Introduction
Diabetic nephropathy (DN) is a serious complication
of diabetes, and is the most common cause of end-stage renal
disease in developed countries (1).
It was estimated that the total number of individuals with diabetic
mellitus worldwide will reach 693 million in 2045 (2). In addition, ~20–40% of all diabetic
patients will develop DN (3). High
glucose (HG) and HG-induced oxidative stress, inflammation and
hemodynamic changes serve an important role in glomerular injury,
which is a hallmark of DN (4,5). In
diabetes, HG affects all cells in the kidneys, including
endothelial cells, renal tubular epithelial cells, renal
interstitial fibroblasts, podocytes and mesangial cells (6–8).
Mesangial cells are specialized smooth muscle cells between
capillary loops of the glomerular capillary, and are involved in
the physiological and pathological changes of glomera function
(9). Mesangial cell injury,
characterized by an increase in apoptosis, excessive inflammatory
cytokine production and extracellular matrix synthesis, is a basic
pathological change of DN (7,9).
Clinical, animal and in vitro studies have revealed that HG
can induce mesangial cell apoptosis, which aggravates the
pathological process of DN (10,11).
The hyperglycemia-induced excessive generation of reactive oxygen
species (ROS) has been recognized as one of the causes for DN
(10–12); however, the precise mechanisms
involved are yet to be fully elucidated.
p66Src homology/collagen (Shc) is a
member of the ShcA protein family and acts as a response protein
that modulates the response to oxidative stress (13). p66Shc is mainly located
in the cytoplasm; however, when it is activated under stress
conditions, it enters the mitochondria and acts with cytochrome
c to produce ROS (14).
Moreover, p66Shc is known to serve a major role in
various kidney diseases, such as drug-induced acute kidney injury
(15), hypertension-induced
nephropathy (16) and DN (17). p66Shc knockout has been
reported to protect mesangial cells from HG-induced apoptosis
(17), as well as protect against
renal tubular injury (18) and
injury to podocytes (19).
Therefore, treatments targeted at p66Shc may be
beneficial for DN.
Piperazine ferulate (PF;
C4H10N2·2C10H10O4;
474.51 g/mol; Fig. 1A), a compound
synthesized by ferulic acid and piperazine, is used in the
treatment of various types of kidney disease including DN,
nephritis (20) and immunoglobulin
(Ig)A nephropathy (21) in China.
Previous research has revealed that PF reduces the levels of blood
urea nitrogen and serum creatinine in rats subjected to 5/6
nephrectomy (22) and exerts
anti-hypertensive effects via the activation of endothelial nitric
oxide synthase (eNOS) (23).
Another previous study reported that PF administration can restore
the HG-induced expression of eNOS in glomerular endothelial cells
and delay the development of DN (24). However, it remains unknown as to
whether PF restores mesangial cell injury under hyperglycemic
conditions by inhibiting p66Shc.
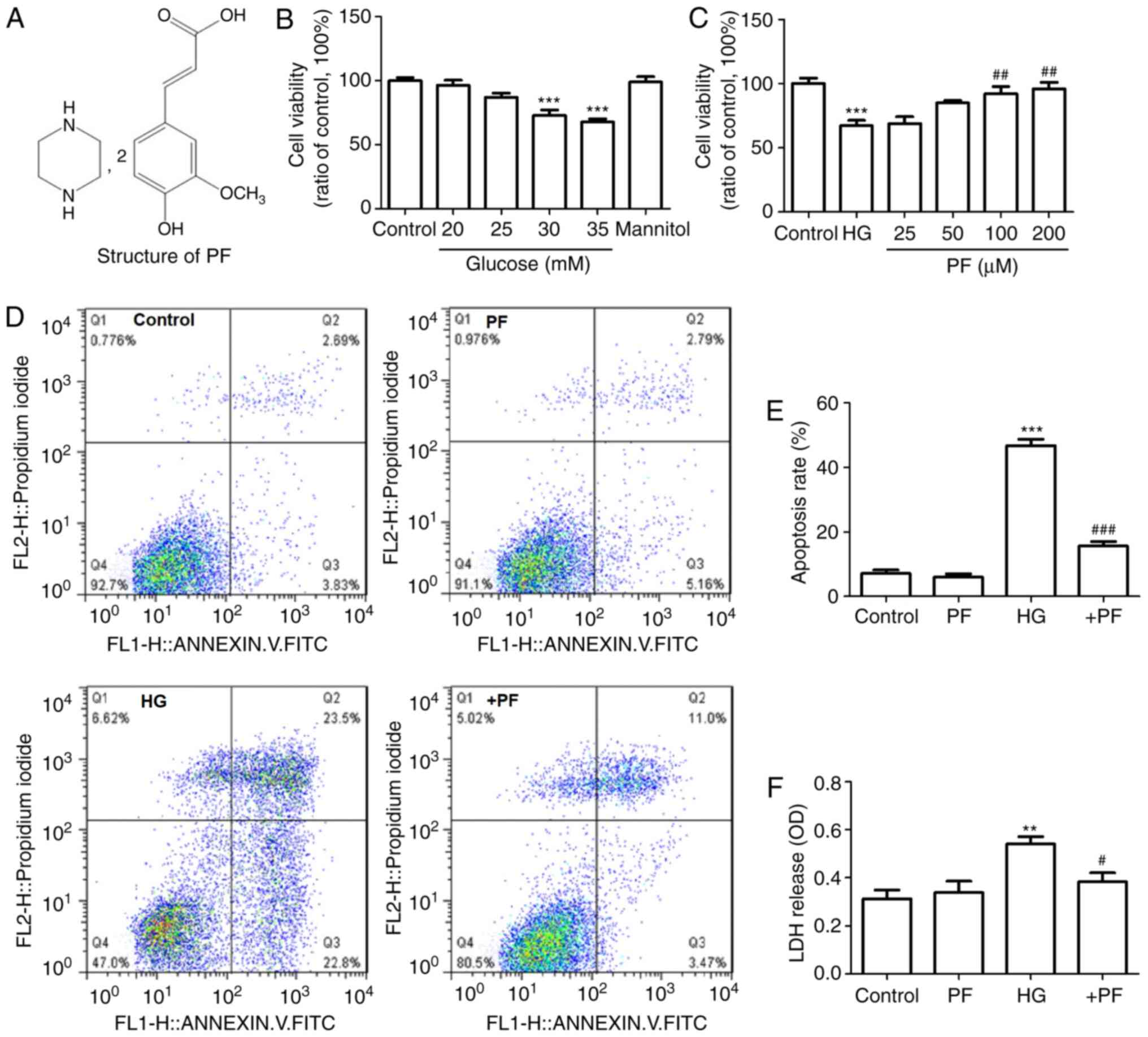 | Figure 1.PF prevents the mesangial cell injury
induce by HG. (A) Structure of PF. (B) Effect of various
concentration of HG on cell viability. (C) Effect of various
concentration of PF on HG-induced cell damage (HG, 30 mM). (D)
Representative flow cytometric results. (E) Statistical analysis of
the flow cytometry data. (F) Level of LDH released in the
supernatant of cultured cells. Data are expressed as the mean ±
SEM, n=3. **P<0.01, ***P<0.001 vs. control group;
#P<0.05, ##P<0.01,
###P<0.001 vs. HG group. PF, piperazine ferulate; HG,
high glucose; OD, optical density; LDH, lactate dehydrogenase; +PF,
HG + PF. |
In the present study, the effects of PF on the
mesangial cell injury under hyperglycemic conditions were
investigated.
Materials and methods
Materials
The lactate dehydrogenase (LDH) activity kit,
TRIzol® reagent, BCA protein assay kit, primary antibody
dilution buffer, RIPA, PMSF, dihydroethidium and the BeyoECL plus
kit were obtained from Beyotime Institute of Biotechnology. The
IL-6 ELISA kit (cat. no. EK0411) and TNF-α ELISA kit (cat. no.
EK0527) were purchased from Boster Biological Technology.
SYBR® Premix Ex Taq™ and the PrimeScript reverse
transcription reagent kit were obtained from Takara Biotechnology
Co., Ltd. The Annexin V-FITC kit was purchased from BD Biosciences.
MTT assay and the JC-1 probe were purchased from Beijing Solarbio
Science & Technology Co., Ltd. PF was obtained from Hunan
QianJinXiangJiang Pharmaceutical Industry Co., Ltd. D-glucose,
D-mannitol, 5% bovine serum albumin (BSA) blocking buffer and the
Masson's Trichrome stain kit (cat. no. G1340) were purchased from
Beijing Solarbio Science & Technology Co., Ltd. All antibodies
used for western blotting are detailed in Table I.
 | Table I.Specific details of the antibodies
used in western blot analysis. |
Table I.
Specific details of the antibodies
used in western blot analysis.
| Target | Supplier (cat.
no.) | Clone | Species | Dilution |
|---|
|
p66Shc | Novus Biological,
LLC (NBP2-20352) | Polyclonal Ab. | Rabbit | 1:500 |
| p-p66Shc
(Ser36) | Abcam
(ab54518) | Monoclonal Ab. | Mouse | 1:1,000 |
| IκBα | Abcam
(ab32518) | Monoclonal Ab. | Rabbit | 1:500 |
| p-NF-κB p65
(S536) | Abcam
(ab76302) | Monoclonal Ab. | Rabbit | 1:1,000 |
| p-IKKα/β (Ser
176) | Santa Cruz
Biotechnology, Inc. (sc-21661) | Polyclonal Ab. | Goat | 1:500 |
| IKKα/β | Affinity
Biosciences (AF6014) | Polyclonal Ab. | Rabbit | 1:300 |
| NF-κB p65 | Beyotime Institute
of Biotechnology (AF1234) | Monoclonal Ab. | Rabbit | 1:500 |
| Caspase-3 | Beyotime Institute
of Biotechnology (AF0081) | Polyclonal Ab. | Rabbit | 1:500 |
| Bcl2 | Boster Biological
Technology (BA0412) | Polyclonal Ab. | Rabbit | 1:300 |
| Fibronectin | Boster Biological
Technology (BA1772) | Polyclonal Ab. | Rabbit | 1:500 |
| Bax | Boster Biological
Technology (A00183) | Polyclonal Ab. | Rabbit | 1:500 |
| β-actin | Boster Biological
Technology (BM0627) | Monoclonal Ab. | Mouse | 1:500 |
Animal experiment
Male C57BL/6J mice (weight, 18–22 g; age, 8 weeks)
were purchased from the Experimental Animal Center of
Silaikejingda. Mice were kept under a 12-h light-dark cycle with a
controlled temperature (24±1°C) and humidity (50±10%), and allowed
free to access to food and water. All experimental protocols were
approved by the Ethics Committee of Animal Experiments of the
Central South University, and were performed in accordance with the
Guidelines for the Care and Use of Laboratory Animals.
A total of 45 mice were randomly divided into three
groups (n=15 mice per group) as follows: A control group, a model
group (DN) and the model + PF group (DN + PF group). Diabetic model
mice were established according to a previously described protocol
(24). PF was dissolved in 0.5%
sodium carboxymethyl cellulose (CMC-Na). DN + PF group mice were
treated intraperitoneally with PF (100 mg/kg) once daily for 12
consecutive weeks. Mice in the control and DN groups were
administered the same amount of CMC-Na (0.5%). Finally, the mice
were anaesthetized with sodium pentobarbital (50 mg/kg body weight)
and sacrificed by exsanguination. The kidney tissue was collected
for histopathological analysis.
Immunohistochemistry analysis
Kidney tissue was fixed with 4% paraformaldehyde
overnight at 4°C, embedded in paraffin blocks and cut into
5-µm-thick sections. The sections were dewaxed in xylene at room
temperature, rehydrated in a descending series of ethanol (100, 95,
80 and 70%), and washed with distilled water at room temperature,
followed by blocking with 3% hydrogen peroxidase at room
temperature for 10 min, and blocking with 5% BSA for 30 min at
37°C. The sections were incubated with fibronectin (1:200) and
collagen 4A1 (Boster Biological Technology; cat. no. BA2174; 1:200)
overnight at 4°C, followed by incubation with the HRP-conjugated
goat anti-rabbit (Boster Biological Technology; cat. no. BA1054;
1:5,000) for 1 h at room temperature. Subsequently, the sections
were incubated with the HRP substrate diaminobenzadine for color
development and stained with hematoxylin (0.5%) for 1 min at room
temperature. All stained sections were evaluated using conventional
light microscopy (magnification, ×400).
Masson's trichrome staining
Masson's trichrome staining was performed using the
Masson's Trichrome staining kit according to the manufacturer's
instructions. Briefly, kidney tissue was fixed with 4%
paraformaldehyde overnight at 4°C, embedded in paraffin blocks and
cut into 5-µm thick sections. The sections were dewaxed with xylene
at room temperature and then rehydrated with gradient ethanol (100,
95, 80 and 50%). After washing with distilled water, sections were
incubated with 0.5% Weigert's iron hematoxylin at room temperature
for 10 min. After rinsing in distilled water, the sections were
stained with 1% hydrochloric acid-alcohol at room temperature for
10 sec, and then stained with 0.7% Ponceau 2R-0.3% acid fuchsin
solution at room temperature for 10 min, followed by staining with
1% phosphomolybdic acid solution for 2 min and 2% aniline blue for
2 min at room temperature. Slides were then dehydrated in graded
ethanol (30, 50, 70, 95 and 100%) and sealed with neutral gum. The
images were captured using a light microscope (magnification,
×400).
Cell culture and treatment
Mouse mesangial cells (SV-40 MES-13 cells) were
purchased from the China Infrastructure of Cell Line Resource, and
cultured in DMEM (HyClone; Cyclone) containing 5% FBS (HyClone;
Cytiva) at 37°C in a humidified incubator with 5% CO2.
To examine the effects of HG on the cell viability, mouse mesangial
cells were treated with various concentrations of HG (20, 25, 30
and 35 mM) or mannitol (24.5 mM D-mannitol + 5.5 mM glucose) at
37°C for 24 h. D-mannitol was used to as an osmotic control for the
HG. MTT method was used to evaluate the cell viability.
Cell viability and apoptosis
assay
The viability of mouse mesangial cells was evaluated
using an MTT assay. Cells were plated in 96-well plates at a
density of 1.0×105 cells per well and treated with HG
(30 mM) with or without PF (25, 50, 100 and 200 µM) at 37°C for 24
h. MTT solution (5 mg/ml) was added to each well, and incubated
with the cells at 37°C for 4 h. Following incubation, 150 µl DMSO
was added to each well. The optical density (OD) value was measured
using a microplate reader at a wavelength of 450 nm.
The level of LDH released in the cell culture
supernatant was detected using the LDH cytotoxicity assay detection
kit. Briefly, the cell culture supernatant was obtained following
centrifugation at 1,000 × g for 10 min at 4°C. The supernatant was
harvested, and the level of LDH was determined following the
manufacturer's instructions of the kit.
Overexpression plasmids
A mouse p66Shc expression plasmid
(pcDNA-p66Shc) was constructed by Hunan Fenghui
Biotechnology Co., Ltd. Mesangial cells grown in 24-well plates
were transfected with the p66Shc overexpression plasmid
(pcDNA3.1-p66Shc; 1 or 2 ng/µl) or vector (pcDNA3.1; 2
ng/µl) using Lipofectamine® 3000 reagent (Invitrogen;
Thermo Fisher Scientific, Inc.) at 37°C for 24 h. Mesangial cells
were treated with HG (30 mM) in the presence or absence of 100 µM
PF at 37°C for 24 h.
Western blot analysis
Protein expression levels were determined via
western blot analysis. Cells or kidney tissue lysates were prepared
with RIPA lysis solution containing 10% PMSF. The protein
concentration was determined using a BCA protein assay kit and each
protein was then denatured at 95°C for 5 min. Protein samples (35
µg each) were separated by SDS-PAGE on 10% gels and transferred
onto a PVDF membrane (EMD Millipore). After blocking with 5% skim
milk in TBS containing 0.1% Tween-20 at room temperature for 1 h,
the membrane was incubated with primary antibodies against
p66Shc, p-P66Shc, caspase-3, p-IKKα/β
(Ser176), IKKα/β, Bcl2, Bax, fibronectin, IκBα, p-NF-κB p65 (S536)
and β-actin overnight at 4°C. Antibodies were diluted with primary
antibody dilution buffer (as indicated in Table I). Subsequently, the membrane was
incubated with HRP-conjugated goat anti-rabbit (Boster Biological
Technology; cat. no. BA1054; 1:5,000), HRP-conjugated goat
anti-mouse (Boster Biological Technology; cat. no. BA1050; 1:5,000)
or HRP-conjugated rabbit anti-goat secondary antibodies (Boster
Biological Technology; cat. no. BA1060; 1:5,000). Blots were
visualized by the BeyoECL Plus kit, and were captured using an
Amersham Imager 600 (Amersham; Cytiva) and semi-quantified using
ImageJ 1.37c software (National Institute of Health). The protein
level was normalized to that of β-actin.
RNA preparation and reverse
transcription-quantitative PCR (RT-qPCR)
Total RNA was extracted from the treated cells or
kidney tissue using TRIzol according to the manufacturer's
instructions. Total RNA (400 ng) was reverse transcribed to cDNA
using the PrimeScript RT reagent kit at 37°C for 15 min and 85°C
for 15 sec. qPCR was performed using SYBR-Green dye I and the
LightCycler96® (Roche Diagnostics GmbH). The sequences
of the RT-qPCR primers were as follows: Fibronectin forward,
5′-AGGCTGGATGATGGTGGACT-3′ and reverse, 5′-TGCTCCACGTGTCTCCAATC-3′;
collagen 4A1 forward, 5′-GGCATTGTGGAGTGTCAACC-3′ and reverse,
5′-ACAGGCAAGGCAGCTCTCTC-3′; and β-actin forward,
5′-ACTGCTCTGGCTCCTAGCAC-3′ and reverse, 5′-ACATCTGCTGGAAGGTGGAC-3′.
The thermal profile settings were as follows: Initial denaturation
at 95°C for 30 sec, followed by 40 cycles of denaturation at 95°C
for 5 sec and annealing at 60°C for 31 sec. The relative mRNA
expression levels were normalized to the expression of β-actin and
calculated using the 2−ΔΔCq method (25).
Determination of inflammatory cytokine
levels
The levels of IL-6 and TNF-α in the cell
supernatants were measured using ELISA kits. Briefly, the cell
culture supernatant was obtained following centrifugation at 1,000
× g for 10 min at 4°C and the levels of IL-6 and TNF-α were
determined according to the manufacturers' instructions. All
samples were measured in triplicate.
ROS measurement
Intracellular ROS levels were measured using
dihydroethidium. Following treatment, mesangial cells were washed
with PBS, and then incubated in serum-free DMEM with 5 µM
dihydroethidium for 30 min at 37°C. The fluorescence of ethidium
was detected using a fluorescence microscope (Carl Zeiss AG) at an
excitation wavelength of 535 nm and emission wavelength of 610
nm.
Immunofluorescence analysis
The cells were fixed in 4% paraformaldehyde for 15
min at room temperature, and then washed three times with 0.01 M
PBS for 5 min each. Following incubation with 0.1% Triton X-100 at
room temperature for 10 min, the cells were blocked in PBS-B
solution and incubated with specific anti-NF-κB p65 primary
antibody at 4°C overnight. The cells were then incubated with Cy3
antibody (Beyotime Institute of Biotechnology; cat. no. A0516;
1:500) for at room temperature for 1 h and the nuclei were stained
with DAPI at room temperature for 10 min. Images were captured
using a fluorescence microscope (magnification, ×400).
JC-1 staining
The mitochondrial membrane potential of the
mesangial cells was determined using a JC-1 kit according to the
manufacturer's instructions. This method is based on the ability of
JC-1 to form red fluorescent in normal mitochondria (aggregates
state of JC-1). The loss of mitochondrial membrane potential
generates the reduction of red fluorescence and a concomitant
increase in green fluorescence (monomeric state of JC-1) (26). The cells were washed with PBS three
times and incubated in JC-1 dye at 37°C for 30 min. Images were
captured using a fluorescence microscope at an excitation/emission
wavelength of 485/590 nm (magnification, ×400).
TUNEL assay
TUNEL staining was used to label apoptotic cells in
the glomerulus, according the manufacturer's protocol. Briefly, the
kidney tissue was fixed with 4% paraformaldehyde at 4°C overnight,
and then dehydrated in an ascending series of ethanol (70, 80, 90,
95 and 100%), embedded in paraffin and sectioned (5-µm thickness).
Subsequently, the sections were deparaffinized with xylene at room
temperature, rehydrated with a descending series of ethanol (100,
90 and 70%), and then incubated with proteinase K for 30 min at
room temperature. After washing with PBS, the sections were
incubated with TUNEL reaction mixture (Beyotime Institute of
Biotechnology; cat. no. C1089) at 37°C for 1 h. After washing with
PBS thrice, the sections were stained with DAPI (5 µg/ml) at room
temperature for 5 min, prior to addition of the antifade mounting
medium (Beyotime Institute of Biotechnology; cat. no. P0126).
Finally, the images were captured using a fluorescence microscope
(magnification, ×400) in six randomly selected fields of view.
Flow cytometry analysis
Following HG treatment, mesangial cells were
collected and washed with PBS three times. A total of
1–5×105 cells were suspended in binding buffer, and 5 µl
Annexin V-FITC and 5 µl PI were added. After mixing, the tube was
incubated in the dark at room temperature for 15 min. The samples
were analyzed using a BD FACSCalibur cell sorting system (BD
Biosciences) within 1 h, and data were analyzed using FlowJo 7.6.1
software (FlowJo LLC).
Statistical analysis
Statistical analysis was performed using SPSS
software (version 17.0; SPSS, Inc.). One-way ANOVA followed by
Tukey's test was performed for multiple groups (≥3) comparisons.
Data are presented as the mean ± SEM from ≥3 independent
experiments. P<0.05 was considered to indicate a statistically
significant difference.
Results
PF inhibits the mesangial cells injury
induced by HG
Mouse mesangial cells were treated with various
concentrations of HG (20, 25, 30 and 35 mM) to examine the effects
of HG on the cell viability. As presented in Fig. 1B, HG (30 or 35 mM) treatment
resulted in a significant decrease in cell viability compared with
the control group, while mannitol (30 mM) exerted no effect on cell
viability.
The effect of various concentrations of PF (25, 50,
100 and 200 µM) on the HG-induced decrease in the viability of
mesangial cells was examined. The results demonstrated that
pre-incubation of the mesangial cells with PF (100 and 200 µM)
caused in an increase in cell viability compared with the HG (30
mM) group (Fig. 1C). The subsequent
experiments were performed using 100 µM concentrations of PF.
The results of flow cytometry and LDH release assay
revealed that cell apoptosis and LDH release were significantly
increased in the HG group, compared with the control group, and
these effects were attenuated by incubation with PF (Fig. 1D-F). These results suggested that PF
prevented mesangial cell injury induced by HG.
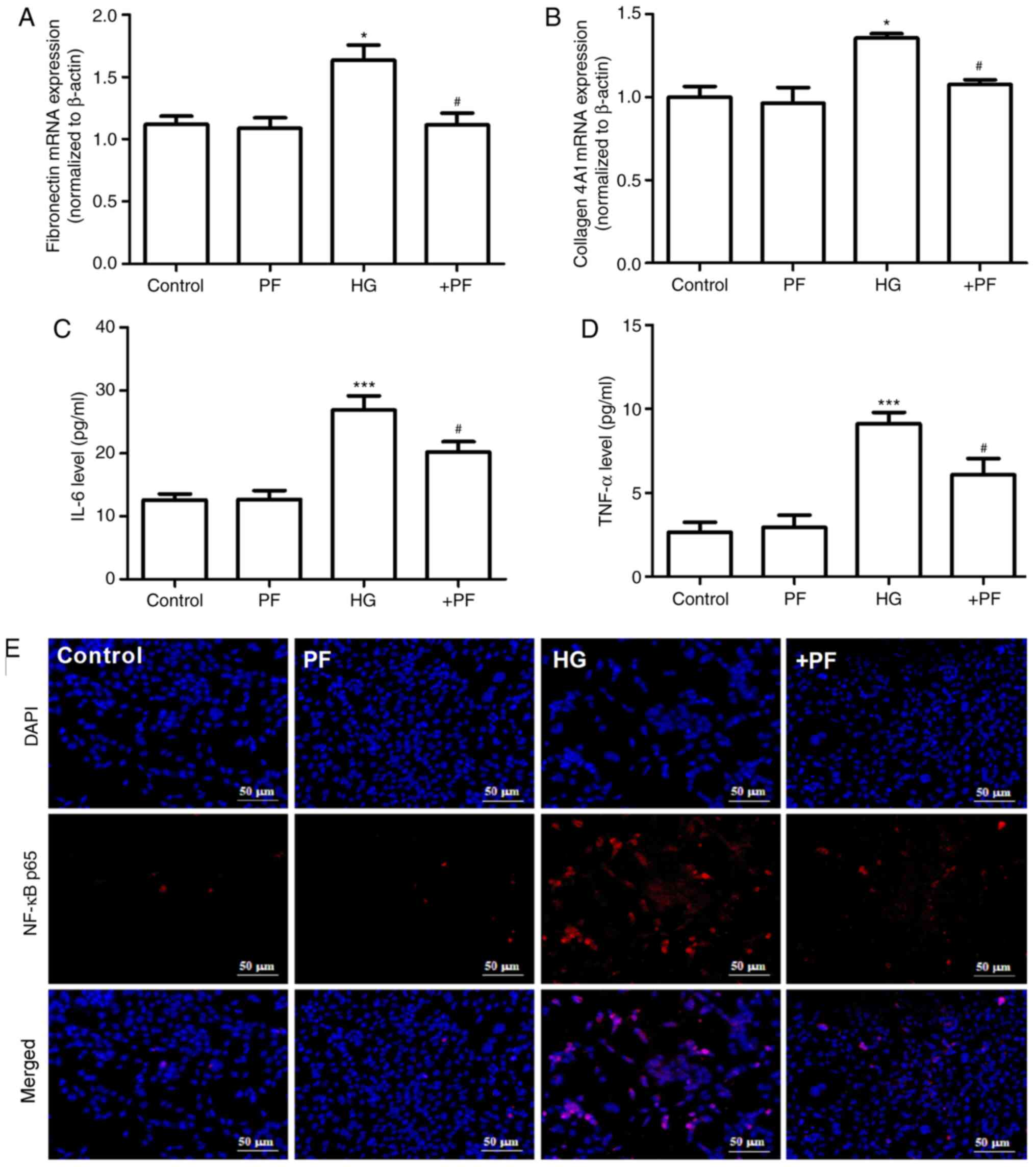
PF attenuates HG-induced inflammatory
cytokine and fibrosis in mesangial cells
The effects of PF on the HG-induced inflammatory
response and fibrosis in mesangial cells were evaluated. As
presented in Fig. 2A and B,
compared with the control group, the mRNA expression levels of
fibronectin and collagen 4A1 in mesangial cells were increased
following HG treatment, and these changes were mitigated when the
mesangial cells were incubated with PF. Likewise, treatment of the
mesangial cells with PF significantly reversed the increase in the
IL-6 and TNF-α levels in the cell supernatant induced by HG
(Fig. 2C and D).
The results of immunofluorescence analysis
demonstrated that, compared with the control group, HG induced p65
translocation from the cytoplasm to the nucleus, and PF attenuated
this effect in the HG-treated mesangial cells (Fig. 2E). Taken together, these results
suggest that PF attenuates the HG-induced production of
inflammatory cytokines and fibrosis in mesangial cells.
PF inhibits the expression of
p-p66Shc in HG-induced mesangial cells
It has been previously reported that
p66Shc is involved in the progression of DN (27), and the phosphorylation of
p66Shc at serine 36 serves a key role in
p66Shc activation (28).
The present study identified that HG induced an increase in
p-p66Shc or p-p66Shc/p66Shc
protein expression levels in mesangial cells compared with the
control group, but had no effect on p66Shc expression.
Moreover, co-incubation with PF decreased the HG-induced
phosphorylation of p66Shc (Fig. 3A-C). Similarly, treatment of the
mesangial cells with PF reversed the increase in the expression
levels of p-IKKα/β/IKKα/β and p-NF-κB p65/NF-κB p65 induced by HG
(Fig. 3D-F). The result of
immunofluorescence analysis demonstrated that PF prevented the
HG-induced ROS generation in mesangial cells (Fig. 3G). These results indicate that PF
inhibits the HG-induced phosphorylation of p66Shc,
IKKα/β and NF-κB p65, as well as ROS generation.
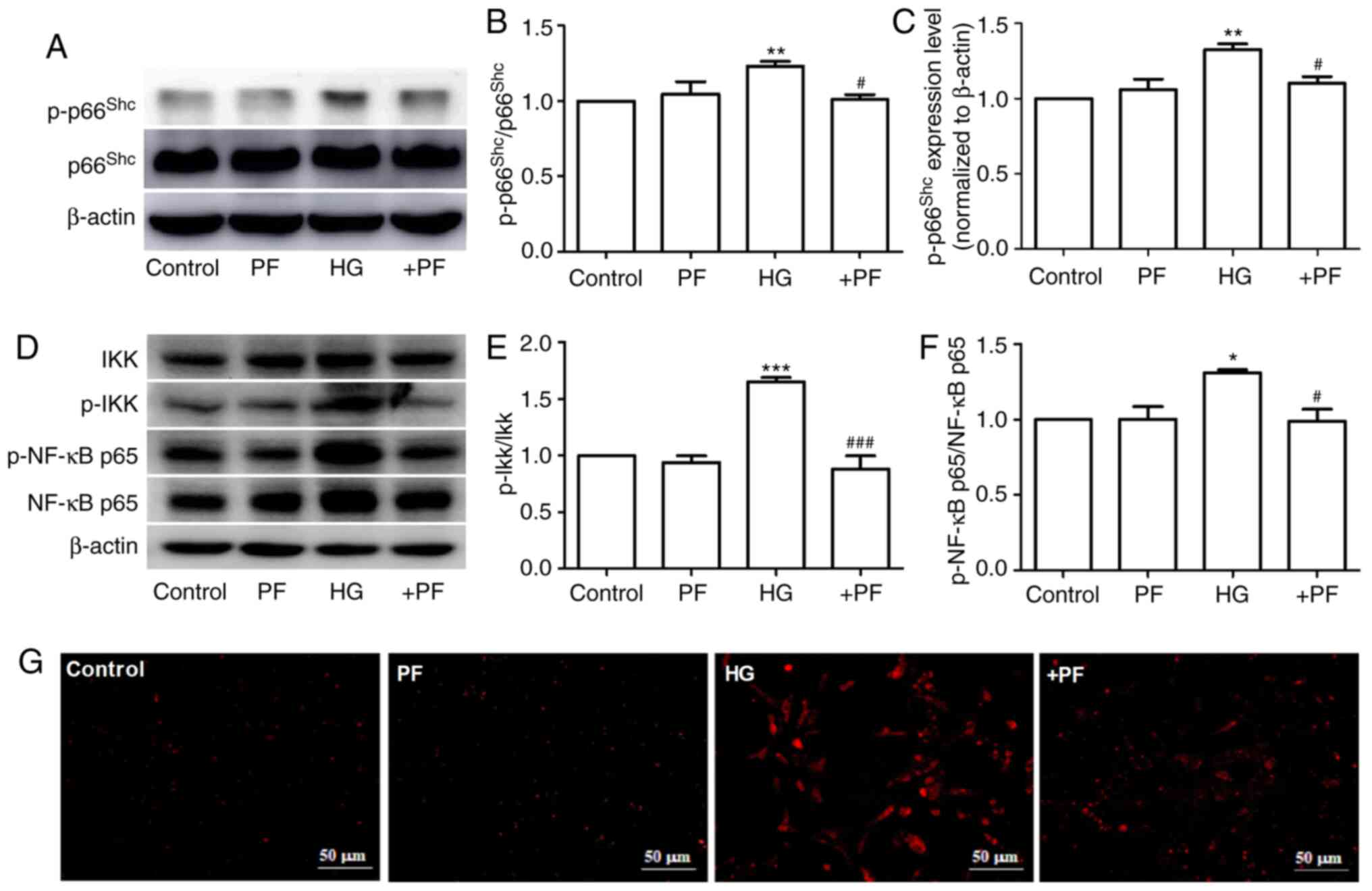 | Figure 3.PF inhibits the expression of
p-p66Shc in HG-exposed mesangial cells. (A) Western blot
analysis of p-p66shc and total p66shc. (B)
Densitometric analyses of p-p66shc/p66shc and
(C) p-p66shc. (D) Western blot analysis of IKKα/β,
p-IKKα/β, NF-κB p65 and p-NF-κB p65. (E) Densitometric analyses of
p-IKKα/β/IKKα/β and (F) p-NF-κB p65/NF-κB p65. (G) Fluorescence
image of ROS detected using dihydroethidium; ×400 magnification.
Data are expressed as the mean ± SEM, n=3. *P<0.05, **P<0.01,
***P<0.001 vs. control group; #P<0.05,
###P<0.001 vs. HG group. PF, piperazine ferulate; HG,
high glucose; p-, phosphorylated; Shc, Src homology/collagen; +PF,
HG + PF. |
PF regulates HG-induced mesangial
cells injury by inhibiting p66Shc
To assess whether p66Shc mediates the
protective effects of PF against mesangial cell damage induced by
HG, the overexpression of the p66Shc gene was induced
using a p66Shc plasmid (Fig.
4A and B). PF treatment attenuated the HG-induced the increased
level of IL-6 in the cell culture supernatant, while co-treatment
of the mesangial cells with pcDNA-p66Shc abolished the
cytoprotective effects of PF (Fig.
4C). PF was demonstrated to attenuate the increase in cleaved
caspase-3, p-IKK, Bax and fibronectin expression levels and the
decrease in Bcl2 expression in mesangial cells exposed to HG, and
co-treatment with pcDNA-p66Shc was able to reverse the
protective effects of PF under HG conditions (Fig. 4D-I). Furthermore, PF treatment
restored the decrease in the mitochondrial membrane potential of
mesangial cells exposed to HG, and the protective effects of PF
were abrogated when the mesangial cells were transfected with
p66Shc plasmid (Fig.
4J). These data indicated that PF exerted its protective
effects via the inhibition of p66Shc.
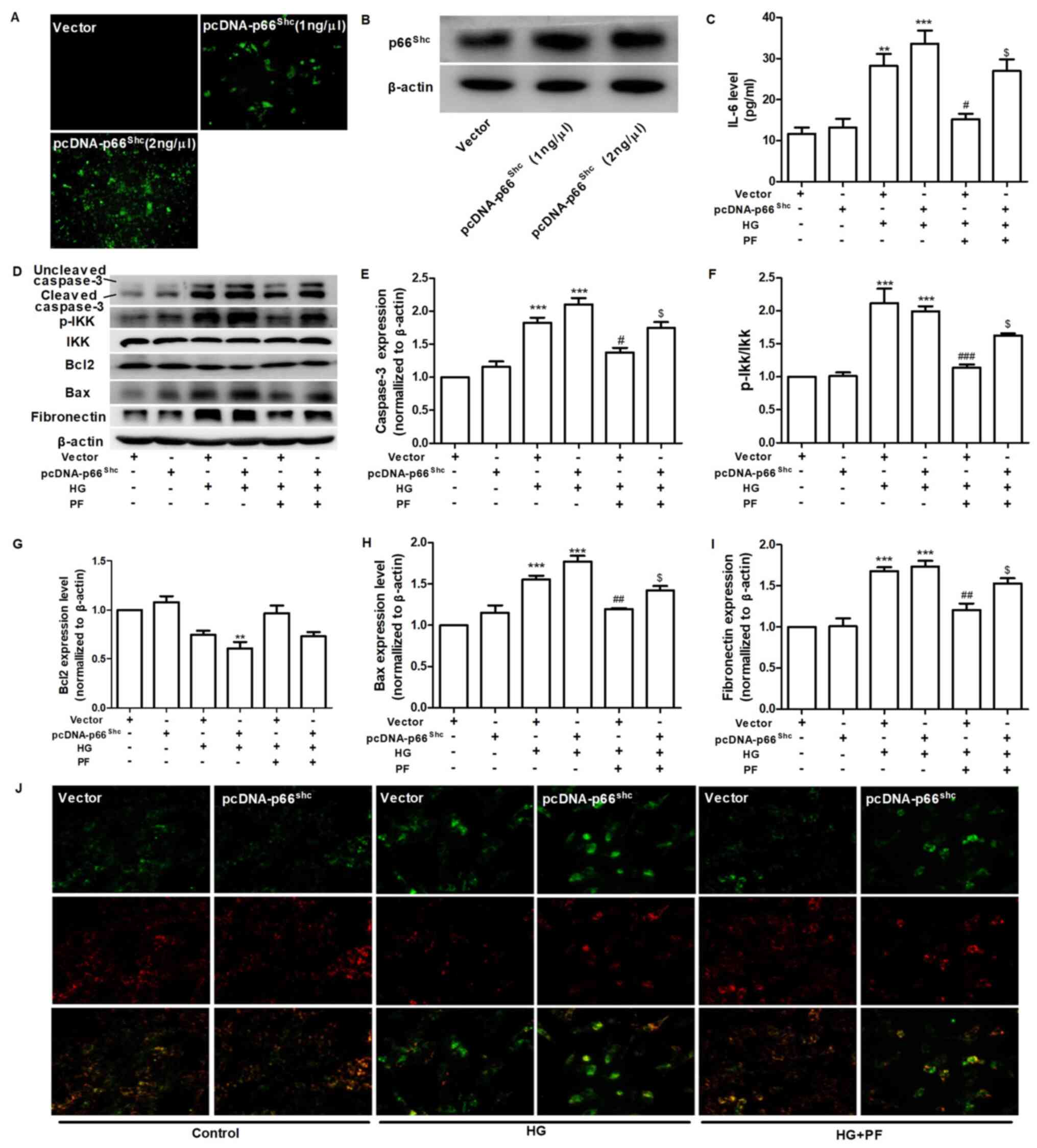 | Figure 4.PF regulates HG-induced mesangial
cell injury by inhibiting p66shc. (A) Effect of
pcDNA-p66shc (1 ng/µl or 2 ng/µl) on the expression of
p66shc. (B) Western blot analysis verified the validity
of the pcDNA-p66Shc. (C) Level of IL-6 in the cell
culture supernatant. (D) Western blot analysis of caspase-3,
p-IKKα/β, total IKKα/β, Bcl2, Bax, fibronectin and β-actin.
Densitometric analyses of (E) caspase-3, (F) p-IKK/IKK, (G) Bcl2,
(H) Bax and (I) fibronectin. (J) Representative image of mesangial
cells stained with JC-1 (red for aggregate form of JC-1 and green
for monomeric form); ×400 magnification. Data are expressed as the
mean ± SEM, n=3. **P<0.01, ***P<0.001 vs. vector group;
#P<0.05, ##P<0.01,
###P<0.001 vs. vector + HG group;
$P<0.01 vs. vector + HG + PF group. PF, piperazine
ferulate; HG, high glucose; vector, empty vector control; p-,
phosphorylated; Shc, Src homology/collagen. |
PF attenuates inflammation and
apoptosis in the kidney tissue of diabetic mice
Subsequently, the effects of PF on apoptosis and
inflammatory signaling molecules in the kidney tissues of diabetic
mice were assessed. As presented in Fig. 5A-D, PF attenuated the HG-induced
increase in the protein expression levels of p66shc,
p-p66shc, p-p66Shc/p66Shc, p-NF-κB
p65/NF-κB p65 and p-IKK/IKK, and restored the loss of IκBα protein
expression observed in kidney tissue of diabetic mice. The results
of TUNEL assay revealed that PF treatment also caused a notable
decrease in glomerular cell apoptosis (Fig. 5E). These results indicated that PF
inhibited the activation of inflammatory signals and apoptosis
in vivo.
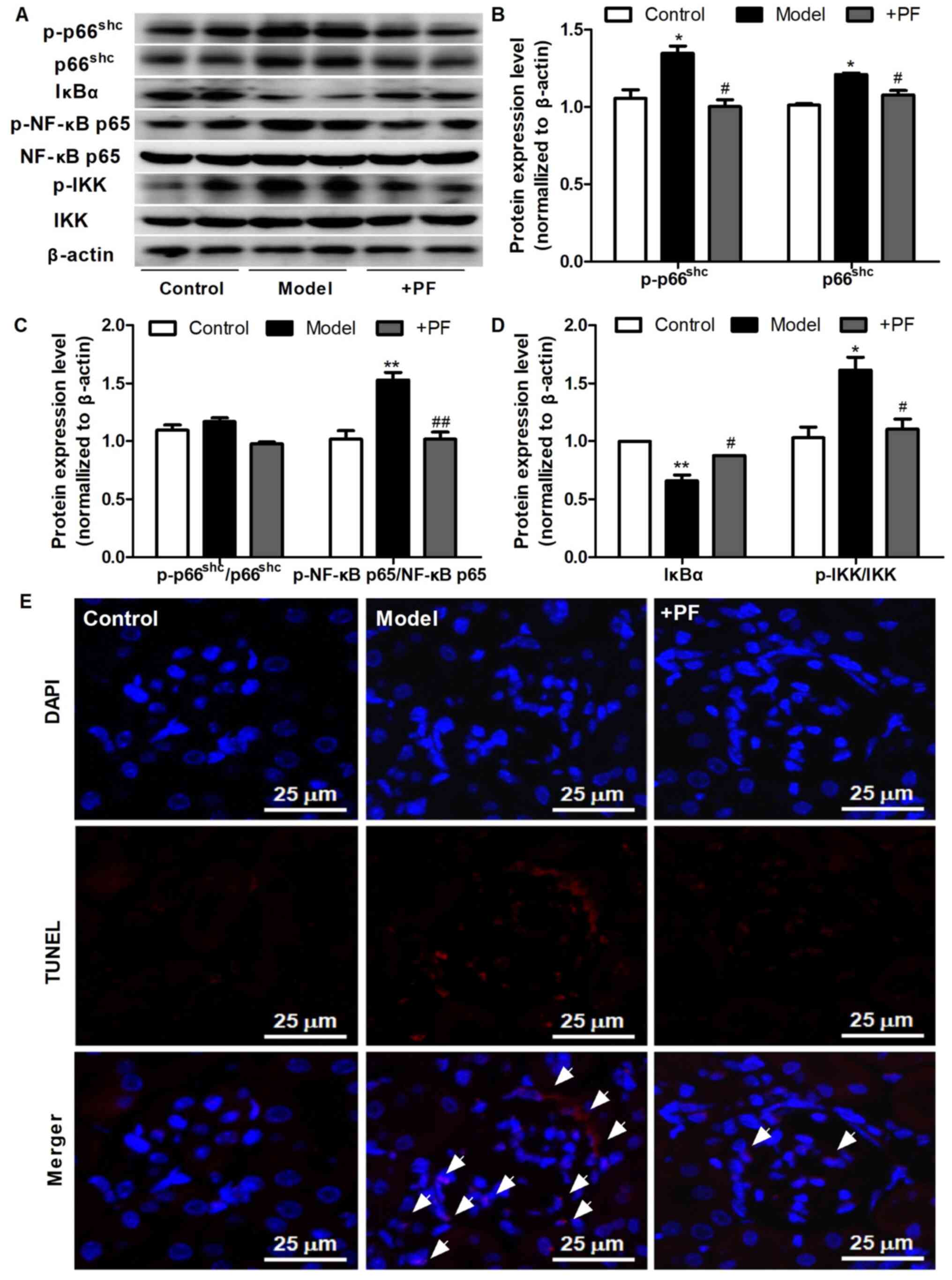 | Figure 5.PF attenuates kidney injury in
diabetic mice. (A) Western blot analysis of p-p66shc,
total p66shc, IκBα, p-NF-κB p65, total NF-κB p65,
p-IKKα/β, total IKKα/β and β-actin. (B) Densitometric analysis of
p-p66shc and total p66shc. (C) Densitometric
analyses of p-p66shc/p66shc and p-NF-κB
p65/NF-κB p65. (D) Densitometric analyses of IκBα and
p-IKKα/β/IKKα/β. (E) Representative image of the TUNEL staining;
×400 magnification; white arrowheads indicate TUNEL-positive cells.
Data are expressed as the mean ± SEM, n=3. *P<0.05, **P<0.01
vs. control group; #P<0.05, ##P<0.01
vs. HG group. PF, piperazine ferulate; HG, high glucose; p-,
phosphorylated; Shc, Src homology/collagen; +PF, diabetic
nephropathy + PF. |
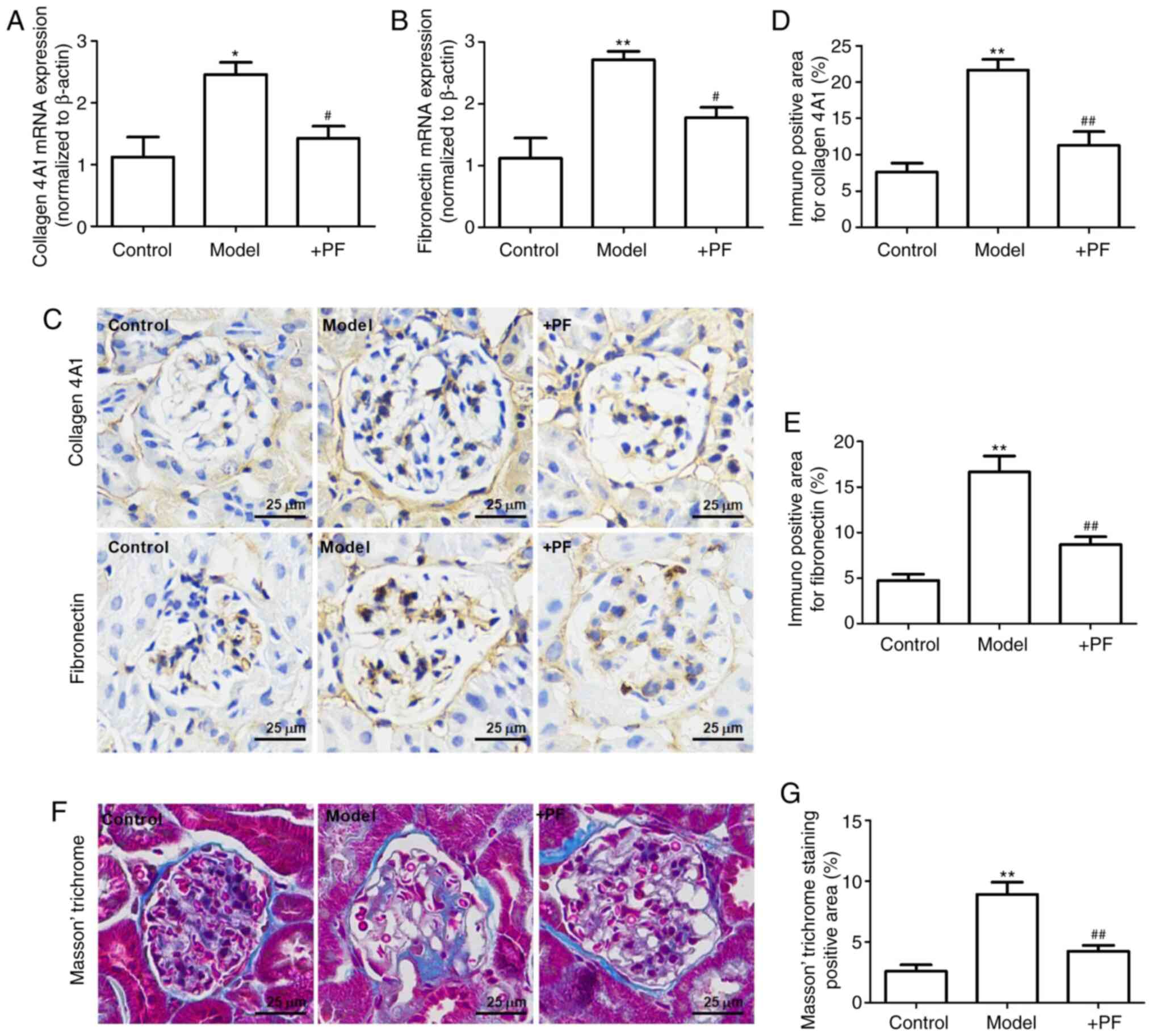
PF attenuates mesangial matrix
expansion in diabetic mice
The results of RT-qPCR revealed that the mRNA
expression levels of collagen 4A1 and fibronectin were increased in
the model group when compared with the control group. Furthermore,
PF reversed these changes in the mRNA expression levels of collagen
4A1 and fibronectin (Fig. 6A and
B). Immunohistochemistry revealed that hyperglycemia increased
the expression levels of collagen 4A1 and fibronectin in the
glomerulus, and these effects were inhibited by PF (Fig. 6C-E). Masson's trichrome staining
identified that PF reversed glomerular fibrosis induced by
hyperglycemia (Fig. 6F and G).
These results suggested that PF reversed the HG-induced mesangial
matrix expansion in the glomerulus.
Discussion
DN is the leading cause of end-stage renal failure,
but the pathogenesis of DN is not yet fully understood. Persistent
hyperglycemia leads to increased glycation end products, oxidative
stress, the production of inflammatory cytokines and hemodynamic
abnormalities (29). These factors
individually and/or synergistically result in the pathological
features of DN, including the thickening of the glomerular basement
membrane, damage to glomerular cells, mesangial matrix expansion
and tubulointerstitial fibrosis (30). HG-induced damage to glomerular
cells, including glomerular endothelial cells, podocytes and
mesangial cells, is a key factor in exacerbating the progression of
DN (11,30–32).
Moreover, damage to glomerular endothelial cells and podocytes
causes filtration barrier damage and accelerates the excretion of
albuminuria (7,31). Mesangial cells maintain the
structure and function of the glomerulus, and also regulate filter
barrier function by controlling the capillary surface area
(31). Therefore, preventing
mesangial cell apoptosis is of utmost importance for the prevention
and treatment of DN.
ROS serve a key role in glomerular mesangial cell
apoptosis under hyperglycemic conditions (33). Some in vivo studies have
reported that antioxidant compounds, such as N-acetylcysteine
(34), folic acid (35) and carnosic acid (36), were able to delay the progression of
DN by regulating the production of ROS. However, none of these
compounds have been approved for clinical use as anti-diabetic
nephropathy drugs. Therefore, the therapeutic targets of DN warrant
further investigation.
p66Shc is predominantly located in the
cytoplasm, with 10–40% located in the mitochondrial membrane space
in a complex with the mitochondrial heat shock protein (37), and is involved in the apoptosis of
mesangial cells under hyperglycemic conditions (17).
In China, PF is used in the treatment of various
kidney diseases, and no severe adverse reactions have been reported
to date. An acute toxicity test revealed that the median lethal
dose of PF to Kunming mice was 3,580.1±251.7 mg/kg. Moreover,
reproductive toxicity studies have reported that PF has no obvious
embryonic side-effects and teratogenic effects (the data are from
the package insert of PF) (38). In
the present study, it was found that PF was an efficient drug
against HG-induced mesangial cell injury by inhibiting
p66Shc. For example, PF decreased the levels of IL-6 and
TNF-α, the generation of ROS and the nuclear translocation of NF-κB
p65 in HG-treated mesangial cells. Furthermore, PF inhibited the
HG-induced upregulation of fibronectin and collagen 4A1 expression
levels in in vitro and in vivo experiments. It was
also demonstrated that the multiple pharmacological functions of
PF, including anti-inflammatory, anti-apoptotic and anti-fibrotic
effects, were abolished when the mesangial cells overexpressed of
p66Shc. The phosphorylation of p66Shc is a
critical step for the production of ROS, and both UV radiation and
H2O2 can phosphorylate the serine residue of
p66Shc (39). The
present study identified that PF decreased the protein expression
level of p-p66Shc in mesangial cells exposed to HG.
However, the present study also has limitations. Although the study
demonstrated that PF could inhibit the phosphorylation level of
p66Shc in both in vivo and in vitro
experiments, the mechanism by which PF inhibits the phosphorylation
of p66Shc is not clear. Further studies are required to
determine the mechanisms via which PF inhibits the phosphorylation
of serine of p66Shc through molecular docking and
molecular-protein interaction test.
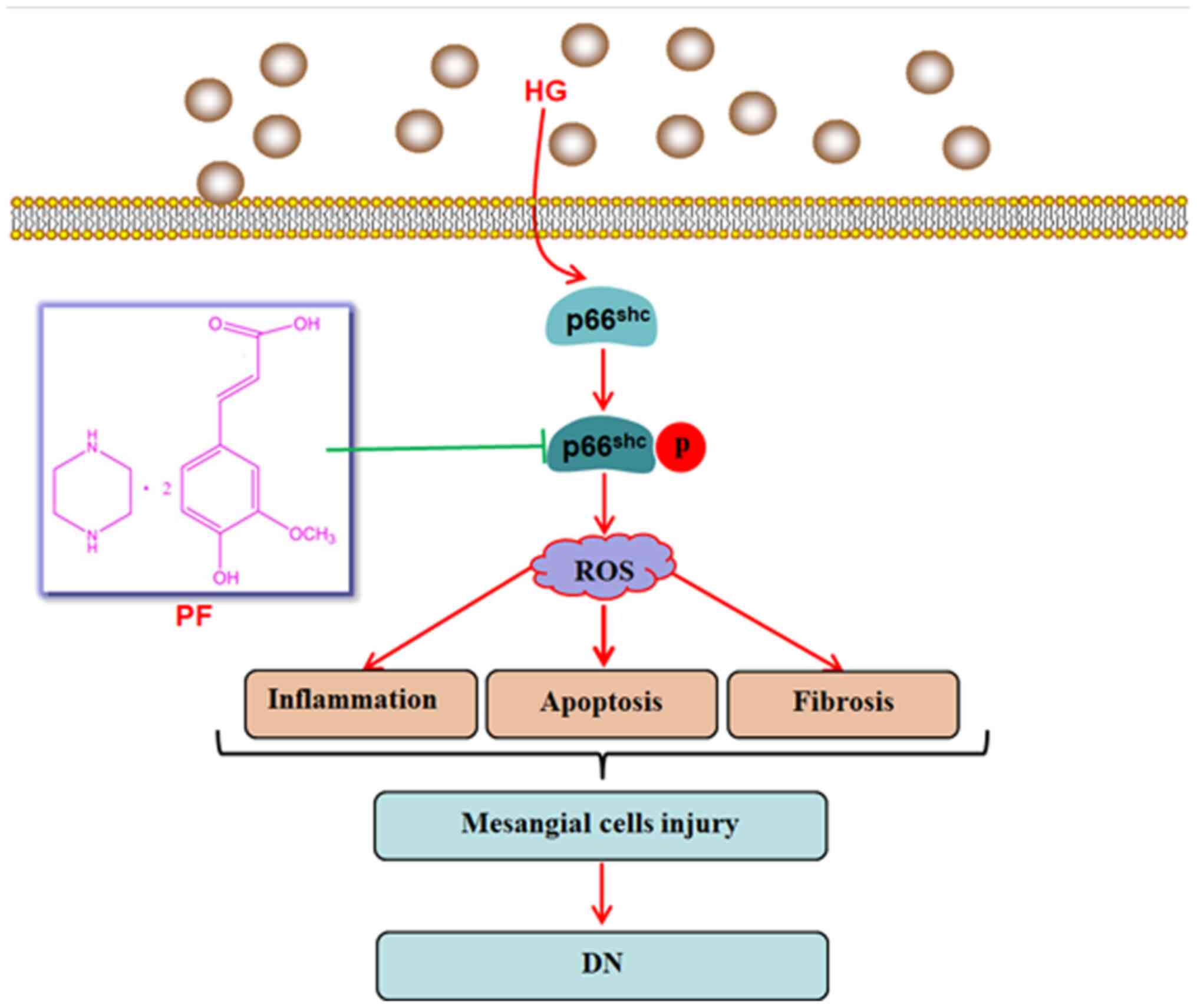
In conclusion, the present study demonstrated that
the inhibition of p66Shc activation may be used as a
therapeutic approach to attenuate mesangial cells damage induced by
hyperglycemia, and that PF attenuated HG-induced mesangial cell
injury by inhibiting p66Shc (Fig. 7). The results provided a potential
mechanism via which PF attenuates the development of DN, but
further studies are warranted to evaluate the exact mechanisms
underlying the regulatory effects of PF on p66Shc.
Acknowledgements
Not applicable.
Funding
The present study was supported financially by the
National Natural Science Foundation of China (grant no. 81603171),
Hunan Provincial Natural Scientific Foundation (grant nos.
2018JJ3743 and 2020JJ5841), Scientific Research Project of Hunan
Provincial Health Commission (grant nos. B2019157 and B2019158) and
the Open Sharing Fund for the Large-scale Instruments of Central
South University (grant no. CSUZC202055).
Availability of data and materials
All data generated or analyzed during the present
study are included in this published article.
Authors' contributions
DXX, LYY and XDY designed the study. YYY and RRD
performed the experiments. YYY and ZC analyzed the data. YYY wrote
the manuscript, and revised the manuscript with XDY. YYY and RRD
confirm the authenticity of all the raw data. All authors read and
approved the final version of the manuscript.
Ethics approval and consent to
participate
All experimental protocols were approved by the
Ethics Committee of Animal Experiments of the Central South
University, and were performed in accordance with the Guidelines
for the Care and Use of Laboratory Animals.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Mou X, Chenv JW, Zhou DY, Liu K, Chen LJ,
Zhou D and Hu YB: A novel identified circular RNA, circ_0000491,
aggravates the extracellular matrix of diabetic nephropathy
glomerular mesangial cells through suppressing miR101b by targeting
TGFbetaRI. Mol Med Rep. 22:3785–3794. 2020.PubMed/NCBI
|
|
2
|
Cho NH, Shaw JE, Karuranga S, Huang Y, da
Rocha Fernandes JD, Ohlrogge AW and Malanda B: IDF diabetes atlas:
Global estimates of diabetes prevalence for 2017 and projections
for 2045. Diabetes Res Clin Pract. 138:271–281. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yang X, Hu C, Wang S and Chen Q: Clinical
efficacy and safety of Chinese herbal medicine for the treatment of
patients with early diabetic nephropathy: A protocol for systematic
review and meta-analysis. Medicine (Baltimore). 99:e206782020.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Jia Q, Yang R, Liu XF, Ma SF and Wang L:
Genistein attenuates renal fibrosis in streptozotocin-induced
diabetic rats. Mol Med Rep. 19:423–431. 2019.PubMed/NCBI
|
|
5
|
Zhang J, Dong XJ, Ding MR, You CY, Lin X,
Wang Y, Wu MJ, Xu GF and Wang GD: Resveratrol decreases high
glucose induced apoptosis in renal tubular cells via suppressing
endoplasmic reticulum stress. Mol Med Rep. 22:4367–4375.
2020.PubMed/NCBI
|
|
6
|
Dou L and Jourde-Chiche N: Endothelial
toxicity of high glucose and its by-products in diabetic kidney
disease. Toxins (Basel). 11:5782019. View Article : Google Scholar
|
|
7
|
Tung CW, Hsu YC, Shih YH, Chang PJ and Lin
CL: Glomerular mesangial cell and podocyte injuries in diabetic
nephropathy. Nephrology (Carlton). 23 (Suppl 4):S32–S37. 2018.
View Article : Google Scholar
|
|
8
|
Chen X, Yang Y, Liu C, Chen Z and Wang D:
Astragaloside IV ameliorates high glucoseinduced renal tubular
epithelial mesenchymal transition by blocking mTORC1/p70S6K
signaling in HK2 cells. Int J Mol Med. 43:709–716. 2019.PubMed/NCBI
|
|
9
|
Maezawa Y, Cina D and Quaggin SE:
Glomerular cell biology. Chapter 22. The Kidney. 5th edition.
Alpern RJ, Moe OW and Caplan M: San Diego: Academic Press; pp.
721–755. 2013
|
|
10
|
Mishra R, Emancipator SN, Kern T and
Simonson MS: High glucose evokes an intrinsic proapoptotic
signaling pathway in mesangial cells. Kidney Int. 67:82–93. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Tsai YC, Kuo MC, Hung WW, Wu LY, Wu PH,
Chang WA, Kuo PL and Hsu YL: High glucose induces mesangial cell
apoptosis through miR-15b-5p and promotes diabetic nephropathy by
extracellular vesicle delivery. Mol Ther. 28:963–974. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ostergaard JA, Cooper ME and
Jandeleit-Dahm KAM: Targeting oxidative stress and anti-oxidant
defence in diabetic kidney disease. J Nephrol. 33:917–929. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Di Lisa F, Giorgio M, Ferdinandy P and
Schulz R: New aspects of p66Shc in ischaemia reperfusion injury and
other cardiovascular diseases. Br J Pharmacol. 174:1690–1703. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Giorgio M, Migliaccio E, Orsini F,
Paolucci D, Moroni M, Contursi C, Pelliccia G, Luzi L, Minucci S,
Marcaccio M, et al: Electron transfer between cytochrome c
and p66Shc generates reactive oxygen species that trigger
mitochondrial apoptosis. Cell. 122:221–233. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Clark JS, Faisal A, Baliga R, Nagamine Y
and Arany I: Cisplatin induces apoptosis through the ERK-p66shc
pathway in renal proximal tubule cells. Cancer Lett. 297:165–170.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Miller B, Palygin O, Rufanova VA, Chong A,
Lazar J, Jacob HJ, Mattson D, Roman RJ, Williams JM, Cowley AWJr,
et al: p66Shc regulates renal vascular tone in hypertension-induced
nephropathy. J Clin Invest. 126:2533–2546. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Menini S, Amadio L, Oddi G, Ricci C, Pesce
C, Pugliese F, Giorgio M, Migliaccio E, Pelicci P, Iacobini C and
Pugliese G: Deletion of p66Shc longevity gene protects against
experimental diabetic glomerulopathy by preventing diabetes-induced
oxidative stress. Diabetes. 55:1642–1650. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhan M, Usman I, Yu J, Ruan L, Bian X,
Yang J, Yang S, Sun L and Kanwar YS: Perturbations in mitochondrial
dynamics by p66Shc lead to renal tubular oxidative injury in human
diabetic nephropathy. Clin Sci (Lond). 132:1297–1314. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Jiang W, Xiao T, Han W, Xiong J, He T, Liu
Y, Huang Y, Yang K, Bi X, Xu X, et al: Klotho inhibits
PKCalpha/p66SHC-mediated podocyte injury in diabetic nephropathy.
Mol Cell Endocrinol. 494:1104902019. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Li D, Li B, Peng LX, Liu R and Zeng N:
Therapeutic efficacy of piperazine ferulate combined with
irbesartan in diabetic nephropathy: A Systematic review and
meta-analysis. Clin Ther. 42:2196–2212. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liu Z, Pan J, Sun C, Zhou J and Li NA:
Clinical effects of perazine ferulate tablets combined with
eucalyptol limonene pinene enteric soft capsules for treatment of
children with IgA nephropathy. Exp Ther Med. 12:169–172. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zheng L, Chen S, Wang F, Huang S, Liu X,
Yang X, Zhou H, Zhao GP, Luo M, Li S and Chen J: Distinct responses
of gut microbiota to Jian-Pi-Yi-Shen decoction are associated with
improved clinical outcomes in 5/6 nephrectomized rats. Front
Pharmacol. 11:6042020. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Jianzhi S, Qizeng W, Bin L, Wenhui L,
Yunpeng C, Chenrong F, Lin Z and Huiting C: Piperazine ferulate
exerts antihypertensive effect and improves endothelial function in
vitro and in vivo via the activation of endothelial nitric oxide
synthase. Cell Mol Biol (Noisy-le-grand). 65:119–124. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yang YY, Shi LX, Li JH, Yao LY and Xiang
DX: Piperazine ferulate ameliorates the development of diabetic
nephropathy by regulating endothelial nitric oxide synthase. Mol
Med Rep. 19:2245–2253. 2019.PubMed/NCBI
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wong YH and Abdul Kadir H: Induction of
mitochondria-mediated apoptosis in Ca Ski human cervical cancer
cells triggered by mollic acid arabinoside isolated from leea
indica. Evid Based Complement Alternat Med. 2012:6847402012.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Cheng YS, Chao J, Chen C, Lv LL, Han YC
and Liu BC: The PKCβ-p66shc-NADPH oxidase pathway plays a crucial
role in diabetic nephropathy. J Pharm Pharmacol. 71:338–347. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Haller M, Khalid S, Kremser L, Fresser F,
Furlan T, Hermann M, Guenther J, Drasche A, Leitges M, Giorgio M,
et al: Novel Insights into the PKCbeta-dependent Regulation of the
Oxidoreductase p66Shc. J Biol Chem. 291:23557–23568. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Dragos D, Manea MM, Timofte D and Ionescu
D: Mechanisms of herbal nephroprotection in diabetes mellitus. J
Diabetes Res. 2020:57105132020. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Rayego-Mateos S, Morgado-Pascual JL,
Opazo-Rios L, Guerrero-Hue M, Garcia-Caballero C, Vazquez-Carballo
C, Mas S, Sanz AB, Herencia C, Mezzano S, et al: Pathogenic
pathways and therapeutic approaches targeting inflammation in
diabetic nephropathy. Int J Mol Sci. 21:37982020. View Article : Google Scholar
|
|
31
|
Davidson A, Berthier C and Kretzler M:
Pathogenetic Mechanisms in lupus nephritis. Chapter 18. Dubois'
Lupus Erythematosus and Related Syndromes. 8th edition. Wallace DJ
and Hahn BH: W.B. Saunders; Philadelphia, PA: pp. 237–255. 2013,
View Article : Google Scholar
|
|
32
|
Fu J, Wei C, Zhang W, Schlondorff D, Wu J,
Cai M, He W, Baron MH, Chuang PY, Liu Z, et al: Gene expression
profiles of glomerular endothelial cells support their role in the
glomerulopathy of diabetic mice. Kidney Int. 94:326–345. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wan C, Su H and Zhang C: Role of NADPH
oxidase in metabolic disease-related renal injury: An update. Oxid
Med Cell Longev. 2016:78130722016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chen X and Fang M: Oxidative stress
mediated mitochondrial damage plays roles in pathogenesis of
diabetic nephropathy rat. Eur Rev Med Pharmacol Sci. 22:5248–5254.
2018.PubMed/NCBI
|
|
35
|
Ebaid H, Bashandy SAE, Abdel-Mageed AM,
Al-Tamimi J, Hassan I and Alhazza IM: Folic acid and melatonin
mitigate diabetic nephropathy in rats via inhibition of oxidative
stress. Nutr Metab (Lond). 17:62020. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Xie Z, Zhong L, Wu Y, Wan X, Yang H, Xu X
and Li P: Carnosic acid improves diabetic nephropathy by activating
Nrf2/ARE and inhibition of NF-κB pathway. Phytomedicine.
47:161–173. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Orsini F, Migliaccio E, Moroni M, Contursi
C, Raker VA, Piccini D, Martin-Padura I, Pelliccia G, Trinei M,
Bono M, et al: The life span determinant p66Shc localizes to
mitochondria where it associates with mitochondrial heat shock
protein 70 and regulates trans-membrane potential. J Biol Chem.
279:25689–25695. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
DRUGDATAEXPY. 2009. Chongqing, .
http://zy.yaozh.com/instruct/20160513-1/35.jpgMarch
1–2021
|
|
39
|
Nemoto S and Finkel T: Redox regulation of
forkhead proteins through a p66shc-dependent signaling pathway.
Science. 295:2450–2452. 2002. View Article : Google Scholar : PubMed/NCBI
|