Epilepsy is a common neurological disease that
affects >50 million people worldwide. Approximately 30% of
epileptic patients are resistant to various antiepileptic drugs and
eventually develop refractory epilepsy (1,2).
Although novel treatment strategies have been developed over the
past few years, certain cases of refractory epilepsy cannot be
controlled yet (3). In addition,
most available treatments are focused on decreasing seizures by
inhibiting the excitability of the nervous system (4). Therefore, the development of new
antiepileptogenic therapies that can resolve seizures is imminent
(5). Recent studies reported that
neuroinflammation plays an important role in the development of
epilepsy and abnormal neural circuits (6–11).
Further, anti-inflammatory therapies were reported to have possible
antiepileptogenic potential (12,13).
Therefore, uncovering the role of inflammatory reactions in
epilepsy can lead to the development of more effective and novel
antiepileptic drugs which can decrease the disability rate and
improve the quality of life in epileptic patients.
The central nervous system can be termed as an
‘immunity exemption zone’ because the blood-brain barrier restricts
the entry of various substances in the blood circulation (14–17).
However, accumulating evidence indicates that the immune exemption
of the central nervous system is relative (18,19).
Engelhardt et al (20)
reported that although immature T cells cannot pass through the
blood-brain barrier, activated T cells can directly attach to the
surface of the cerebral vascular endothelium and pass through the
blood-brain barrier in the direction of blood flow. After crossing
the blood-brain barrier, T cells can mediate inflammation in the
central nervous system (20).
The central nervous system has a special ‘immune
defense line’. Cells involved in the brain's inflammatory reaction
include the microglia, astrocytes and endothelial cells. Microglia
are the immune sentinels of the central nervous system; they can be
activated by injury stimuli and induce the corresponding
inflammatory reaction, which maintains homeostasis in the central
nervous system (21). The number of
microglia is relatively small, accounting for ~5% of the glial
population (22–24). Following infection or damage to the
central nervous system, the resting microglia acquire antigenicity,
their shape becomes extended, and they are activated into
macrophages to participate in the inflammatory reaction together
with T cells inoculated within the blood circulation (25). However, hyperactivated microglia are
important sources of pro-inflammatory factors as well as oxidative
stress, and can cause neurotoxicity.
Astrocytes are also important contributors to the
inflammatory reaction in the nervous system (26,27).
Astrocytes can play a role in neuronal migration movement, maintain
the potassium concentration in the central nervous system, regulate
neuronal excitability, and present antigens to autoreactive T
cells.
Microglia are activated leading to the production of
inflammatory factors following trauma or other injuries. Activated
microglia can release cytotoxic substances and cytokines (28). Further, tissue injury could lead to
the infiltration of circulating immune cells (29). Additionally, brain tissues are also
exposed to systemic inflammatory response, which further aggravates
the immune response and leads to secondary neuronal damage
(30).
Activation of the immune system and an excessive
inflammatory response play crucial roles in the development of
chronic seizures (31–33). Moreover, neuronal inflammation
associated with inflammatory cytokines signaling pathways may
trigger epileptogenesis (34). Of
note, patients diagnosed with relapsing remitting multiple
sclerosis with epilepsy showed more extensive cortical inflammation
compared with patients diagnosed with relapsing remitting multiple
sclerosis without epilepsy (35).
Therefore, examining the association between neuroinflammation and
epilepsy can help uncover the pathogenesis. Clinical and animal
studies suggested that the immune response is triggered during the
pathophysiology of epilepsy and that the inflammatory reaction
within the brain may be involved in the development of epilepsy
(36,37). Further, the dysregulation of
immunoinflammatory reactions during the pathological course of
epilepsy was associated with seizure-induced plasticity (10). Rana and Musto (37) reported that neuronal inflammation
generated by neural death and astrocytes proliferation was
associated with the microglial activation in damaged areas such as
the amygdala, piriform and hippocampus in a rat model of
lithium-pilocarpine-induced epilepsy (38). This dysfunction can facilitate the
occurrence of epileptic seizures or epilepsy-induced neuronal
damage (39). Indeed, inflammatory
reactions were found to increase the propensity for seizures,
change neuronal excitability, damage the blood-brain barrier, and
mediate neuronal apoptosis and synaptic remodeling by activating
intracellular signaling pathways (40). Seizures can activate microglia and
neurons in the brain, and produce a series of inflammatory
reactions without additional exogenous stimuli (32,33).
Consequently, microglia and neurons secrete large amounts of
pro-inflammatory factors and prostaglandins, and activate the
complement system, which ultimately promotes neuronal death and
synapse regeneration, leading to chronic spontaneous seizures
(10,39).
Noteworthy, adenosine triphosphate (ATP) was shown
to activate the sterile inflammatory process through interactions
with purinergic receptors (40).
The ATP-gated ionotropic P2X7 receptor (P2X7R) can mediate the
regulation of neuroinflammation and immune reactions in the central
nervous system (41). Neuronal
injury was reported to activate microglia and increase the
expression of P2X7R (42,43). Monif et al (44) reported that the overexpression of
P2X7R is sufficient to drive microglial activation. Moreover, the
Fas ligand derived from microglia exacerbates P2X7-mediated
microglial activation and triggers a vicious cycle of neuronal
death (45,46). Low concentrations of ATP act as
chemotactic agents for microglia recognition and migration to guide
them to the site of injury (47).
ATP-stimulated and P2X4R-mediated microglial activation might have
an initial protective effect. Activated microglia can remove
potentially necrotic cell debris and promote tissue repair, thereby
contributing to neuroprotection. Further, activated microglia
release neurotrophic factors through activated P2X4Rs and
contribute to neuronal survival (48). P2X7R-activated microglia in
neuron-microglia co-culture protect neurons from glutamate toxicity
primarily by releasing tumor necrosis factor (TNF)-α. The depletion
of microglia can lead to an increase in the levels of cytokines and
chemokines such as interleukin (IL)-1β, TNF-α, cytokine-induced
neutrophil chemoattractant 1 and monocyte chemoattractant protein-1
in the brain, which aggravates brain damage (49). In later stages of brain injury, ATP
can stimulate the overexpression of microglia P2X7R, leading to
microglia activation and proliferation, as well as cell death
(50). Over-activated microglia
upregulate the expression of surface immunomodulatory proteins and
release of neurotoxic proinflammatory factors such as IL-1β, IL-6,
and TNF-α, which can promote further activation of microglia.
Long-term inflammatory reactions result in neuronal death that
affects both healthy and damaged cells (51,52).
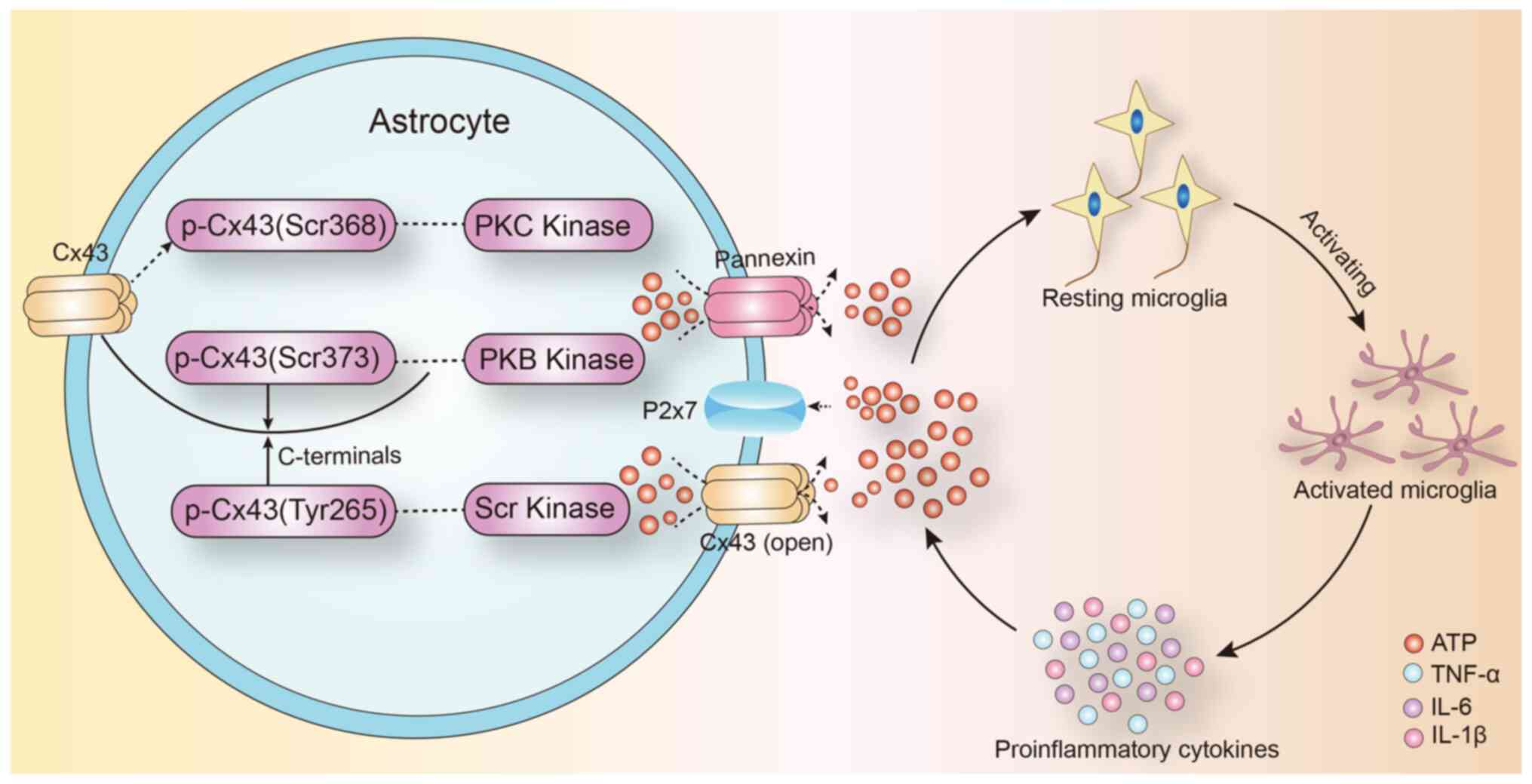
The expression of connexin 43 (Cx43) in astrocytes
is affected by inflammatory cytokines (53). Treatment with IL-1β, for 24 h,
inhibited the gap junction communication between the human
embryonic astrocytes and decreased Cx43 mRNA and protein expression
(54). IL-1β had a transient
inhibitory effect on gap junction communication between primary
astrocytes, and this inhibitory effect was produced through
p38-dependent signaling pathway (55). Therefore, Cx43-mediated gap junction
communication in astrocytes is closely correlated to the
inflammatory response in the central nervous system. They regulate
inflammation in the brain and selectively regulate the opening of
gap junction communication by intracranial inflammatory factors
(Fig. 1).
Astrocytes are the largest glial cell population in
the central nervous system. Cx43 is a gap junction protein that is
mainly expressed in astrocytes and mediates over 95% of the gap
junction communication in the brain (57). Under physiological conditions, gap
junctions allow the exchange of small molecules (<1.5 kDa)
between cells. ATP mediates the migration of activated microglia to
the injured area, especially in the initial phase of inflammation
(41). Further, extracellular ATP
induces the release of endogenous ATP from microglia and attracts
distant microglia to the injury site, which leads to the promotion
of the inflammatory cascade. ATP released by astrocyte hemichannels
establishes an ATP gradient in the extracellular environment that
can trigger microglial activation (58,59).
Increased extracellular ATP concentration in the injury site
mediates the activation of microglia around the lesion (60). Of note, the local injection of ATP
mimicked the traumatic brain injury-induced microglial activation
and the administration of the gap junction channel blocker,
carbenic acid, significantly inhibited the microglial activation
(61,62). Following injury, extracellular ATP
is released from the open hemichannels and mediates a rapid
reaction to microglial damage (63). Jesus et al (64) demonstrated that targeted knockout of
astrocyte Cx43 expression decreased proinflammatory cytokine levels
in the brain following lipopolysaccharide injection. In addition,
hemichannel modulators like Cx43 mimetic peptide and Cx43 antisense
oligonucleotides could inhibit the inflammatory response mediated
by microglial activation following spinal cord injury (65). Taken together, it is plausible that
astrocyte gap junction channel can act as a ‘switch’ in the
inflammatory signaling cascade by promoting the release of ATP into
the extracellular space. The inflammatory response can affect
neuronal excitability, neuronal apoptosis and synaptic remodeling.
These factors can lead to the development of abnormal neural
excitability, which contributes to the pathogenesis of epilepsy. At
the same time, pro-inflammatory cytokines can directly affect the
expression of Cx43 protein in astrocytes (Fig. 1).
The occurrence, development and maintenance of
epileptic seizures progress through a complicated process. The
neuroinflammatory reaction can aggravate epilepsy and maintain
recurrent episodes by increasing neuronal excitability, mediating
neuronal apoptosis and remodeling the synapses. Therefore,
controlling the neuroinflammatory reaction can mitigate the
downstream cascade. The gap junction pathway mediated by astrocyte
Cx43 can play a crucial role in controlling the epilepsy-induced
neuroinflammatory cascade. Therefore, Cx43 can be a potential
target for managing epileptic inflammatory reactions. Studies that
examine the correlation between neuroinflammation and gap junctions
will lead to a better understanding of epilepsy pathogenesis and
can uncover new treatment targets.
Not applicable.
This study was funded by the National Natural
Science Foundation of China (grant nos. 81801284, 81771396 and
31371125) and Jilin Provincial Ring-fenced Funding for Industrial
Innovation Project (grant no. 2017C029-1).
Not applicable.
CX and JX searched for literature; GW and JW wrote
this review; JL and XW revised this review. All authors read and
approved the final manuscript. Data authentication is not
applicable.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
|
1
|
Beghi E and Giussani G: Treatment of
epilepsy in light of the most recent advances. Lancet Neurol.
18:7–8. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Brewer MK, Grossman TR, McKnight TR,
Goldberg YP, Landy H and Gentry MS: The 4th International Lafora
Epilepsy Workshop: Shifting paradigms, paths to treatment, and hope
for patients. Epilepsy Behav. 90:284–286. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Conway CR, Udaiyar A and Schachter SC:
Neurostimulation for depression in epilepsy. Epilepsy Behav.
88S:25–32. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Johnson EL: Seizures and epilepsy. Med
Clin North Am. 103:309–324. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Klein P, Dingledine R, Aronica E, Bernard
C, Blümcke I, Boison D, Brodie MJ, Brooks-Kayal AR, Engel J Jr,
Forcelli PA, et al: Commonalities in epileptogenic processes from
different acute brain insults: Do they translate? Epilepsia.
59:37–66. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Vezzani A, Balosso S and Ravizza T:
Neuroinflammatory pathways as treatment targets and biomarkers in
epilepsy. Nat Rev Neurol. 15:459–472. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hodges SL and Lugo JN: Therapeutic role of
targeting mTOR signaling and neuroinflammation in epilepsy.
Epilepsy Res. 161:1062822020. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Eastman CL, D'Ambrosio R and Ganesh T:
Modulating neuroinflammation and oxidative stress to prevent
epilepsy and improve outcomes after traumatic brain injury.
Neuropharmacology. 172:1079072020. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Terrone G, Balosso S, Pauletti A, Ravizza
T and Vezzani A: Inflammation and reactive oxygen species as
disease modifiers in epilepsy. Neuropharmacology. 167:1077422020.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sanz P and Garcia-Gimeno MA: Reactive glia
inflammatory signaling pathways and epilepsy. Int J Mol Sci.
21:40962020. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Mukherjee S, Arisi GM, Mims K,
Hollingsworth G, O'Neil K and Shapiro LA: Neuroinflammatory
mechanisms of post-traumatic epilepsy. J Neuroinflammation.
17:1932020. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Korff CM and Dale RC: The immune system in
pediatric seizures and epilepsies. Pediatrics. 140:e201635342017.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Bauer J, Becker AJ, Elyaman W, Peltola J,
Rüegg S, Titulaer MJ, Varley JA and Beghi E: Innate and adaptive
immunity in human epilepsies. Epilepsia. 58 (Suppl 3):57–68. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Tamura R, Yoshida K and Toda M: Current
understanding of lymphatic vessels in the central nervous system.
Neurosurg Rev. 43:1055–1064. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kipnis J: Multifaceted interactions
between adaptive immunity and the central nervous system. Science.
353:766–771. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Klein RS, Garber C and Howard N:
Infectious immunity in the central nervous system and brain
function. Nat Immunol. 18:132–141. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wesselingh R, Butzkueven H, Buzzard K,
Tarlinton D, O'Brien TJ and Monif M: Innate immunity in the central
nervous system: A missing piece of the autoimmune encephalitis
puzzle? Front Immunol. 10:20662019. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Peralta Ramos JM, Iribarren P, Bousset L,
Melki R, Baekelandt V and Van der Perren A: Peripheral inflammation
regulates CNS immune surveillance through the recruitment of
inflammatory monocytes upon systemic α-synuclein administration.
Front Immunol. 10:802019. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Prinz M and Priller J: The role of
peripheral immune cells in the CNS in steady state and disease. Nat
Neurosci. 20:136–144. 2017. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Engelhardt B, Vajkoczy P and Weller RO:
The movers and shapers in immune privilege of the CNS. Nat Immunol.
18:123–131. 2017. View
Article : Google Scholar : PubMed/NCBI
|
|
21
|
Greenhalgh AD, Zarruk JG, Healy LM, Baskar
Jesudasan SJ, Jhelum P, Salmon CK, Formanek A, Russo MV, Antel JP,
McGavern DB, et al: Peripherally derived macrophages modulate
microglial function to reduce inflammation after CNS injury. PLoS
Biol. 16:e20052642018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Bohlen CJ, Friedman BA, Dejanovic B and
Sheng M: Microglia in brain development, homeostasis, and
neurodegeneration. Annu Rev Genet. 53:263–288. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Tan YL, Yuan Y and Tian L: Microglial
regional heterogeneity and its role in the brain. Mol Psychiatry.
25:351–367. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Illes P, Rubini P, Ulrich H, Zhao Y and
Tang Y: Regulation of microglial functions by purinergic mechanisms
in the healthy and diseased CNS. Cells. 9:11082020. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hsu M, Rayasam A, Kijak JA, Choi YH,
Harding JS, Marcus SA, Karpus WJ, Sandor M and Fabry Z:
Neuroinflammation-induced lymphangiogenesis near the cribriform
plate contributes to drainage of CNS-derived antigens and immune
cells. Nat Commun. 10:2292019. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wheeler MA, Jaronen M, Covacu R, Zandee
SEJ, Scalisi G, Rothhammer V, Tjon EC, Chao CC, Kenison JE, Blain
M, et al: Environmental control of astrocyte pathogenic activities
in CNS inflammation. Cell. 176:581–596.e18. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Rothhammer V and Quintana FJ: Control of
autoimmune CNS inflammation by astrocytes. Semin Immunopathol.
37:625–638. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Anderson WD, Greenhalgh AD, Takwale A,
David S and Vadigepalli R: Novel influences of IL-10 on CNS
inflammation revealed by integrated analyses of cytokine networks
and microglial morphology. Front Cell Neurosci. 11:2332017.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chiang N and Serhan CN: Structural
elucidation and physiologic functions of specialized pro-resolving
mediators and their receptors. Mol Aspects Med. 58:114–129. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Dupuis N and Auvin S: Inflammation and
epilepsy in the developing brain: Clinical and experimental
evidence. CNS Neurosci Ther. 21:141–151. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Mukhtar I: Inflammatory and immune
mechanisms underlying epileptogenesis and epilepsy: From
pathogenesis to treatment target. Seizure. 82:65–79. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wyatt-Johnson SK and Brewster AL: Emerging
Roles for Microglial Phagocytic Signaling in Epilepsy. Epilepsy
Curr. 20:33–38. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Hiragi T, Ikegaya Y and Koyama R:
Microglia after seizures and in epilepsy. Cells. 7:262018.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Paudel YN, Shaikh MF, Shah S, Kumari Y and
Othman I: Role of inflammation in epilepsy and neurobehavioral
comorbidities: Implication for therapy. Eur J Pharmacol.
837:145–155. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Calabrese M, De Stefano N, Atzori M,
Bernardi V, Mattisi I, Barachino L, Rinaldi L, Morra A, McAuliffe
MM, Perini P, et al: Extensive cortical inflammation is associated
with epilepsy in multiple sclerosis. J Neurol. 255:581–586. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Drion CM, van Scheppingen J, Arena A,
Geijtenbeek KW, Kooijman L, van Vliet EA, Aronica E and Gorter JA:
Effects of rapamycin and curcumin on inflammation and oxidative
stress in vitro and in vivo - in search of potential
anti-epileptogenic strategies for temporal lobe epilepsy. J
Neuroinflammation. 15:2122018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Rana A and Musto AE: The role of
inflammation in the development of epilepsy. J Neuroinflammation.
15:1442018. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Jung KH, Chu K, Lee ST, Kim JH, Kang KM,
Song EC, Kim SJ, Park HK, Kim M, Lee SK, et al: Region-specific
plasticity in the epileptic rat brain: A hippocampal and
extrahippocampal analysis. Epilepsia. 50:537–549. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Vezzani A and Rüegg S: Introduction to the
2nd Meeting on Immunity and Inflammation in Epilepsy (IIE2016).
Epilepsia. 58 (Suppl 3):7–10. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Mazarati AM, Lewis ML and Pittman QJ:
Neurobehavioral comorbidities of epilepsy: Role of inflammation.
Epilepsia. 58 (Suppl 3):48–56. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Allsopp RC, Dayl S, Schmid R and Evans RJ:
Unique residues in the ATP gated human P2X7 receptor define a novel
allosteric binding pocket for the selective antagonist AZ10606120.
Sci Rep. 7:7252017. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Nuka E, Ohnishi K, Terao J and Kawai Y:
ATP/P2X7 receptor signaling as a potential anti-inflammatory target
of natural polyphenols. PLoS One. 13:e02042292018. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Zumerle S, Calì B, Munari F, Angioni R, Di
Virgilio F, Molon B and Viola A: Intercellular calcium signaling
induced by ATP potentiates macrophage phagocytosis. Cell Rep.
27:1–10.e4. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Monif M, Burnstock G and Williams DA:
Microglia: Proliferation and activation driven by the P2X7
receptor. Int J Biochem Cell Biol. 42:1753–1756. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Li R, Shang Y, Hu X, Yu Y, Zhou T, Xiong W
and Zou X: ATP/P2X7r axis mediates the pathological process of
allergic asthma by inducing M2 polarization of alveolar
macrophages. Exp Cell Res. 386:1117082020. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Ryoden Y, Fujii T, Segawa K and Nagata S:
Functional expression of the P2X7 ATP receptor requires eros. J
Immunol. 204:559–568. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Burnstock G and Knight GE: The potential
of P2X7 receptors as a therapeutic target, including inflammation
and tumour progression. Purinergic Signal. 14:1–18. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Amadio S, Parisi C, Piras E, Fabbrizio P,
Apolloni S, Montilli C, Luchetti S, Ruggieri S, Gasperini C,
Laghi-Pasini F, et al: Modulation of P2X7 receptor during
inflammation in multiple sclerosis. Front Immunol. 8:15292017.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Wei L, Syed Mortadza SA, Yan J, Zhang L,
Wang L, Yin Y, Li C, Chalon S, Emond P, Belzung C, et al:
ATP-activated P2X7 receptor in the pathophysiology of mood
disorders and as an emerging target for the development of novel
antidepressant therapeutics. Neurosci Biobehav Rev. 87:192–205.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Beamer E, Fischer W and Engel T: The
ATP-gated P2X7 receptor as a target for the treatment of
drug-resistant epilepsy. Front Neurosci. 11:212017. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Kaczmarek-Hajek K, Zhang J, Kopp R,
Grosche A, Rissiek B, Saul A, Bruzzone S, Engel T, Jooss T,
Krautloher A, et al: Re-evaluation of neuronal P2X7 expression
using novel mouse models and a P2X7-specific nanobody. Elife.
7:e362172018. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Kim Y, Davidson JO, Gunn KC, Phillips AR,
Green CR and Gunn AJ: Role of hemichannels in CNS inflammation and
the inflammasome pathway. Adv Protein Chem Struct Biol. 104:1–37.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Das A, Wallace GC IV, Holmes C, McDowell
ML, Smith JA, Marshall JD, Bonilha L, Edwards JC, Glazier SS, Ray
SK, et al: Hippocampal tissue of patients with refractory temporal
lobe epilepsy is associated with astrocyte activation,
inflammation, and altered expression of channels and receptors.
Neuroscience. 220:237–246. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
John GR, Scemes E, Suadicani SO, Liu JS,
Charles PC, Lee SC, Spray DC and Brosnan CF: IL-1beta
differentially regulates calcium wave propagation between primary
human fetal astrocytes via pathways involving P2 receptors and gap
junction channels. Proc Natl Acad Sci USA. 96:11613–11618. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Zvalova D, Cordier J, Mesnil M, Junier MP
and Chneiweiss H: p38/SAPK2 controls gap junction closure in
astrocytes. Glia. 46:323–333. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
De Bock M, Wang N, Decrock E, Bultynck G
and Leybaert L: Intracellular cleavage of the Cx43 C-terminal
domain by matrix-metalloproteases: A novel contributor to
inflammation? Mediators Inflamm. 2015:2574712015. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Jourdeuil K and Taneyhill LA: The gap
junction protein connexin 43 controls multiple aspects of cranial
neural crest cell development. J Cell Sci. 133:jcs2354402020.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Nikolic L, Shen W, Nobili P, Virenque A,
Ulmann L and Audinat E: Blocking TNFα-driven astrocyte purinergic
signaling restores normal synaptic activity during epileptogenesis.
Glia. 66:2673–2683. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Ghaemi A, Alizadeh L, Babaei S, Jafarian
M, Khaleghi Ghadiri M, Meuth SG, Kovac S and Gorji A:
Astrocyte-mediated inflammation in cortical spreading depression.
Cephalalgia. 38:626–638. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Webster KM, Sun M, Crack P, O'Brien TJ,
Shultz SR and Semple BD: Inflammation in epileptogenesis after
traumatic brain injury. J Neuroinflammation. 14:102017. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Illes P, Burnstock G and Tang Y:
Astroglia-derived ATP modulates CNS neuronal circuits. Trends
Neurosci. 42:885–898. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Nakamura Y, Park JH and Hayakawa K:
Therapeutic use of extracellular mitochondria in CNS injury and
disease. Exp Neurol. 324:1131142020. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Li W, Li J, Sama AE and Wang H:
Carbenoxolone blocks endotoxin-induced protein kinase R (PKR)
activation and high mobility group box 1 (HMGB1) release. Mol Med.
19:203–211. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Jesus LB, Santos AB, Jesus EEV, Santos
RGD, Grangeiro MS, Bispo-da-Silva A, Arruda MR, Argolo DS, Pinheiro
AM, El-Bachá RS, et al: IDO, COX and iNOS have an important role in
the proliferation of Neospora caninum in neuron/glia co-cultures.
Vet Parasitol. 266:96–102. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Kwan T, Floyd CL, Kim S and King PH: RNA
binding protein human antigen R is translocated in astrocytes
following spinal cord injury and promotes the inflammatory
response. J Neurotrauma. 34:1249–1259. 2017. View Article : Google Scholar : PubMed/NCBI
|