Introduction
Ischemic stroke is one of the most important causes
of death and disability worldwide (1,2). In
China, stroke became the top leading cause of mortality in 2017.
Approximately 1.5 million people die from strokes each year
(3). In ischemic stroke, the
insufficient supply of oxygen and glucose causes brain damage
within the subsequent few hours (4,5). The
primary method is to restore the blood supply of ischemic brain
tissue as soon as possible. However, the reopening of the occluded
cerebrovascular usually leads to pathological damage in the
ischemic tissue, which may further aggravate or potentially make
the damage irreversible. Neuronal injury caused by
ischemia/reperfusion is a complex process involving various
mechanisms, including apoptosis, oxygen-free radicals, glutamic
acid toxicity and [Ca2+] overload (6). It remains an area of interest to
perform drug research on stroke treatment (7).
Apoptosis is an essential mechanism of ischemic
stroke (8,9). Apoptosis is regulated through the
death receptor pathway, mitochondrial pathway and endoplasmic
reticulum pathway. In the majority of vertebrates, apoptosis is
regulated by the mitochondrial pathway (10,11).
Mitochondria are the active centers of apoptosis regulation and the
executors (12). The pro-apoptotic
factor causes the mitochondrial permeability transition pore (MPTP)
to open excessively, decreasing the membrane potential, releasing
cytochrome c from the mitochondria to the cytosol. Next, caspase is
activated and cells undergo apoptosis (13,14).
Mu-Xiang-You prescription is a classic prescription
of Hui medicine, which affects ischemic stroke (15). Aucklandia lappa Decne is the
main drug in the prescription (45% by weight); therefore, we
hypothesized that Aucklandia lappa Decne is an essential
drug in treating ischemic stroke. CT
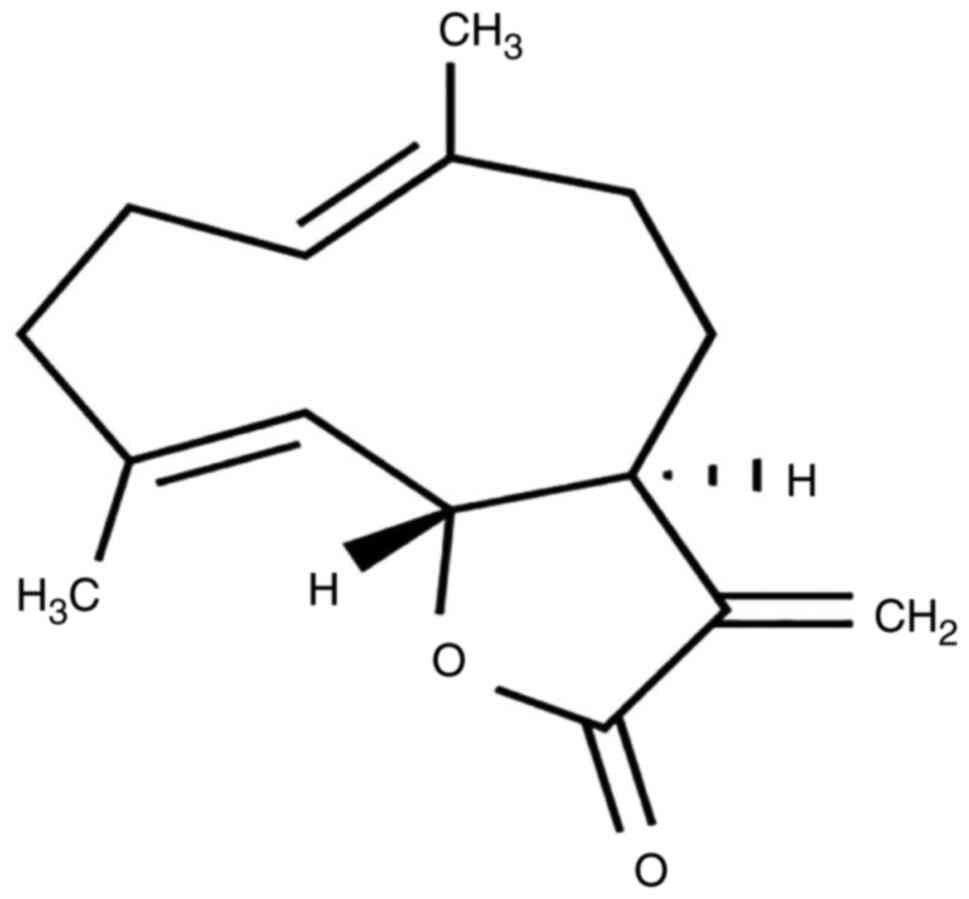
(C15H20O2; Fig. 1), an active sesquiterpene lactone,
is one of the main useful components of Aucklandia lappa
Decne. CT is used to control the quality of Aucklandia lappa
Decne in Chinese Pharmacopoeia. We hypothesized that the effect of
this prescription in the treatment of ischemic stroke is associated
with CT. CT has multiple pharmacological activities, including
inhibiting tumour cell proliferation (16) and decreasing the inflammatory
response (17,18). However, it is unclear whether CT may
attenuate cerebral ischemia/reperfusion injury and its
anti-apoptosis mechanism. Therefore, the present study aimed to
investigate the effects of CT and mechanisms on OGD/R-induced PC12
cell injury.
Materials and methods
Drugs and reagents
CT (cat. no. 447-43-0) was provided by the PUSH
Bio-Technology. Fetal bovine serum and Dulbecco's modified Eagle's
medium (DMEM) were offered by Gibco (Thermo Fisher Scientific,
Inc.). Streptomycin and penicillin were purchased from HyClone (GE
Healthcare Life Sciences). Nimodipine injection (cat. no. 12301323)
was provided by Bayer Schering Pharma AG. Lactate dehydrogenase
(LDH) assay kit (cat. no. 20090723), total protein extraction (cat.
no. 20150323) and BCA protein quantification kits (kit no.
20150323) were provided by Nanjing Jiancheng Bioengineering
Institute. ZSGB-BIO supplied horseradish peroxidase-conjugated goat
anti-rabbit IgG secondary antibody (cat. no. ZB-2301). Primary
antibodies against α-tubulin (cat. no. 2148), Apaf-1 (cat. no.
8723), Bcl-2 (cat. no. 2876) and Bax (cat. no. 2772) were provided
by Cell Signaling Technology, Inc. Antibodies against procaspase-9
(cat. no. ab2013), procaspase-7 (cat. no. ab25900),
cleaved-caspase-3 (cat. no. ab32042) and procaspase-3 (cat. no.
ab44976) were provided by Abcam. Cleaved-caspase-7 (cat. no.
AF4203), cleaved-caspase-9 (cat. no. AF5240) were supplied by
Affinity Biosciences.
Molecular docking
The mol2, a molecular structure recording format
designed by Sybyl molecular simulation software (version no. 2.1.1;
Certara), the mol2 format of CT was downloaded from PubChem
(http://pubchem.ncbi.nlm.nih.gov). CT and
its interacting protein crystals (the structure is a protein
complex with an inhibitor) were introduced into the Maestro 11.1
software ligprep module (Schrödinger, Inc.). The energy
optimization of CT in the first field (OPLS-2005) was minimized.
The structures of caspase-9, caspase-3 and caspase-7 with
endogenous ligands were obtained from the Protein Data Bank. Using
the Maestro 11.1 software Glide module, the target protein is
modified, dehydrated and hydrogenated under default parameters. The
active site of docking is generated by centering on the original
ligand. Finally, CT is molecularly docked with the target protein
(Fig. 2).
Cell culture
The PC12 cells were obtained from the rat adrenal
medulla of pheochromocytoma and have been widely used as an in
vitro cellular model of neurological diseases and cell signal
transduction pathways due to its sympathetic neuron's physiological
characteristics. PC12 cells were provided by the Chinese Academy of
Sciences Shanghai Cell Biology Institute. PC12 cells were cultured
in high-glucose DMEM supplemented with 5% fetal calf serum,
streptomycin (100 µg/ml), penicillin (100 U/ml) and 5%
CO2 at 37°C (19,20).
Drug treatment
A stock solution was prepared using dimethyl
sulfoxide, CT concentrations were set at 2.5, 5 and 10 µM according
to our preliminary experimental results. Nimodipine is a calcium
channel blocker, and it was used as a positive control drug in this
experiment at a concentration of 5 µM (21). Prior to the investigation, the cells
were inoculated in culture plates at a density of
1.0×105 cells/ml. After 12 h of adhering, to initiate
oxygen-glucose deprivation (OGD) by replacing the cell culture
medium with the glucose-free medium, the cells were incubated for 2
h in an oxygen-free chamber (5% CO2; 95% N2)
at 37°C. At the end of the OGD period, the cells were incubated
under normal growth conditions (5% CO2 and 95%
O2) for 24 h to achieve reperfusion (R) (22). Cells in the experimental groups were
treated with CT (2.5, 5 or 10 µM) and Nimodipine (5 µM) during the
entire period of OGD/R.
Morphological changes in cells
Following OGD/R, the cells were washed three times
with PBS, and the inverted microscope (Olympus Corporation;
TH4-200; magnification, ×200) was used to capture the images.
CCK-8 assay and LDH release assay
A CCK-8 assay measured the cell viability, according
to the manufacturer's protocols. PC12 cells were cultured in
96-well plates (1×104 cells/well) and, following OGD/R,
CCK-8 (10 µl per well) was added, and cells were incubated for 4 h
at 37°C. The OD value at the wavelength of 450 nm was detected by
an enzyme labeling instrument.
Cytotoxicity was assessed by measuring the level of
LDH in the culture medium. Following reperfusion, the medium was
collected for LDH level measurement. The LDH level was measured by
spectrophotometry determination at 440 nm, according to the
manufacturer's protocol. The release of LDH reflects the degree of
cell damage.
Flow cytometric apoptosis assay
The percentage of the apoptotic cells was determined
using an Annexin V-FITC/PI kit (Nanjing KeyGen Biotech Co., Ltd.).
The PC12 cells were cultured in 6-well plates (1×106
cells/well), washed twice with ice-cold PBS following treatment,
collected by trypsinization without EDTA. Next, cells were
suspended in 400 µl 1X Annexin V and then Annexin V-FITC staining
fluid 5 µl was added in the dark, at 4°C for 15 min, prior to 10 µl
PI staining fluid being added at 4°C for 5 min. Finally, the cells
were analyzed by flow cytometry (BD Biosciences) using BD Accuri C6
software (version no. 1.0.264.21; BD Biosciences) The total
apoptosis rate is equal to the early apoptotic rate plus the dead
cells rate.
Mitochondrial membrane potential (MMP)
measurement
Disruption of the MMP is one of the earliest
intracellular changes following apoptosis induction (19). It has been reported that the
decrease in MMP is associated with the apoptosis of numerous types
of cells (23). JC-1 is a probe
commonly used to detect the MMP of cells. When the MMP is high,
JC-1 may accumulate in the mitochondrial matrix and form polymers
to generate red fluorescence. When the MMP is low, the monomer
state glows green. Following reperfusion, the cells were washed
twice with PBS. Next, the cells were pretreated and stained
according to the method of the MMP detection kit (Nanjing KeyGen
Biotech Co., Ltd.). The fluorescence microscope (IX-73; Olympus
Corporation; magnification, ×200) detected the intensity of
fluorescence in each group.
Intracellular [Ca2+]
measurement
Fluo-3 AM is the most commonly used for the
detection of intracellular [Ca2+]. Following
reperfusion, the PC12 cells were washed twice with PBS and
incubated with 5 µM/l Fluo-3/AM staining solution at 37°C for 30
min in the dark. Subsequently, the cells were washed twice with
PBS. Next, with a fluorescence microscope (IX-73; Olympus
Corporation; magnification, ×200) to observe the fluorescence
intensity.
Western blot analysis
The protein expression of Apaf-1, cleaved-caspase-7,
procaspase-7, cleaved-caspase-3, procaspase-3, cleaved-caspase-9,
procaspase-9, Bcl-2 and Bax in the cells following OGD/R injury
were detected by Western blotting. Following treatment, cells were
collected and cell lysates were prepared by incubation in RIPA
buffer containing a protease inhibitor cocktail (Beijing Leagene
Biotech Co., Ltd.) according to the manufacturer's protocols.
Following protein concentration estimation using a BCA protein
quantitation assay kit (Nanjing KeyGen Biotech Co., Ltd.), equal
amounts (30 µg) of proteins were separated via 10% SDS-PAGE, prior
to being transferred to a PVDF membrane. The membranes were blocked
with 5% skimmed dry milk in tris-buffer solution (TBST) for 2 h at
room temperature. Membranes were incubated with primary antibodies
for overnight at 4°C, and the primary antibodies used were as
follows: Bcl-2 (dilution, 1:1,000), Bax (dilution, 1:300), Apaf-1
(dilution, 1:300), procaspase-9 (dilution, 1:1,000), procaspase-7
(dilution, 1:500), procaspase-3 (dilution, 1:500),
cleaved-caspase-3 (dilution, 1:500), cleaved-caspase-7 (dilution,
1:500), cleaved-caspase-9 (dilution, 1:500) and α-tubulin
(dilution, 1:1,000). Next, labelled membranes were washed with TBST
three times, and the membrane was probed with an HRP-labeled
secondary antibody (dilution, 1:2,000; BIOSS) for 1 h at room
temperature. The bands were then visualized using the Amersham
Imager 600 imaging system (GE Healthcare Life Sciences).
Immunofluorescence detection
Immunofluorescence was detected to investigate the
expression of Bcl-2 and Bax. Following reperfusion, the cells were
washed 3 times with PBS, then post-fixed in 4% paraformaldehyde at
room temperature for 20 min and incubated in 0.3% Triton X-100 for
a further 20 min. Cells were washed with PBS and blocked with 1%
BSA at 37°C for 2 h. Next, Bcl-2 (dilution, 1:100) and Bax
(dilution, 1:100) antibodies were added and incubated overnight.
Cells were washed with PBS, fluorescent second antibody (dilution,
1:500) was added and incubated for 2 h in the dark at room
temperature, the cells were washed 3 times with PBS, and DAPI
(dilution, 1:500) was added for 30 min. Finally, cells were viewed
using a fluorescence microscope (Olympus IX71; Olympus Corporation;
magnification, ×100).
Statistical analysis
The data were analyzed by Image Proplus and SPSS
17.0 software (SPSS, Inc.). Data are presented as the mean ±
standard deviation, and the changes in variable parameters were
analyzed by one-way analysis of variance, followed by the Dunnett's
test. P<0.05 was considered to indicate a statistically
significant difference.
Results
Results of molecular docking
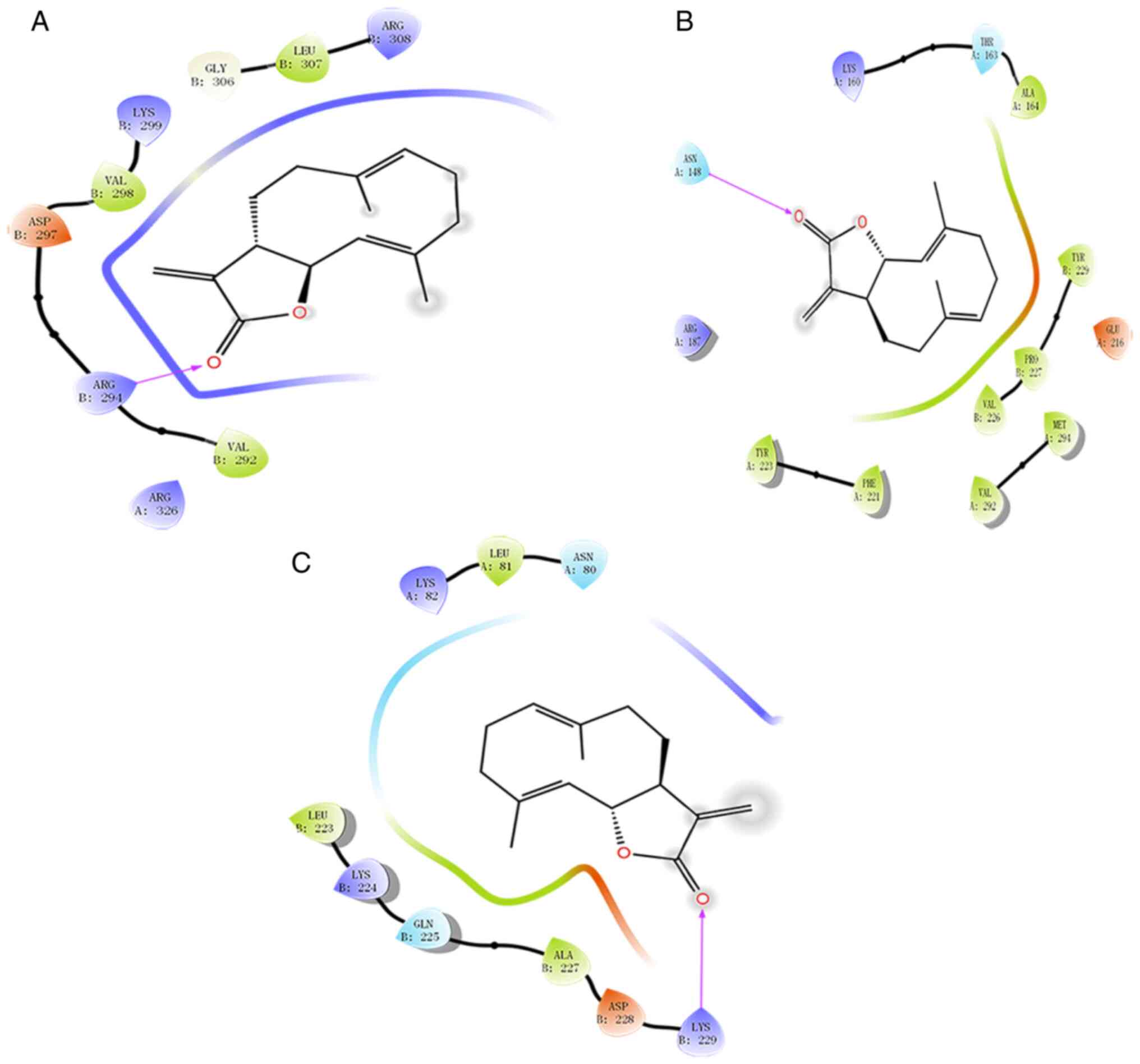
Molecular docking was used to predict the probable
targets of CT. The docking results revealed a high affinity of CT
towards Caspase-9, Caspase-3 and Caspase-7 (Fig. 2). Caspase-9 showed interactions
between CT and Gly306, Leu307, Arg308, Val292, Arg294, Asp297,
Val298 and Lys299 forming a hydrogen bond effect. Specifically, CT
and an active site with residues Arg294 forms a hydrogen bond
effect. Caspase-7 was interacting with CT through Val226, Pro227,
Tyr229, Lys160, Thr163, Ala164, Tyr223 and Phe221. Specifically, CT
and an active site with residues, Asn148, forms a hydrogen bond
effect. CT was interacting with the Lys82, Leu81, Asn80, Lbu223,
Lys224, Gln225, Ala227 and Asp228 residues of Caspase-3 form a
strong hydrophobic effect. Specifically, CT and an active site with
residues, Lys229, form a hydrogen bond effect.
Effect of CT on PC12 cell
morphological changes
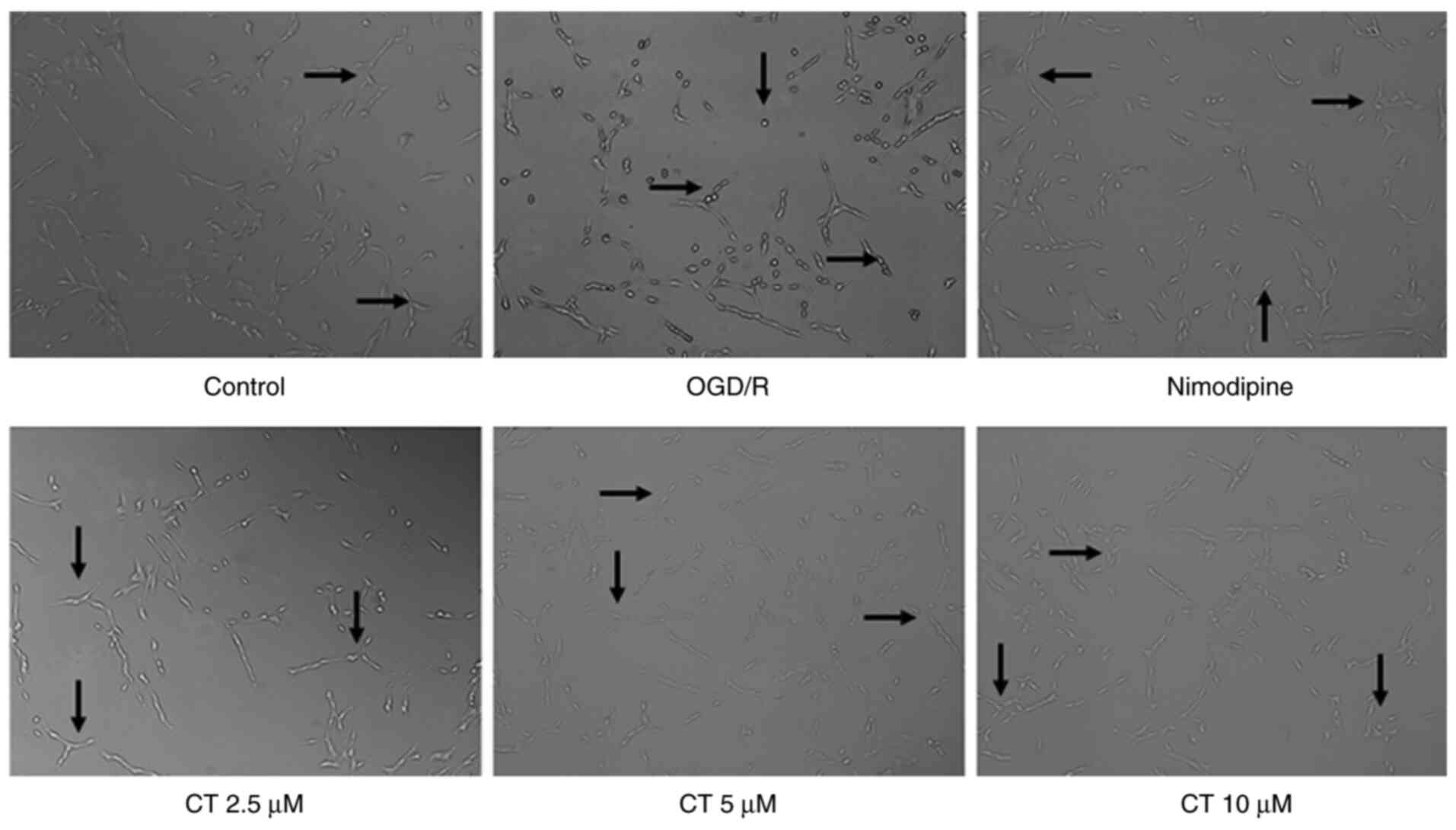
The results demonstrated that the number of PC12
cells significantly decreased following OGD/R treatment, certain
cells became round or floating, and the cells exhibited typical
swelling. Compared with the OGD/R group, in the OGD/R+CT group
(2.5, 5 and 10 µM), the cell body was relatively full, the membrane
was smooth and intact, and the adhesion was good (Fig. 3). The results indicated that CT has
a protective effect on PC12 cell injury induced by OGD/R.
Effect of CT on PC12 cell viability
and cell cytotoxicity
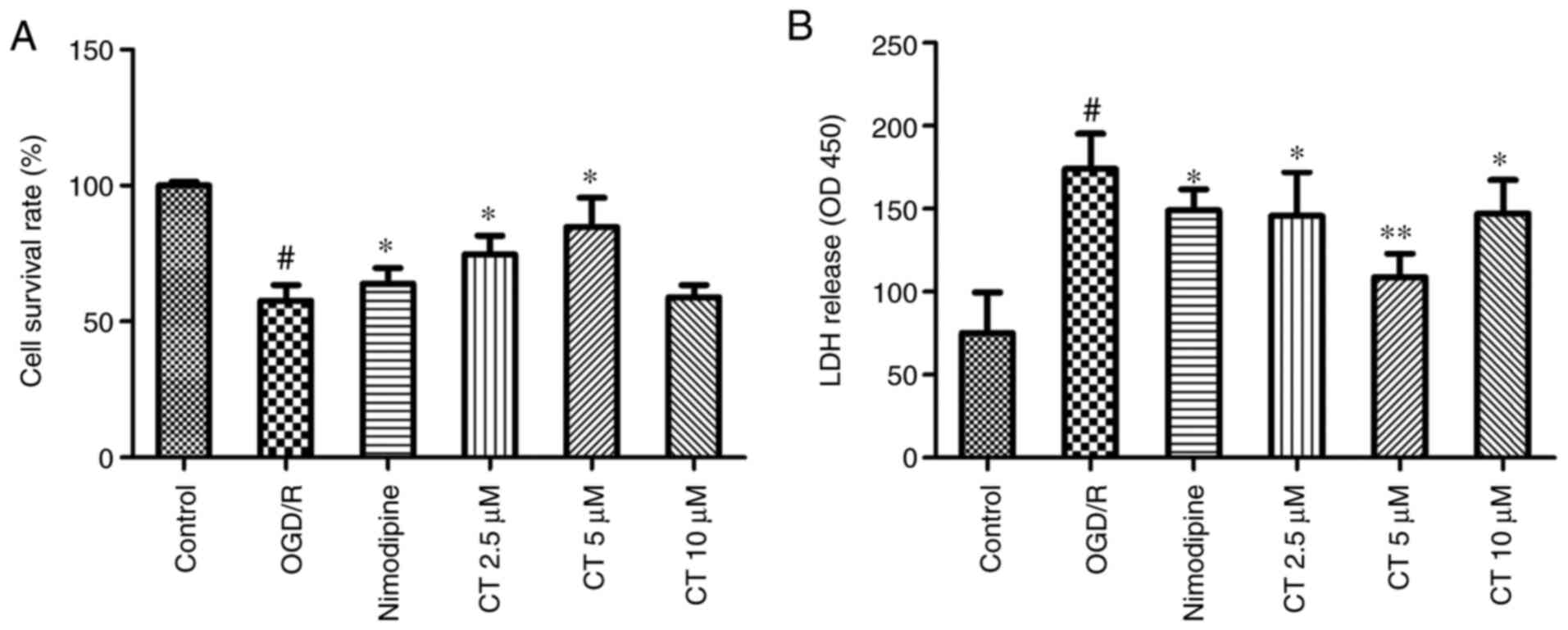
As shown in Fig. 4A,
PC12 cell viability was significantly decreased following the cell
being exposed to OGD for 2 h and reperfusion for 24 h. Following
treatment with CT (2.5 and 5 µM), PC12 cell viability was
significantly increased (P<0.05). LDH assays were performed as
shown in Fig. 4B, and it was
revealed that CT (2.5, 5 and 10 µM) significantly attenuated
OGD/R-induced LDH leakage.
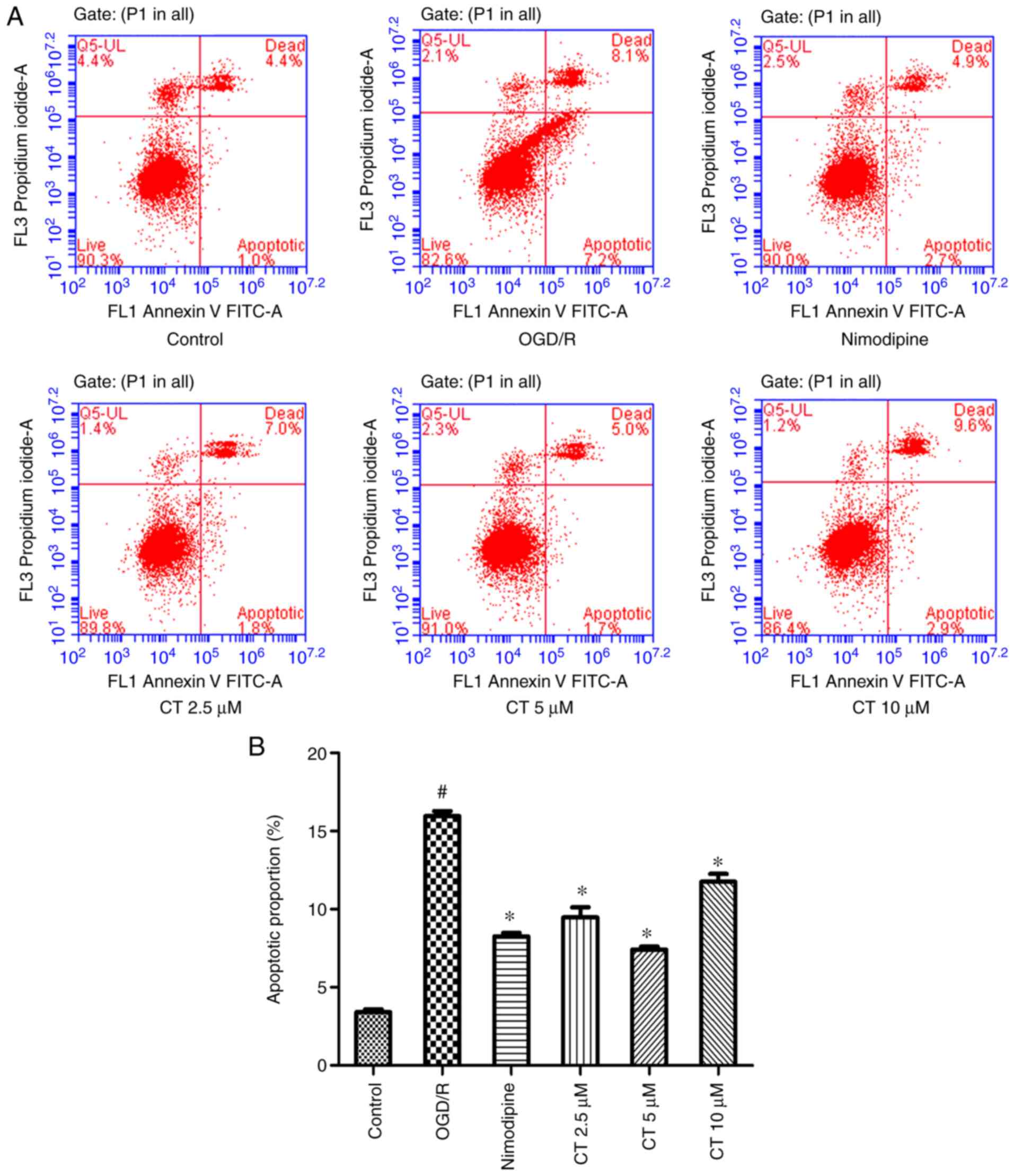
Effects of CT on apoptosis in PC12
cells
FITC-Annexin V/PI double staining was used with flow
cytometry to analyze the anti-apoptotic capacity of CT under OGD/R
conditions (Fig. 5). The results
demonstrated that the cell apoptotic rate was significantly
decreased following treatment with CT (2.5, 5 and 10 µM). The
apoptotic cell rate with CT (10 µM) was higher than CT (5 µM),
consistent with the effect of CT on the PC12 cell viability assay
following OGD/R injury.
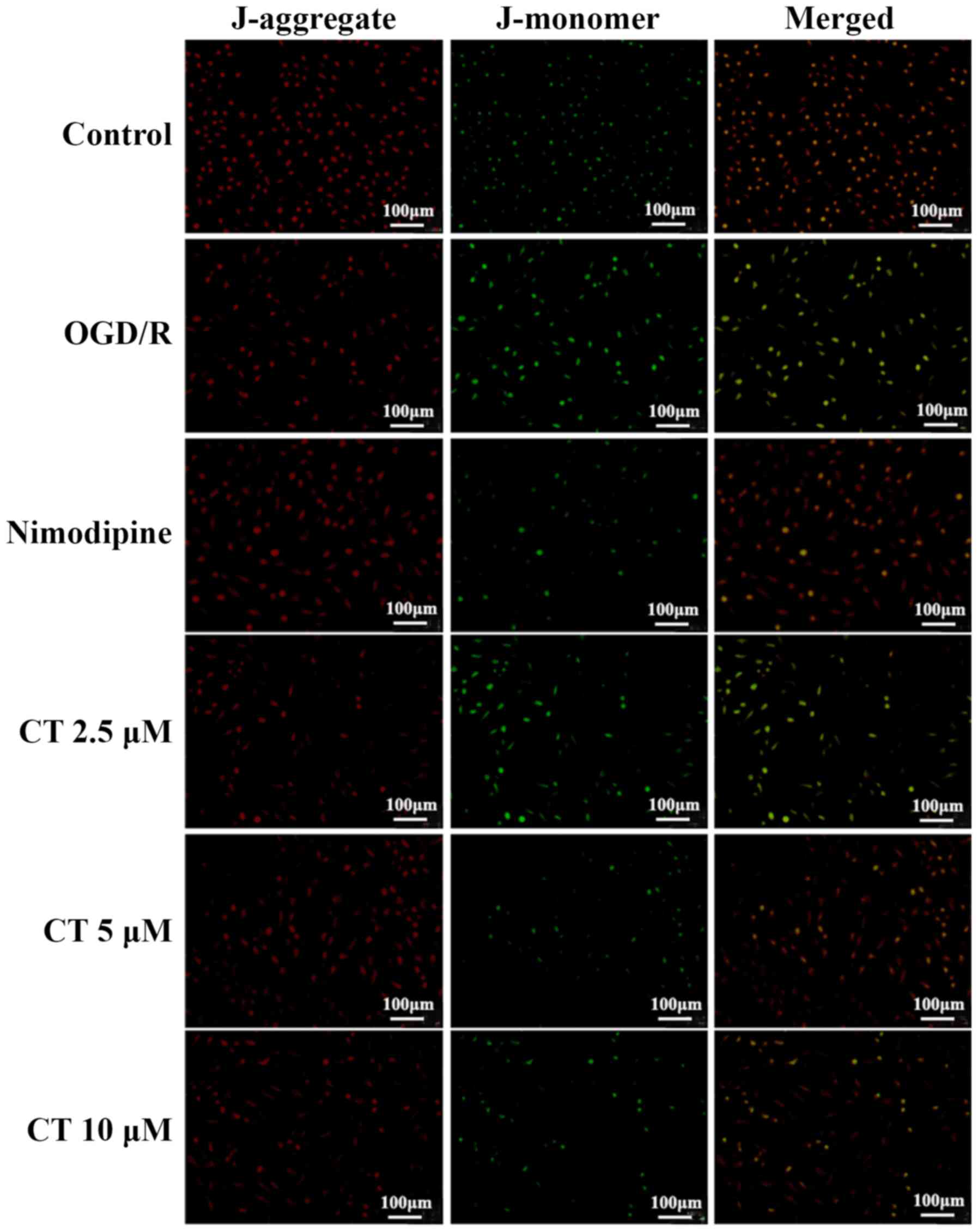
Effects of CT on MMP in PC12
cells
Fluorescence microscopy was used to detect MMP.
Following treatment with the CT (2.5, 5 and 10 µM), red
fluorescence increased in varying degrees, respectively, compared
with the OGD/R group (Fig. 6).
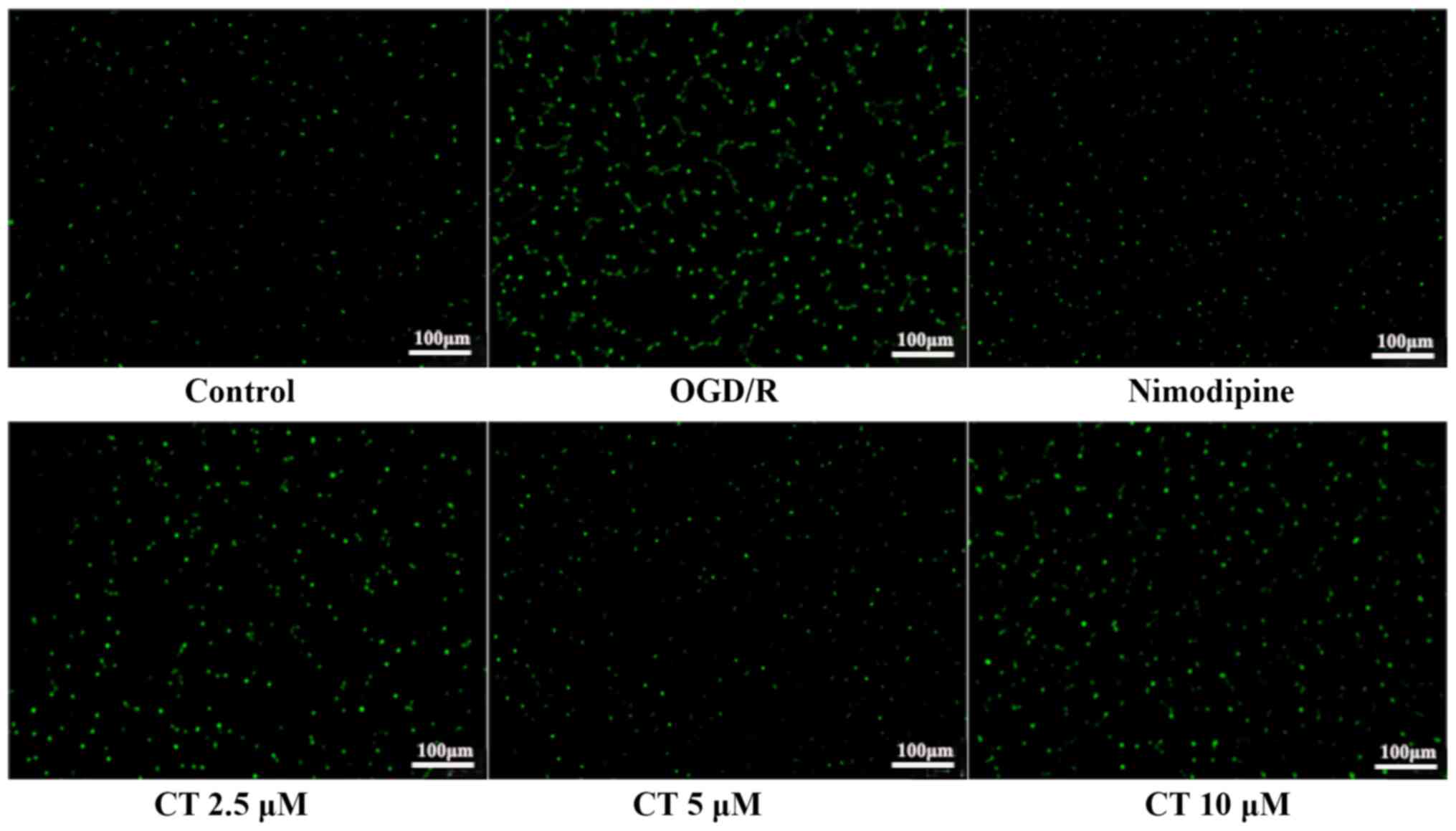
Effects of CT on intracellular
[Ca2+] in PC12 cells
As a second messenger, [Ca2+] regulates
signal transduction and neurotransmitter release (24,25).
Compared with the control group, the green fluorescence of the
OGD/R group was very strong, which indicated that OGD/R increased
the [Ca2+] concentration in PC12 cells. The green
fluorescence of cells treated with CT (2.5, 5 and 10 µM) was
decreased, suggesting that CT decreased intracellular
[Ca2+] concentration. The results demonstrated that CT
could inhibit the increase of intracellular [Ca2+]
induced by OGD/R injury (Fig.
7).
Effects of CT on the expression of
Apaf-1, cleaved-Caspase-3, Caspase-3, cleaved-Caspase-7, Caspase-7,
cleaved-Caspase-9, Caspase-9, Bax and Bcl-2 in PC12 cells following
OGD/R
The expression of Bax and Apaf-1 were significantly
increased following OGD/R. CT (2.5, 5 and 10 µM) inhibited the
expression of Bax and Apaf-1. Additionally, following treatment of
PC12 cells with OGD/R, the expression of Bcl-2 was decreased,
treatment with CT (2.5, 5 and 10 µM) significantly attenuated the
decrease in the Bcl-2 expression (Fig.
8).
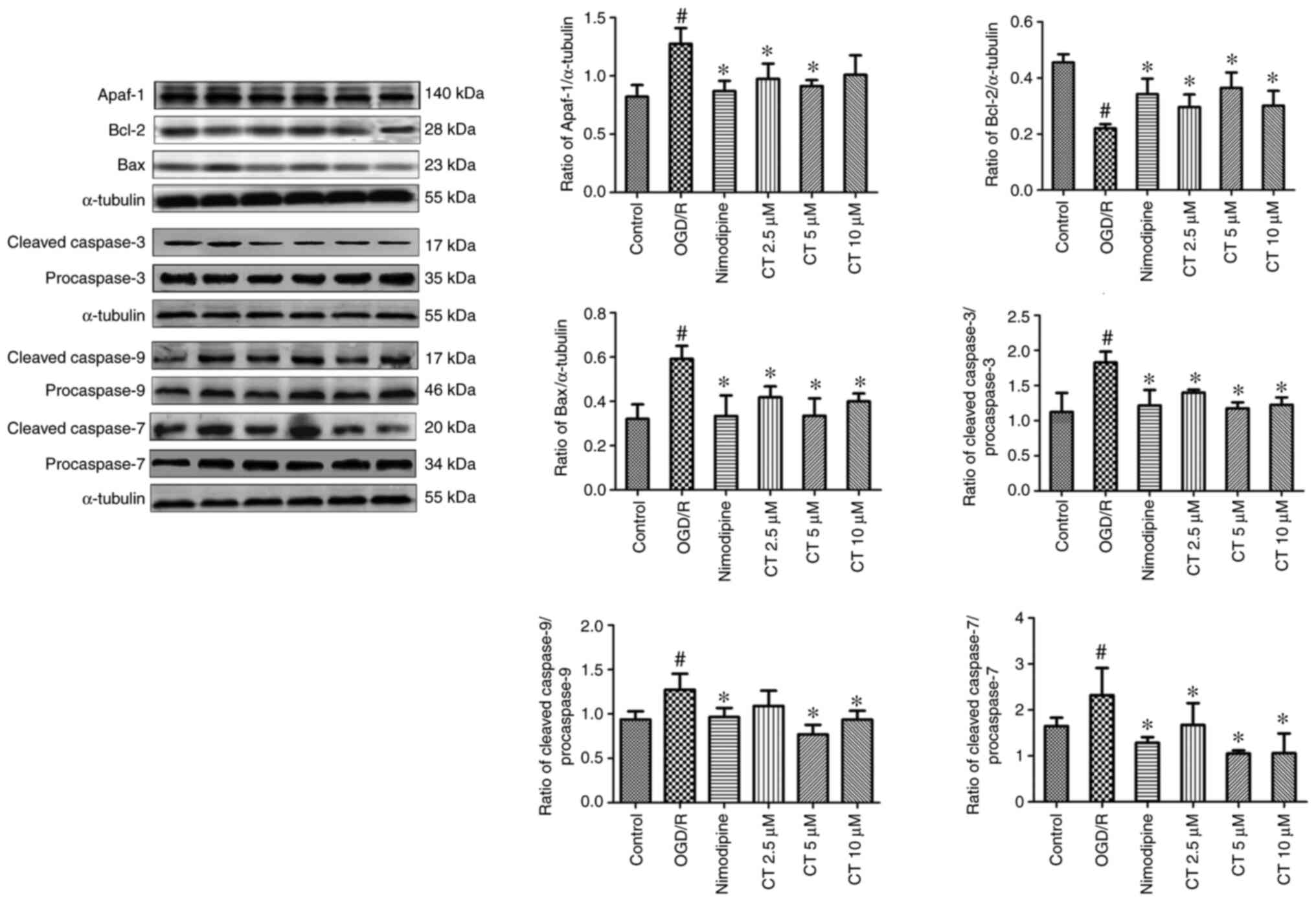 | Figure 8.Effects of CT on the expression of
Apaf-1, Bcl-2, Bax, cleaved-Caspase-9, cleaved-Caspase-7,
cleaved-Caspase-3 in PC12 cells following OGD/R. The stripe diagram
represents Western blot analysis of Apaf-1, Bcl-2, Bax,
cleaved-Caspase-9, cleaved-Caspase-7, cleaved-Caspase-3,
procaspase-9, procaspase-7 and procaspase-3. α-tubulin was usedd as
the loading control. The bar chart represents the quantitative
analysis of Apaf-1, Bcl-2, Bax, cleaved-Caspase-9/procaspase-9,
cleaved-Caspase-7/procaspase-7 and cleaved-Caspase-3/procaspase-3
expression. Values are presented as the mean ± standard deviation
(n=3). #P<0.05, compared with the control group;
*P<0.05, compared with OGD/R group. CT, costunolide; OGD/R,
oxygen-glucose deprivation/reperfusion. |
In the present study, the expression of
cleaved-Caspase-3, cleaved-Caspase-7, cleaved-Caspase-9,
procaspase-3, procaspase-7 and procaspase-9 was investigated. As
shown in Fig. 8, compared with the
control group, the expression of cleaved-Caspase-9,
cleaved-Caspase-3 and cleaved-Caspase-7 were significantly enhanced
following OGD/R. CT (2.5, 5 and 10 µM) may markedly inhibit the
expression of these proteins.
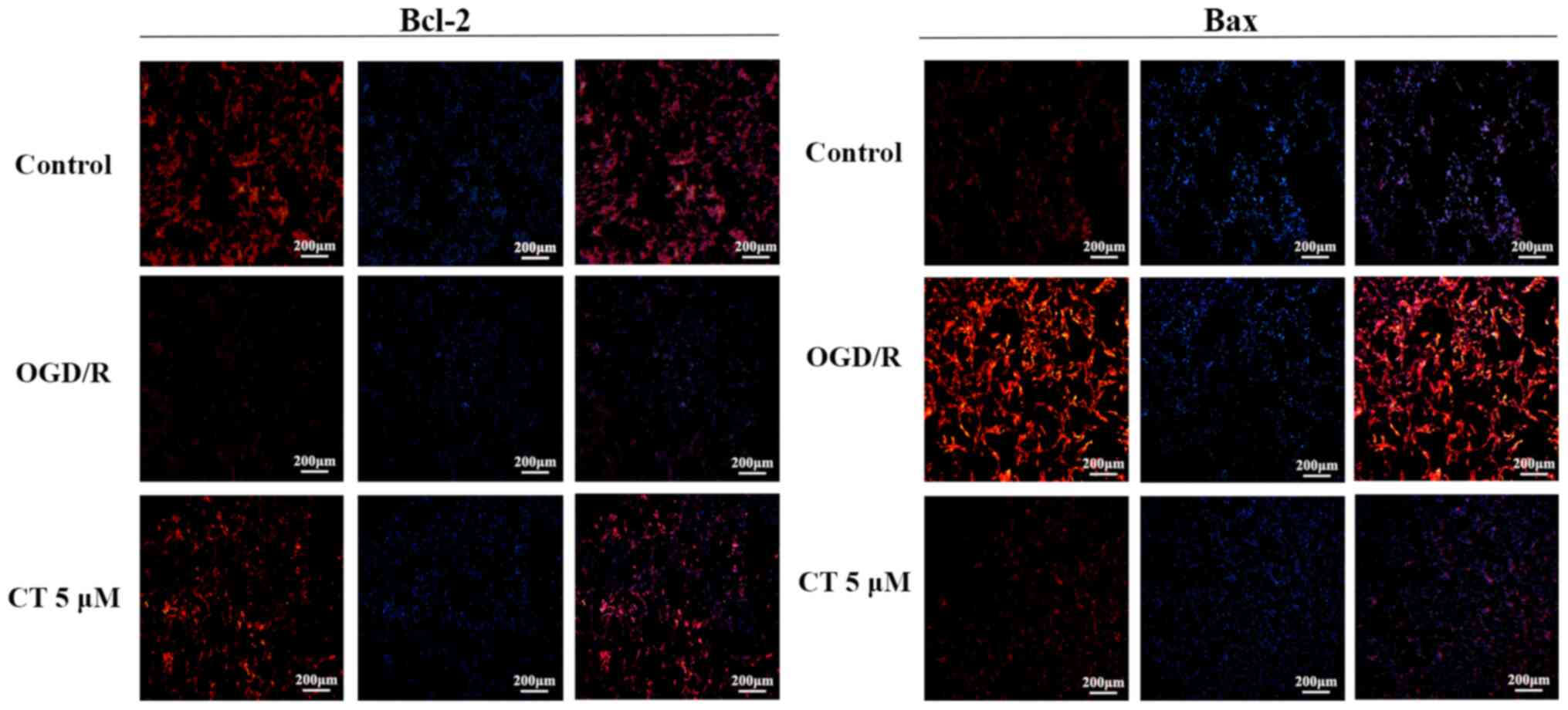
Detection of Bcl-2 and Bax in PC12
cells following OGD/R by cellular immunofluorescence
The expression of Bcl-2 protein was significantly
decreased following OGD/R. However, following the intervention with
CT (5 µM), red fluorescence was enhanced, which indicated that CT
may improve the expression of Bcl-2 (Fig. 9). As shown in Fig. 9, the expression of Bax was notably
increased following OGD/R compared with the control group. Compared
with the OGD/R group, red fluorescence became weaker, which
indicated that CT may inhibit the expression of Bax.
Discussion
Molecular docking is a drug design method that
follows the characteristics of the receptor, and the interaction
between the receptor and the drug molecule (26). Proteins may undergo conformational
transitions in physiological environments, and these transitions
may stabilize following ligand binding (27). Drugs are small compounds that act by
binding to or interacting with a protein kinase. The intermolecular
interactions between protein and medicines may be due to
hydrophobic interactions, electrostatic and van der Waals forces
(28). The results demonstrated
that CT contains binding sites with Caspase-9, Caspase-7 and
Caspase-3. It suggested that these proteins may be the target of CT
anti-apoptosis.
According to a previous study, the PC12 injury model
induced by OGD/R is often used to mimic cerebral
ischemia/reperfusion injury (29).
This model was used in the present study to investigate the effect
and mechanisms of CT. The results demonstrated that CT may protect
PC12 cells from OGD/R-induce injury by enhancing cell viability and
inhibiting LDH leakage.
Apoptosis plays an essential role in maintaining
metabolic balance and controlling neuron damage. Available evidence
has suggested that apoptosis serves a significant role in the
cerebral ischemia/reperfusion process (30,31).
The results of the present study demonstrated that the number of
apoptotic cells increased significantly following OGD/R. CT (2.5, 5
and 10 µM) significantly decreased apoptosis cells. However, the
anti-apoptotic effect of 10 µM CT was not as significant as that of
5 µM CT. In early experiments on the cytotoxicity of CT on PC12
cells, it was found that after the CT dose increased to 10 µM, it
became cytotoxic, which led to an increase in apoptosis. Therefore,
it may cause the apoptotic cell rate with 10 µM CT to be higher
than 5 µM CT, but the toxicity of 10 µM CT is minimal, and it has a
specific protective effect on PC12 cells following OGD/R injury.
Therefore, 5 µM CT has a better anti-apoptotic effect than 10 µM
CT. MMP is a sensitive indicator reflecting mitochondrial function.
The decrease in MMP leads to instability of neuronal cell
structure, thereby causing cell damage and ultimately results in
cell death (22). A recent study
has confirmed that OGD/R may cause intracellular [Ca2+]
overload and lead to mitochondrial dysfunction, resulting in
unstable neuronal cell structure and ultimately leading to neuronal
apoptosis (32). Following
treatment with OGD/R in PC12 cells, the MMP decreased, and the
intracellular [Ca2+] increased. However, CT decreased
intracellular [Ca2+] overload and enhanced MMP. These
results indicated that CT has a protective role in PC12 cell injury
induced by OGD/R.
A mitochondria-mediated apoptosis signalling pathway
is one of the main pathways in the apoptotic of cerebral
ischemia-reperfusion injury. The Bcl-2 protein family and its
members form an incredibly complex network of interactions that
regulate apoptosis. These proteins serve an essential role in
regulating and controlling apoptosis (33). Bcl-2 is an essential anti-apoptotic
protein that may stabilize mitochondrial membrane function and
prevent mitochondria release Cyto-c (34). Bax is a pro-apoptotic protein in the
cytoplasm under normal conditions. Bax increases the permeability
of the mitochondrial membrane, which leads to the release of Cyto-c
under certain stimulating conditions (35,36).
Next, in the presence of dATP, Cyto-c binds to Apaf-1 to catalyze
the activation of Caspase-9, and then activate downstream effector
Caspases, including Caspase-3, Caspase-6 and Caspase-7, which in
turn execute apoptosis by cleaving cellular proteins following
specific Asp residues (37). In the
mitochondria-mediated pathway, Caspase-3 is the main influencing
factor in the process of apoptosis, and its activation is a sign
that the cell is entering an irreversible stage of apoptosis
(15). In the present study, the
expression levels of Bcl-2 decreased, while the protein levels of
Bax, Apaf-1, cleaved-Caspase-9, cleaved-Caspase-3 and
cleaved-Caspase-7 significantly increased in OGD/R-treated PC12
cells. These results were consistent with those reported in the
literature (38,39). Reversal of these CT trends suggested
that the protective effect of CT may be associated with the
inhibition of mitochondrial-mediated apoptosis in PC12 cells.
Costunolide had a protective effect against
OGD/R-induced PC12 cell injury, and the mechanism may be associated
with the inhibition of mitochondrial-mediated apoptosis.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of Ningxia (grant no. 2020AAC02017) and
the National Natural Science Foundation of China (grant no.
81660700).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
LM, HM and JM established the PC12 cell injury model
induced by OGD/R, and were responsible for detecting cell
viability, LDH, [Ca2+] concentration, mitochondrial
membrane potential and protein expression. TL and HM analyzed and
interpreted the data. LM wrote the manuscript. YZ and QZ designed
the study, supervised the research group and revised the manuscript
critically for important intellectual content. YZ and QZ confirmed
the authenticity of all the raw data. The final version of the
manuscript was read and approved by all authors.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Katan M and Luft A: Global burden of
stroke. Semin Neurol. 38:208–211. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Nitzsche A, Poittevin M, Benarab A,
Philippe Bonnin P, Faraco G, Uchida H, Favre J, Garcia-Bonilla L,
Garcia MCL, Léger PL, et al: Endothelial S1P signaling counteracts
infarct expansion in ischemic stroke. Circ Res. 128:363–382. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Tu WJ, Zeng XW, Deng A, Zhao SJ, Luo DZ,
Ma GZ, Wang H and Liu Q: Circulating FABP4 (Fatty Acid-Binding
Protein 4) is a novel prognostic biomarker in patients with acute
ischemic stroke. Stroke. 48:1531–1538. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Brassai A, Suvanjeiev RG, Bán EG and
Lakatos M: Role of synaptic and nonsynaptic glutamate receptors in
ischaemia induced neurotoxicity. Brain Res Bull. 112:1–6. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Maier O, Menze BH, Von der Gablentz J,
Ḧani L, Heinrich MP, Liebrand M, Winzeck S, Basit A, Bentley P,
Chen L, et al: ISLES 2015-A public evaluation benchmark for
ischemic stroke lesion segmentation from multispectral MRI. Med
Image Anal. 35:250–269. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhao J, Bai Y, Zhang C, Zhang X, Zhang YX,
Chen J, Xiong L, Shi M and Zhao G: Cinepazide maleate protects PC12
cells against oxygen-glucose deprivation-induced injury. Neurol
Sci. 35:875–881. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhao QP, Chen AL, Wang XB, Zhang ZH, Zhao
YH, Huang Y, Ren SG and Zhu Y: Protective effects of
dehydrocostuslactone on rat hippocampal slice injury induced by
oxygen-glucose deprivation/reoxygenation. Int J Mol Med.
42:1190–1198. 2018.PubMed/NCBI
|
|
8
|
Li ZR, Yang L, Zhen J, Zhao Y and Lu ZN:
Nobiletin protects PC12 cells from ERS-induced apoptosis in OGD/R
injury via activation of the PI3K/AKT pathway. Exp Ther Med.
16:1470–1476. 2018.PubMed/NCBI
|
|
9
|
Meng X, Xie W, Xu Q, Liang T, Xu X, Sun G
and Sun X: Neuroprotective effects of radix scrophulariae on
cerebral ischemia and reperfusion injury via MAPK pathways.
Molecules. 23:24012018. View Article : Google Scholar
|
|
10
|
Kerr JF and Searle J: A suggested
explanation for the paradoxically slow growth rate of basal-cell
carcinomas that contain numerous mitotic figures. J Pathol.
107:41–44. 1972. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Green DR and Kroemer G: The
pathophysiology of mitochondrial cell death. Science. 305:626–629.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zamzami N, Hirsch T, Dallaporta B, Petit
PX and Kroemer G: Mitochondrial implication in accidental and
programmed cell death: Apoptosis and necrosis. J Bioenerg Biomembr.
29:185–193. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Song XF, Tian H, Zhang P and Zhang ZX:
Expression of Cyt-c-mediated mitochondrial apoptosis-related
proteins in rat renal proximal tubules during development. Nephron.
135:77–86. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Malladi S, Challa-Malladi M, Fearnhead HO
and Bratton SB: The Apaf-1*procaspase-9 apoptosome complex
functions as a proteolytic-based molecular timer. EMBO J.
28:1916–1925. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhao Q, Cheng X, Wang X, Wang J, Zhu Y and
Ma X: Neuroprotective effect and mechanism of Mu-Xiang-You-Fang on
cerebral ischemia-reperfusion injury in rats. J Ethnopharmacol.
192:140–147. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Cai H, He X and Yang C: Costunolide
promotes imatinib-induced apoptosis in chronic myeloid leukemia
cells via the Bcr/Abl-Stat5 pathway. Phytother Res. 32:1764–1769.
2018. View
Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chen Z, Zhang D, Li M and Wang B:
Costunolide ameliorates lipoteichoic acid-induced acute lung injury
via attenuating MAPK signaling pathway. Int Immunopharmacol.
61:283–289. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Saraswati S, Alhaider AA and Abdelgadir
AM: Costunolide suppresses inflammatory angiogenic response in a
subcutaneous murine sponge model. APMIS. 126:257–266. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang C, Li C, Chen S, Li Z, Jia X, Wang
K, Bao J, Liang Y, Wang X, Chen M, et al: Berberine protects
against 6-OHDA-induced neurotoxicity in PC12 cells and zebrafish
through hormetic mechanisms involving PI3K/AKT/Bcl-2 and Nrf2/HO-1
pathways. Redox Biol. 11:1–11. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang JF, Zhang L, Shi LL, Zhao ZH, Xu H,
Liang F, Li HB, Zhao Y, Xu X, Yang K and Tian YF: Parthenolide
attenuates cerebral ischemia/reperfusion injury via
Akt/GSK-3βpathway in PC12 cells. Biomed Pharmacother. 89:1159–1165.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang H, Wei W, Lan XB, Liu N, Li Y, Ma H,
Sun T, Peng X, Zhuang C and Yu J: Neuroprotective effect of
swertiamain on cerebral ischemia/reperfusion injury by inducing the
Nrf2 protective pathway. ACS Chem Neurosci. 10:2276–2286. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ma HX, Hou F, Chen AL, Li TT, Zhu YF and
Zhao QP: Mu-Xiang-You-Fang protects PC12 cells against
OGD/R-induced autophagy via the AMPK/mTOR signaling pathway. J
Ethnopharmacol. 252:1125832020. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Jangholi E, Sharifi ZN, Hoseinian M,
Zarrindast MR, Rahimi HR, Mowla A, Aryan H, Javidi MA, Parsa Y,
Ghaffarpasand F, et al: Verapamil inhibits mitochondria-induced
reactive oxygen species and dependent apoptosis pathways in
cerebral transient global ischemia/reperfusion. Oxid Med Cell
Longev. 2020:58726452020. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Farajdokht F, Mohaddes G, Karimi-Sales E,
Kafshdooz T, Mahmoudi J, Aberoumandi SM and Karimi P: Inhibition of
PTEN protects PC12 cells against oxygen-glucose deprivation induced
cell death through mitoprotection. Brain Res. 1692:100–109. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Seta K, Kim HW, Ferguson T, Kim R,
Pathrose P, Yuan Y, Lu G, Spicer Z and Millhorn DE: Genomic and
physiological analysis of oxygen sensitivity and hypoxia tolerance
in PC12 cells. Ann N Y Acad Sci. 971:379–388. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kuntz ID, Blaney JM, Oatley SJ, Langridge
R and Ferrin TE: A geometric approach to macromolecule-ligand
interactions. J Mol Biol. 161:269–288. 1982. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Overington JP, Bissan AL and Hopkins AL:
How many drug targets are there? Nat Rev Drug Discov. 5:993–996.
2006. View
Article : Google Scholar : PubMed/NCBI
|
|
28
|
Naqvi AAT, Mohammad T, Hasan GM and Hassan
MI: Advancements in docking and molecular dynamics simulations
towards ligand-receptor interactions and structure-function
relationships. Curr Top Med Chem. 8:1755–1768. 2018. View Article : Google Scholar
|
|
29
|
Feng LY, Gao JM, Liu YG, Shi JS and Gong
Q: Icariside II alleviates oxygen-glucose deprivation and
reoxygenation-induced PC12 cell oxidative injury by activating
Nrf2/SIRT3 signaling pathway. Biomed Pharmacother. 103:9–17. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Broughton BR, Reutens DC and Sobey CG:
Apoptotic mechanisms after cerebral ischemia. Stroke. 40:e331–e339.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhu JR, Tao YF, Lou S and Wu ZM:
Protective effects of ginsenoside Rb(3) on oxygen and glucose
deprivation-induced ischemic injury in PC12 cells. Acta Pharmacol
Sin. 31:273–280. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
de Pablo Y, Nilsson M, Pekna M and Pekny
M: Intermediate filaments are important for astrocyte response to
oxidative stress induced by oxygen-glucose deprivation and
reperfusion. Histochem Cell Biol. 140:81–91. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zheng YQ, Liu JX, Li XZ, Xu L and Xu YG:
RNA interference-mediated downregulation of Beclin1 attenuates
cerebral ischemic injury in rats. Acta Pharmacol Sin. 30:919–927.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Adams KW and Cooper GM: Rapid turnover of
mcl-1 couples translation to cell survival and apoptosis. J Biol
Chem. 282:6192–6200. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kim J, Parrish AB, Kurokawa M, Matsuura K,
Freel CD, Andersen JL, Johnson CE and Kornbluth S: Rsk-mediated
phosphorylation and 14-3-3βbinding of Apaf-1 suppresses cytochrome
c-induced apoptosis. EMBO J. 31:1279–1292. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Liu X, Zhu X, Chen M, Ge Q, Shen Y and Pan
S: Resveratrol protects PC12 cells against OGD/R-induced apoptosis
via the mitochondrial-mediated signaling pathway. Acta Biochim
Biophys Sin (Shanghai). 48:342–353. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Boatright KM and Salvesen GS: Mechanisms
of caspase activation. Curr Opin Cell Biol. 15:725–731. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Guo H, Chen L, Cui H, Peng X, Fang J, Zuo
Z, Deng J, Wang X and Wu B: Research advances on pathways of
nickel-induced apoptosis. Int J Mol Sci. 17:10–19. 2015. View Article : Google Scholar
|
|
39
|
Haddad JJ: The role of Bax/Bcl-2 and
pro-caspase peptides in hypoxia/reperfusion-dependent regulation of
MAPKERK: Discordant proteomic effect of MAPK(p38). Protein Pept
Lett. 14:361–371. 2007. View Article : Google Scholar : PubMed/NCBI
|