Introduction
Diabetic nephropathy (DN) is a type of severe
complication of diabetes mellitus (DM) and is the main cause of
chronic kidney disease (1). Various
of mechanism are reported to be involved in the pathogenesis and
development of DN, such as hyperglycemia, oxidative stress
(2), mitochondrial dysfunction
(3), endoplasmic reticulum stress,
chronic inflammation and abnormal autophagy (4). Recently, accumulating evidence has
revealed that lipid metabolism abnormality serves an important role
in the pathogenesis of DN (5). In
both diabetic animals and patients, renal mesangial cells, renal
tubular cells and podocytes experience lipid accumulation, which
induces extracellular matrix (ECM) deposition, including
fibronectin, collagen I, collagen 3 mainly located in renal tubular
interstitium and collagen 4 mainly located in renal glomeruli, and
fibrosis (6,7).
Sterol regulatory element-binding protein 1
(SREBP-1) is a key transcription factor of lipid metabolism,
especially fatty acid synthesis (8). In a previous study, SREBP-1 was proven
to regulate the lipid metabolism abnormality of DN in renal tubular
cells (9). Hyperglycemia increases
SREBP-1 expression, leading to fatty acid synthase, acetol-CoA
carboxylase upregulation and triglyceride augment in renal tubular
cells (9). However, the exact
mechanism involved in hyperglycemia-induced increased SREBP-1 is
yet to be fully elucidated.
F-box and WD repeat domain containing 7 (FBXW7) is a
type of E3 ubiquitin ligase that belongs to the SCF E3 ubiquitin
ligase family, and has multiple downstream targets, such as c-Myc,
MCL1 apoptosis regulator, BCL2 family member, c-Jun, Notch1 and
cyclin E (10). Moreover, FBXW7 is
a multifunctional protein and regulates cell proliferation,
apoptosis, survival and aging (11). FBXW7 also serves a role in affecting
tumor growth in neoplasm genesis and progression, acting as tumor
suppressor gene (12). Increasing
evidence has revealed that SREBP-1 is another downstream target of
FBXW7 (13). Therefore, we
hypothesized that FBXW7 may mediate high glucose-induced SREBP-1
upregulation in renal tubular cells.
In the present study, diabetic mice and a human
renal tubular cell line (HKC) were chosen to detect the expression
levels of FBXW7 and SREBP-1. Furthermore, FBXW7 expression was
upregulated or downregulated using expression plasmid or short
hairpin (sh)RNA plasmid to investigate the direct relationship
between FBXW7 and SREBP-1. Considering that the PI3K/Akt pathway is
the critical cell signaling pathway affecting DN (14), the potential role of the PI3K/Akt
pathway on FBXW7 expression in HKC cells was determined.
Materials and methods
Reagents and materials
The antibody against SREBP-1 (cat. no. sc-13551,
dilution 1:200 for immunohistochemistry, immunocytochemistry and
immunofluorescence; dilution 1:1,000 for western blotting) was
purchased from Santa Cruz Biotechnology, Inc. The antibody against
FBXW7 was obtained from Aviva Systems Biology Corp (cat. no.
ARP47419_P050, dilution 1:200 for immunohistochemistry,
immunocytochemistry and immunofluorescence, 1:1,000 for western
blotting). The antibodies against phosphorylated (p)-Akt (Ser 473;
cat. no. 4060, dilution 1:1,000 for western blotting) and Akt (cat.
no. 4691, dilution 1:1,000 for western blotting) were purchased
from Cell Signaling Technology, Inc. The antibody against nestin
was bought from Abcam (cat. no. ab221660, dilution 1:200 for
immunofluorescence). The antibody against β-actin was purchased
from ABclonal Biotechnology Co., Ltd. (cat. no. AC026, dilution
1:1,000 for western blotting). Streptozotocin (STZ) was obtained
from Sigma-Aldrich (Merck KGaA), while LY294002 was purchased from
MedChemExpress. Lipofectamine® 2000 was purchased from
Thermo Fisher Scientific, Inc. and TRIzol® from
Invitrogen (Thermo Fisher Scientific, Inc.). The PrimeScript™
reverse transcription (RT) reagent kit with gDNA Eraser and
SYBR® Premix Ex Taq™ II (Tli RNaseH Plus) were obtained
from Takara Bio, Inc. An immunohistochemistry kit was purchased
from Beijing Zhongshan Golden Bridge Technology Co., Ltd. DyLight
488 or 594-labelled goat secondary antibodies were purchased from
KPL, Inc. The FBXW7 expression plasmid (pcDNA3.0-FBXW7) was bought
from Sino Biological. The pGenesil-1 plasmid (Wuhan An Di Jing Sai
Bio-Technology Co., Ltd.) for shRNA plasmid construction was stored
in the authors' lab. Glucose and mannitol were bought from Beijing
Solarbio Science & Technology Co., Ltd.
Diabetic mice
A total of 20 male CD1 mice aged six weeks (weight
20–25 g) were purchased from Beijing Vital River Laboratory Animal
Technology Co., Ltd. and were fed with free access to water and
food. All animal experiments were according to the rules of
Institutional Animal Care and Use Committee of the Third Hospital
of Hebei Medical University (permit no. Guo A 2017-019-1; period,
2017–2020). All animals were housed with 12-h light/dark cycle,
20±3°C and 35–55% humidity. A total of 10 mice were
intraperitoneally injected with STZ (150 mg/kg body weight) to
establish diabetic models (15) and
the corresponding 10 control mice were injected with equivalent
volume sodium citrate solution (1.29% sodium citrate, 1.18% citric
acid). Then, 3 day after the injection, mice with measured blood
glucose levels of >16.7 mmol/l were regarded as successfully
established models. All animals were fed for 16 weeks and then
sacrificed. Mice were euthanized by exsanguination after deep 4%
isoflurane anesthesia, and death was confirmed by the lack of
reflexes of paw withdrawal and eye blink. Mice with a lack of
breathing and heartbeat were considered dead, and the renal cortex
was collected for the subsequent experiments.
Cell culture and grouping
A human renal tubular cell line (HKC) was kindly
provided by Professor Chen Xiang-Mei, Division of Nephropathy, 301
Hospital (Beijing, China). The HKC cell line was originally
established by Racusen et al (16) who isolated human renal tubule
epithelial cells and exposed them in culture to a hybrid
immortalizing virus, adenovirus 12-SV40. The cells were cultured as
described previously (9) and
randomly divided into three groups to examine the effect of high
glucose: Normal glucose group (5.5 mmol/l glucose; N), high glucose
group (30 mmol/l glucose; H) and mannitol group (30 mmol/l
mannitol; M). After the indicated times (at 37°C for 36, 48 and 72
h), the various assays were performed.
To investigate the effect of FBXW7 overexpression on
SREBP-1 expression, HKC cells were divided into untransfected
group, pcDNA3.0 group and pcDNA3.0-FBXW7 group. Moreover, to
determine the effect of FBXW7 knockdown on SREBP-1 expression, HKC
cells were divided into untransfected group, pGenesil-1 group,
pGenesil-1-FBXW7-1 group and pGenesil-1-FBXW7-2 group. To evaluate
the role of the PI3K/Akt pathway in regulating FBXW7 expression,
HKC cells were divided into N group, H group, H + DMSO group
(1:1,000 DMSO) and H + LY294002 group (20 µmol/l LY294002). After
48 h at 37°C, the related detections were performed.
shRNA plasmid construction
The pGenesil-1 plasmid was used to construct FBXW7
shRNA recombinant plasmid. A total of two single-stranded DNA
oligonucleotides aimed at the FBXW7 gene were annealed to form
double-stranded DNA, which was linked with pGenesil-1 digested with
BamHI and HindIII using a T4 ligase at 16°C
overnight. Then, the mixture was transfected into E. coil
strain DH5 α for cloning and plasmid extraction. Finally, two
constructs were designated pGenesil-1-FBXW7-1 and
pGenesil-1-FBXW7-2 after sequencing identification. The
pGenesil-1-FBXW7-1 targeted the sequence 5′-ACAGGACAGTGTTTACAAA-3′,
and pGenesil-1-FBXW7-2 targeted the sequence
5′-CAACAACGACGCCGAATTA-3′.
Plasmid transfection
Lipofectamine 2000 was used in HKC cells to perform
transient transfection. HKC cells seeded on cover slides in 6-well
plate were treated at 37°C for 5 h with 250 µl serum-free DMEM
(Gibco; Thermo Fisher Scientific, Inc.) containing 3.0 µg plasmid
and 10 µl Lipofectamine 2000. After 5 h, this medium was replaced
with normal DMEM containing 10% FBS (Biological Industries). At 48
h after transfection, the cells were collected for the subsequent
experimentations.
Immunohistochemistry and
immunocytochemistry
Renal cortex tissues were resected from the kidneys
and fixed immediately in 4% paraformaldehyde for 48 h at room
temperature. Subsequently, 4 µm paraffin-embedded sections were
made for immunohistochemistry. After deparaffinization (100%
xylene, room temperature, 30 min), hydration (gradient ethanol
series, room temperature, 10 min each), antigen recovery (citrate
buffer pH 6.0, 100°C, 5 min) and blocking (5% goat serum; cat. no.
SP9000; OriGene Technologies, Inc.), 37°C, 20 min), sections were
incubated with primary antibodies against FBXW7 or SREBP-1 at 4°C
overnight. The next morning sections were washed with PBS and then
incubated with ready-to-use biotin-labelled secondary antibody
(cat. no. SP9000; OriGene Technologies, Inc.) for 30 min at 37°C.
After incubation with horseradish peroxidase (HRP)-labelled
streptavidin for 30 min at 37°C, sections was visualized using DAB
(OriGene Technologies, Inc.) for 10 sec at room temperature. A
negative control was established by replacing specific antibody
with PBS. The sections were imaged with Olympus light microscope
(Olympus Corporation) at ×400 magnification.
Immunocytochemistry was used to investigate the
expression levels of FBXW7 and SREBP-1 in HKC cells. After fixation
(4% paraformaldehyde, room temperature, 15 min), permeabilization
(0.3% Triton X-100, 37°C, 10 min) and blocking (5% goat serum,
37°C, 30 min), cells were incubated with specific primary antibody
of FBXW7 overnight at 4°C. The following day, cells were incubated
with biotin-labelled secondary antibody at 37°C for 30 min and
HRP-labelled streptavidin at 37°C for 30 min, in turn. Finally,
cells were stained with DAB at room temperature for 20 sec,
followed by image capture using light microscope at ×400
magnification.
Immunofluorescence
After fixation using 4% paraformaldehyde at room
temperature for 48 h, 4 µm sections were made according to the
standard procedure. After deparaffinization (100% xylene, room
temperature, 30 min), hydration (gradient ethanol series, room
temperature, 10 min each) and blocking (5% goat serum, 37°C, 20
min), tissue sections were co-incubated with primary antibodies of
FBXW7 and SREBP-1 at 4°C overnight. Then, after rinsing with PBS,
sections were co-incubated with DyLight 488 labelled-goat
anti-rabbit secondary antibody and DyLight 594 labelled-goat
anti-mouse secondary antibody at 37°C for 30 min. After
counter-staining with DAPI at room temperature for 10 min, positive
signals were observed under fluorescence microscope and images were
captured at ×400 magnification.
For cultured cells, after fixation, permeabilization
and blocking, cells were incubated with primary antibody against
FBXW7 overnight at 4°C and DyLight 488-labelled secondary antibody
at 37°C for 2 h, in turn. Positive signals were observed under
fluorescence microscope at ×400 magnification following DAPI
staining at room temperature for 10 min.
RT-quantitative (RT-q)PCR
Total RNA was extracted from HKC cells using TRIzol
reagent and reverse transcribed using RT primer mix and RT enzyme
mix (PrimeScript™ reverse transcription reagent kit with gDNA
Eraser, Takara Bio, Inc.) at 37°C for 15 min after quantification.
Equal amounts of RT reaction product (2 µl) were subjected to PCR
amplification and the SYBR-Green (Takara Bio, Inc.) qPCR method was
used to determine FBXW7 mRNA expression. The thermocycling
conditions were as follows: Initial denaturation for 3 min at 95°C,
followed by 40 cycles of denaturation at 95°C for 5 sec, annealing
at 55°C for 30 sec and extension at 72°C for 30 sec. The final
extension was performed at 72°C for 10 min. GAPDH was used as a
housekeeping gene. The results were analyzed using the
2−∆∆Cq method (17). The
primers sequences are presented in Table I.
 | Table I.Primers of human SREBP-1 and
GAPDH. |
Table I.
Primers of human SREBP-1 and
GAPDH.
| Gene | Forward primer | Reverse primer |
|---|
| Human SREBP-1 |
5′-AAAGAGTTGTTAGCGGTTCTCG-3′ |
5′-CCACATGGATACCATCAAACTG-3′ |
| Human GAPDH |
5′-GTCAACGGATTTGGTCGTATTG-3′ |
5′-TGTAGTTGAGGTCAATGAAGGG-3′ |
Western blotting
Total protein extracted from HKC cells (RIPA lysis
buffer; Beijing Solarbio Science & Technology Co., Ltd.) was
quantified using a Bio-Rad Quick Start Bradford Protein Assay kit
(Bio-Rad Laboratories, Inc.). An equal amount of protein (30 µg)
was separated via 10% SDS-PAGE. Then, protein was transferred onto
PVDF membranes (EMD Millipore), which were blocked with 5% BSA
(Biosharp Life Sciences) for 1 h at 37°C. Next, blots were
incubated with primary antibodies for FBXW7, SREBP-1 and β-actin
overnight at 4°C. The following morning, blots were washed with
TBS-0.1% Tween-20 (TBST) and incubated with HRP-conjugated
secondary antibody for 2 h at room temperature. After rinsing with
TBST, blots were incubated with ECL solution (Tiangen Biotech Co.,
Ltd.) for 2 min at room temperature and bands were visualized. For
semi-quantitative analysis, bands were evaluated with LabWorks v4.5
software (Analytik Jena UVP) and normalized to β-actin density. The
data are representative of three independent experiments.
Statistical analysis
Data are presented as the mean ± SD and were
analyzed with SPSS 13.0 for Windows (SPSS, Inc.). One-way ANOVA
followed by Bonferroni post hoc test was used to determine
significant differences among >2 groups. Unpaired Student's
t-test was performed to determine the significance between two
groups. P<0.05 was considered to indicate a statistically
significant difference. All experiments were independently repeated
three times.
Results
FBXW7 and SREBP-1 expression levels in
renal tubular cells of diabetic mice
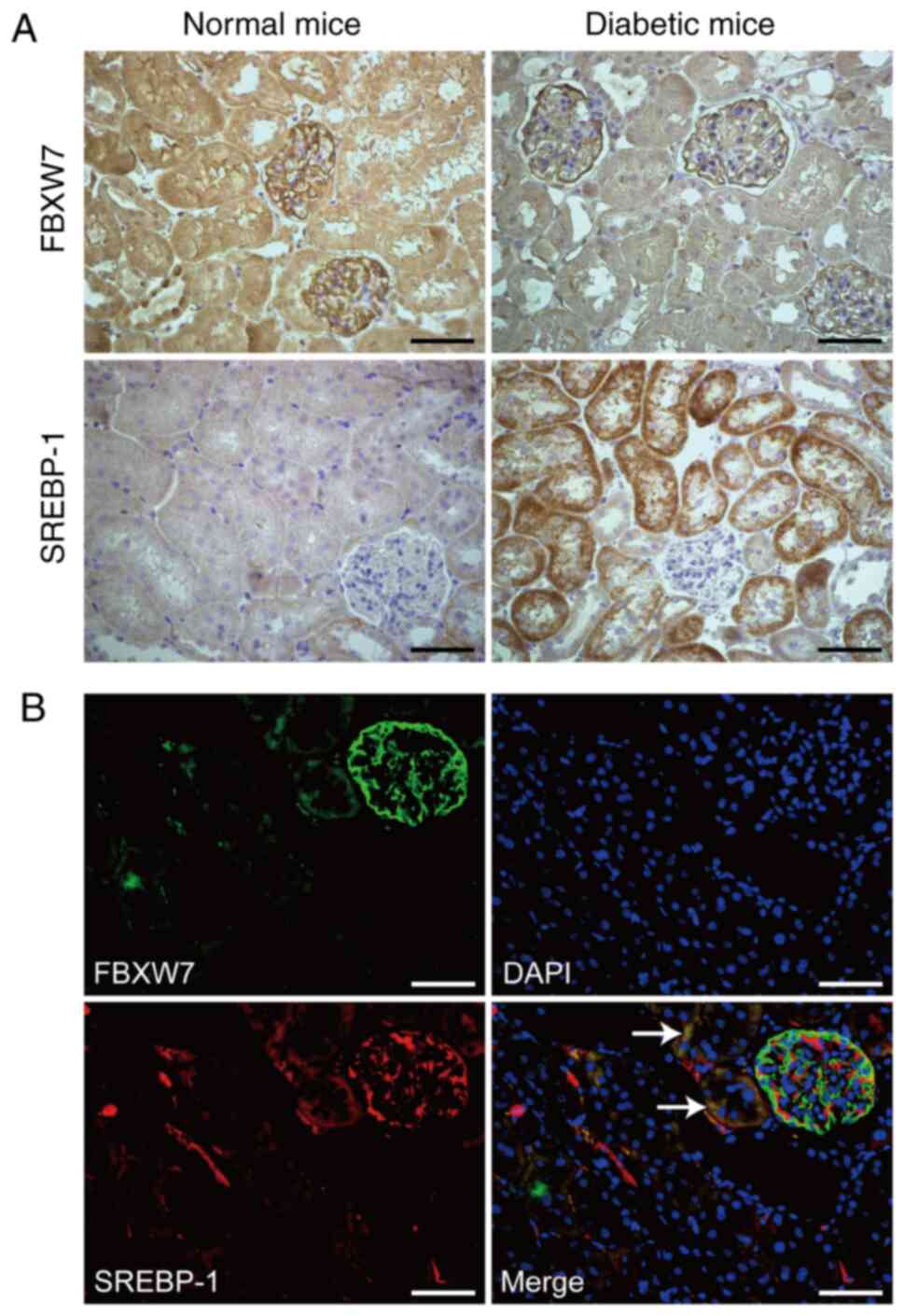
FBXW7 and SREBP-1 expression levels were detected
via immunohistochemistry in normal mice and diabetic mice. As
presented in Fig. 1A, FBXW7 protein
expression was located in the glomeruli and renal tubular cells of
normal mice and diabetic mice. Compared with normal mice, diabetic
mice had a lower expression of FBXW7 in renal tubular cells. On the
contrary, SREBP-1 expression was increased in renal tubular cells
of diabetic mice in comparison with normal mice. Furthermore, the
potential co-expression of FBXW7 and SREBP-1 was analyzed using a
double staining immunofluorescence technique, and it was identified
that there was a definite co-expression of FBXW7 and SREBP-1 in
renal tubular cells. Different to the immunohistochemistry
findings, the results of immunofluorescence also identified the
expression of SREBP-1 in the glomeruli, which may be due to the
high sensitivity of immunofluorescence. However, there was no
co-expression of FBXW7 and SREBP-1 in glomeruli, which suggested
the diverse location of FBXW7 and SREBP-1 in renal glomeruli
(Fig. 1B). The exact location of
FBXW7 in glomeruli was further examine using double staining
immunofluorescence with antibodies against FBXW7 and nestin
(biomarker of podocyte). The partial co-expression of FBXW7 and
nestin suggested that FBXW7 was also located in podocytes, besides
renal tubular cells (Fig. S1).
High glucose decreases FBXW7
expression and increases SREBP-1 expression in HKC cells
Hyperglycemia is the most important characteristic
of DM (18), and high glucose was
used to treat HKC cells in the present study. It was identified
that FBXW7 was decreased after treatment with high glucose for 36,
48 and 72 h compared with the corresponding N group (Fig. 2A). Statistical analysis demonstrated
that FBXW7 expression was decreased by 70% in HKC cells treated
with high glucose for 72 h compared with those cells treated with
normal glucose. Moreover, there was no significant difference in
the expression of FBXW7 between N group and M group at the
indicated time points. Similarly, the results of immunofluorescence
demonstrated that FBXW7 was located in the cytoplasm of HKC cells
and was downregulated after treatment with high glucose in
comparison with normal glucose treatment (Fig. 2B).
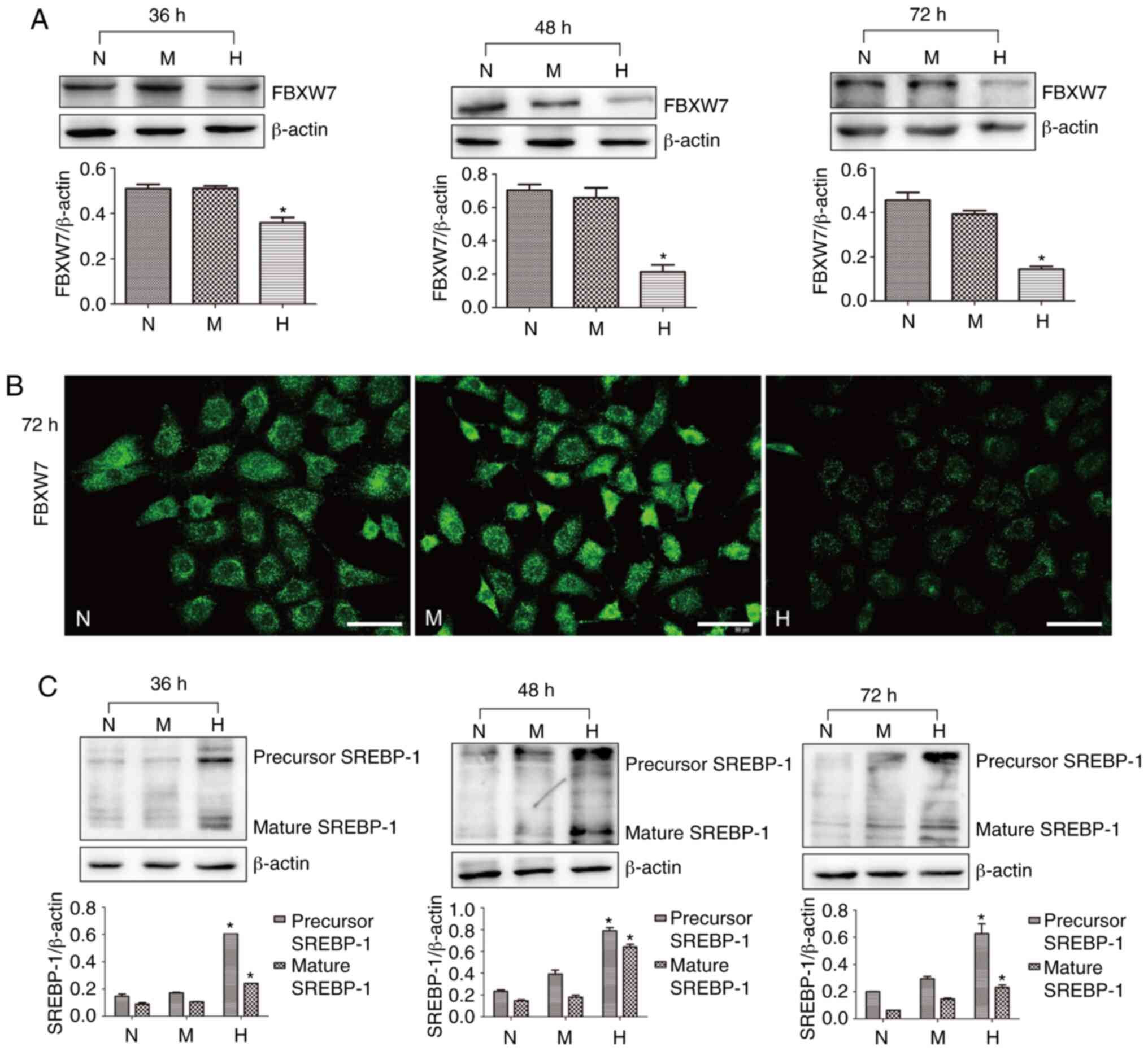 | Figure 2.Expression levels of FBXW7 and
SREBP-1 in H-cultured HKC cells. (A) Western blotting and
statistical analysis of FBXW7 expression in HKC cells treated with
N, H and M for 36, 48 and 72 h. (B) Immunofluorescence of FBXW7
expression in HKC cells treated with N, H and M for 48 h. Scale
bar, 50 µm. (C) Western blotting and statistical analysis of
precursor segment of SREBP-1 and mature segment of SREBP-1 in HKC
cells treated with N, H and M for 36, 48 and 72 h. *P<0.05 vs. N
group. N, normal glucose; M, mannitol; FBXW7, F-box and WD repeat
domain containing 7; SREBP-1, sterol regulatory element-binding
protein 1. |
SREBP-1 expression was detected via western
blotting, and the immunoblots were representative of three
independent experiments. The precursor segment of SREBP-1 was
increased by 3.75, 4.16 and 3.15 fold after treatment with high
glucose for 36, 48 and 72 h, respectively, compared with the N
group. Furthermore, the mature segment of SREBP-1 was enhanced by
2.67, 4.27 and 3.83 fold after treatment with high glucose
(Fig. 2C).
Overexpression of FBXW7 decreases
SREBP-1 expression in HKC cells
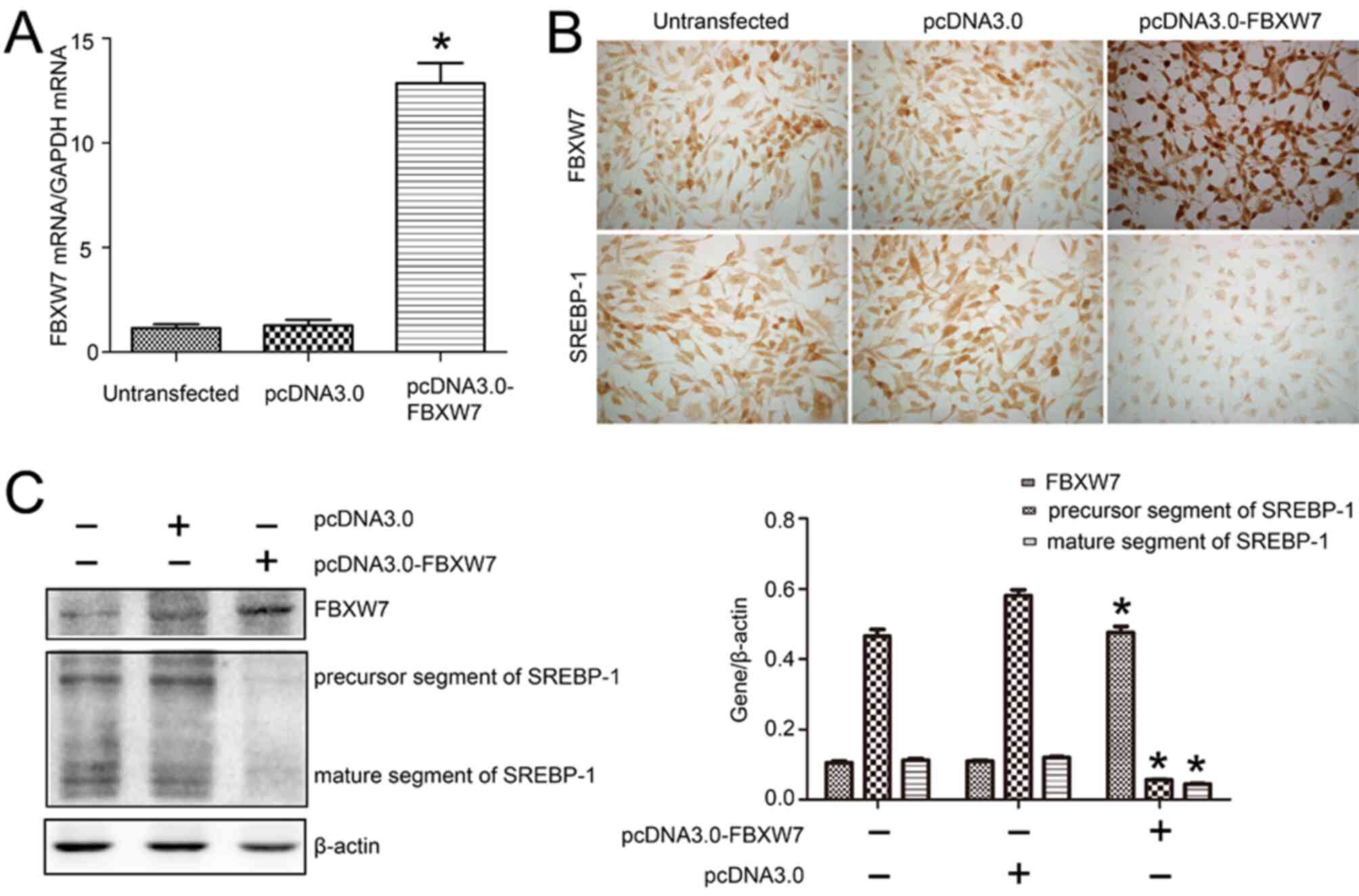
To elucidate the direct effect of FBXW7 on SREBP-1,
FBXW7 expression was overexpression via transfection with
pcDNA3.0-FBXW7 plasmid in HKC cells. The RT-qPCR results identified
a 10.12-fold increase in FBXW7 mRNA expression (Fig. 3A). Furthermore, the
immunocytochemistry demonstrated that the overexpression of FBXW7
appeared as increased brown staining in HKC cells transfected with
pcDNA3.0-FBXW7, compared with that observed in cells transfected
with pcDNA3.0. It was found that SREBP-1 expression was
downregulated, as detected via immunocytochemistry (Fig. 3B). Overexpression of FBXW7 also led
to smaller-sized HKC cells compared with those found in the
pcDNA3.0 blank plasmid transfection group.
Subsequently, western blotting was conducted and the
results indicated that pcDNA3.0-FBXW7 plasmid transfection
increased FBXW7 expression by 1.29-fold compared with the pcDNA3.0
transfection. Moreover, the precursor segment and mature segment of
SREBP-1 were decreased by 11.11 and 2.63 fold, respectively, in HKC
cells transfected with pcDNA3.0-FBXW7 plasmid compared with those
transfected with pcDNA3.0 (Fig.
3C).
Downregulation of FBXW7 expression
increases SREBP-1 expression in HKC cells
The effect of FBXW7 on SREBP-1 expression in HKC
cells was further examined by knocking down FBXW7 expression via
transfection with shRNA plasmid. As presented in Fig. 4A, the transfection efficiency of
pGenesil-1, pGenesil-1-FBXW7-1 and pGenesil-1-FBXW7-2 was >75%,
as indicated by GFP expression. RT-qPCR results demonstrated that
FBXW7 mRNA expression was effectively decreased in both
pGenesil-1-FBXW7-1-transfected and pGenesil-1-FBXW7-2-transfected
HKC cells compared with pGenesil-1-transfected cells. Statistical
analysis identified a 54% and a 56% downregulation in FBXW7 mRNA
expression in pGenesil-1-FBXW7-1-transfected cells and
pGenesil-1-FBXW7-2-transfected cells, respectively (Fig. 4B). In line with RT-qPCR results,
immunocytochemistry analysis found that pGenesil-1-FBXW7-1 and
pGenesil-1-FBXW7-2 downregulated FBXW7 protein expression (Fig. 4C). Conversely, SREBP-1 expression
was markedly enhanced with the transfection of pGenesil-1-FBXW7-1
and pGenesil-1-FBXW7-2 in HKC cells (Fig. 4D). In addition, the western blotting
results found a 29% and a 41% downregulation in FBXW7 expression
after the transfection of pGenesil-1-FBXW7-1 and pGenesil-1-FBXW7-2
in cells, compared with that in pGenesil-1 in HKC cells. A 2.84 and
a 2.54 fold-increase in precursor SREBP-1 and mature SREBP-1
expression levels was identified after pGenesil-1-FBXW7-1
transfection, as well as a 3.52 and a 3.64 fold-increase after
pGenesil-1-FBXW7-2 transfection (Fig.
4E).
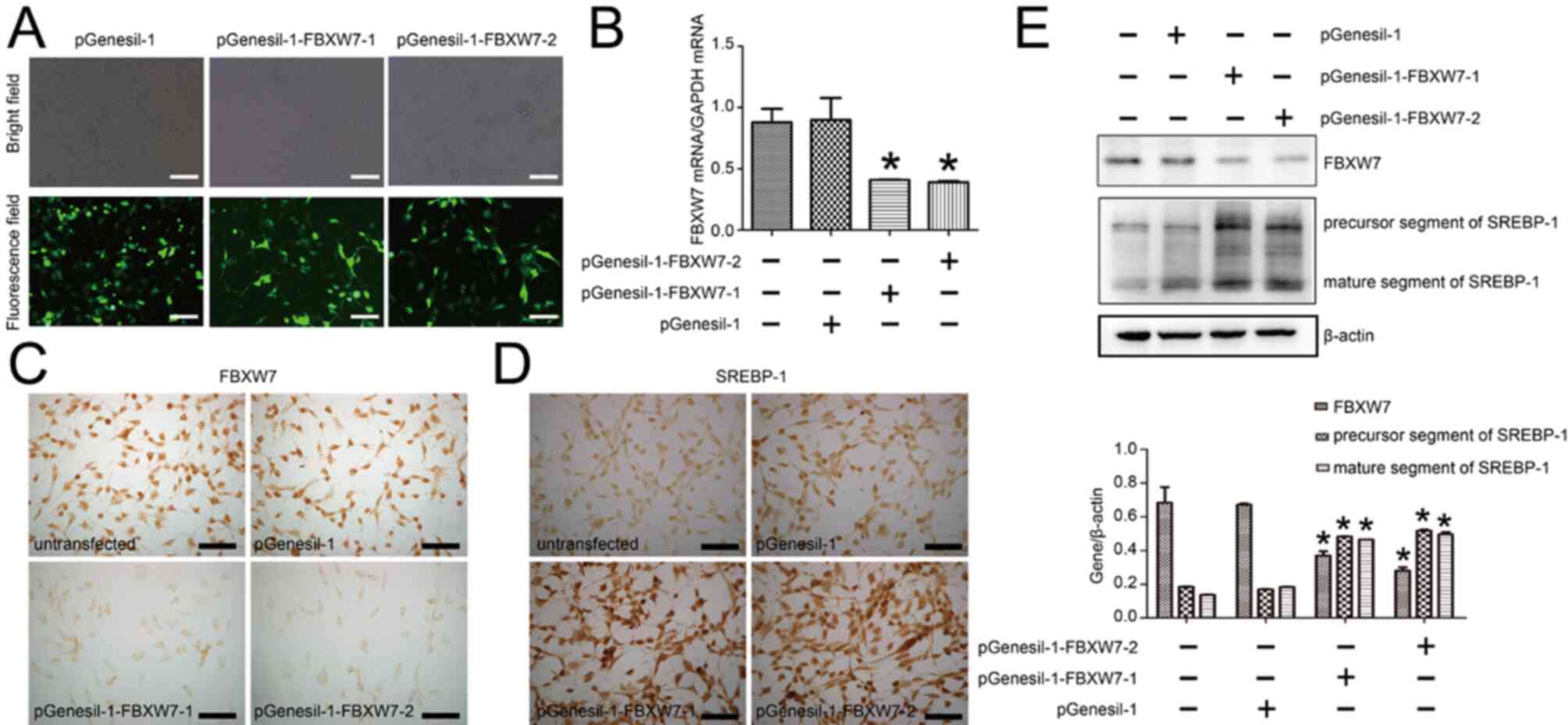 | Figure 4.Knockdown of FBXW7 increases SREBP-1
expression in HKC cells. (A) Transfection efficiency of HKC cells
transfected with pGenesil-1, pGenesil-1-FBXW7-1 and
pGenesil-1-FBXW7-2. Scale bar, 100 µm. (B) Reverse
transcription-quantitative PCR results of FBXW7 mRNA expression in
HKC cells of untransfected group, pGenesil-1 group,
pGenesil-1-FBXW7-1 group and pGenesil-1-FBXW7-2 group. (C)
Immunocytochemistry results of FBXW7 expression in HKC cells of
untransfected group, pGenesil-1 group, pGenesil-1-FBXW7-1 group and
pGenesil-1-FBXW7-2 group. Scale bar, 50 µm. (D) Immunocytochemistry
results of SREBP-1 expression in HKC cells of untransfected group,
pGenesil-1 group, pGenesil-1-FBXW7-1 group and pGenesil-1-FBXW7-2
group. Scale bar, 50 µm. (E) Western blotting and statistical
analysis of FBXW7, precursor segment of SREBP-1 and mature segment
of SREBP-1 expression levels in HKC cells of untransfected group,
pGenesil-1 group, pGenesil-1-FBXW7-1 group and pGenesil-1-FBXW7-2
group. *P<0.05 vs. pGenesil-1 group. FBXW7, F-box and WD repeat
domain containing 7; SREBP-1, sterol regulatory element-binding
protein 1. |
Inhibition of the PI3K/Akt pathway
enhances FBXW7 expression and decreases SREBP-1 expression in HKC
cells
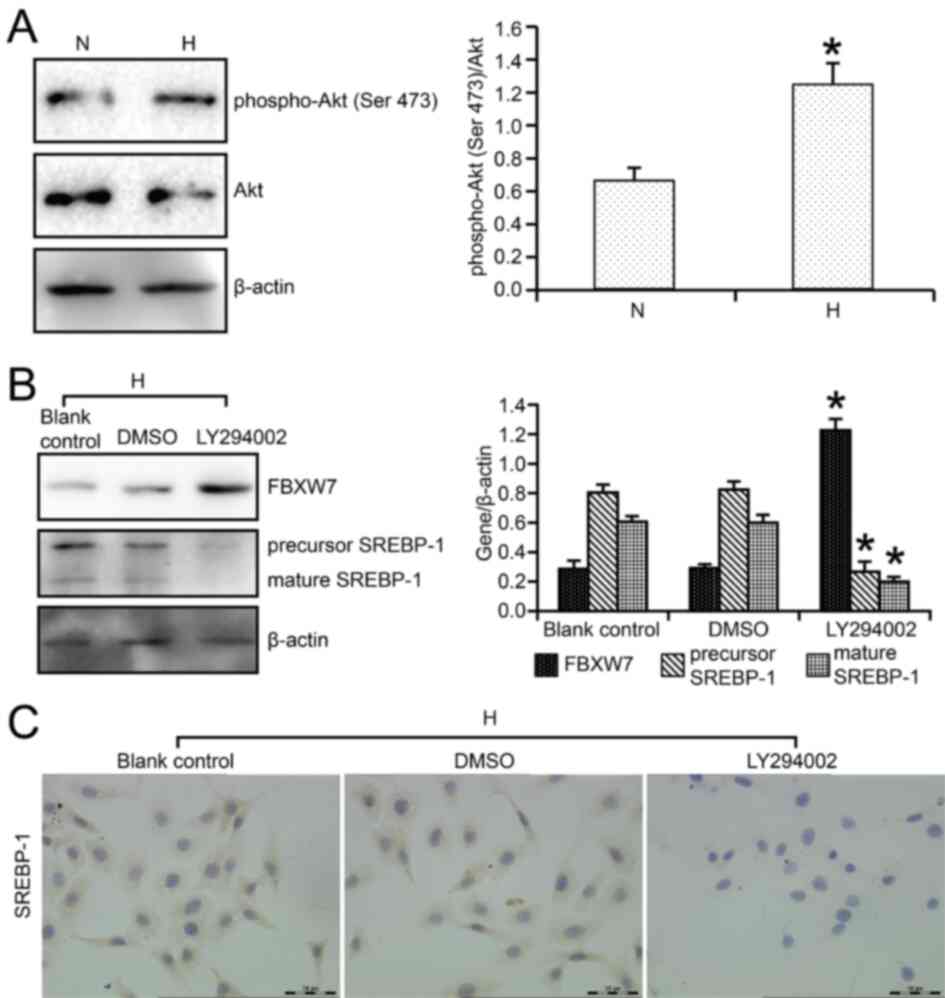
PI3K/Akt signaling has been revealed to be the main
cell signaling pathway that regulates cell function in renal
tubular cells of DN, such as lipid metabolism,
epithelial-mesenchymal transition and autophagy (14,19).
The results indicated that in high glucose-cultured HKC cells,
p-Akt (Ser 473)/Akt ratio was significantly increased compared with
that in normal glucose-cultured HKC cells (Fig. 5A). Moreover, LY294002, a known
PI3K/Akt pathway inhibitor, was used to determine the effect of the
PI3K/Akt pathway on FBXW7 and SREBP-1 expression levels in HKC
cells. It was found that FBXW7 expression was upregulated by 4.21
fold after LY294002 treatment compared with DMSO treatment
(Fig. 5B). Furthermore, the
precursor segment and mature segment of SREBP-1 were downregulated
by 3.10 and 3.00 fold, respectively, by LY294002 treatment,
compared with DMSO (Fig. 5B).
Immunocytochemistry also demonstrated that LY294002 markedly
decreased SREBP-1 expression, which presented as brown granules, in
high glucose-cultured HKC cells (Fig.
5C).
Discussion
FBXW7 has been reported to be associated with the
pathogenesis of DM. Brenachot et al (20) revealed that IL-1β and IFN-γ
downregulated FBXW7 expression in vitro insulin cells,
followed by NF-κB pathway activation and cell apoptosis, which led
to type I DM. Moreover, in type II DM, Zhao et al (21) observed that loss of FBXW7 in hepatic
cells resulted in hyperglycemia, insulin resistance and impaired
glucose tolerance. The present study identified the decreased
expression of FBXW7 in renal tubular cells of DN. Similarly, Tu
et al (22) identified FBXW7
downregulation in the later stages of diabetic cardiomyopathy
(>12 weeks). Therefore, these findings suggest that FBXW7
expression abnormality serves a role in the pathogenesis of DM and
the corresponding complications.
The present study examined the potential regulatory
factor of FBXW7 expression in renal tubular cells of DN using in
vitro cultured cells. The results demonstrated that
hyperglycemia was the main factor to inhibit FBXW7 expression in
HKC cells. In line with the current findings, Gao et al
(23) also reported that
hyperglycemia was a regulatory factor of FBXW7 and could
downregulate FBXW7 expression in renal mesangial cells, leading to
diminished autophagy and increased inflammatory cytokines.
Therefore, it was suggested that high glucose was the main factor
to decrease FBXW7 and increase SREBP-1 expression levels in renal
tubular cells of DN. Using double staining immunofluorescence in
renal tissue it was also found that FBXW7 was located in podocytes,
which indicated that, alongside renal tubular cells, the function
of FBXW7 in podocytes requires further study.
The present study identified a negative relationship
between FBXW7 and SREBP-1 expression in renal tubular cells of
diabetic mice and in vitro HKC cells. This type of negative
association between FBXW7 and SREBP-1 was also revealed in a study
by Tu et al (24). These
authors reported that in C57BL/6J mice fed with high fat diets the
mRNA and protein expression levels of Fbxw7 were significantly
decreased in liver tissues, compared with normal diets group, which
was negatively correlated with SREBP-1, indicating the role of the
FBXW7/SREBP-1 axis in the development of non-alcoholic fatty liver
disease (24). Moreover, the
present study observed the co-location of FBXW7 and SREBP-1 in
renal tubular cells of diabetic mice, but not in glomeruli, using
immunofluorescence double staining. In line with this finding, our
previous study revealed that SREBP-1 was mainly expressed in renal
tubular cells, which could mediate lipid metabolism abnormality and
ECM accumulation in DN (9,25). However, immunofluorescence detection
demonstrated the positive expression of SREBP-1 in glomeruli, while
immunohistochemistry did not detect a positive signal of SREBP-1 in
glomeruli. It was suggested that the sensitivity difference of the
experimental methods may be the main cause of this discrepancy.
Therefore, considering the negative relationship and co-expression
of FBXW7 and SREBP-1, it was suggested that FBXW7 may negatively
regulate SREBP-1 expression in renal tubular cells of DN.
To elucidate the direct regulation of FBXW7 on
SREBP-1 in HKC cells, the present study overexpressed FBXW7, and
subsequently found that SREBP-1 was downregulated. However, when
FBXW7 was knocked down by RNA interference, SREBP-1 was
upregulated. As an E3 ubiquitin ligase, FBXW7 has been reported to
enhance SREBP-1a, SREBP-1c and SREBP-2 ubiquitination and
degradation in a manner dependent on the phosphorylation of SREBPs,
which is regulated by GSK-3 (26).
Moreover inactivation of FBXW7 results in the stabilization of
SREBPs, enhancing the synthesis of cholesterol and fatty acids via
SREBPs downstream genes (26,27).
However, in a previous study, FBXW7 was also revealed to be
regulated by SREBP-2 via microRNAs (miR), miR-182 and miR-96, and
accumulating miR-182 and miR-96 negatively regulated FBXW7 and
insulin induced gene-2 expression levels (28). Thus, there may be a regulatory loop
between FBXW7 and SREBPs. Therefore, along with the present data,
it was suggested that FBXW7 may be the upstream regulator of
SREBP-1 in renal tubular cells of DN. Furthermore, the present
results indicated that overexpression of FBXW7 caused smaller-sized
HKC cells. Based on a previous study (29), it was suggested that FBXW7
upregulation may affect cell proliferation via its target c-Myc, as
well as lipid metabolism via SREBP-1.
The PI3K/Akt pathway is a multifunctional signaling
pathway and serves an important role in the pathogenesis and
development of DN (30). In our
previous study, the PI3K/Akt pathway was revealed to regulate
SREBP-1 expression in renal tubular cells of DN (14). Furthermore, downstream proteins of
Akt, GSK-3β and mTOR were found to be involved in PI3K/Akt
pathway-regulated SREBP-1 expression and lipid metabolism in renal
tubular cells of DN (31,32). The present study demonstrated that
PI3K/Akt pathway activation also regulated FBXW7 expression in HKC
cells. LY294002, a known PI3K/Akt pathway inhibitor, improved the
high glucose-induced decline in FBXW7 expression. Similarly, Suryo
Rahmanto et al (33)
reported that in medulloblastoma cells, pharmacological inhibition
of the PI3K/Akt pathway enhanced FBXW7 expression and destabilized
SOX9, rendering cells sensitive to cisplatin treatment. Therefore,
the PI3K/Akt signaling pathway may be the upstream pathway that
regulates FBXW7 expression in renal tubular cells of DN. However,
to further elucidate the function and regulation of FBXW7 in DM,
additional in vivo experiments using lentivirus containing
FBXW7 CDS (coding sequence) or shRNA targeted at FBXW7 are
required.
In conclusion, the present findings demonstrated
that high glucose caused FBXW7 downregulation in renal tubular
cells of diabetic mice, especially at late stage, and was
negatively associated with SREBP-1 expression. Moreover, FBXW7 may
be the upstream regulator of SREBP-1 in renal tubular cells of DN,
and it was found that the PI3K/Akt pathway mediated the high
glucose-induced downregulation of FBXW7 expression in renal tubular
cells. Therefore, combined therapies targeted at both FBXW7 and
SREBP-1 may be an effective strategy to prevent DN by regulating
lipid metabolism.
Supplementary Material
Supporting Data
Acknowledgements
The authors would like to thank Professor Huachuan
Zheng (Department of Experimental Oncology and Animal Center,
Shengjing Hospital of China Medical University) for helpful
guidance in biotechnology.
Funding
This work was supported by a grant from the Natural
Science Foundation of Hebei Province (grant no. H2018206096).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
JH and LZ designed the research. LL, JY, FL and FG
conducted the experiment. LL, LZ and JH analyzed the data. LZ and
JH wrote the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
All mice were treated according to the guidelines of
Institutional Animal Care and Use Committee of the Third Hospital
of Hebei Medical University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Ramzy MM, Abdalla AM, Zenhom NM, Okasha
AM, Abdelkafy AE and Saleh RK: Therapeutic effect of liraglutide on
expression of CTGF and BMP-7 in induced diabetic nephropathy. J
Cell Biochem. 120:17512–17519. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Yaribeygi H, Mohammadi MT, Rezaee R and
Sahebkar A: Fenofibrate improves renal function by amelioration of
NOX-4, IL-18, and p53 expression in an experimental model of
diabetic nephropathy. J Cell Biochem. 119:7458–7469. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Forbes JM and Thorburn DR: Mitochondrial
dysfunction in diabetic kidney disease. Nat RevNephrol. 14:291–312.
2018.
|
|
4
|
Cybulsky AV: Endoplasmic reticulum stress,
the unfolded protein response and autophagy in kidney diseases. Nat
Rev Nephrol. 13:681–696. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yang W, Luo Y, Yang S, Zeng M, Zhang S,
Liu J, Han Y, Liu Y, Zhu X, Wu H, et al: Ectopic lipid
accumulation: Potential role in tubular injury and inflammation in
diabetic kidney disease. Clin Sci (Lond). 132:2407–2422. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Fornoni A, Merscher S and Kopp JB: Lipid
biology of the podocyte-new perspectives offer new opportunities.
Nat Rev Nephrol. 10:379–388. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Brosius FC III: New insights into the
mechanisms of fibrosis and sclerosis in diabetic nephropathy. Rev
Endocr Metab Disord. 9:245–254. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Soyal SM, Nofziger C, Dossena S, Paulmichl
M and Patsch W: Targeting SREBPs for treatment of the metabolic
syndrome. Trends Pharmacol Sci. 36:406–416. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Jun H, Song Z, Chen W, Zanhua R, Yonghong
S, Shuxia L and Huijun D: In vivo and in vitro effects of SREBP-1
on diabetic renal tubular lipid accumulation and RNAi-mediated gene
silencing study. Histochem Cell Biol. 131:327–345. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhou Z, He C and Wang J: Regulation
mechanism of Fbxw7-related signaling pathways (Review). Oncol Rep.
34:2215–2224. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kourtis N, Strikoudis A and Aifantis I:
Emerging roles for the FBXW7 ubiquitin ligase in leukemia and
beyond. Curr Opin Cell Biol. 37:28–34. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yumimoto K and Nakayama KI: Recent insight
into the role of FBXW7 as a tumor suppressor. Semin Cancer Biol.
67:1–15. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Fiore D, Piscopo C, Proto MC, Vasaturo M,
Piaz FD, Fusco BM, Pagano C, Laezza C, Bifulco M and Gazzerro P:
N6-Isopentenyladenosine inhibits colorectal cancer and improves
sensitivity to 5-fluorouracil-targeting FBXW7 tumor suppressor.
Cancers (Basel). 28:14562019. View Article : Google Scholar
|
|
14
|
Hao J, Liu S, Zhao S, Liu Q, Lv X, Chen H,
Niu Y and Duan H: PI3K/Akt pathway mediates high glucose-induced
lipogenesis and extracellular matrix accumulation in HKC cells
through regulation of SREBP-1 and TGF-β1. Histochem Cell Biol.
135:173–181. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Du W, Wang N, Li F, Jia K, An J, Liu Y,
Wang Y, Zhu L, Zhao S and Hao J: STAT3 phosphorylation mediates
high glucose-impaired cell autophagy in an HDAC1-dependent and
-independent manner in schwann cells of diabetic peripheral
neuropathy. FASEB J. 33:8008–8021. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Racusen LC, Monteil C, Sgrignoli A,
Lucskay M, Marouillat S, Rhim JG and Morin JP: Cell lines with
extended in vitro growth potential from human renal proximal
tubule: Characterization, response to inducers, and comparison with
established cell lines. J Labo Clin Med. 129:318–329. 1997.
View Article : Google Scholar
|
|
17
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ceriello A: Postprandial hyperglycemia and
diabetes complications: Is it time to treat? Diabetes. 541:1–7.
2005. View Article : Google Scholar
|
|
19
|
Zhuang L, Jin G, Hu X, Yang Q and Shi Z:
The inhibition of SGK1 suppresses epithelial-mesenchymal transition
and promotes renal tubular epithelial cell autophagy in diabetic
nephropathy. Am J Transl Res. 118:4946–4956. 2019.
|
|
20
|
Brenachot X, Ramadori G, Ioris RM,
Veyrat-Durebex C, Altirriba J, Aras E, Ljubicic S, Kohno D,
Fabbiano S, Clement S, et al: Hepatic protein tyrosine phosphatase
receptor gamma links obesity-induced inflammation to insulin
resistance. Nat Commun. 8:18202017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhao J, Xiong X, Li Y, Liu X, Wang T,
Zhang H, Jiao Y, Jiang J, Zhang H, Tang Q, et al: Hepatic F-box
protein FBXW7 maintains glucose homeostasis through degradation of
fetuin-A. Diabetes. 67:818–830. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tu Lu-Mei LM, Wang Y and Yan-Ling MU:
Expression of FBXW7 in diabetic cardiomyopathy. Chin J
Pathophysiol. 34:2271–2276. 2018.
|
|
23
|
Gao C, Fan F, Chen J, Long Y, Tang S,
Jiang C and Xu Y: FBW7 regulates the autophagy signal in mesangial
cells induced by high glucose. Biomed Res Int. 2019:60615942019.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Tu K, Zheng X, Yin G, Zan X, Yao Y and Liu
Q: Evaluation of Fbxw7 expression and its correlation with
expression of SREBP-1 in a mouse model of NAFLD. Mol Med Rep.
6:525–530. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hao J, Zhu L, Zhao S, Liu S, Liu Q and
Duan H: PTEN ameliorates high glucose-induced lipid deposits
through regulating SREBP-1/FASN/ACC pathway in renal proximal
tubular cells. Exp Cell Res. 317:1629–1639. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Sundqvist A, Bengoechea-Alonso MT, Ye X,
Lukiyanchuk V, Jin J, Harper JW and Ericsson J: Control of lipid
metabolism by phosphorylation-dependent degradation of the SREBP
family of transcription factors by SCF(Fbw7). Cell Metab.
1:379–391. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Bengoechea-Alonso MT and Ericsson J: The
phosphorylation-dependent regulation of nuclear SREBP1 during
mitosis links lipid metabolism and cell growth. Cell Cycle.
15:2753–2765. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Jeon TI, Esquejo RM, Roqueta-Rivera M,
Phelan PE, Moon YA, Govindarajan SS, Esau CC and Osborne TF: An
SREBP-responsive microRNA operon contributes to a regulatory loop
for intracellular lipid homeostasis. Cell Metab. 18:51–61. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yeh CH, Bellon M and Nicot C: FBXW7: a
critical tumor suppressor of human cancers. Mol Cancer. 17:1152018.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lei S, Su W, Xia ZY, Wang Y, Zhou L, Qiao
S, Zhao B, Xia Z and Irwin M: Hyperglycemia-Induced Oxidative
Stress Abrogates Remifentanil Preconditioning-Mediated
Cardioprotection in Diabetic Rats by Impairing Caveolin-3-Modulated
PI3K/Akt and JAK2/STAT3 Signaling. Oxid Med Cell Longev.
2019:98363022019. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hao J, Zhu L, Li F, Liu Q, Zhao X, Liu S,
Xing L, Feng X and Duan H: Phospho-mTOR: A novel target in
regulation of renal lipid metabolism abnormality of diabetes. Exp
Cell Res. 319:2296–2306. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Liu W, Hao J, Zhu L, Li F, Liu Q, Liu S,
Zhao S, Li H and Duan H: Phospho-GSK-3β is involved in the
high-glucose-mediated lipid deposition in renal tubular cells in
diabetes. Int J Biochem Cell Biol. 45:2066–2075. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Suryo Rahmanto A, Savov V, Brunner A,
Bolin S, Weishaupt H, Malyukova A, Rosén G, Čančer M, Hutter S and
Sundström A: FBW7 suppression leads to SOX9 stabilization and
increased malignancy in medulloblastoma. EMBO J. 35:2192–2212.
2016. View Article : Google Scholar : PubMed/NCBI
|