates NF-kappaB activation. Endocrine.
35:341–346. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Navarro-González JF, Sánchez-Niño MD,
Donate-Correa J, Martín-Núñez E, Ferri C, Pérez-Delgado N, Górriz
JL, Martínez-Castelao A, Ortiz A and Mora-Fernández C: Effects of
pentoxifylline on soluble Klotho concentrations and renal tubular
cell expression in diabetic kidney disease. Diabetes Care.
41:1817–1820. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hong EH, Lee SJ, Kim JS, Lee KH, Um HD,
Kim JH, Kim SJ, Kim JI and Hwang SG: Ionizing radiation induces
cellular senescence of articular chondrocytes via negative
regulation of SIRT1 by p38 kinase. J Biol Chem. 285:1283–1295.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Muñoz-Espín D and Serrano M: Cellular
senescence: From physiology to pathology. Nat Rev Mol Cell Biol.
15:482–496. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yang H and Fogo AB: Cell senescence in the
aging kidney. J Am Soc Nephrol. 21:1436–1439. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Moreno JA, Izquierdo MC, Sanchez-Niño MD,
Suárez-Alvarez B, Lopez-Larrea C, Jakubowski A, Blanco J, Ramirez
R, Selgas R, Ruiz-Ortega M, et al: The inflammatory cytokines TWEAK
and TNFα reduce renal klotho expression through NFκB. J Am Soc
Nephrol. 22:1315–1325. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
John GB, Cheng C-Y and Kuro-o M: Role of
Klotho in aging, phosphate metabolism, and CKD. Am J Kidney Dis.
58:127–134. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Xu Y and Sun Z: Molecular basis of Klotho:
From gene to function in aging. Endocr Rev. 36:174–193. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Pavik I, Jaeger P, Ebner L, Wagner CA,
Petzold K, Spichtig D, Poster D, Wüthrich RP, Russmann S and Serra
AL: Secreted Klotho and FGF23 in chronic kidney disease Stage 1 to
5: A sequence suggested from a cross-sectional study. Nephrol Dial
Transplant. 28:352–359. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lu X and Hu MC: Klotho/FGF23 axis in
chronic kidney disease and cardiovascular disease. Kidney Dis.
3:15–23. 2017. View Article : Google Scholar
|
|
34
|
Kuro-O M: The Klotho proteins in health
and disease. Nat Rev Nephrol. 15:27–44. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Hu MC, Kuro-o M and Moe OW: Klotho and
chronic kidney disease. Contrib Nephrol. 180:47–63. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Palau V, Pascual J, Soler MJ and Riera M:
Role of ADAM17 in kidney disease. Am J Physiol Renal Physiol.
317:F333–F342. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Ponnaiya B, Cornforth MN and Ullrich RL:
Radiation-induced chromosomal instability in BALB/c and C57BL/6
mice: The difference is as clear as black and white. Radiat Res.
147:121–125. 1997. View
Article : Google Scholar : PubMed/NCBI
|
|
38
|
Hamasaki K, Imai K, Hayashi T, Nakachi K
and Kusunoki Y: Radiation sensitivity and genomic instability in
the hematopoietic system: Frequencies of micronucleated
reticulocytes in whole-body X-irradiated BALB/c and C57BL/6 mice.
Cancer Sci. 98:1840–1844. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Kim JH, Hwang KH, Park KS, Kong ID and Cha
SK: Biological role of anti-aging protein Klotho. J Lifestyle Med.
5:1–6. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kal HB and van Kempen-Harteveld ML: Renal
dysfunction after total body irradiation: dose-effect relationship.
Int J Radiat Oncol Biol Phys. 65:1228–1232. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Mitobe M, Yoshida T, Sugiura H, Shirota S,
Tsuchiya K and Nihei H: Oxidative stress decreases klotho
expression in a mouse kidney cell line. Nephron, Exp Nephrol.
101:e67–e74. 2005. View Article : Google Scholar
|
|
42
|
Kala J: Radiation-induced kidney injury. J
Onco-Nephrology. 3:160–167. 2019. View Article : Google Scholar
|
|
43
|
Fujiu K, Manabe I and Nagai R: Renal
collecting duct epithelial cells regulate inflammation in
tubulointerstitial damage in mice. J Clin Invest. 121:3425–3441.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Puri TS, Shakaib MI, Chang A, Mathew L,
Olayinka O, Minto AW, Sarav M, Hack BK and Quigg RJ: Chronic kidney
disease induced in mice by reversible unilateral ureteral
obstruction is dependent on genetic background. Am J Physiol Renal
Physiol. 298:F1024–F1032. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
van Kooten C, Daha MR and van Es LA:
Tubular epithelial cells: A critical cell type in the regulation of
renal inflammatory processes. Exp Nephrol. 7:429–437. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Jevnikar AM, Brennan DC, Singer GG, Heng
JE, Maslinski W, Wuthrich RP, Glimcher LH and Kelley VE: Stimulated
kidney tubular epithelial cells express membrane associated and
secreted TNF α. Kidney Int. 40:203–211. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Black RA, Rauch CT, Kozlosky CJ, Peschon
JJ, Slack JL, Wolfson MF, Castner BJ, Stocking KL, Reddy P,
Srinivasan S, et al: A metalloproteinase disintegrin that releases
tumour-necrosis factor-α from cells. Nature. 385:729–733. 1997.
View Article : Google Scholar : PubMed/NCBI
|