Introduction
Neuroblastoma (NB) originates from undifferentiated
neural crest cells and is the most common extracranial solid tumor
in children, accounting for 15% of deaths from pediatric tumors
(1). The clinical manifestations of
NB are variable, ranging from a spontaneously developing mass to
aggressive drug-resistant tumors, associated with high levels of
metastasis and poor prognosis rates (2). In ~50% of patients with NB, metastasis
occurs in the bone marrow, skin, liver and lymph nodes (3). Amplification of the N-myc
proto-oncogene (MYCN) has been identified in 20–30% of patients,
and is associated with advanced disease stage and poor prognosis
(4). The transcription factor N-Myc
(encoded by MYCN) contributes to the malignant phenotype by both
up- and downregulating target genes (5). According to previous studies, N-myc
directly interacts with the microRNA (miRNA/miR)-17–92 promoter
site. Furthermore, there is a positive association between MYCN
expression and members of the miR-17- 92 cluster in NB cell lines
and primary tumors (6–8).
miRNAs belong to a group of single-stranded,
non-coding RNAs of 18–25 nucleotides in length, which regulate gene
expression at the post-transcriptional level (9). Abnormal regulation of miRNA expression
has been linked to the development of various diseases, including
NB (10–12). These molecules exert their
regulatory effects by interacting with the 3′untranslated regions
(3′UTRs) of mRNAs, and either inhibiting translation or
facilitating mRNA degradation (13). Exosomes are membranous extracellular
microvesicles with a diameter of 30–150 nm that contain various
macromolecules, including miRNAs (14). These microvesicles originate from a
variety of different cell types, and can act as key mediators
between cancerous cells and facilitate stroma intercellular
communication by delivering molecular cargo into the tumor
microenvironment (15).
Exosome-mediated miRNA transfer is considered an efficient strategy
for miRNAs to exert their effects on different tumor progression
processes (16). A number of
recently reported studies have identified exosomal miRNAs as key
biomarkers for the detection and prognosis of NB (17,18).
Although exosomes from MYCN-amplified NB cells are able transmit
aggressive cancer cell phenotypes (19), their regulatory mechanisms in NB
cell functional variation and miRNA expression modification are not
fully understood.
The present study aimed to investigate the transfer
of aggressive cell phenotypes from MYCN-amplified to non-MYCN
amplified NB cells via exosome-mediated miRNAs. It was hypothesized
that anti-oncogenes in recipient cells are targeted by exosomal
miRNAs, which are present in high levels in the exosomes of
MYCN-amplified NB cells.
Materials and methods
Cell culture
Human SH-SY5Y NB cells were procured from American
Type Culture Collection, and SK-N-BE(2) cells were purchased from
The Cell Bank of Type Culture Collection of The Chinese Academy of
Sciences. The cells were cultured in DMEM (Hyclone; Cytiva)
containing FBS (10%; Gibco; Thermo Fisher Scientific, Inc.),
penicillin and streptomycin (100 U/ml and 100 µg/ml, respectively),
and maintained at 37°C in an atmosphere of 5% CO2.
Routine passage was conducted when the cells had reached 80–90%
confluency.
Bioinformatics analysis
The miRNA expression profiles of MYCN-amplified and
non-MYCN amplified NB tissues were obtained directly from the
original text of three previous studies (6–8), with
no additional processing. To identify the differential expression
of miR-17-92a between these tissue types, miRNAs with significantly
different expression levels were summarized from the results of
these three studies, and then subjected to further analysis. The
TargetScan (http://www.targetscan.org/vert_72/), DIANA (http://diana.imis.athena-innovation.gr/DianaTools/index.php?r=tarbase/index)
and miRanda (http://mirdb.org/) databases were then
used to predict the target genes of these miRNAs. The
immunohistochemical profiles of PTEN (determined using a tissue
microarray containing human NB sections) obtained from the results
of a previously published article (20) were statistically analyzed to
identify the differential expression of PTEN in MYC-amplified NB
tissues (compared with their non-MYCN amplified counterparts).
Reverse transcription-quantitative
(RT-q) PCR
Total RNA was extracted from cells using a
TRIzol® RNA purification kit (Thermo Fisher Scientific,
Inc.). Reverse transcription was performed with 1 µg total RNA
using the miRNA 1st Strand cDNA Synthesis kit (Vazyme Biotech Co.,
Ltd.) for mature miRNA expression, and the TransScript®
qPCR kit (TransGen Biotech Co., Ltd.) for mRNA expression. Reverse
transcription was performed according to the manufacturer's
protocols. qPCR were subsequently performed with the ChamQ
Universal SYBR qPCR Master Mix (Vazyme Biotech Co., Ltd.),
according to the manufacturer's protocols. The thermocycling
conditions were 94 C for 5 min, then 40 cycles of 93°C for 30 sec,
55°C for 30 sec and 72°C for 60 sec. Relative mRNA and miRNA
expression levels were evaluated using the 2−∆∆Cq method
(21), and normalized to the levels
of GAPDH and U6, respectively. The primer sequences are presented
in Table I.
 | Table I.Primer sequences used for reverse
transcription- quantitative PCR. |
Table I.
Primer sequences used for reverse
transcription- quantitative PCR.
| Gene | Primer sequences
(5′→3′) |
|---|
| miR-17-5p | F:
GCGCAAAGTGCTTACAGTGC |
|
| R:
AGTGCAGGGTCCGAGGTATT |
| miR-18a | F:
CGCGTAAGGTGCATCTAGTGC |
|
| R:
AGTGCAGGGTCCGAGGTATT |
| miR-19a | F:
GCGTGTGCAAATCTATGCAA |
|
| R:
AGTGCAGGGTCCGAGGTATT |
| miR-19b | F:
CGTGTGCAAATCCATGCAA |
|
| R:
AGTGCAGGGTCCGAGGTATT |
| miR-20a | F:
GCGCGTAAAGTGCTTATAGTGC |
|
| R:
AGTGCAGGGTCCGAGGTATT |
| miR-92a | F:
GCGTATTGCACTTGTCCCG |
|
| R:
AGTGCAGGGTCCGAGGTATT |
| U6 | F:
AGAGAAGATTAGCATGGCCCCTG |
|
| R:
ATCCAGTGCAGGGTCCGAGG |
| PTEN | F:
GACCAGAGACAAAAAGGGAGTA |
|
| R:
ACAAACTGAGGATTGCAAGTTC |
| GAPDH | F:
TCCAGAGTGCAAGGCTTCAG |
|
| R:
GACAGCACGCAGTAGCAGTAG |
Isolation of exosomes by
ultracentrifugation
Exosomes were isolated from the cell culture
supernatants based on a previously reported procedure (22). Briefly, when the cells had reached
60–80% confluency, the media were replaced with DMEM containing 10%
exosome-depleted FBS (Systems Biosciences, LLC). After incubation
for 48 h, the supernatants were removed and centrifuged at 2,000 ×
g (10 min) and 10,000 × g (30 min) for the removal of the cells and
cellular debris, respectively. The resulting supernatant was
centrifuged again for 70 min (110,000 × g) to pellet the exosomal
fraction. The pellets were washed with PBS, followed by
ultracentrifugation for a further 70 min at 110,000 × g, and then
resuspended in PBS. The pellets were stored −80°C and each
centrifugation step was carried out at 4°C. For subsequent
experimentation, 50 µg SK-N-BE(2) cell exosomes were incubated with
50,000 SH-SY5Y cells for 48 h at 37°C. The untreated cells were
used as a control group.
Exosome characterization
Transmission electron microscopy
(TEM)
Isolated exosomes (10 µl) were placed in copper
grids and allowed to settle for 2 min at room temperature. The
samples were blotted to remove excess fluid, followed by staining
with phosphotungstic acid (3%, v/v) in ddH2O for 1 min
at room temperature. The samples were carefully blotted with soft
paper before being left in a dry environment for 10–15 min at room
temperature. Finally, the samples were evaluated using the
JEM1200-EX transmission electron microscope (JEOL, Ltd.) at a
voltage of 80 kV. In total, five fields of view were randomly
selected for each sample (magnification, ×200,000).
Nanoparticle tracking analysis
(NTA)
To obtain an estimated number of vesicles
(≤1×107), exosomal dilutions were prepared with PBS at a
factor of 500. Exosome concentration and size were determined using
the ZetaView® PMX 110 and the corresponding software
(ZetaView® 8.04.02 SP2; both Analytik Jena AG).
Western blot analysis
Total cellular proteins were extracted using RIPA
lysis buffer (Wuhan Servicebio Technology Co., Ltd.) 96 h after
transfection. The native protein lysate was collected, and the
protein concentration was measured using a BCA protein
concentration assay kit (Wuhan Servicebio Technology Co., Ltd.),
according to the manufacturer's instructions. Proteins (20–40
µg/per well) were separated via SDS-PAGE on a 10% gel. The
separated proteins were transferred to PVDF membranes (EMD
Millipore), which were then blocked for 2 h in non-fat milk (5%) in
PBS with 0.1% Tween-20 (PBST) at room temperature. The membranes
were treated with the following primary antibodies: Anti-β-actin
(1:1,000; cat. no. GB12001; Wuhan Servicebio Technology Co., Ltd.),
anti-CD63 (1:1,000; cat. no. NBP2-67425; Novus Biologicals, LLC),
anti-phosphorylated (p)-Akt Ser473 (1:1,000; cat. no. 4060; Cell
Signaling Technology, Inc.) anti-tumor susceptibility gene 101
protein (TSG101; 1:1,000; cat. no. ab125011; Abcam), anti-PTEN
(1:1,000; cat. no. 9188; Cell Signaling Technology, Inc.) and
anti-Akt (1:1,000; cat. no. 4691; Cell Signaling Technology, Inc.).
After incubation at 4°C for 24 h, the membranes were washed three
times with TBS with 0.1% Tween-20, followed by a 1-h incubation at
~25°C with a HRP-conjugated goat anti-rabbit IgG secondary antibody
(1:1,000; cat. no. GB23303; Wuhan Servicebio Technology Co., Ltd.).
An ECL immunoblotting kit (Dalian Meilun Biology Technology Co.,
Ltd.) was used to detect protein expression levels relative to the
β-actin loading control. The proteins were visualized using
chemiluminescent film and analyzed using ImageJ software (version
1.52; National Institutes of Health). Each experiment was
independently conducted in triplicate.
Transfection
The miR-17-5p mimics
(5′-CAAAGUGCUUACAGUGCAGGUAG-3′), inhibitor
(5′-CUACCUGCACUGUAAGCACUUUG-3′) and negative control [mimics NC
(5′-UUCUCCGAACGUGUCACGUTT-3′) and inhibitor NC
(5′-CAGUACUUUUGUGUAGUACAA-3′)] were acquired from Shanghai
GenePharma Co., Ltd. For subsequent experimentation, SH-SY5Y cells
were seeded on a 6-well plate at a density of 1×106
cells/ml and cultured overnight at 37°C. The next day, cells were
transfected with these molecular products (100 nM mimic, inhibitor
or NC) using TransIntro™ EL Transfection Reagent (Beijing Transgen
Biotech Co., Ltd.), following the manufacturer's protocols. The
media were replaced with fresh growth medium and transfection was
carried out for 6 h. Cells were cultured at 37°C with 5%
CO2, and the medium was replaced after 24 h. Then, 48 h
after changing the medium, the cells were harvested for subsequent
experiments. Transfection efficiency was confirmed by RT-qPCR.
Cellular proliferation assay
In vitro cellular proliferative capacity was
measured using the Cell Counting Kit-8 (CCK-8; Dalian Meilun
Biology Technology Co., Ltd.), according to the manufacturer's
protocols. Transfected cells were seeded into 96-well plates to a
final volume of 100 µl complete medium (3×103
cells/well; five wells per concentration) and cultured for 4 days;
10 µl CCK-8 reagent was added to each well at the indicated time
intervals (0, 1, 2, 3 and 4 days), after which the cultured plates
were incubated for a further 2 h at 37°C. The OD of each well was
recorded at 450 nm using a microplate reader (Synergy H1; BioTek
Instruments, Inc.).
Colony formation assay
Transfected cells (500 cells/well) were seeded into
6-well plates in DMEM enriched with 10% FBS, followed by incubation
at 37°C (5% CO2). After culturing for 2 weeks, the cells
were washed with PBS, fixed with 4% paraformaldehyde for 15 min at
37°C and stained with 1% crystal violet for 25 min at room
temperature. Colonies were defined as >50 cells and counted
under a fluorescence microscope (magnification, ×400) in five
randomly selected fields.
Cellular migration assay
The migratory capacity of SH-SY5Y cells was observed
using Transwell chambers with a pore size of 8 µm. After 24 h,
transfected cells were isolated and resuspended in serum-free DMEM.
The resuspended cells (~2×104 cells/well) were added
into the upper chambers, and the lower chambers were filled with
500 µl DMEM containing 20% FBS. The plates were cultured for 12–20
h at 37°C with 5% CO2. The migrated cells were fixed
with 4% paraformaldehyde for 25 min, followed by staining with 1%
crystal violet for 20 min. Both the fixing and staining assays were
conducted at room temperature. Migratory cells were counted in five
randomly selected fields, and images were captured with a light
microscope (magnification, ×200; Carl Zeiss AG).
Dual-luciferase reporter assay
miR-17-5p was predicted to interact with the 3′-UTR
of PTEN. The wild-type (wt) and mutant (mut) 3′-UTR of PTEN mRNA
were purchased from Shanghai GenePharma Co., Ltd., and were
introduced between the NotI and XhoI restriction
sites of the psiCHECK-2 vector (Promega Corporation), and the
resulting vectors were termed psiCHECK-2-PTEN-wt and
psiCHECK-2-PTEN-mut (the binging site ‘GCA CUU U’ was replaced by
‘CGU GAA A’), accordingly. The following primers were used for the
amplification of particular fragments: PTEN-wt forward,
5′-CACAACTCGAGGCCCTGTACCATCCCAAGTC-3′ and reverse,
5′-AAGGAAAAAAGCGGCCGCACTGGCAGGTAGAAGGCAAC-3′; and PTEN-mut forward,
5′-CTAGAAATTTTCACGTTAATGTTCATAACGATGGCTGT-3′ and reverse,
5′-ACATTAACGTGAAAATTTCTAGAACTAAACATTAAAC-3′. 293T cells
(1×105 cells/well; American Type Culture Collection)
were co-transfected with 0.1 mg psiCHECK-2-PTEN-wt or 0.1 mg
psiCHECK-2-PTEN-mut and 10 nm miR-17-5p mimics or the corresponding
NC mimic using LipoFiter™ (Hanbio Biotechnology Co., Ltd.); 48 h
post-transfection, the cells were collected and evaluated using the
Dual-Luciferase reporter assay (Promega Corporation), according to
the manufacturer's protocols. Firefly luciferase activity was
normalized to that of Renilla luciferase.
Statistical analysis
The obtained data were statistically analyzed using
SPSS version 21.0 (IBM Corp.) and graphically represented using
GraphPad Prism 7 (GraphPad Software, Inc.). Each experiment was
conducted in triplicate and the data are presented as the mean ±
SD. An unpaired t-test was used to evaluate comparisons between two
independent groups. One-way ANOVA was used to assess variations
among multiple groups, followed by Tukey's multiple comparison
tests. The P-values were two-sided, and P<0.05 was considered to
indicate a statistically significant difference.
Results
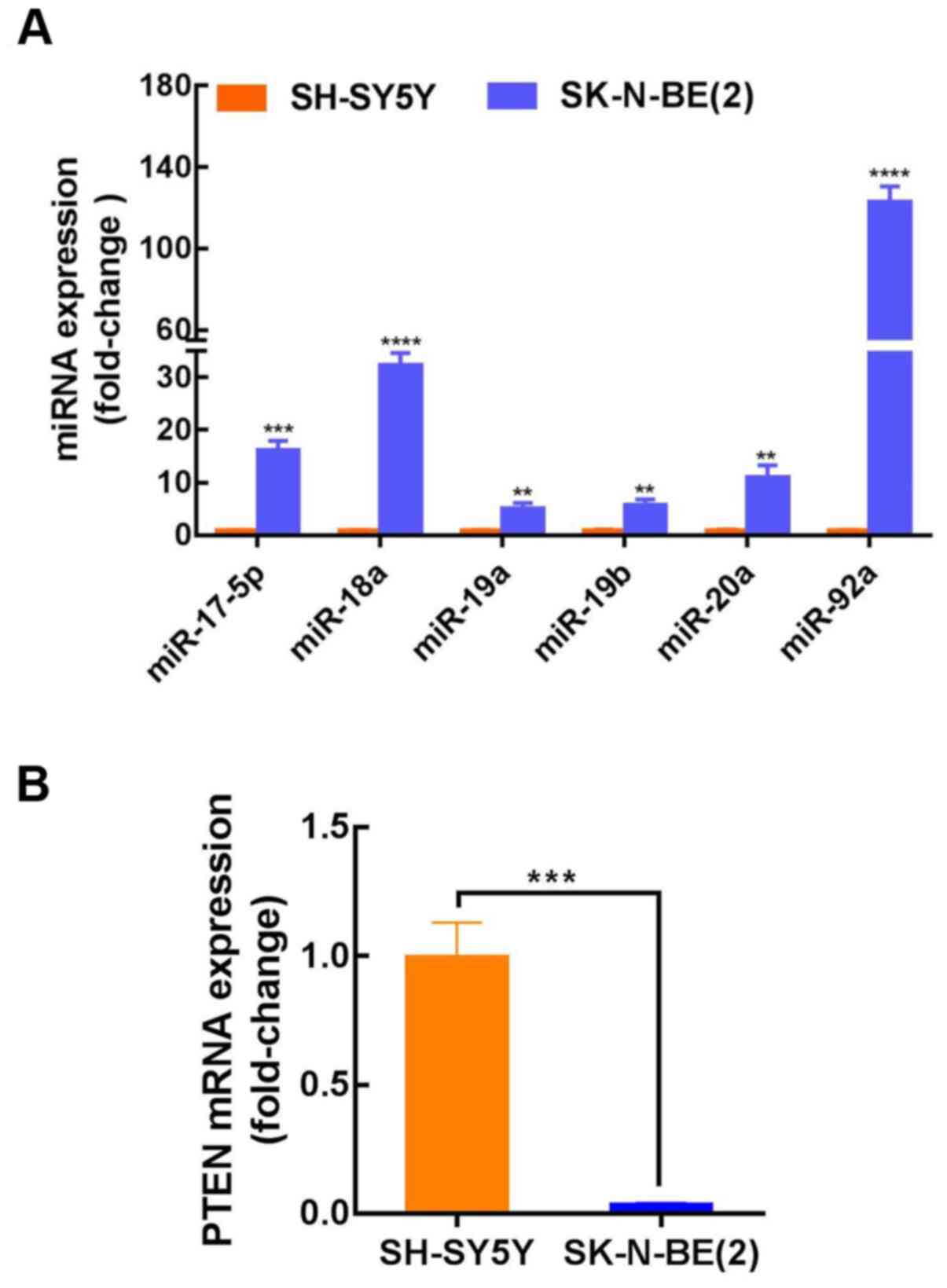
PTEN and miR-17–92 are involved in
NB
The published results from three independent studies
were collected and analyzed (Table
II), and the expression of the miR-17–92 cluster in
MYCN-amplified NB tissues was found to be considerably higher than
that in non-MYCN amplified NB. For further clarification of the
miR-17–92 cluster mechanism in NB, three prediction databases
(TargetScan, DIANA and miRanda) were used to identify potential
binding targets. The target gene prediction results indicated a
conserved binding site for miR-17-5p, miR-92a, miR-20a, miR-19a and
miR-19b within the PTEN 3′-UTR (Table
III). However, no PTEN binding sites were identified for
miR-18a. SK-N-BE(2) and SH-SY5Y cells were then used to further
investigate the role of the miR-17–92 cluster in NB progression.
SK-N-BE(2) cells exhibit MYCN amplification and are known to be
more aggressive than non-MYCN-amplified SH-SY5Y cells. The results
indicated that the expression level of miR-17-92 in SK-N-BE(2)
cells (MYCN-amplified) was considerably higher than that in SH-SY5Y
cells (non-MYCN amplified); however, the expression of PTEN was
considerably decreased (Fig. 1A and
B). These findings demonstrated that the expression of PTEN was
lower, while miR-17-92 expression was elevated in MYCN-amplified
NB, and that miR-17-5p, miR-92a, miR-20a, miR-19a and miR-19b may
target PTEN. To confirm these findings, miR-17-5p was selected for
further investigation. The expression level of miR-17-5p in
SK-N-BE(2) cells was ~16-fold higher than that in SH-SY5Y cells,
though there are almost no published reports on the role of
miR-17-5p in NB.
| Table II.Differentially expressed miRNAs in
human NB tumor. |
Table II.
Differentially expressed miRNAs in
human NB tumor.
| miRNA | Levels | Tissue | Refs. |
|---|
| miR-17-5p | Up | MYCN-amplified
NB | (6,7) |
| miR-18a | Up | MYCN-amplified
NB | (6–8) |
| miR-19a | Up | MYCN-amplified
NB | (7,8) |
| miR-19b | Up | MYCN-amplified
NB | (7) |
| miR-20a | Up | MYCN-amplified
NB | (7,8) |
| miR-92a | Up | MYCN-amplified
NB | (6–8) |
| Table III.Putative binding sites for miRNAs in
the 3′UTR of PTEN. |
Table III.
Putative binding sites for miRNAs in
the 3′UTR of PTEN.
| Position in the
3′UTR of PTEN | Predicted
consequential pairing of target region and miRNA |
|---|
| Position
272–278 |
|
| PTEN
3′UTR |
5′-GGAUUAAUAAAGAUGGCACUUUC-3′ |
|
hsa-miR-17-5p |
3′-GAUGGACGUGACUUCGUGAAAC-5′ |
| Position
272–278 |
|
| PTEN
3′UTR |
5′-GGAUUAAUAAAGAUGGCACUUUC-3′ |
|
hsa-miR-20a-5p |
3′-GAUGGACGUGAUAUUCGUGAAAU-5′ |
| Position
1221–1228 |
|
| PTEN
3′UTR |
5′-AAUGAAUUUUGCAGUUUUGCACA-3′ |
|
hsa-miR-19a-3p |
3′-AGUCAAAACGUAUCUAAACGUGU-5′ |
| Position
1221–1228 |
|
| PTEN
3′UTR |
5′-AAUGAAUUUUGCAGUUUUGCACA-3′ |
|
hsa-miR-19b-3p |
3′-AGUCAAAACGUACCUAAACGUGU-5′ |
| Position
4003–4010 |
|
| PTEN
3′UTR |
5′-AGUAAAUGAAAAAAUGUGCAAUA-3′ |
|
hsa-miR-92a-3p |
3′-UGUCCGGCCCUGUUCACGUUAU-5′ |
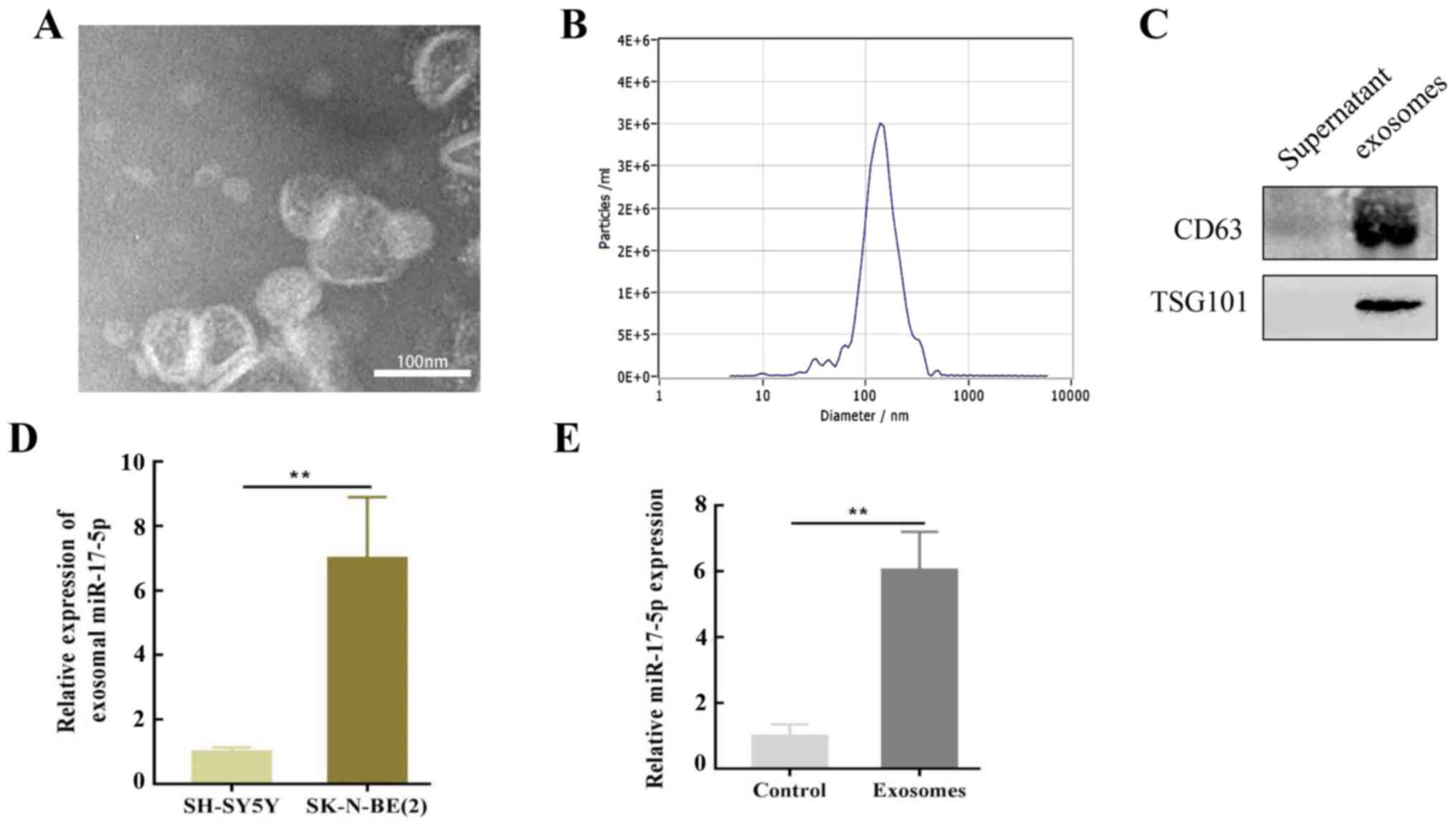
SK-N-BE(2) cells deliver miR-17-5p to
SH-SY5Y cells via exosomes
It was hypothesized that SK-N-BE(2) cells may
influence SH-SY5Y cells via exosomes secreted into the culture
medium. To verify this assumption, exosomes were isolated from
SK-N-BE(2) cells using differential ultracentrifugation and
characterized by TEM. The TEM results revealed that the purified
exosomes were primarily cup-shaped in structure, with a diameter of
50–150 nm. An abnormal lipid bilayer was also observed (Fig. 2A), and NTA revealed that the number
of purified exosomes peaked at a mean diameter of 50–150 nm
(Fig. 2B). Moreover, these vesicles
were enriched with exosomal markers, including CD63 and
TSG101 (Fig. 2C).
To determine the molecular mechanism of SK-N-BE(2)
cell-induced regulation, SH-SY5Y cells were co-cultured with
exosomes isolated from SK-N-BE (2)
cells, and RT-qPCR was performed to evaluate the levels of
miR-17-5p expression. The results revealed that miR-17-5p was
upregulated in SK-N-BE(2)-associated exosomes compared with those
from SH-SY5Y cells (Fig. 2D). This
upregulation in MYCN-amplified NB cell exosomes may reveal that
oncomiRs and the cancerous phenotype can be transferred to other
cells. Notably, a high expression level of miR-17-5p was observed
in SH-SY5Y cells co-cultured with SK-N-BE(2)-derived exosomes
(Fig. 2E).
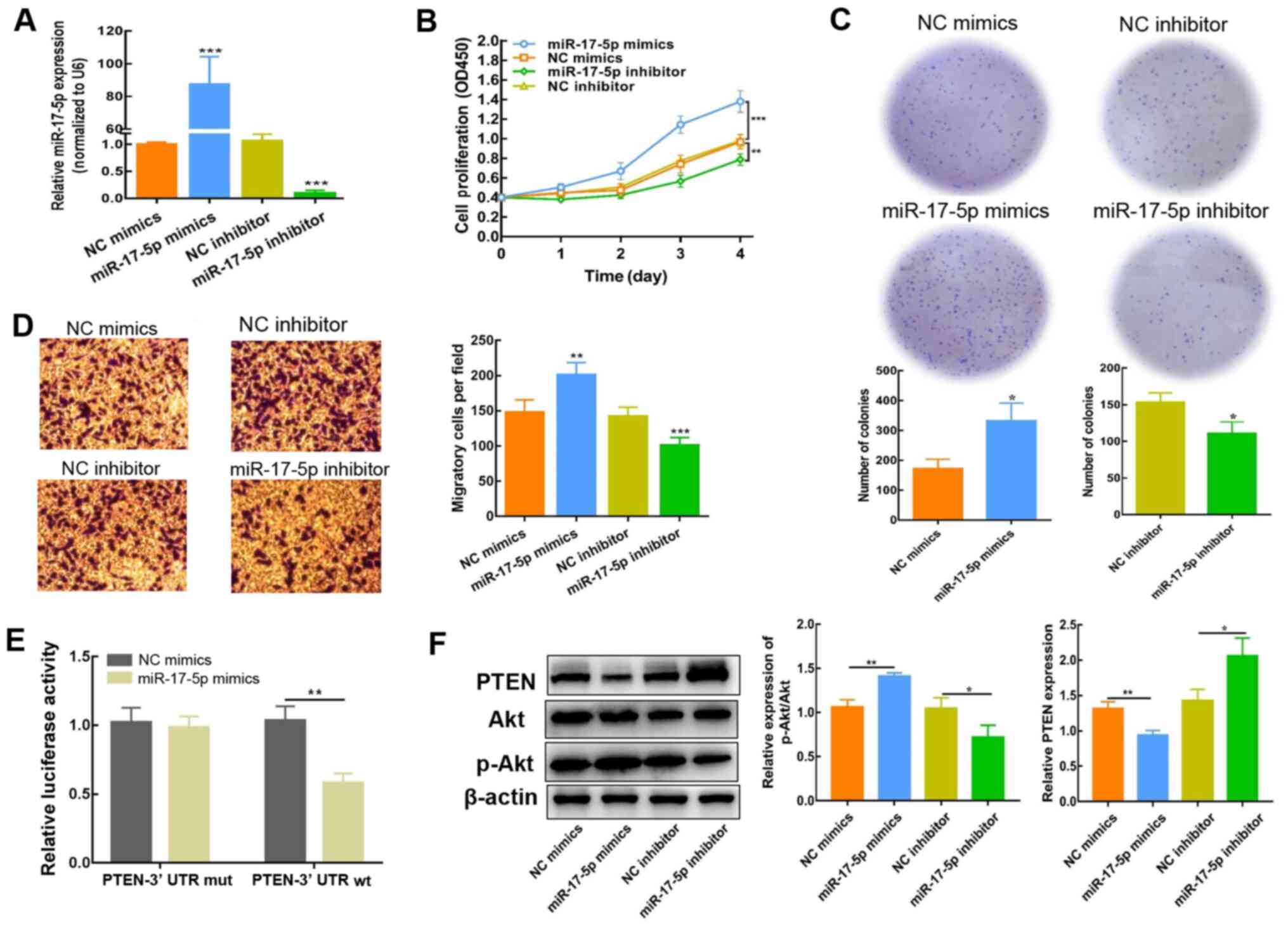
miR-17-5p upregulation enhances the
proliferative and migratory potential of SH-SY5Y cells
To determine whether miR-17-5p impacts the
biological activity of NB cells, SH-SY5Y cells were transfected
with miR-17-5p mimics or inhibitor; miR-17-5p expression was
detected using RT-qPCR 2 days post-transfection. The expression of
miR-17-5p was significantly increased following transfection with
miR-17-5p mimics, but reduced as a result of miR-17-5p inhibitor
transfection (Fig. 3A). CCK-8,
colony formation and Transwell assays were subsequently conducted
to evaluate the influence of miR-17-5p expression on the
proliferation and migration of SH-SY5Y cells. As depicted in
Fig. 3B-D, high miR-17-5p
expression enhanced SH-SY5Y cell proliferation and migration. By
contrast, inhibiting miR-17-5p expression resulted in a decreased
level of proliferation and migration. These findings demonstrated
that miR-17-5p significantly influenced the regulation of NB
cellular motility, including proliferation and metastatic
capacity.
miR-17-5p stimulates the PI3K/Akt
signaling cascade in NB cells by downregulating PTEN
For further evaluation of the regulatory effect of
miR-17-5p on PTEN, the predicted interaction sites between
miR-17-5p and PTEN were identified using TargetScan. As indicated
in Table III, the PTEN 3′UTR
contains a number of miR-17-5p interaction sites. Moreover, a
dual-luciferase reporter assay was conducted using the PTEN-Wt
3′-UTR sequence, which was co-transfected into NB cells with
miR-17-5p mimics or NC. Compared with the NC mimics, the luciferase
activity of PTEN-Wt was blocked in the presence of miR-17-5p mimics
(Fig. 3E), which indicated that
miR-17-5p interacted with PTEN. Consistently, the results of
western blot analysis showed that the expression of PTEN protein in
SH-SY5Y cells was decreased, and that the p-Akt/Akt ratio was
significantly elevated after transfection with miR-17-5p mimics
(Fig. 3F). By contrast, PTEN
protein expression levels were elevated, whereas the p-Akt/Akt
ratio was reduced, following miR-17-5p inhibition, demonstrating
that PTEN expression was attenuated via miR-17-5p. Collectively,
these results indicated that the PI3K/Akt signaling cascade was
activated by miR-17-5p via the downregulation of PTEN.
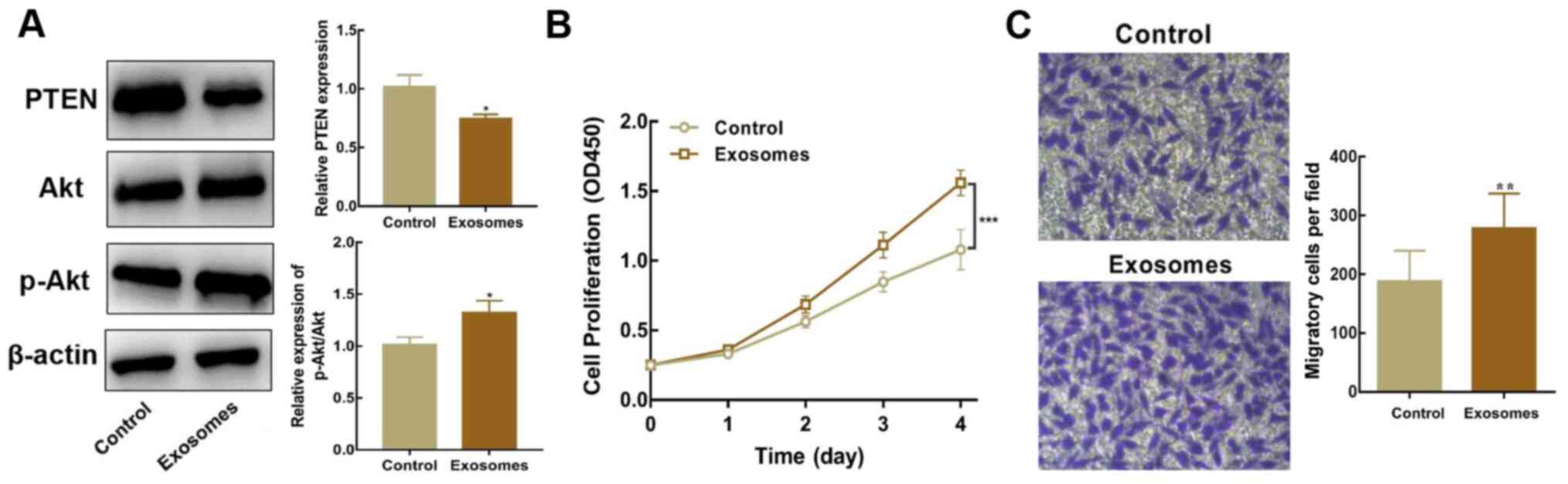
Exosomal miR-17-5p enhances the
proliferative and migratory abilities of SH-SY5Y cells
Exosomes isolated from SK-N-BE(2) cells were
co-cultured with SH-SY5Y cells to identify whether exosomal
miR-17-5p facilitates the activation of the PI3K/Akt signaling
cascade (PTEN-mediated), and subsequently regulates the
proliferation and migration of SH-SY5Y cells. Western blot analysis
revealed that PTEN expression was lower, whereas the p-Akt/Akt
ratio was higher, in SH-SY5Y cells co-cultured with exosomes than
in untreated cells (Fig. 4A).
Moreover, SH-SY5Y cells co-cultured with exosomes overexpressing
miR-17-5p exhibited higher proliferative and migratory abilities
(Fig. 4B and C). In summary, these
results indicated that inhibiting PTEN expression and stimulating
the PI3K/Akt signaling cascade via exosomal miR-17-5p enhanced the
proliferative and migratory potential of SH-SY5Y cells.
Discussion
MYCN amplification is the most effective prognostic
marker of adverse disease outcome in NB (23). MYCN expression has been found to
positively correlate with metastatic behavior,
epithelial-mesenchymal transition, cell cycle progression and
impaired immune surveillance in NB (4). Due to its secondary structure and no
obvious sites for the binding of small molecules, the
identification of therapeutics targeting MYCN has been challenging.
To date, the direct targeting of myc family proteins has been
unsuccessful, hence the potential for indirect targeting is
currently under investigation (2,4).
Exosomes are cell-secreted nanoscale vesicles that are derived from
NB cells (MYCN-amplified), which mediate intercellular
communication and impart aggressive NB phenotypes (24). Exosomes secreted from cancerous
cells may serve to mediate the transfer of these phenotypes to
sensitive recipient cells via miRNAs (25,26).
However, there is still ambiguity regarding the ability of NB cells
to transfer phenotypes in this manner. The present study revealed
that exosomes derived from MYCN-amplified NB cells, with elevated
miR-17-5p expression, inhibited PTEN and activated the PI3K/Akt
signaling cascade, thus enhancing the proliferative and migratory
capacities of non-MYCN amplified cells.
There is evidence to suggest that N-myc directly
interacts with the promoter region of the miR-17-92 cluster, which
results in its upregulation (27).
The precursor transcript of the miR-17-92 gene comprises six tandem
stem-loop hairpins that produce six mature miRNAs, including
miR-92a, miR-20a, miR-19a, miR-19b, miR-18a and miR-17-5p (28). Consistently, the present study
demonstrated that miR-17-92 cluster expression was upregulated in
MYCN-amplified SK-N-BE(2) cells. Moreover, SK-N-BE(2)-derived
exosomes were found to transfer miR-17-5p from the oncogenic
miR-17-92 cluster, promoting the elevation of miR-17-5p expression
in SH-SY5Y cells (non-MYCN amplified). Notably, the transfer of
miR-17-5p via SK-N-BE(2)-derived exosomes enhanced the suppression
of PTEN expression in SH-SY5Y cells.
Haug et al (24) analyzed the miRNA profiles of
exosomes isolated from MYCN-amplified NB cells. Functional
enrichment analysis revealed several well-characterized pathways,
including the PTEN/PI3K/Akt signaling cascade. In previous years,
several reports have also established the importance of
PTEN/PI3K/Akt signaling in NB survival, angiogenesis, proliferation
and invasion, including its association with MYCN (29,30).
In the present study, the results of the dual-luciferase reporter
assay demonstrated that PTEN was targeted, and its expression was
downregulated, by miR-17-5p. Furthermore, Paul et al
(20) immunohistochemically
assessed the expression of PTEN using a tissue microarray
containing human NB sections, revealing that PTEN was significantly
downregulated in MYCN-amplified NB tissues. In the current study,
miR-17-5p-upregulated exosomes derived from MYCN-amplified NB cells
increased the proliferative and migratory potential of non-MYCN
amplified NB cells in vitro. Previous studies have revealed
that GDNF family receptor α2 plays a key role in the regulation of
NB development by suppressing PTEN via the activation of the
PI3K/Akt signaling cascade (31–33).
In NB cells, the upregulation of miR-17-5p tends to elevate the
levels of Akt phosphorylation, which indicates that in NB, the
PI3K/Akt signaling cascade is activated via miR-17-5p.
It is commonly known that within a single tumor, the
presence of different cell cancer subsets with unique genotypes
results in intra-tumor heterogeneity (34,35).
Clonal subsets of tumor cells with different mutant states may also
exist in a single NB tumor. Exosomes may facilitate the interaction
between different subsets of tumor cell clones, as well as with
non-cancerous cells in the tumor microenvironment (19). The present study was also conducted
in the context of intra-tumor heterogeneity. However, as the
research is still at the preclinical stage, the internalization of
PKH67-labeled exosomes by SH-SY5Y cells was not observed. Although
several articles have reported that miR-17-5p can be regulated by
N-myc (36–38), this was not confirmed using SH-SY5Y
cells in the present study, and the mechanisms by which exosomal
miRNAs influence NB are yet to be elucidated. Therefore, additional
experimental approaches, such as Transwell invasion assays,
apoptosis analysis and animal studies, are required to reveal the
intrinsic pathways of exosomal miR-17-5p in NB, both in
vitro and in vivo.
In conclusion, the results of the present study
suggested that exosomes derived from MYCN-amplified SK-N-BE(2)
cells transfer miR-17-5p into non-MYCN amplified SH-SY5Y cells,
promoting the proliferation and migration of non-MYCN amplified NB
cells. Therefore, exosomes from SK-N-BE(2) cells with upregulated
miR-17-5p expression may be a key candidate target for the
treatment of NB.
Acknowledgements
Not applicable.
Funding
This work was supported by Qingdao Minsheng Science
and Technology Plan Project Task Book (grant no. 18-6-1-71-nsh) and
Qingdao Outstanding Health Professional Development.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
LH and HL conceived the project and supervised the
experiments. WC, BY, YZ and YH performed the experiments. JY, XH
and JZ provided technical support. WC, LY and LS performed
statistical analysis. WC and XH wrote the manuscript with help from
all of the authors. WC, XH and BY confirm the authenticity of all
the raw data. HL and LH drafted the article and critically revised
it for important intellectual content. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Maris JM, Hogarty MD, Bagatell R and Cohn
SL: Neuroblastoma. Lancet. 369:2106–2120. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Pastor ER and Mousa SA: Current management
of neuroblastoma and future direction. Crit Rev Oncol Hematol.
138:38–43. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Johnsen JI, Dyberg C and Wickstrom M:
Neuroblastoma-A neural crest derived embryonal malignancy. Front
Mol Neurosci. 12:92019. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Huang M and Weiss WA: Neuroblastoma and
MYCN. Cold Spring Harb Perspect Med. 3:a0144152013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Buechner J and Einvik C: N-myc and
noncoding RNAs in neuroblastoma. Mol Cancer Res. 10:1243–1253.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Schulte JH, Horn S, Otto T, Samans B,
Heukamp LC, Eilers UC, Krause M, Astrahantseff K, Klein-Hitpass L,
Buettner R, et al: MYCN regulates oncogenic MicroRNAs in
neuroblastoma. Int J Cancer. 122:699–704. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bray I, Bryan K, Prenter S, Buckley PG,
Foley NH, Murphy DM, Alcock L, Mestdagh P, Vandesompele J, Speleman
F, et al: Widespread dysregulation of miRNAs by MYCN amplification
and chromosomal imbalances in neuroblastoma: Association of miRNA
expression with survival. PLoS One. 4:e78502009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Mestdagh P, Fredlund E, Pattyn F, Schulte
JH, Muth D, Vermeulen J, Kumps C, Schlierf S, De Preter K, Van Roy
N, et al: MYCN/c-MYC-induced microRNAs repress coding gene networks
associated with poor outcome in MYCN/c-MYC-activated tumors.
Oncogene. 29:1394–1404. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Babaei K, Shams S, Keymoradzadeh A, Vahidi
S, Hamami P, Khaksar R, Norollahi SE and Samadani AA: An insight of
microRNAs performance in carcinogenesis and tumorigenesis; an
overview of cancer therapy. Life Sci. 240:1170772020. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhou X, Lu H, Li F, Hao X, Han L, Dong Q
and Chen X: MicroRNA-429 inhibits neuroblastoma cell proliferation,
migration and invasion via the NF-κB pathway. Cell Mol Biol Lett.
25:52020. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhao X, Gu H, Wang L, Zhang P, Du J, Shen
L, Jiang D, Wang J, Li X, Zhang S, et al: MicroRNA-23a-5p mediates
the proliferation and differentiation of C2C12 myoblasts. Mol Med
Rep. 22:3705–3714. 2020.PubMed/NCBI
|
|
12
|
Tang D, Gao W, Yang J, Liu J, Zhao J, Ge
J, Chen Q and Liu B: miR181d promotes cell proliferation via the
IGF1/PI3K/AKT axis in glioma. Mol Med Rep. 22:3804–3812.
2020.PubMed/NCBI
|
|
13
|
Galardi A, Colletti M, Businaro P,
Quintarelli C, Locatelli F and Di Giannatale A: MicroRNAs in
neuroblastoma: Biomarkers with therapeutic potential. Curr Med
Chem. 25:584–600. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Pegtel DM and Gould SJ: Exosomes. Annu Rev
Biochem. 88:487–514. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
McAndrews KM and Kalluri R: Mechanisms
associated with biogenesis of exosomes in cancer. Mol Cancer.
18:522019. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sun Z, Shi K, Yang S, Liu J, Zhou Q, Wang
G, Song J, Li Z, Zhang Z and Yuan W: Effect of exosomal miRNA on
cancer biology and clinical applications. Mol Cancer. 17:1472018.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Morini M, Cangelosi D, Segalerba D,
Marimpietri D, Raggi F, Castellano A, Fruci D, de Mora JF, Canete
A, Yanez Y, et al: Exosomal microRNAs from longitudinal liquid
biopsies for the prediction of response to induction chemotherapy
in high-risk neuroblastoma patients: A proof of concept SIOPEN
Study. Cancers (Basel). 11:14762019. View Article : Google Scholar
|
|
18
|
Ma J, Xu M, Yin M, Hong J, Chen H, Gao Y,
Xie C, Shen N, Gu S and Mo X: Exosomal hsa-miR199a-3p promotes
proliferation and migration in neuroblastoma. Front Oncol.
9:4592019. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Fonseka P, Liem M, Ozcitti C, Adda CG, Ang
CS and Mathivanan S: Exosomes from N-Myc amplified neuroblastoma
cells induce migration and confer chemoresistance to non-N-Myc
amplified cells: Implications of intra-tumour heterogeneity. J
Extracell Vesicles. 8:15976142019. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Paul P, Qiao J, Kim KW, Romain C, Lee S,
Volny N, Mobley B, Correa H and Chung DH: Targeting
gastrin-releasing peptide suppresses neuroblastoma progression via
upregulation of PTEN signaling. PLoS One. 8:e725702013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Purushothaman A: Exosomes from cell
Culture-Conditioned medium: Isolation by ultracentrifugation and
characterization. Methods Mol Biol. 1952:233–244. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zafar A, Wang W, Liu G, Wang X, Xian W,
McKeon F, Foster J, Zhou J and Zhang R: Molecular targeting
therapies for neuroblastoma: Progress and challenges. Med Res Rev.
Nov 6–2020.(Epub ahead of print). doi: 10.1002/med.21750.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Haug BH, Hald ØH, Utnes P, Roth SA, LØkke
C, FlÆgstad T and Einvik C: Exosome-like extracellular vesicles
from MYCN-amplified neuroblastoma cells contain oncogenic miRNAs.
Anticancer Res. 35:2521–2530. 2015.PubMed/NCBI
|
|
25
|
Su T, Xiao Y, Xiao Y, Guo Q, Li C, Huang
Y, Deng Q, Wen J, Zhou F and Luo XH: Bone marrow mesenchymal stem
Cells-Derived Exosomal miR-29b-3p Regulates Aging-Associated
insulin resistance. ACS Nano. 13:2450–2462. 2019.PubMed/NCBI
|
|
26
|
Babuta M, Furi I, Bala S, Bukong TN, Lowe
P, Catalano D, Calenda C, Kodys K and Szabo G: Dysregulated
autophagy and lysosome function are linked to exosome production by
Micro-RNA 155 in alcoholic liver disease. Hepatology. 70:2123–2141.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Loven J, Zinin N, Wahlstrom T, Muller I,
Brodin P, Fredlund E, Ribacke U, Pivarcsi A, Pahlman S and
Henriksson M: MYCN-regulated microRNAs repress estrogen
receptor-alpha (ESR1) expression and neuronal differentiation in
human neuroblastoma. Proc Natl Acad Sci USA. 107:1553–1558. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Khuu C, Utheim TP and Sehic A: The three
paralogous MicroRNA clusters in development and disease, miR-17-92,
miR-106a-363, and miR-106b-25. Scientifica (Cairo).
2016:13796432016.PubMed/NCBI
|
|
29
|
Opel D, Poremba C, Simon T, Debatin KM and
Fulda S: Activation of Akt predicts poor outcome in neuroblastoma.
Cancer Res. 67:735–745. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hogarty MD and Maris JM: PI3King on MYCN
to improve neuroblastoma therapeutics. Cancer Cell. 21:145–147.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Li Z, Xie J, Fei Y, Gao P, Xie Q, Gao W
and Xu Z: GDNF family receptor alpha 2 promotes neuroblastoma cell
proliferation by interacting with PTEN. Biochem Biophys Res Commun.
510:339–344. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lee YR, Chen M and Pandolfi PP: The
functions and regulation of the PTEN tumour suppressor: New modes
and prospects. Nat Rev Mol Cell Biol. 19:547–562. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wu Q, Shang Y, Shen T, Liu F, Xu Y and
Wang H: Neuroprotection of miR-214 against isoflurane-induced
neurotoxicity involves the PTEN/PI3K/Akt pathway in human
neuroblastoma cell line SH-SY5Y. Arch Biochem Biophys.
678:1081812019. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Gerlinger M, Rowan AJ, Horswell S, Math M,
Larkin J, Endesfelder D, Gronroos E, Martinez P, Matthews N,
Stewart A, et al: Intratumor heterogeneity and branched evolution
revealed by multiregion sequencing. N Engl J Med. 366:883–892.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Barranha R, Costa JL, Carneiro F and
Machado JC: Genetic heterogeneity in colorectal cancer and its
clinical implications. Acta Med Port. 28:370–375. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Lu R, Zhao G, Yang Y, Jiang Z, Cai J,
Zhang Z and Hu H: Long noncoding RNA HOTAIRM1 inhibits cell
progression by regulating miR-17-5p/ PTEN axis in gastric cancer. J
Cell Biochem. 120:4952–4965. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Liang W and Yue Z: Lycium barbarum
polysaccharides promote osteoblasts viability by regulating
microRNA-17/PTEN. Life Sci. 225:72–78. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Wang X, Li Z, Bai J, Song W and Zhang F:
miR-17-5p regulates the proliferation and apoptosis of human
trabecular meshwork cells by targeting phosphatase and tensin
homolog. Mol Med Rep. 19:3132–3138. 2019.PubMed/NCBI
|