Introduction
Despite efforts to promote spinal cord injury (SCI)
repair (1), it is still regarded as
an incurable condition with limited treatment strategies (2). SCI is a disabling injury condition
that manifests as partial or complete loss of sensory and motor
function below the level of injury (3,4).
Moreover, >50% of SCIs result from trauma, including traffic
accidents, sports injuries and gun shots, whereas other factors,
such as inflammation, cancer and vascular diseases, are responsible
for non-traumatic SCI (5). SCI
results in a considerable burden on patients and treatment costs
are high (6). The pathophysiology
of SCI consists of both primary and secondary injury. Primary
injury refers to the mechanical injury to the spinal cord, which
includes central and peripheral nervous systems damage,
microvasculature damage, axonal disruption, spinal cord swelling,
spinal shock and hypotension (7).
Secondary injury consists of excitotoxicity, free radical-induced
lipid peroxidation, blood-brain barrier alterations, inflammation,
and the death of neurons and glial cells (8). Therefore, the development of improved
treatment strategies is important for patients with SCI.
Promoting axonal regeneration serves a pivotal role
in SCI treatment (9). A previous
study reported that astrocyte activation primarily mediates glial
scar formation following SCI, which prevents axonal regeneration
(10). Glial fibrillary acidic
protein (GFAP) is a cytokine secreted by activated astrocytes, thus
it is considered as a marker for astrocyte activation (11,12).
Growth-associated protein-43 (GAP-43, neuromodulin) participates in
the regulation of axonal growth (13). Neurotrophins, including nerve growth
factor (NGF) and brain-derived neurotrophic factor (BDNF), have
been shown to promote axonal regrowth and regeneration of injured
spinal cord (14). Therefore, GFAP,
GAP-43, NGF and BNDF are often assessed to evaluate axonal
regeneration.
Transcranial magnetic stimulation (TMS) is a medical
technology based on bioelectromagnetics, which was first introduced
by Barker et al (15). TMS
refers to the stimulation of the central nervous system,
particularly the brain, with a pulsed magnetic field (16). Due to its numerous advantages,
including its non-invasive nature, convenience and lack of pain,
interesting results have been observed for the use of TMS treatment
for paralysis, Parkinson's disease, epilepsy and depression
(17,18). Increasing evidence demonstrates that
TMS might serve as a potential neurophysiological technique for SCI
diagnosis and treatment, and clinical trials have reported that
high frequency repetitive TMS promotes motor rehabilitation and
modulates brainstem reflexes in individuals with SCI (19,20).
Embryonic, fetal neural and induced pluripotent stem
cells may be used to repair SCI; however, the clinical application
of stem cell therapies is limited by immunological rejection and
ethical concerns (21–23). Bone marrow mesenchymal stem cells
(BMSCs), which can be easily obtained from bone marrow, display the
potential to differentiate into cartilage cells, osteoblasts, fat
cells, myoblasts and nerve cells (24). BMSCs are widely used in cellular
transplantation due to their easy acquisition, low immunogenicity
and lack of ethical restraints (25). Additionally, a previous study
demonstrated that BMSCs could differentiate into neuronal-like
cells and promote SCI repair in mice (26). The Raf/MEK/ERK signaling pathway is
a signaling cascade involved in various physiological processes,
including cellular proliferation and differentiation (27). Raf/MEK/ERK signaling is also
involved in neuronal apoptosis in the hippocampus following
subarachnoid hemorrhage (28).
Previous studies revealed that Raf/MEK/ERK signaling was
upregulated in SCI model rats (29), and that the Raf inhibitor dabrafenib
displays potential to facilitate SCI recovery (30). Therefore, the present study aimed to
investigate the feasibility of allogenic BMSCs and Raf inhibition
for SCI treatment. In the present study, a rat model was
established to mimic SCI. Subsequently, rats were treated with
BMSCs, Raf inhibitor (RafI), TMS, TMS+BMSC or TMS+RafI. The effects
of monotherapy and combination therapy on functional recovery,
pathological alterations, neuronal apoptosis and the expression
levels of neuromodulin, neurotrophins, astrocyte markers and
Raf/MEK/ERK signaling were assessed.
Materials and methods
Animal model establishment and
treatment
Female Sprague Dawley rats (age, 8 weeks; weight,
220±20 g) were purchased from Liaoning Changsheng Biotechnology
Co., Ltd. Rats were housed in cages at 22±1°C with 12-h light/dark
cycles, 45–55% relative humidity, and ad libitum access to
food and water. Following adaptive feeding for one week, 84 rats
were randomly divided into the following 7 groups (12 rats per
group): i) Sham; ii) SCI; iii) SCI+BMSCs; iv) SCI+RafI; v) SCI+TMS;
vi) SCI+TMS+BMSCs; and vii) SCI+TMS+RafI. Rats were anesthetized by
the intraperitoneal injection of 50 mg/kg Nembutal. Subsequently,
the soft tissue around vertebral level T10 was removed to expose
the spinous processes and laminae of T10 and part of T9/T11. Then,
a 10-g impactor was used to induce spinal cord contusion injury
(100 g × cm × force) by the weight-drop method (31). Following SCI induction, the incision
was sutured layer by layer under sterilized conditions. Rats were
intraperitoneally injected with penicillin (40,000 U/day/rat) for 3
days post-surgery, and the bladder of each rat was extruded for
urination twice daily. In the sham group, incisions were directly
sutured without SCI induction, and the rats did not undergo manual
urination. Following SCI induction, rats received the following
treatment: i) Rats in the sham group were injected at the same site
with 5 µl DMEM/F12 medium (Gibco; Thermo Fisher Scientific, Inc.)
supplemented with 10% fetal calf serum (Hyclone; Cytiva) and 10 µl
normal saline on day 7 post-surgery, followed by 10 µl normal
saline daily from day 8–28 post-surgery; ii) rats in the SCI group
were injected into the SCI site with 5 µl DMEM/F12 medium
supplemented with 10% fetal calf serum and 10 µl normal saline on
day 7 post-surgery, followed by 10 µl normal saline daily from day
8–28 post-surgery; iii) rats in the SCI+BMSCs group were injected
into the SCI with 5 µl BMSC suspension (1×105 cells/µ1)
and 10 µl normal saline on day 7 post-surgery, followed by 10 µl
normal saline daily from day 8–28 post-surgery; iv) rats in the
SCI+RafI group were injected into the SCI site with 5 µl DMEM/F12
medium supplemented with 10% fetal calf serum and 10 µl PLX4720 (1
µg/µl; Aladdin) on day 7 post-surgery, followed by 10 µl PLX4720
daily from day 8–28 post-surgery; v) following stimulation with 10
Hz TMS at 24 h post-surgery, and then once a day, 5 days a week,
for 4 weeks, rats in the SCI+TMS group were injected into the SCI
site with 5 µl DMEM/F12 medium supplemented with 10% fetal calf
serum and 10 µl normal saline on day 7 post-surgery, followed by 10
µl normal saline daily from day 8–28 post-surgery; vi) rats in the
SCI+TMS+BMSCs group received the same TMS treatment as the SCI+TMS
group, and were injected into the SCI site with 5 µl BMSC
suspension (1×105 cells/µl) and 10 µl normal saline on
day 7 post-surgery, followed by 10 µl normal saline daily from day
8–28 post-surgery; vii) rats in the SCI+TMS+RafI group received the
same TMS treatment as the SCI+TMS group, and were injected into the
SCI site with 5 µl DMEM/F12 medium supplemented with 10% fetal calf
serum and 10 µl PLX4720 (1 µg/µ1) on day 7 post-surgery, followed
by 10 µl PLX4720 daily from day 8–28 post-surgery.
Rats were evaluated using the Basso, Beattie and
Bresnahan locomotor rating (BBB) scale before surgery and at day 1,
14 and 28 post-surgery (32,33). A
total of six live rats per group were used for BBB scoring and
subsequent experiments. Subsequently, rats were euthanized by the
intraperitoneal injection of 200 mg/kg Nembutal. A total of six
rats were used to assess hematoxylin and eosin (H&E) staining,
TUNEL and neuronal nuclei (NeuN) double staining as well as
immunofluorescence staining; another six rats were used to perform
western blotting. Spinal cord tissues were fixed with 4%
paraformaldehyde at room temperature for 48 h or stored at −70°C.
All animal experiments were performed according to Guide for the
Care and Use of Laboratory Animals 8th edition (34), and approved by the ethics committee
of Shengjing Hospital of China Medical University (approval no.
2020PS697K).
Isolation and identification of
BMSCs
Femurs and tibias were collected from each rat and
the soft tissues surrounding the femurs and tibias were removed.
The marrow cavity was repeatedly syringed with DMEM/F12 culture
medium to harvest bone marrow cells. Bone marrows cells were
resuspended and lysed using red cell lysis buffer (Beijing Solarbio
Science & Technology Co., Ltd.). Following washing twice with
PBS, cells were resuspended in DMEM/F12 culture medium supplemented
with 10% fetal calf serum. Then, cells (3×105/well) were
seeded into a 6-well plate. For identification, BMSCs were
collected at passage 3 and incubated with anti-CD11b (cat. no.
12-0110-80), anti-CD29 (cat. no. 11-0291-80), anti-CD90 (cat. no.
11-0900-81) and anti-CD45 (cat. no. 11-0461-80) monoclonal
antibodies (all eBioscience; Thermo Fisher Scientific, Inc.) at
37°C for 30 min. BMSCs were identified by flow cytometric detection
(35) using an Accuri C6 flow
cytometer (BD Biosciences) and Accuri C6 software (version
1.0.264.21; BD Biosciences).
H&E staining
Rat spinal cord tissues were embedded in paraffin
and cut into 5-µm thick sections. Tissue sections were stained
using a H&E staining kit (Wanleibio Co., Ltd.) according to the
manufacturer's instructions. Stained sections were visualized using
a fluorescence microscope (Olympus Corporation; magnification,
×400).
TUNEL and NeuN double staining
Paraffin-embedded spinal cord tissues were cut into
5-µm thick sections, de-paraffinized by heating to 60°C for 2 h
followed by xylene immersion, re-hydrated in a descending ethanol
series (95, 85, and 75%) and washed with PBS. Following heating in
a microwave (low power) with citrate buffer containing 0.0018
citric acid and 0.0082 mol/l sodium citrate for 10 min for antigen
revival, tissue sections were stained using the In Situ Cell Death
Detection Kit (Roche Diagnostics) according to the manufacturer's
protocol. After washing with PBS, the tissue sections were blocked
with 100% goat serum (Beijing Solarbio Science & Technology
Co., Ltd.) for 30 min at room temperature and then incubated with
an anti-NeuN mouse monoclonal primary antibody (cat. no. ab104224;
1:400; Abcam) overnight at 4°C. After washing with PBS, tissue
sections were incubated with a Cy3-labelled goat anti-mouse IgG
(H+L) secondary antibody (cat. no. A0521; 1:200; Beyotime Institute
of Biotechnology) for 1 h at room temperature. Tissue sections were
then stained with DAPI for 5 min at room temperature (Biosharp Life
Sciences). Following sealing with anti-fluorescence quenching
reagent (Beijing Solarbio Science & Technology Co., Ltd.),
stained tissue sections were observed using a fluorescence
microscope (Olympus Corporation; magnification, ×400) and three
random fields of view were observed for each section. The
quantification procedure was conducted by Image-pro plus software
(version 6.0; Media Cybernetics, Inc.).
Immunofluorescence staining
Paraffin-embedded spinal cord tissues were cut into
5-µm thick sections, de-paraffinized by heating to 60°C for 2 h
followed by xylene immersion, re-hydrated in a descending ethanol
series (95, 85 and 75%), washed with PBS, heated in a microwave
(low power) with citrate buffer for 10 min for antigen revival.
Following washing with PBS, tissue sections were blocked with 100%
goat serum for 15 min at room temperature. Subsequently, tissue
sections were incubated with anti-GFAP rabbit polyclonal (cat. no.
WL0836; 1:100; Wanleibio Co. Ltd.) or anti-GAP-43 rabbit monoclonal
(cat. no. ab75810; 1:200; Abcam) primary antibodies overnight at
4°C. Following primary antibody incubation, tissue sections were
incubated with Cy3-labelled goat anti-rabbit IgG (H+L) secondary
antibody (cat. no. A0516; 1:300; Beyotime Institute of
Biotechnology) for 1 h at room temperature. After staining with
DAPI for 5 min at room temperature, tissue sections were sealed
with anti-fluorescence quenching reagent and stained sections were
observed using a fluorescence microscope (Olympus Corporation;
magnification, ×400).
Western blotting
Total protein was extracted from rat spinal cord
tissues using the Whole Cell Lysis Assay Kit (Wanleibio Co., Ltd.)
and quantified using the BCA Protein Assay Kit (Wanleibio Co.,
Ltd.). Proteins (40 µg per well) were separated via 8–12% SDS-PAGE
and transferred onto PVDF membranes (EMD Millipore). After blocking
with 5% skimmed milk in TBST (0.15% Tween-20) at room temperature
for 1 h, the membranes were incubated overnight at 4°C with the
following primary antibodies: Rabbit polyclonal anti-NGF (cat. no.
A13922; 1:1,000; ABclonal Biotech Co., Ltd.), rabbit monoclonal
anti-BDNF (cat. no. BM4113; 1:200; Wuhan Boster Biological
Technology, Ltd.), rabbit monoclonal anti-C-Raf (cat. no. BM4108;
1:1,000; Wuhan Boster Biological Technology, Ltd.), rabbit
monoclonal anti-phosphorylated (p)-C-Raf (phospho ser259; cat. no.
Bsm-52194R; 1:1,000; BIOSS), rabbit polyclonal anti-MEK1/2 (cat.
no. WL02258; 1:500; Wanleibio Co. Ltd.), rabbit monoclonal
anti-p-MEK1/2 (phosphor ser221; cat. no. 2338; 1:2,000; Cell
Signaling Technology, Inc.), rabbit monoclonal anti-ERK1/2 (cat.
no. BM4326; 1:200; Wuhan Boster Biological Technology, Ltd.),
rabbit polyclonal anti-p-ERK1/2 (phospho thr202/tyr204; cat. no.
WLP1512; 1:500; Wanleibio Co. Ltd.) and rabbit polyclonal
anti-β-actin (cat. no. WL01845; 1:1,000; Wanleibio Co. Ltd.).
Following washing with TBST, the membranes were incubated with
HRP-conjugated goat anti-rabbit IgG secondary antibody (1:5,000;
cat. no. WLA023; Wanleibio Co. Ltd.) for 45 min at 37°C. Proteins
were visualized using ECL luminescent reagent (Wanleibio Co. Ltd.).
Protein expression levels were semi-quantified using
Gel-Pro-Analyzer (version 4.0; Media Cybernetics, Inc.) with
β-actin as the loading control.
Statistical analysis
Statistical analyses were performed using GraphPad
Prism 7 software (GraphPad Software, Inc.). Data are presented as
the mean ± SD (n≥6). BBB scores among the 7 groups were compared
using the Kruskal-Wallis test followed by Dunn's post hoc test.
Comparisons among multiple groups were analyzed using one-way ANOVA
followed by Tukey's post hoc test. P<0.05 was considered to
indicate a statistically significant difference.
Results
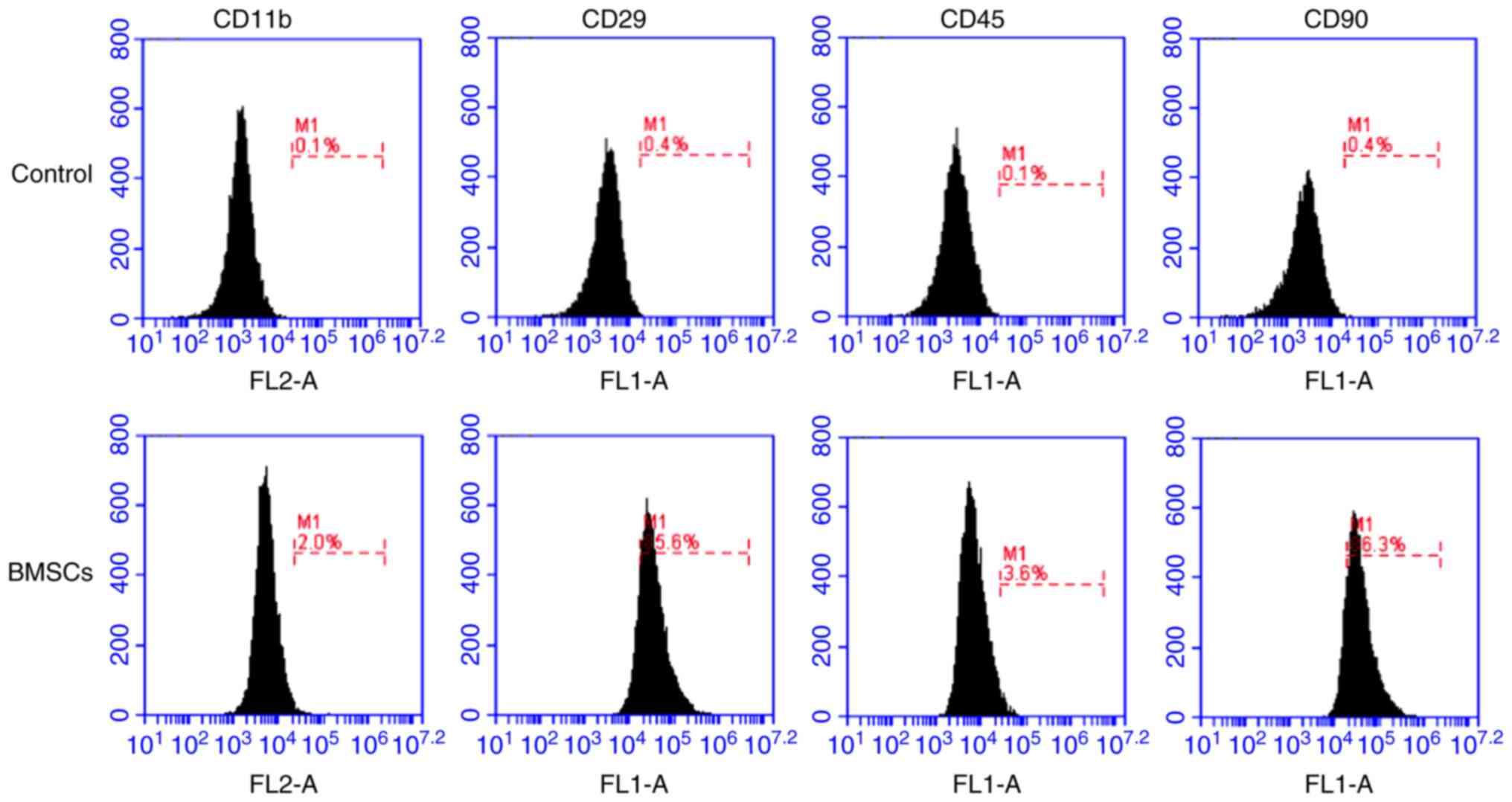
Successful isolation of BMSCs
BMSCs were isolated from rat bone marrow and
identified via flow cytometry at passage 3. The flow cytometry
results demonstrated that the isolated cells were
CD11b−, CD45−, CD29+ and
CD90+ (Fig. 1),
indicating that the isolated cells were BMSCs.
Combination treatment of TMS+BMSCs or
TMS+RafI further alleviates locomotor function, pathological
alterations and neuronal apoptosis in SCI model rats
To investigate the effect of TMS+BMSCs and TMS+RafI
combination therapies on SCI model rats, BBB scoring was employed
to evaluate functional recovery (Fig.
2A). Following monotherapy or combination therapy, the BBB
scores of SCI model rats at day 28 post-surgery were significantly
increased (SCI+BMSCs, 14.00; SCI+RafI, 14.33; SCI+TMS, 13.17;
SCI+TMS+BMSCs, 15.33; and SCI+TMS+RafI, 16.17) compared with
untreated SCI model rats (7.83), which suggested that all therapies
were beneficial for the recovery of locomotor function following
SCI. In addition, morphological alterations in the spinal cord were
detected by performing H&E staining. The spinal cord tissues of
the SCI group displayed a little congestion and obvious structural
disorders at the site of injury, whereas monotherapy slightly
alleviated the intensity of spinal cord lesions (Fig. 2B). TMS+BMSCs and TMS+RafI
combination therapies notably relieved SCI-induced structural
damage (Fig. 2B). Neuronal
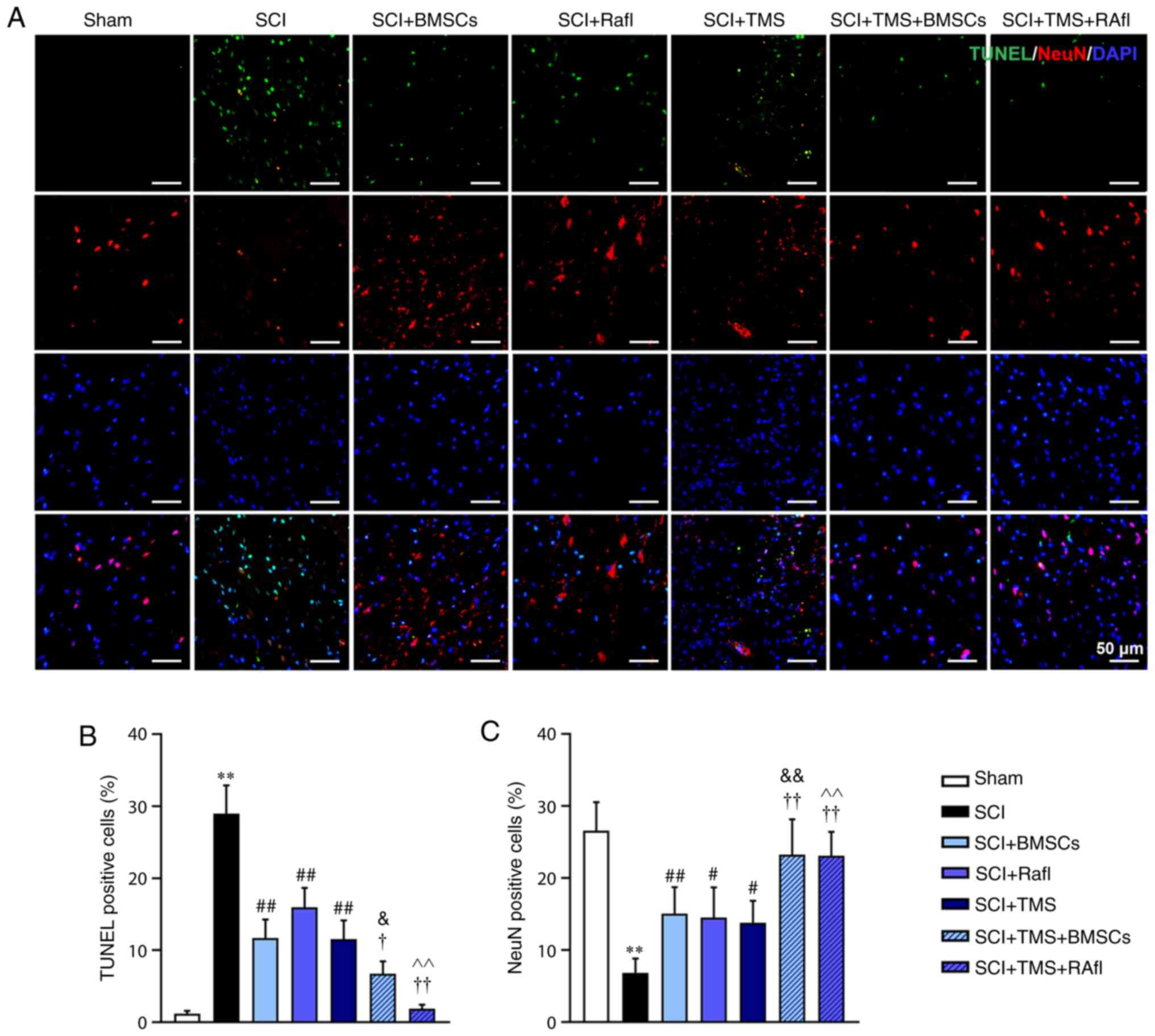
apoptosis was determined by conducting TUNEL-NeuN double staining.
The SCI group displayed a significantly increased number of
apoptotic cells (28.94%) and fewer NeuN+ cells (6.85%)
compared with the sham group (TUNEL+ cells, 1.19%;
NeuN+ cells, 26.58%), indicating that SCI enhanced
neuronal apoptosis (Fig. 3A-C).
However, compared with the SCI group, all therapies significantly
increased the number of NeuN+ cells (SCI+BMSCs, 15.09%;
SCI+RafI, 14.51%; SCI+TMS, 13.81%; SCI+TMS+BMSCs, 23.26%;
SCI+TMS+RafI, 23.10%) and decreased the number of TUNEL+
cells (SCI+BMSCs, 11.66%; SCI+RafI, 15.97%; SCI+TMS, 11.55%;
SCI+TMS+BMSCs, 6.75%; SCI+TMS+RafI, 1.88%), which was significantly
enhanced in the SCI+TMS+BMSCs and SCI+TMS+RafI groups compared with
the corresponding monotherapy groups (Fig. 3A-C). Collectively, the results
suggested that the combination therapies were more effective at
alleviating SCI-induced pathological alterations and neuronal
apoptosis in SCI model rats compared with monotherapy.
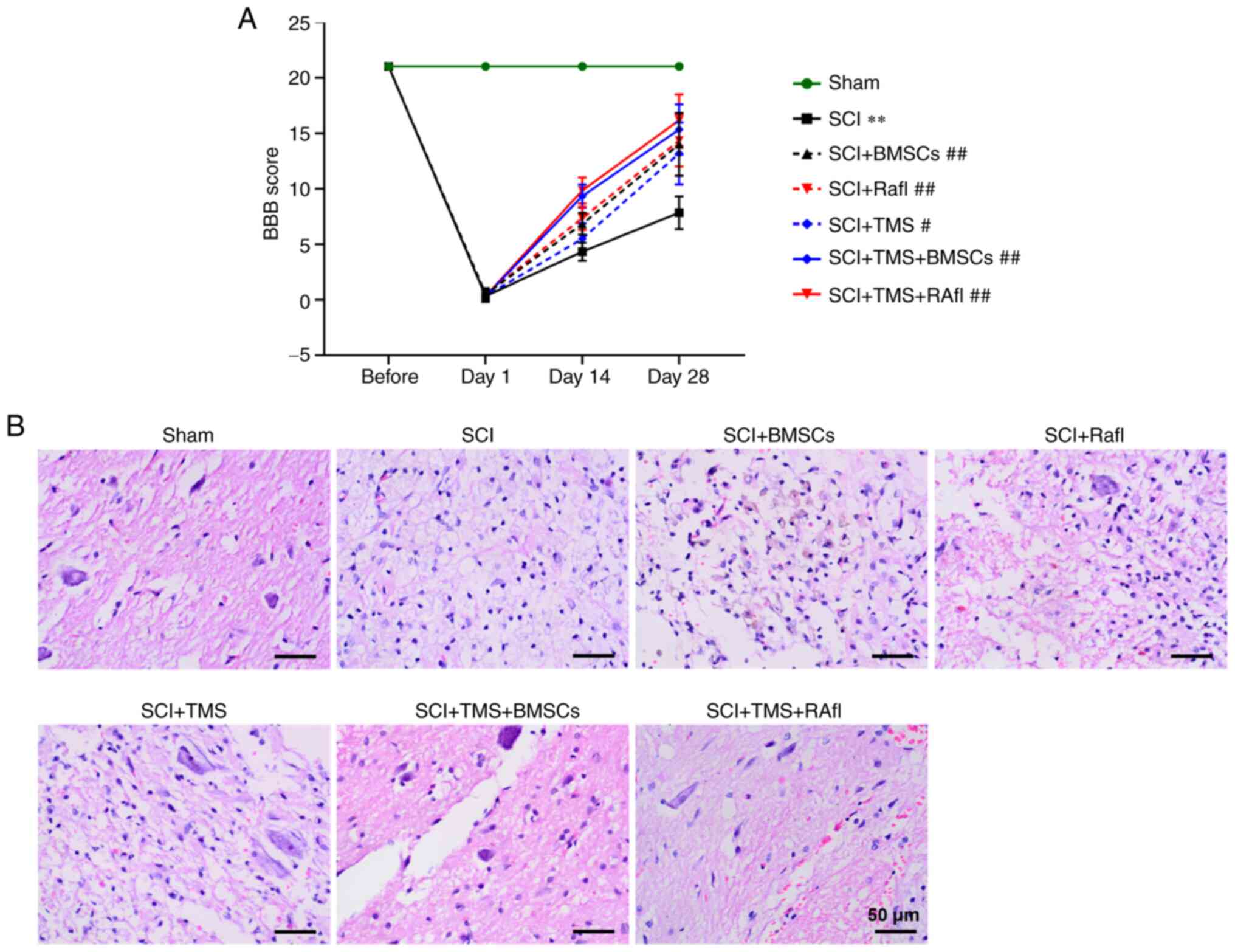 | Figure 2.Effects of combination therapy on
locomotor functional recovery and pathological alterations in SCI
model rats. (A) BBB scoring was used to evaluate the recovery of
locomotor function. (B) Hematoxylin and eosin staining was
performed to evaluate spinal injury (scale bar, 50 µm). Green
arrow, neurons; red arrow, congestion; yellow asterisk,
vacuolation. **P<0.01 vs. sham (day 28); #P<0.05
and ##P<0.01 vs. SCI (day 28). SCI, spinal cord
injury; BBB, Basso, Beattie and Bresnahan locomotor rating; TMS,
transcranial magnetic stimulation; BMSCs, bone marrow mesenchymal
stem cells; RafI, Raf inhibitor. |
Combination treatment of TMS+BMSCs or
TMS+RafI further increases the expression of neuromodulin and
neurotrophins, and decreases the expression of astrocyte markers in
SCI model rats
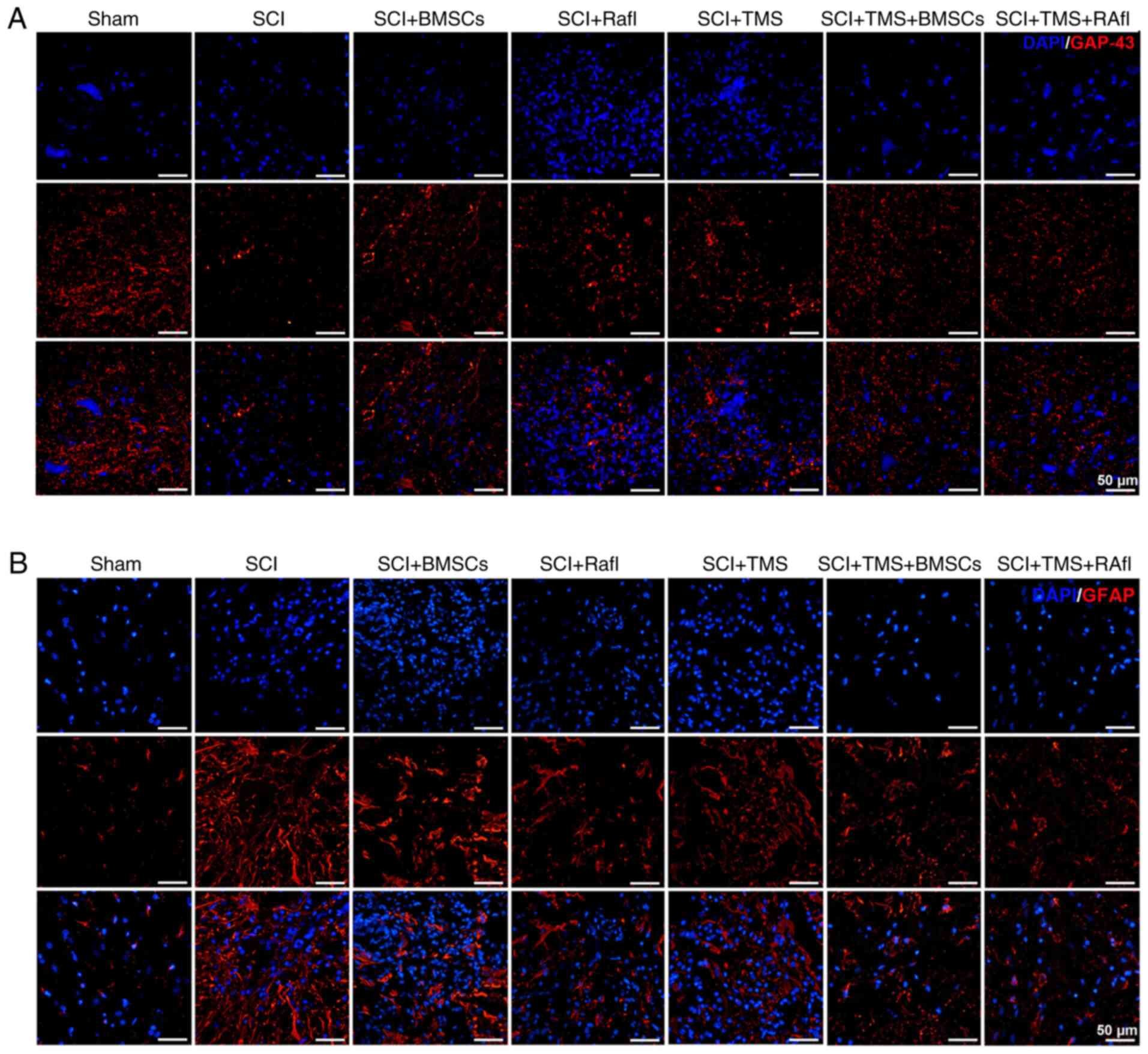
GAP-43 and GFAP expression levels were detected by
performing an immunofluorescence assay. GAP-43 expression levels
were notably lower in the SCI group compared with the sham group
(Fig. 4A). After monotherapy or
combination therapy, GAP-43 expression was markedly increased in
SCI model rats, which was more notable in the SCI+TMS+BMSCs and
SCI+TMS+RafI groups (Fig. 4A). By
contrast, the expression of GFAP was obviously upregulated in the
SCI group compared with the sham group, which was decreased
following monotherapy or combination therapy, where the reduction
was more remarkable in the RafI, SCI+TMS+BMSCs and SCI+TMS+RafI
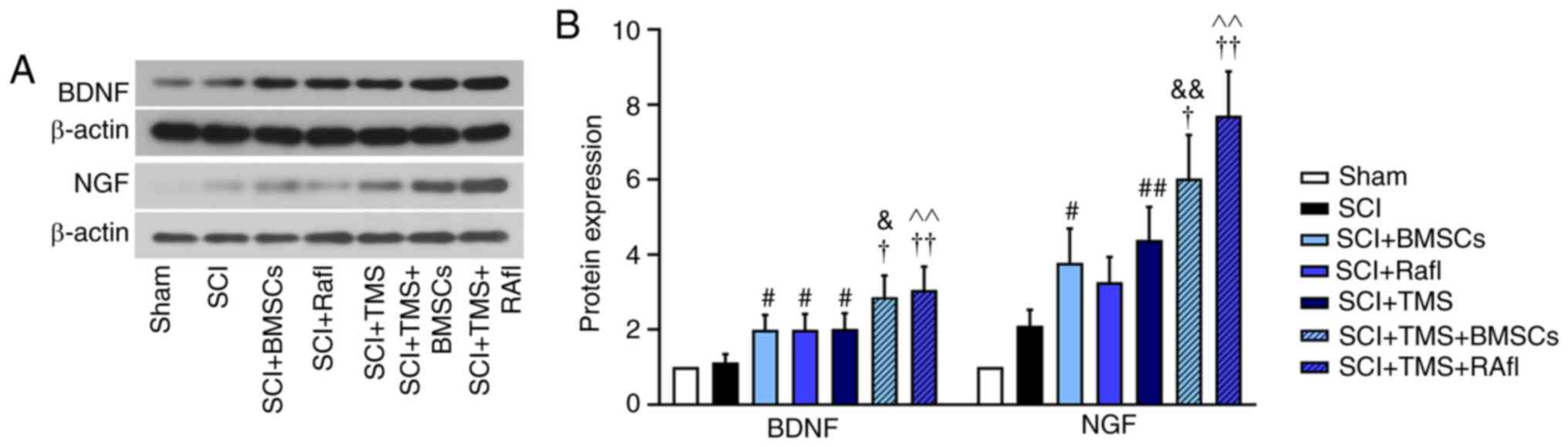
groups (Fig. 4B). Subsequently, the
relative expression levels of NGF and BDNF were detected via
western blotting (Fig. 5A and B).
With the exception of RafI monotherapy, all other monotherapies and
combination therapies significantly increased the relative
expression levels of NGF (SCI+BMSCs, 3.78; SCI+TMS, 4.4;
SCI+TMS+BMSCs, 6.02; SCI+TMS+RafI, 7.52) and BDNF (SCI+BMSCs, 2.00;
SCI+TMS, 2.02; SCI+TMS+BMSCs, 2.86; SCI+TMS+RafI, 3.05) in SCI
model rats compared with untreated SCI model rats (NGF, 2.10; and
BDNF, 1.12). Compared with the SCI group, RafI monotherapy
increased the relative expression of NGF (3.26) and significantly
upregulated BDNF (1.99) expression levels (Fig. 5A and B). Moreover, the increases in
NGF and BDNF expression levels were significantly enhanced in the
combination therapy groups compared with the corresponding
monotherapy groups (Fig. 5A and
B).
Combination treatment of TMS+BMSCs or
TMS+RafI further inhibits Raf/MEK/ERK signaling in SCI model
rats
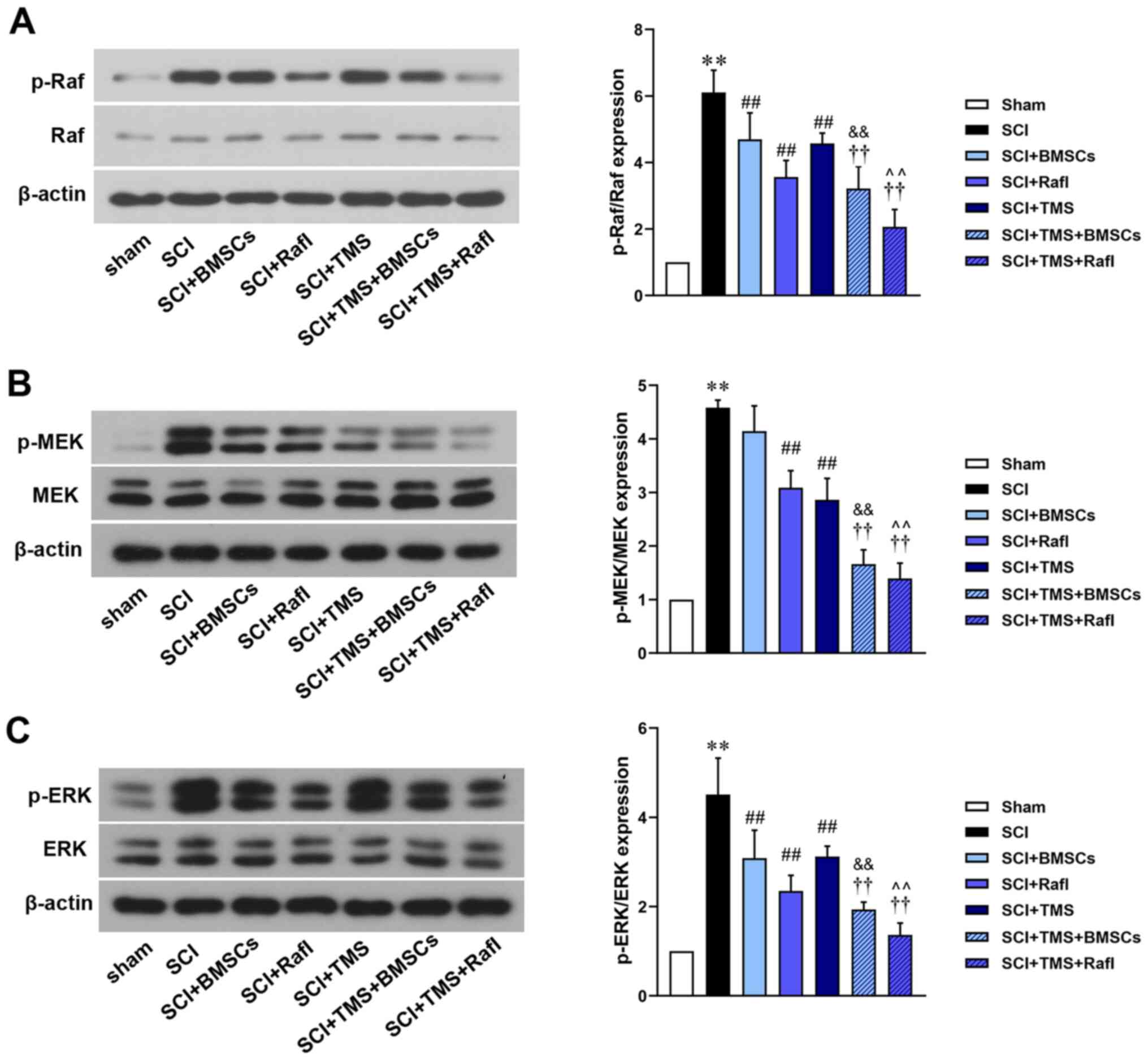
The relative expression levels of Raf, p-Raf, MEK,
p-MEK, ERK and p-ERK were detected (Fig. 6). Compared with the SCI group
(p-Raf/Raf, 6.11; p-ERK/ERK, 4.51), all therapies significantly
reduced the relative expression levels of p-Raf/Raf (SCI+BMSCs,
4.69; SCI+RafI, 3.56; SCI+TMS, 4.57; SCI+TMS+BMSCs, 3.22;
SCI+TMS+RafI, 2.07) and p-ERK/ERK (SCI+BMSCs, 3.08; SCI+RafI, 2.35;
SCI+TMS, 3.12; SCI+TMS+BMSCs, 1.93; SCI+TMS+RafI, 1.36). With the
exception of BMSC monotherapy (4.14), all other therapies
significantly reduced the relative expression levels of p-MEK/MEK
(SCI+RafI, 3.09; SCI+TMS, 2.86; SCI+TMS+BMSCs, 1.66; SCI+TMS+RafI,
1.39) compared with the SCI group (4.58). BMSC monotherapy
downregulated the relative expression levels of p-MEK/MEK compared
with the SCI group. The downregulation of p-Raf/Raf, p-MEK/MEK and
p-ERK/ERK expression levels was significantly enhanced in the
combination treatment groups compared with the corresponding
monotherapy groups. No notable alterations in the relative
expression levels of Raf, MEK and ERK were observed among the
treatment groups.
Discussion
SCI commonly results from accidents and acts of
violence, and patients with SCI experience a series of
complications, including cerebrovascular damage, cervical spine
injury and brain injury (36,37).
At present, there is no satisfactory treatment for SCI (2). Since its discovery in 1985, TMS has
been widely used to treat a diverse range of neurodegenerative
disorders (15). Previous studies
have suggested that TMS might serve as a promising therapeutic
strategy for SCI (38,39). BMSCs, which do not result in
immunological rejection, are appropriate for the treatment of SCI
(40). Emerging evidence has also
demonstrated that BMSCs display the potential to regenerate neural
function (41), and a previous
study reported that BMSCs and TMS exerted synergistic effects in
vascular dementia model rats (42).
Recovery of the memory and learning ability of rats with vascular
dementia was more significant following the combined application of
TMS and BMSCs compared with either therapy alone, indicating that
the combination of TMS and BMSCs may be more effective for the
treatment of neurological diseases (42). However, whether the combined use of
TMS and BMSCs is more effective compared with either therapy alone
in treating SCI is not completely understood. Raf inhibition has
also been reported as a potential therapeutic strategy for SCI
(30), but whether the combination
of TMS and Raf inhibition displays synergistic effects on SCI
recovery requires further investigation. Therefore, the present
study aimed to compare the effects of TMS, BMSC transplantation,
RafI, TMS+BMSCs and TMS+RafI treatment on SCI in vivo.
SCI can result in long-term motor deficits, thus the
recovery of motor dysfunction is vital for patients with SCI
(43). The BBB scale is commonly
used to evaluate motor function deficit and recovery from various
injuries (44). Furthermore, TMS
was reported to facilitate motor recovery in rats with T10-11
vertebrae injuries, as evidenced by a higher BBB score in the
TMS-treated group (45). Moreover,
BMSC transplantation treatment displayed a significant effect on
motor function recovery (46),
which was consistent with the results of the present study. In the
present study, the BBB scores of SCI model rats receiving TMS, BMSC
or RafI treatment alone were significantly higher compared with
untreated SCI model rats. Although the combined application of
TMS+BMSCs or TMS+RafI did not further improve motor recovery in SCI
model rats, the two combined treatments displayed no negative
effects on motor recovery. In the present study, two key
limitations were that the time interval for BBB scoring was
potentially too long and there was a lack of additional time
points. Moreover, histopathological lesions of the spinal cord were
also attenuated by monotherapy and combination therapy to varying
degrees.
Apoptosis, programmed cell death, is the process of
cellular self-destruction (47).
Apoptosis is implicated in secondary spinal cord injury, further
influencing neural damage and functional deficits (48). In the present study, all therapies
inhibited SCI-induced neuronal apoptosis, as evidenced by the
decreased number of TUNEL+ cells and the increased
number of NeuN+ cells in the treatment groups compared
with untreated SCI model rats. Notably, the combined application of
TMS+BMSCs or TMS+RafI significantly decreased the number of
TUNEL+ cells and significantly increased the number of
NeuN+ cells compared with the corresponding monotherapy
groups, indicating that combination treatment displayed synergistic
effects on the attenuation of neuronal apoptosis at the site of
SCI. Apoptosis is one of the key determining factors of the extent
of neuronal loss post-SCI (49),
thus blocking neuronal apoptosis may improve functional recovery
(50–52). Therefore, the attenuated functional
deficits induced by the therapies investigated in the present study
may be partly attributed to inhibition of neuronal apoptosis.
GAP-43 is primarily expressed in developing and
regenerating neurons, which is associated with neuronal growth
(53). The astrocyte marker GFAP is
involved in astroglial development and differentiation (54). In the present study, compared with
untreated SCI model rats, all therapies markedly upregulated the
expression of GAP-43 and notably downregulated the expression of
GFAP in SCI model rats, particularly RafI, TMS+BMSCs and TMS+RafI.
NGF and BDNF are neurotrophic growth factors that are critical for
the development and survival of neurons (55). The results of the present study
demonstrated that all therapies, excluding RafI monotherapy,
significantly upregulated NGF and BDNF expression levels in SCI
model rats compared with untreated SCI model rats. Moreover, the
combination of TMS+BMSCs or TMS+RafI displayed an enhanced ability
to upregulate NGF and BDNF expression levels compared with the
corresponding monotherapy groups. The results suggested that
combination therapies might promote neuronal growth, development
and survival, and inhibit SCI-induced astroglial activation more
effectively compared with monotherapies.
Emerging evidence has indicated that Raf/MEK/ERK
signaling might promote the neural cell apoptosis (56). In a previous study, p-Raf/Raf,
p-MEK/MEK and p-ERK/ERK expression levels were upregulated in SCI
model rats, indicating that Raf/MEK/ERK signaling is involved in
the development of SCI (29). C-Raf
overexpression displayed an inhibitory effect on the development
and maturation of spinal cord neurons (28,29).
In the present study, treatment with TMS, BMSCs or RafI alone
inhibited SCI-induced activation of the Raf/MEK/ERK signaling
pathway. Furthermore, the combined application of TMS+BMSCs or
TMS+RafI displayed a significantly enhanced potential to inhibit
SCI-induced activation of Raf/MEK/ERK signaling compared with the
corresponding monotherapy groups. Due to the pivotal role of
Raf/MEK/ERK signaling in SCI, it was hypothesized that combination
therapy might display an improved therapeutic effect on SCI by
further suppressing Raf/MEK/ERK signaling.
In conclusion, the results of the present study
demonstrated that the therapeutic effect of TMS combined with BMSC
transplantation or RafI on SCI recovery in rats was more efficient
compared with monotherapy; therefore, combination therapy may serve
as a novel therapeutic strategy for SCI.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
LZ conceptualized and designed the study. SF
performed the histological examination of the spinal cord and
immunofluorescence staining and revised the manuscript. SW
performed western blotting. SF and SW established the animal model
of SCI and performed all treatments. SS and HS analyzed the data.
HS drafted the manuscript. LZ and SF confirm the authenticity of
all the raw data. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
All animal experiments were performed according to
Guide for the Care and Use of Laboratory Animals 8th edition, and
approved by the ethics committee of Shengjing Hospital of China
Medical University (approval no. 2020PS697K).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
SCI
|
spinal cord injury
|
|
TMS
|
transcranial magnetic stimulation
|
|
BMSCs
|
bone marrow mesenchymal stem cells
|
|
NeuN
|
neuronal nuclei
|
|
GAP-43
|
growth-associated protein 43
|
|
NGF
|
nerve growth factor
|
|
BDNF
|
brain derived neurotrophic factor
|
|
GFAP
|
glial fibrillary acidic protein
|
|
TUNEL
|
TdT-mediated dUTP Nick-End
Labeling
|
|
BBB scale
|
Basso, Beattie and Bresnahan locomotor
rating scale
|
References
|
1
|
Kim YH, Ha KY and Kim SI: Spinal cord
injury and related clinical trials. Clin Orthop Surg. 9:1–9. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wang YT, Lu XM, Chen KT, Shu YH and Qiu
CH: Immunotherapy strategies for spinal cord injury. Curr Pharm
Biotechnol. 16:492–505. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Manchikanti L, Singh V, Datta S, Cohen SP
and Hirsch JA; American Society of Interventional Pain Physicians,
: Comprehensive review of epidemiology, scope, and impact of spinal
pain. Pain Physician. 12:E35–E70. 2009.PubMed/NCBI
|
|
4
|
Kwon BK, Tetzlaff W, Grauer JN, Beiner J
and Vaccaro AR: Pathophysiology and pharmacologic treatment of
acute spinal cord injury. Spine J. 4:451–464. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Spinal Cord Injury (SCI) 2016 facts and
figures at a glance. J Spinal Cord Med. 39:493–494. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ditunno JF Jr: Functional outcomes in
spinal cord injury (SCI): Quality care versus cost containment. J
Spinal Cord Med. 20:1–7. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
McDonald JW and Sadowsky C: Spinal-cord
injury. Lancet. 359:417–425. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Rowland JW, Hawryluk GW, Kwon B and
Fehlings MG: Current status of acute spinal cord injury
pathophysiology and emerging therapies: Promise on the horizon.
Neurosurg Focus. 25:E22008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ren Z, Chen X, Yang J, Kress BT, Tong J,
Liu H, Takano T, Zhao Y and Nedergaard M: Improved axonal
regeneration after spinal cord injury in mice with conditional
deletion of ephrin B2 under the GFAP promoter. Neuroscience.
241:89–99. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yang T, Dai Y, Chen G and Cui S:
Dissecting the dual role of the glial scar and scar-forming
astrocytes in spinal cord injury. Front Cell Neurosci. 14:782020.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Benninger F, Glat MJ, Offen D and Steiner
I: Glial fibrillary acidic protein as a marker of astrocytic
activation in the cerebrospinal fluid of patients with amyotrophic
lateral sclerosis. J Clin Neurosci. 26:75–78. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Okada S, Hara M, Kobayakawa K, Matsumoto Y
and Nakashima Y: Astrocyte reactivity and astrogliosis after spinal
cord injury. Neurosci Res. 126:39–43. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Curtis R, Green D, Lindsay RM and Wilkin
GP: Up-regulation of GAP-43 and growth of axons in rat spinal cord
after compression injury. J Neurocytol. 22:51–64. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Keefe KM, Sheikh IS and Smith GM:
Targeting neurotrophins to specific populations of neurons: NGF,
BDNF, and NT-3 and their relevance for treatment of spinal cord
injury. Int J Mol Sci. 18:5482017. View Article : Google Scholar
|
|
15
|
Barker AT, Jalinous R and Freeston IL:
Non-invasive magnetic stimulation of human motor cortex. Lancet.
1:1106–1107. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Pashut T, Wolfus S, Friedman A, Lavidor M,
Bar-Gad I, Yeshurun Y and Korngreen A: Mechanisms of magnetic
stimulation of central nervous system neurons. PLoS Comput Biol.
7:e10020222011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Rothwell JC: Techniques and mechanisms of
action of transcranial stimulation of the human motor cortex. J
Neurosci Methods. 74:113–122. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Cantello R: Applications of transcranial
magnetic stimulation in movement disorders. J Clin Neurophysiol.
19:272–293. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
de Araujo AVL, Barbosa VRN, Galdino GS,
Fregni F, Massetti T, Fontes SL, de Oliveira Silva D, da Silva TD,
Monteiro CBM, Tonks J and Magalhães FH: Effects of high-frequency
transcranial magnetic stimulation on functional performance in
individuals with incomplete spinal cord injury: Study protocol for
a randomized controlled trial. Trials. 18:5222017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kumru H, Kofler M, Valls-Sole J and Vidal
J: Brainstem reflex excitability after high-frequency repetitive
transcranial magnetic stimulation in healthy and spinal cord injury
subjects. Brain Res Bull. 147:86–91. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Aznar J and Sanchez JL: Embryonic stem
cells: Are useful in clinic treatments? J Physiol Biochem.
67:141–144. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ogawa Y, Sawamoto K, Miyata T, Miyao S,
Watanabe M, Nakamura M, Bregman BS, Koike M, Uchiyama Y, Toyama Y
and Okano H: Transplantation of in vitro-expanded fetal neural
progenitor cells results in neurogenesis and functional recovery
after spinal cord contusion injury in adult rats. J Neurosci Res.
69:925–933. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Tsuji O, Miura K, Okada Y, Fujiyoshi K,
Mukaino M, Nagoshi N, Kitamura K, Kumagai G, Nishino M, Tomisato S,
et al: Therapeutic potential of appropriately evaluated
safe-induced pluripotent stem cells for spinal cord injury. Proc
Natl Acad Sci USA. 107:12704–12709. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Miao C, Lei M, Hu W, Han S and Wang Q: A
brief review: The therapeutic potential of bone marrow mesenchymal
stem cells in myocardial infarction. Stem Cell Res Ther. 8:2422017.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Fehlings MG and Vawda R: Cellular
treatments for spinal cord injury: The time is right for clinical
trials. Neurotherapeutics. 8:704–720. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Mannoji C, Koda M, Kamiya K, Dezawa M,
Hashimoto M, Furuya T, Okawa A, Takahashi K and Yamazaki M:
Transplantation of human bone marrow stromal cell-derived
neuroregenrative cells promotes functional recovery after spinal
cord injury in mice. Acta Neurobiol Exp (Wars). 74:479–488.
2014.PubMed/NCBI
|
|
27
|
Roberts PJ and Der CJ: Targeting the
Raf-MEK-ERK mitogen-activated protein kinase cascade for the
treatment of cancer. Oncogene. 26:3291–3310. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Cao FJ, Zhang X, Liu T, Li XW, Malik M and
Feng SQ: Up-regulation of Ras/Raf/ERK1/2 signaling in the spinal
cord impairs neural cell migration, neurogenesis, synapse
formation, and dendritic spine development. Chin Med J (Engl).
126:3879–3885. 2013.PubMed/NCBI
|
|
29
|
Liu T, Cao FJ, Xu DD, Xu YQ and Feng SQ:
Upregulated Ras/Raf/ERK1/2 signaling pathway: A new hope in the
repair of spinal cord injury. Neural Regen Res. 10:792–796. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Sugaya T, Kanno H, Matsuda M, Handa K,
Tateda S, Murakami T, Ozawa H and Itoi E: B-RAF(V600E) inhibitor
dabrafenib attenuates RIPK3-mediated necroptosis and promotes
functional recovery after spinal cord injury. Cells. 8:15822019.
View Article : Google Scholar
|
|
31
|
Woller SA, Malik JS, Aceves M and Hook MA:
Morphine self-administration following spinal cord injury. J
Neurotrauma. 31:1570–1583. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Basso DM, Beattie MS and Bresnahan JC:
Graded histological and locomotor outcomes after spinal cord
contusion using the NYU weight-drop device versus transection. Exp
Neurol. 139:244–256. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Sandhir R, Gregory E, He YY and Berman NE:
Upregulation of inflammatory mediators in a model of chronic pain
after spinal cord injury. Neurochem Res. 36:856–862. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
National Research Council (US) Committee
for the Update of the Guide for the Care and Use of Laboratory
Animals, . Guide for the Care and Use of Laboratory Animals, 8th
edition. National Academies Press (US); Washington, DC: 2011
|
|
35
|
Chen Q, Duan X, Xu M, Fan H, Dong Y, Wu H,
Zhang M, Liu Y, Nan Z, Deng S and Liu X: BMSC-EVs regulate Th17
cell differentiation in UC via H3K27me3. Mol Immunol. 118:191–200.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Eckert MJ and Martin MJ: Trauma: Spinal
cord injury. Surg Clin North Am. 97:1031–1045. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Lee BB, Cripps RA, Fitzharris M and Wing
PC: The global map for traumatic spinal cord injury epidemiology:
Update 2011, global incidence rate. Spinal Cord. 52:110–116. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Awad BI, Carmody MA, Zhang X, Lin VW and
Steinmetz MP: Transcranial magnetic stimulation after spinal cord
injury. World Neurosurg. 83:232–235. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Cha HG, Ji SG and Kim MK: Effect of
high-frequency repetitive transcranial magnetic stimulation on
motor cortical excitability and sensory nerve conduction velocity
in subacute-stage incomplete spinal cord injury patients. J Phys
Ther Sci. 28:2002–2004. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Lin L, Lin H, Bai S, Zheng L and Zhang X:
Bone marrow mesenchymal stem cells (BMSCs) improved functional
recovery of spinal cord injury partly by promoting axonal
regeneration. Neurochem Int. 115:80–84. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Wu S, Suzuki Y, Ejiri Y, Noda T, Bai H,
Kitada M, Kataoka K, Ohta M, Chou H and Ide C: Bone marrow stromal
cells enhance differentiation of cocultured neurosphere cells and
promote regeneration of injured spinal cord. J Neurosci Res.
72:343–351. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Wang F, Zhang C, Hou S and Geng X:
Synergistic effects of mesenchymal stem cell transplantation and
repetitive transcranial magnetic stimulation on promoting autophagy
and synaptic plasticity in vascular dementia. J Gerontol A Biol Sci
Med Sci. 74:1341–1350. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Anderson KD, Abdul M and Steward O:
Quantitative assessment of deficits and recovery of forelimb motor
function after cervical spinal cord injury in mice. Exp Neurol.
190:184–191. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Basso DM, Beattie MS and Bresnahan JC: A
sensitive and reliable locomotor rating scale for open field
testing in rats. J Neurotrauma. 12:1–21. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Poirrier AL, Nyssen Y, Scholtes F, Multon
S, Rinkin C, Weber G, Bouhy D, Brook G, Franzen R and Schoenen J:
Repetitive transcranial magnetic stimulation improves open field
locomotor recovery after low but not high thoracic spinal cord
compression-injury in adult rats. J Neurosci Res. 75:253–261. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Ning GZ, Song WY, Xu H, Zhu RS, Wu QL, Wu
Y, Zhu SB, Li JQ, Wang M, Qu ZG and Feng SQ: Bone marrow
mesenchymal stem cells stimulated with low-intensity pulsed
ultrasound: Better choice of transplantation treatment for spinal
cord injury: Treatment for SCI by LIPUS-BMSCs transplantation. CNS
Neurosci Ther. 25:496–508. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Elmore S: Apoptosis: A review of
programmed cell death. Toxicol Pathol. 35:495–516. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Mazarakis ND, Edwards AD and Mehmet H:
Apoptosis in neural development and disease. Arch Dis Child Fetal
Neonatal Ed. 77:F165–F170. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Kim DH, Vaccaro AR, Henderson FC and
Benzel EC: Molecular biology of cervical myelopathy and spinal cord
injury: Role of oligodendrocyte apoptosis. Spine J. 3:510–519.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Lan WB, Lin JH, Chen XW, Wu CY, Zhong GX,
Zhang LQ, Lin WP, Liu WN, Li X and Lin JL: Overexpressing
neuroglobin improves functional recovery by inhibiting neuronal
apoptosis after spinal cord injury. Brain Res. 1562:100–108. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Zhang H, Wu F, Kong X, Yang J, Chen H,
Deng L, Cheng Y, Ye L, Zhu S, Zhang X, et al: Nerve growth factor
improves functional recovery by inhibiting endoplasmic reticulum
stress-induced neuronal apoptosis in rats with spinal cord injury.
J Transl Med. 12:1302014. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Rong W, Wang J, Liu X, Jiang L, Wei F, Hu
X, Han X and Liu Z: Naringin treatment improves functional recovery
by increasing BDNF and VEGF expression, inhibiting neuronal
apoptosis after spinal cord injury. Neurochem Res. 37:1615–1623.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Madsen JR, MacDonald P, Irwin N, Goldberg
DE, Yao GL, Meiri KF, Rimm IJ, Stieg PE and Benowitz LI: Tacrolimus
(FK506) increases neuronal expression of GAP-43 and improves
functional recovery after spinal cord injury in rats. Exp Neurol.
154:673–683. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Brenner M, Kisseberth WC, Su Y, Besnard F
and Messing A: GFAP promoter directs astrocyte-specific expression
in transgenic mice. J Neurosci. 14:1030–1037. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Blais M, Levesque P, Bellenfant S and
Berthod F: Nerve growth factor, brain-derived neurotrophic factor,
neurotrophin-3 and glial-derived neurotrophic factor enhance
angiogenesis in a tissue-engineered in vitro model. Tissue Eng Part
A. 19:1655–1664. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Feng D, Wang B, Ma Y, Shi W, Tao K, Zeng
W, Cai Q, Zhang Z and Qin H: The Ras/Raf/erk pathway mediates the
subarachnoid hemorrhage-induced apoptosis of hippocampal neurons
through phosphorylation of p53. Mol Neurobiol. 53:5737–5748. 2016.
View Article : Google Scholar : PubMed/NCBI
|