Introduction
Pancreatic cancer (PC) is characterized by high
mortality, low survival rate and poor prognosis. The statistics of
the global cancer database (GLOBOCAN) in 2018 revealed 458,918 new
patients with PC and 432,242 mortalities (1). PC incidence and mortality trends over
the period of 2018–2040 are hypothesized to demonstrate an
increasing trend of incidence (+77.7% with 356,358 new cases) and
mortality (+79.9%, 345,181). Hence, PC ranks 14th in terms of
cancer incidence and 7th in terms of cancer mortality (2). Despite the continuous development of
medical diagnosis and treatment technology, the 5-year survival
rate of PC patients remains <10% and the median overall survival
period is only 6–9 months (3). The
cause of this clinical diagnosis and treatment problem is the
unclear understanding of the relevant mechanism of PC cell death.
Existing research results are not ideal to improve the prognosis of
PC, and effective targets for inducing PC cell death remain lacking
(4). Therefore, exploring the
mechanism of PC cell death, the key target genes and related
molecular biological mechanisms is an attractive focus for
research.
In 2012, Dixon et al (5) first reported a new type of cell death
mode other than apoptosis, necrosis and autophagy. This new type is
characterized by the accumulation of reactive oxygen species (ROS)
produced by iron metabolism and lipid peroxidation products. ROS
causes oxidative damage to tissue cells and leads to cell death
(6). As the process of the
occurrence of ROS requires cells to be rich in available iron, the
researchers termed it ferroptosis (5). This new mode of cell death has
received an increasing amount of attention from researchers in
recent years (7–11). Ferroptosis occurs in a variety of
tumors, including PC, and may possess an anticancer effect
(4,12–14).
However, the occurrence and regulatory mechanism of ferroptosis in
PC remain to be elucidated. Identifying new ways to treat PC has
been challenging. Gemcitabine, a first-line drug, is used alone or
in combination for the treatment of patients with advanced
pancreatic ductal adenocarcinoma (15). Heat shock 70-kDa protein 5 improves
the anticancer activity of gemcitabine by inducing ferroptosis
(16). As well as gemcitabine,
certain traditional Chinese herbs induce ferroptosis in PC.
Furthermore, a number of molecules have been demonstrated to induce
ferroptosis in PC cells. These may offer new options for PC
treatment (4).
KAI1/CD82, which expresses membrane proteins, is an
important tumor suppressor gene. It is a prostate cancer
metastasis-related suppressor gene discovered by Dong et al
in 1995 (17). It belongs to the
transmembrane 4 superfamily family and is located in the human
chromosome 1lpl 1.2. It consists of 10 exons and 9 introns and is
approximately 80 kb in length (17). The KAI1 gene is used to inhibit
metastasis in PC (18–23). This gene has been recently
identified to regulate the autophagy of PC cells (24,25). A
number of studies have provided evidence to support the association
between autophagy and ferroptosis in human diseases (26,27).
Therefore, studying the regulatory effect of the KAI1 gene on
ferroptosis in PC is warranted.
In the present study, the roles of KAI1 and
ferroptosis in the onset of PC were evaluated in vitro. The
objective was to provide evidence related to the KAI1 gene and its
potential as a novel therapy target.
Materials and methods
Cell culture
Human PC cell lines of MIA PaCa-2 were provided by
the Shanghai Institute of Cell Biology, Chinese Academy of
Sciences. The cells were grown as sub-confluent monolayers in
Dulbecco's modified Eagle medium (Cytiva) supplemented with 10%
fetal calf serum (Cytiva), 2 mmol/l L-glutamine, 100 IU/ml
penicillin G and 100 µg/ml streptomycin (Beijing Solarbio Science
& Technology Co., Ltd.) in an incubator (37°C, 5%
CO2 and saturated humidity). Cells were passaged at 85%
confluence or were harvested for western blot analysis.
Cell transfection
KAI1 overexpression plasmid (pCMV-KAI1 DNA) was
obtained from Dr Dong, Emory University School of Medicine,
Atlanta, Georgia, USA (28) as a
gift. According to a previous study (22), pCMV-KAI1 DNA and pCMV DNA [empty
pCMV vector (Beijing Biosynthesis Biotechnology Co., Ltd.)] were
transfected with a low expression cell line of KAI1: MIA PaCa-2.
Cells (1×105) were plated in 96-well plates. When the
cells reached 90–95% confluence, 1.0 µg pCMV-KAI1 DNA or pCMV DNA,
1 µl Lipofectamine® 2000 reagent (Invitrogen; Thermo
Fisher Scientific, Inc.), and 100 µl medium without serum were
mixed and incubated for 15 min at room temperature and then added
to each well. Cells were diluted to 1,000 cells/ml following
transfection. The lowest concentration of G418 was selected in the
concentration range of 100–1,000 µg/ml. Positive clone screening
was performed with 100 µg/ml of G418 for 4 weeks (21–23).
The transfected cells were obtained through screening and passage.
At 85% confluence, cells were lysed using radioimmunoprecipitation
assay buffer (Beijing Solarbio Science & Technology Co., Ltd.).
The expression of the KAI1 protein was confirmed by western blot
analysis.
Detection of cell viability by Cell
Counting Kit-8 (CCK-8) assay
Cells in the logarithmic phase were divided into
liproxstein-treated (liproxstein+) and non-liproxstein-treated
(liproxstein-) groups and then inoculated into 6-well plates for 24
h. Cells were washed twice with PBS and digested with trypsin (2.5
g/l; containing 0.2 g/l EDTA). Cells were collected and centrifuged
at 104.5 × g for 5 min at room temperature. The supernatant was
discarded and the medium was gently mixed by pipetting. Cells were
inoculated into 96-well plates following adjustment of the final
concentration to 1×105 cells/ml (100 µl in each well).
After 24 h of incubation, 10 µl CCK-8 solution was added, and
incubation was performed in an incubator for 2 h. The absorbance
was measured at a wavelength of 450 nm using a Multiskan FC
microplate reader (Thermo Fisher Scientific, Inc.).
Detection of intracellular ROS by flow
cytometric analysis
Cell suspensions from the logarithmic cells were
inoculated into 6-well plates at 2 ml per well. When the cells
completely adhered to the wall, the old culture medium was
discarded. The cells were washed twice with PBS and mixed with
serum-free culture medium containing H2DCFH-DA fluorescent dye
(Beijing Solarbio Science & Technology Co., Ltd.) for 30 min in
suspension according to the manufacturer's protocol. The cells were
incubated for 20 min in the dark at room temperature and then
washed twice with precooled PBS to remove the excess dye. Cells
were collected and mixed gently with 700 µl PBS. The mean
fluorescence intensity of intracellular ROS was detected
immediately by flow cytometry (excitation wavelength: 485 nm;
emission wavelength: 530 nm) using a BD FACSAria II flow cytometer
(BD Biosciences). The results were analyzed with FlowJo software
(version 6.1.1; FlowJo LLC).
Western blot analysis
The total proteins of logarithmic growth cells were
lysed using radioimmunoprecipitation assay buffer (Beijing Solarbio
Science & Technology Co., Ltd.) and collected. Protein
concentration was determined with the bicinchoninic acid method
(Beijing Solarbio Science & Technology Co., Ltd.). Each sample
(30 µg/lane) was resolved by 12% sodium dodecyl
sulphate-polyacrylamide gel electrophoresis, followed by transfer
onto a polyvinylidene difluoride membranes (EMD Millipore). The
membranes were then blocked with 5% skimmed milk for 2 h at room
temperature. Membranes were incubated with rabbit polyclonal
anti-KAI1 (1:200 dilution; cat. no. sc-7179; Santa Cruz
Biotechnology, Inc.), rabbit polyclonal anti-ferroportin (FPN;
1:100 dilution; cat. no. bs-3579R; BIOSS), rabbit polyclonal
anti-glutathione peroxidase 4 (GPX4; 1:100 dilution; cat no
bs-3884R; BIOSS) or rabbit polyclonal anti-GAPDH (1:200 dilution;
cat. no. bs-0755R; BIOSS) primary antibodies overnight at 4°C,
followed by horseradish peroxidase-conjugated goat anti-rabbit IgG
secondary antibody (1:2,000 dilution; cat. no. sc-2749; Santa Cruz
Biotechnology, Inc.) for 1 h at room temperature. Immunoreactive
bands were visualized using a Western Lightning Chemiluminescence
Reagent Plus kit (PerkinElmer, Inc.). The signals were detected
using a Las-4000 mini CCD camera (GE Healthcare). With GAPDH as the
internal reference, the relative expression of protein was analyzed
by densitometry using Quantity One 1-D analysis software (version
4.6.9; Bio-Rad Laboratories, Inc.) following internal reference
calibration. Protein expression was derived from the grey ratio of
the target band to the internal reference band.
Statistical analysis
Data were expressed as the mean ± standard deviation
of at least three independent experiments. Statistical analysis
within groups was performed using repeated measures of analysis of
variance followed by Bonferroni's post hoc test. Between groups
analysis was performed using Student's paired t-test. SPSS 16.0
(SPSS, Inc.) was used for all statistical analyses. P<0.05 was
considered to indicate a statistically significant difference.
Results
Construction of KAI1 stable
expression
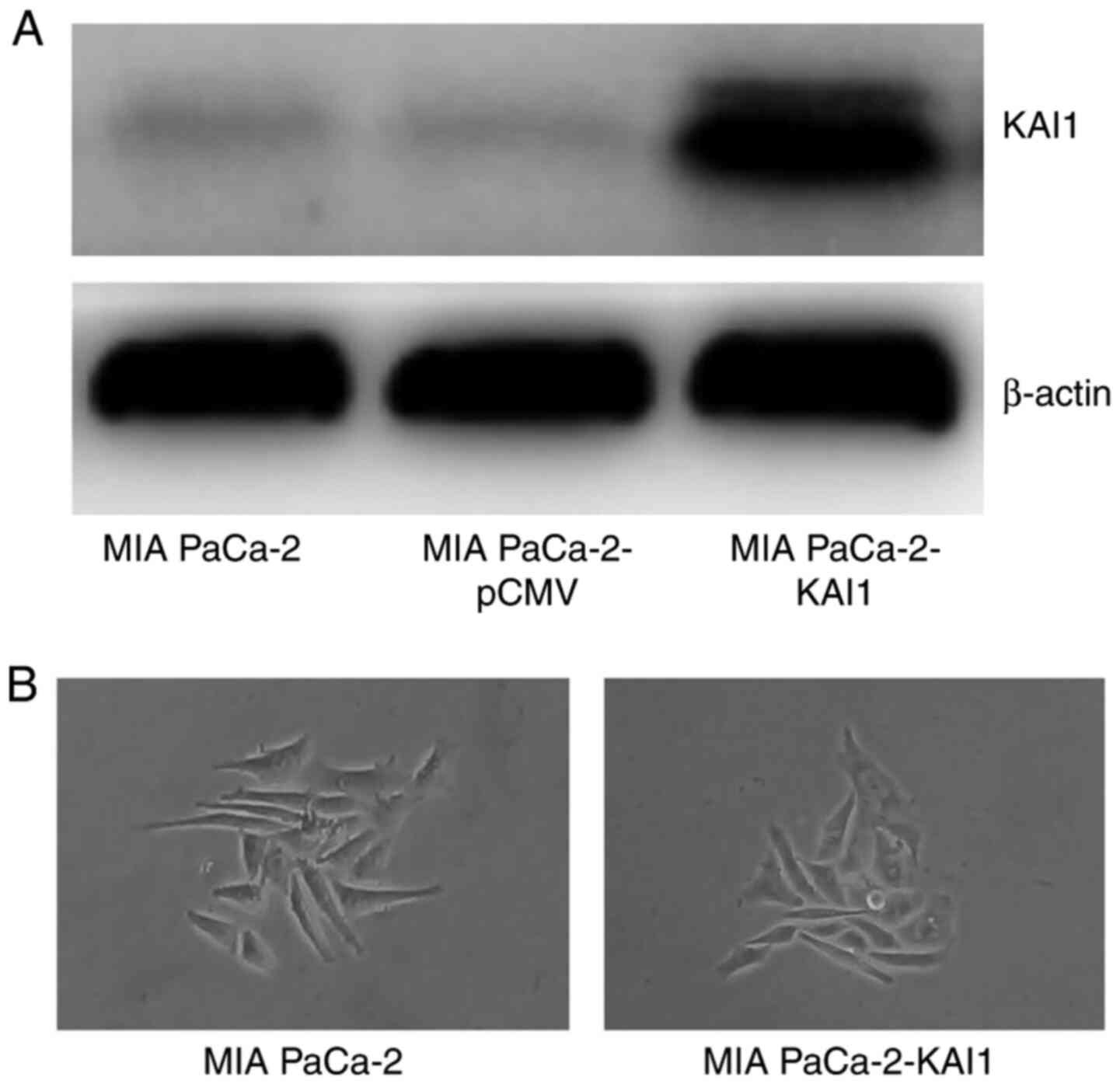
Western blot analysis revealed that the MIA PaCa-2
line transfected with KAI1 plasmid exhibited strong protein bands,
whereas the untransfected cells and transfected with empty pCMV
vector cells exhibited weak bands (Fig.
1A). MIA PaCa-2-KAI1 cell morphology did not show the
scattering and acquisition of fusiform and fibroblastic phenotype
compared with MIA PaCa-2 cells (Fig.
1B). The MIA PaCa-2 cells that demonstrated a stable expression
of the KAI1 gene were labelled as MIA PaCa-2-KAI1.
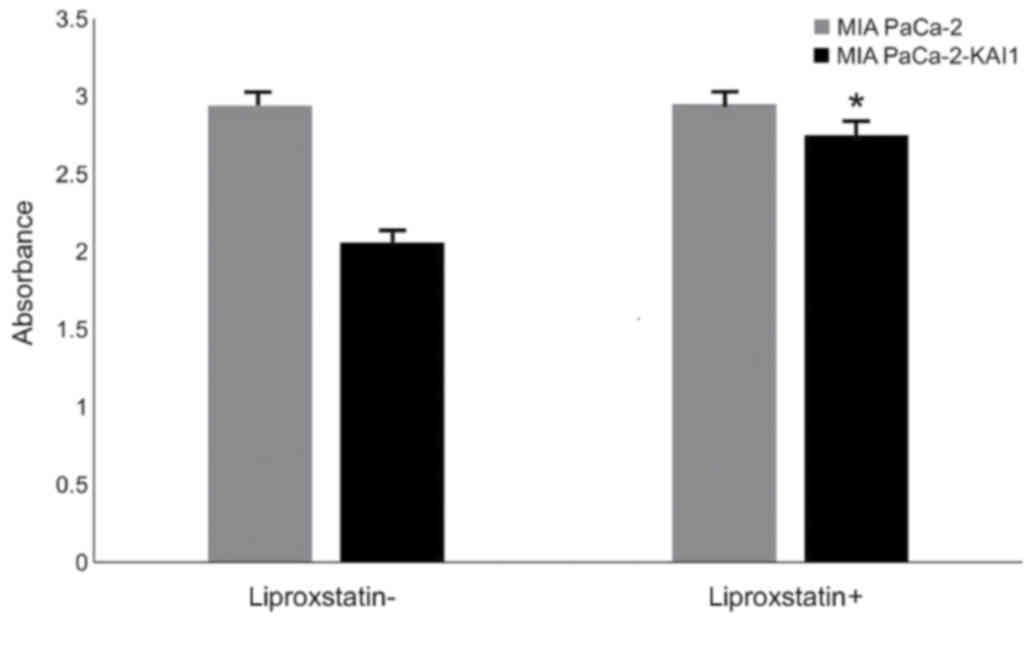
Cell viability
CCK-8 detection revealed that KAI1 overexpression
had no significant effect on the viability of MIA PaCa-2. Following
treatment with liproxstein, a type of ferroptosis inhibitor, for 24
h, the increased viability of MIA PaCa-2-KAI1 (from 2.06±0.02 to
2.75±0.02) was more apparent compared with that of MIA PaCa-2 (from
2.94±0.02 to 2.95±0.02; P<0.05; Fig.
2), thereby indicating that KAI1 may promote ferroptosis.
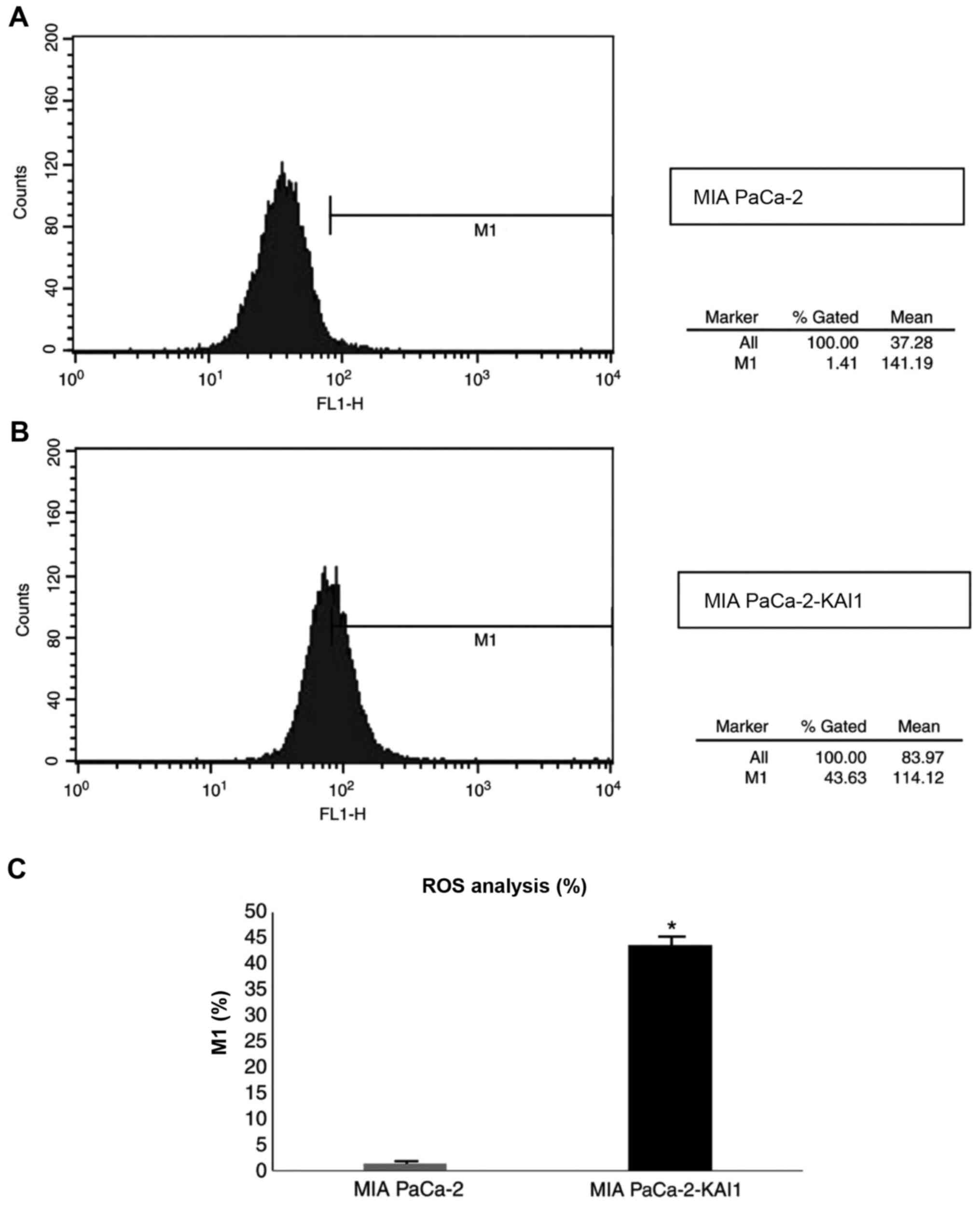
ROS are affected by KAI1
As demonstrated by flow cytometry results (Fig. 3), the fluorescence intensity of ROS
in MIA PaCa-2-KAI1 (43.63%) was significantly higher compared with
that in MIA PaCa-2 (1.42%; P<0.05). This suggested that KAI1
overexpression can induce ROS production in MIA PaCa-2 cells.
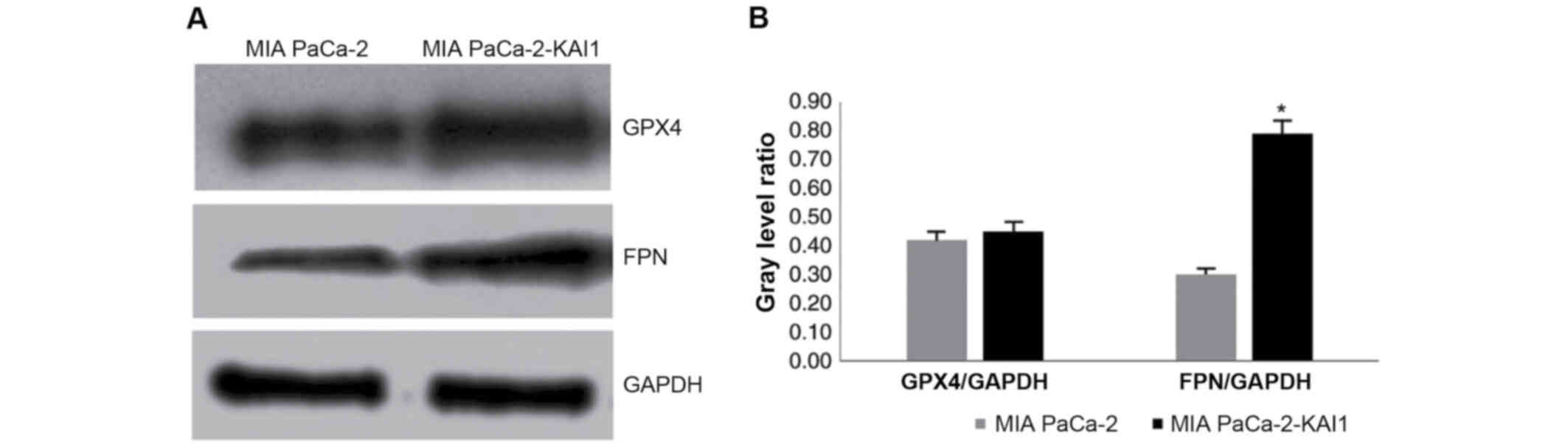
KAI1 gene promotes FPN and GPx4
protein expressions
Western blot analysis was used to detect the
proteins in the MIA PaCa-2 and MIA PaCa-2-KAI1 cells. FPN and GPX4
protein expressions were higher in MIA PaCa-2-KAI1 compared with
MIA PaCa-2 (Fig. 4A). The
comparison of the grey values, with GAPDH protein as the internal
control, indicated that KAI1 promoted FPN protein expression
(Fig. 4B).
Discussion
Previous studies have shown that ferroptosis
inhibits the growth and proliferation of PC cells, thereby
providing a new approach for the treatment of PC (18–25).
In the present study, the widely-studied KAI1 gene was the research
object. The KAI1 gene was highly expressed in the MIA PaCa-2 cells
by transfection and the occurrence of ferroptosis was detected. In
the CCK-8 experiment, it was identified that the viability of MIA
PaCa-2 cells was insignificantly inhibited by KAI1 overexpression
and that the inhibition was weakened by further treatment with the
ferroptosis inhibitor liproxstein. It was hypothesized that the
inhibitor blocked KAI1-induced ferroptosis. KAI1 induced the MIA
PaCa-2 cells to produce ROS extensively, as detected by flow
cytometry. Previous studies have confirmed that ROS serve an
important role in promoting ferroptosis (29–32).
Previous research in our laboratory demonstrated that KAI1 induced
autophagy in MIA PaCa-2 cells (24,25).
Previous studies have shown that the activation of the autophagy
pathway promotes ferroptosis in fibroblasts and cancer cells
(26,27). On the basis of the experimental
results from the present study and literature reports, it was
hypothesized that KAI1 overexpression can promote the ferroptosis
of PC cells.
To identify the possible mechanism underlying this
promoting effect, the changes in cell protein expression was
detected. Genes, such as FPN, GPX4, system Xc-, transferrin,
transferrin receptor 1, divalent metal transporter 1, ferritin
heavy chain 1, and ferritin lightchain, are the critical mediators
in the ferroptosis procedure (33).
FPN and GPX4 were markedly expressed from the MIA PaCa-2 cells,
which highly express KAI1. In addition, the expression of FPN was
greatly increased according to the densitometry analysis. Previous
studies have confirmed that FPN is a multidomain transmembrane
protein encoded by SLC40A and is the only existing transmembrane
iron export protein (6,34). Studies on a variety of tumor cells
have shown that the high expression of FPN can cause iron outflow,
which can lead to iron deficiency in cells and inhibit cancer cell
growth (35–38). Therefore, the results from the
present study, indicating the promotion of FPN expression by KAI1,
further confirmed that KAI1 overexpression can inhibit the
proliferation of PC cells. In addition, increasing iron intake and
reducing iron storage may lead to iron overload, which may in turn
lead to ferroptosis (39,40). Therefore, it was hypothesized that
the high expression of KAI1 promoted FPN expression and led to the
transportation of excessive Fe2+ out of the cells,
thereby causing the cancer cells to absorb large amounts of
Fe3+. This condition breaks the iron homeostasis in
cancer cells. Excessive iron transforms H2O2
and lipoperoxidation into ROS through the Fenton reaction and
finally leads to ferroptosis (6).
The loss of GPX4 activity is another major source of
ROS and is a key factor for inducing ferroptosis (41–43).
The results of the present study demonstrated that KAI1 can promote
the expression of GPX4, although it may inhibit the occurrence of
ferroptosis. Nevertheless, the effect of KAI1 on GPX4 expression
was weaker compared with that on FPN expression. Therefore, the
high expression of the KAI1 gene promoted the occurrence of
ferroptosis in PC cells through its comprehensive effects on FPN
and GPX4. The limitation of the present study that should be
acknowledged when interpreting the results is the lack of research
on the in-depth mechanism underlying the effects of KAI1 on FPN and
GPX4, as well as the relationship between KAI1 and other
ferroptosis-related mediators.
As indicated by the results of the proliferation
experiments, KAI1-induced ferroptosis did not significantly inhibit
the proliferation of PC cells. According to our previous findings
on the KAI1 gene, it inhibits the epithelial-mesenchymal transition
and the migration and lymphangiogenesis of PC cells (23). It was concluded that although the
inhibition of proliferation was not significant, it still limited
the metastasis of PC cells to some extent. The present study added
to the theoretical basis of KAI1 as a tumor suppressor gene and
provided a new experimental basis for its clinical application.
Acknowledgements
The authors would like to thank Dr Dong Jintang
(Emory University School of Medicine, Atlanta, USA) for providing
the KAI1 overexpression plasmid (pCMV-KAI1 DNA).
Funding
The present study was supported by grants from the
Science Foundation of Liaoning (grant no. 2019-MS-352) and the
Science and Technology Foundation of Shenyang (grant no.
19-112-4-046).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XL and XG designed the study. XL and JC performed
the experiments and wrote the first draft. XL, HL and HH collected
and analyzed the data. All authors contributed to the design and
interpretation of the study and to further drafts of the
manuscript. All authors reviewed and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2018. CA Cancer J Clin. 68:7–30. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Rawla P, Sunkara T and Gaduputi V:
Epidemiology of pancreatic cancer: Global trends, etiology and risk
factors. World J Oncol. 10:10–27. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chen G, Guo GQ, Zhou XD and Chen HX:
Potential mechanism of ferroptosis in pancreatic cancer. Oncol
Lett. 19:579–587. 2020.PubMed/NCBI
|
|
5
|
Dixon SJ, Lemberg KM, Lamprecht MR, Skouta
R, Zaitsev EM, Gleason CE, Patel DN, Bauer AJ, Cantley AM, Yang WS,
et al: Ferroptosis: An iron-dependent form of nonapoptotic cell
death. Cell. 5:1060–1072. 2012. View Article : Google Scholar
|
|
6
|
Kajarabille N and Latunde-Dada GO:
Programmed cell-death by ferroptosis: Antioxidants as mitigators.
Int J Mol Sci. 19:49682019. View Article : Google Scholar
|
|
7
|
Hasegawa M, Takahashi H, Rajabi H, Alam M,
Suzuki Y, Yin L, Tagde A, Maeda T, Hiraki M, Sukhatme VP and Kufe
D: Functional interactions of the cystine/glutamate antiporter,
CD44v and MUC1-C oncoprotein in triple-negative breast cancer
cells. Oncotarget. 7:11756–11769. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lin R, Zhang Z, Chen L, Zhou Y, Zou P,
Feng C, Wang L and Liang G: Dihydroartemisinin (DHA) induces
ferroptosis and causes cell cycle arrest in head and neck carcinoma
cells. Cancer Lett. 381:165–175. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Tang M, Chen Z, Wu D and Chen L:
Ferritinophagy/ferroptosis: Iron-related newcomers in human
diseases. J Cell Physiol. 233:9179–9190. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Nie J, Lin B, Zhou M, Wu L and Zheng T:
Role of ferroptosis in hepatocellular carcinoma. J Cancer Res Clin
Oncol. 144:2329–2337. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Xia XJ, Fan XP, Zhao MY and Zhu P: The
relationship between ferroptosis and tumors-a novel landscape for
therapeutic approach. Curr Gene Ther. 19:117–124. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yamaguchi Y, Kasukabe T and Kumakura S:
Piperlongumine rapidly induces the death of human pancreatic cancer
cells mainly through the induction of ferroptosis. Int J Oncol.
3:1011–1022. 2018.
|
|
13
|
Wang K, Zhang Z, Wang M, Cao X, Qi J, Wang
D, Gong A and Zhu H: Role of GRP78 inhibiting artesunate-induced
ferroptosis in KRAS mutant pancreatic cancer cells. Drug Des Devel
Ther. 13:2135–2144. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Song Z, Xiang X, Li J, Deng J, Fang Z,
Zhang L and Xiong J: Ruscogenin induces ferroptosis in pancreatic
cancer cells. Oncol Rep. 43:516–524. 2020.PubMed/NCBI
|
|
15
|
Binenbaum Y, Na'ara S and Gil Z:
Gemcitabine resistance in pancreatic ductal adenocarcinoma. Drug
Resist Updat. 23:55–68. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhu S, Zhang Q, Sun X, Zeh HJ III, Lotze
MT, Kang R and Tang D: HSPA5 regulates ferroptotic cell death in
cancer cells. Cancer Res. 77:2064–2077. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Dong JT, Lamb PW, Rinker-Schaeffer CW,
Vukanovic J, Ichikawa T, Isaacs JT and Barrett JC: KAI1, a
metastasis suppressor gene for prostate cancer on human chromosome
11p11.2. Science. 268:884–886. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Guo XZ, Friess H, Shao XD, Liu MP, Xia YT,
Xu JH and Buchler MW: KAI1 gene is differently expressed in
papillary and pancreatic cancer influence on metastasis. World J
Gastroenterol. 6:866–871. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Xu JH, Guo XZ, Ren LN, Shao LC and Liu MP:
KAI1 is potential target on anti-metastasis in pancreatic cancer
cells. World J Gastroenterol. 14:1126–1132. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Liu X, Guo XZ, Zhang WW, Lu ZZ, Zhang QW,
Duan HF and Wang LS: KAI1 inhibits HGF-induced invasion of
pancreatic cancer by sphingosine kinase activity. Hepatobiliary
Pancreat Dis Int. 10:201–208. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liu X, Guo XZ, Li HY, Chen J, Ren LN and
Wu CY: KAI1 inhibits lymphangiogenesis and lymphatic metastasis of
pancreatic cancer in vivo. Hepatobiliary Pancreat Dis Int.
13:87–92. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Liu X, Guo X, Li H, Chen J and Qi X:
Src/STAT3 signaling pathways are involved in KAI1-induced
downregulation of VEGF-C expression in pancreatic cancer. Mol Med
Report. 13:4774–4778. 2016. View Article : Google Scholar
|
|
23
|
Liu X, Guo XZ, Li HY and Chen J: KAI1
reverses the epithelial-mesenchymal transition in human pancreatic
cancer cells. Hepatobiliary Pancreat Dis Int. 18:471–477. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wu CY, Yan J, Yang YF, Xiao FJ, Li QF,
Zhang QW, Wang LS, Guo XZ and Wang H: Overexpression of KAI1
induces autophagy and increases MiaPaCa-2 cell survival through the
phosphorylation of extracellular signal-regulated kinases. Biochem
Biophys Res Commun. 404:802–808. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wu CY, Guo XZ and Li HY: Hypoxia and serum
deprivation protected MiaPaCa-2 cells from KAI1-induced
proliferation inhibition through autophagy pathway activation in
solid tumors. Clin Transl Oncol. 17:201–208. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hou W, Xie Y, Song X, Sun X, Lotze MT, Zeh
HJ III, Kang R and Tang D: Autophagy promotes ferroptosis by
degradation of ferritin. Autophagy. 12:1425–1428. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhou B, Liu J, Kang R, Klionsky DJ,
Kroemer G and Tang D: Ferroptosis is a type of autophagy-dependent
cell death. Semin Cancer Biol. 66:89–100. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Gao AC, Lou W, Dong JT, Barrett JC,
Danielpour D and Isaacs JT: Defining regulatory elements in the
human KAI1 (CD 82) metastasis suppressor gene. Prostate.
57:256–260. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Xie Y, Hou W, Song X, Yu Y, Huang J, Sun
X, Kang R and Tang D: Ferroptosis: Process and function. Cell Death
Differ. 23:369–379. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lu B, Chen XB, Ying MD, He QJ, Cao J and
Yang B: The role of ferroptosis in cancer development and treatment
response. Front Pharmacol. 8:9922018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Dixon SJ and Stockwell BR: The role of
iron and reactive oxygen species in cell death. Nat Chem Biol.
10:9–17. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Li J, Cao F, Yin HL, Huang ZJ, Lin ZT, Mao
N, Sun B and Wang G: Ferroptosis: Past, present and future. Cell
Death Dis. 11:882020. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Mou Y, Wang J, Wu J, He D, Zhang C, Duan C
and Li B: Ferroptosis, a new form of cell death: Opportunities and
challenges in cancer. J Hematol Oncol. 12:34–50. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Camaschella C, Nai A and Silvestri L: Iron
metabolism and iron disorders revisited in the hepcidin era.
Haematologica. 105:260–272. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Manz DH, Blanchette NL, Paul BT, Torti FM
and Torti SV: Iron and cancer: Recent insights. Ann N Y Acad Sci.
1368:149–161. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Petronek MS, Spitz DR, Buettner GR and
Allen BG: Linking cancer metabolic dysfunction and genetic
instability through the lens of iron metabolism. Cancers (Basel).
11:10772019. View Article : Google Scholar
|
|
37
|
Kazan HH, Urfali-Mamatoglu C and Gunduz U:
Iron metabolism and drug resistance in cancer. Biometals.
30:629–641. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Thévenod F: Iron and its role in cancer
defense: A double-edged sword. Met Ions Life Sci. 18:2018.
|
|
39
|
Chen Y, Fan Z, Yang Y and Gu C: Iron
metabolism and its contribution to cancer (Review). Int J Oncol.
54:1143–1154. 2019.PubMed/NCBI
|
|
40
|
Wang YF, Yu L, Ding J and Chen Y: Iron
metabolism in cancer. Int J Mol Sci. 20:952019. View Article : Google Scholar
|
|
41
|
Forcina GC and Dixon SJ: GPX4 at the
crossroads of lipid homeostasis and ferroptosis. Proteomics.
19:e18003112019. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Seibt TM, Proneth B and Conrad M: Role of
GPX4 in ferroptosis and its pharmacological implication. Free Radic
Biol Med. 133:144–152. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Yang WS, SriRamaratnam R, Welsch ME,
Shimada K, Skouta R, Viswanathan VS, Cheah JH, Clemons PA, Shamji
AF, Clish CB, et al: Regulation of ferroptotic cancer cell death by
GPX4. Cell. 156:317–331. 2014. View Article : Google Scholar : PubMed/NCBI
|