Introduction
Lung cancer is one of the most common causes of
cancer-related mortality, accounting for 18.4% of cancer-related
deaths worldwide (1). In total,
~85% of lung cancer cases are diagnosed as non-small cell lung
cancer (NSCLC) and ~15% of lung cancer cases are diagnosed as small
cell lung cancer (2). At present,
the most common treatment methods for lung cancer include surgical
resection, chemotherapy, radiotherapy, immunotherapy and targeted
drugs (3). Although targeted
therapies, such as EGFR inhibitors, and immunotherapy drugs, such
as anti-programmed cell death protein 1/programmed death-ligand 1,
have significantly improved the treatment of lung cancer, only a
small number of patients are eligible to be treated with targeted
therapies, thus the global 5-year survival rate of lung cancer
remains at ~20% (4–8). Therefore, identifying specific and
sensitive prognostic markers, therapeutic targets and devising
novel therapeutic drugs to improve the survival time and quality of
life of patients with lung cancer remains a priority.
The Warburg effect is defined by the altered
metabolism of glucose, whereby tumor cells produce large amounts of
lactate, even in the presence of oxygen and fully functioning
mitochondria (9). The intrinsic
mechanism of the Warburg effect is complex, and is associated with
the activation of numerous oncogenes, inactivation of numerous
tumor suppressor genes, the abnormal expression of glycosylase and
changes in the tumor microenvironment, amongst other factors
(10–12). However, the specific mechanism
requires further investigation. It is well established that the
cell death program (apoptosis) exerts an anticancer effect
(13). The Warburg effect is
closely associated with the occurrence and development of cancer,
as it provides a growth advantage to tumor cells, helping them to
escape apoptosis and create a suitable environment for tumor
metastasis to occur (14–16). The inhibition of the Warburg effect
promotes cell apoptosis and inhibits tumor proliferation and
metastasis (17). Several key
metabolic enzymes [pyruvate kinase M2 (PKM2), pyruvate
dehydrogenase kinase 1 (PDK1) and hexokinase 2] have been found to
play an important role in the Warburg effect (18–20).
The changes in key enzymes can lead to an enhanced glycolytic
ability, promote glucose uptake into tumor cells and increase the
accumulation of lactic acid, thus further supporting tumor growth
and development (21).
6-Phosphofructo-2-kinase/fructose-2,6-biphosphatase (PFKFB2) is an
enzyme that regulates the synthesis and degradation of
fructose-2,6-bisphosphate (Fru-2, 6-P2), which is widely expressed
in a variety of cancer cells, such as ovarian, breast and
pancreatic cancer cells (22,23).
By regulating the expression of Fru-2, 6-P2 (a signaling metabolite
that participates in glycolysis), the PFKFB family has been found
to regulate intracellular glycolysis in cancer by acting as an
oncogene (24,25).
Shikonin is a naphthalene quinone compound extracted
from Lithospermum erythrorhizon (an herbaceous plant), which
has demonstrated anti-inflammatory, antiviral, antitumor and wound
healing potential, amongst other biological activities (26). The present study aimed to
investigate the effects of shikonin on cell proliferation,
migration, invasion, apoptosis and aerobic glycolysis in lung
cancer cells, and further determined the potential underlying
molecular mechanisms of shikonin. The current findings may provide
a foundation for the clinical use of shikonin in the treatment of
lung cancer.
Materials and methods
Patient studies
A total of 20 NSCLC and adjacent normal tissues were
obtained from patients (10 women and 10 men; age range, 50–79
years) who underwent surgical resection at The Affiliated Hospital
of Qingdao University (Qingdao, China) between January 2019 and
December 2019. Patients who had received chemotherapy or
radiotherapy prior to surgery were excluded from the study.
Patients who were diagnosed by positron emission tomography/CT scan
and were eligible for radical surgery were included in the study.
All samples were stored at −80°C until required for RNA extraction.
The study protocols were approved by the Ethics Committee of The
Affiliated Hospital of Qingdao University, and written informed
consent was obtained from all patients prior to participation.
Cell lines and culture
The Beas-2B lung epithelial cell line and lung
cancer cell lines, A549 and H446, were obtained from The Cell Bank
of Type Culture Collection of The Chinese Academy of Sciences. All
cells were cultured at 37°C (5% CO2) in DMEM (Gibco;
Thermo Fisher Scientific, Inc.) supplemented with 10% FBS (Gibco;
Thermo Fisher Scientific, Inc.) and 1% penicillin/streptomycin
(Gibco; Thermo Fisher Scientific, Inc.).
Cell transfection
Small interfering RNA (siRNA)-negative control (NC)
(sense, 5′-UUCUCCGAACGUGUCACGUTT-3′ and antisense,
5′-ACGUGACACGUUCGGAGAATT-3′), siRNA-PFKFB2 (sense,
5′-AUUGUCAUGCCGAAAGAAGUC-3′ and antisense,
5′-CUUCUUUCGGCAUGACAAUGA-3′), pcDNA3.1-PFKFB2 and pcDNA3.1-NC (all
Shanghai GenePharma Co., Ltd.) were used to knockdown or
overexpress PFKFB2, respectively. Cells were transfected with siRNA
(30 nM) or overexpression vectors (1 µg) using
Lipofectamine® 2000 reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) for 48 h at 37°C. Cells were harvested 48 h
post-transfection for use in subsequent experiments.
Reverse transcription-quantitative PCR
(RT-qPCR)
Transfected or untransfected A549 and H446 cells
were treated with or without 50 µM shikonin (Shanghai Yuanye
Biotechnology Co., Ltd.) for 24 h at 37°C. Total RNA was extracted
from clinical specimens or cells using TRIzol® reagent
(Invitrogen; Thermo Fisher Scientific, Inc.), according to the
manufacturer's protocol. Total RNA was reverse transcribed into
cDNA using a PrimeScript RT kit (Takara Biotechnology Co., Ltd.),
according to the manufacturer's protocol. qPCR was subsequently
performed on an ABI 7500 Real-Time PCR system (Applied Biosystems;
Thermo Fisher Scientific, Inc.) using a SYBR® Premix Ex
Taq™ kit (Takara Biotechnology Co., Ltd.). The following
thermocycling conditions were used for the qPCR: Initial
denaturation at 95°C for 3 min; followed by 40 cycles at 95°C for 5
sec and 60°C for 30 sec. The following primers pairs (Sangon
Biotech Co., Ltd.) were used for the qPCR: PFKFB2 forward,
5′-GCTGCTTGGTGGGAGTGATAA-3′ and reverse,
5′-TGAGAAGCCAAGTGTCAGGG-3′; and β-actin forward,
5′-GAGGACCCTGGATGTGACAG-3′ and reverse, 5′-AAGACCTGTACGCCAACACA-3′.
Relative mRNA expression levels were quantified using the
2−ΔΔCq method (27) and
normalized to β-actin.
Western blotting
A549 and H446 cells were treated with shikonin [0
(control), 10, 20 or 50 µM] for 24 h at 37°C, then total protein
was extracted from cells using RIPA lysis buffer (Santa Cruz
Biotechnology, Inc.) supplemented with protease and phosphatase
inhibitor mixture. Total protein was quantified using a BCA assay
and 50 µg protein/lane was separated via 10% SDS-PAGE. The
separated proteins were subsequently transferred onto PVDF
membranes (EMD Millipore) and blocked with 5% skim milk for 2 h at
room temperature. The membranes were then incubated with the
following primary antibodies at 4°C overnight: Anti-PFKFB2
(1:1,000; cat. no. ab234865; Abcam), anti-PDK1 (1:2,000; cat. no.
ab202468; Abcam), anti-glucose transporter 1 (GLUT1; 1:200; cat.
no. ab150299; Abcam), anti-phosphoglycerate kinase 2 (PGK2;
1:1,000; cat. no. ab183031; Abcam), anti-lactate dehydrogenase A
(LDHA; 1:1,000; cat. no. ab101562; Abcam), anti-PKM2 (1:1,000; cat.
no. ab137852; Abcam), anti-GLUT3 (1:8,000; cat. no. ab41525;
Abcam), anti-pyruvate dehydrogenase (PDH; 1:1,000; cat. no. 3205;
Cell Signaling Technology, Inc.), phosphorylated (p)-PDH (1:1,000;
cat. no. 31866; Cell Signaling Technology, Inc.) and anti-GAPDH
(1:2,500; cat. no. ab9485; Abcam). Following the primary antibody
incubation, the membranes were incubated with an anti-rabbit
HRP-conjugated secondary antibody (1:5,000; cat. no. sc-2357; Santa
Cruz Biotechnology, Inc.) for 1 h at room temperature. Protein
bands were visualized using an enhanced chemiluminescence kit (EMD
Millipore).
Cell proliferation assay
Following transfection, A549 and H446 cells were
plated into 96-well plates at a density of 2×103
cells/well and cultured in DMEM supplemented with 10% FBS. For
certain experiments, cells were also treated with shikonin [0, 10,
20 or 50 µM] for 24, 48, 72 or 96 h at 37°C. At each time point,
Cell Counting Kit-8 (CCK-8) reagent (Dojindo Molecular
Technologies, Inc.) was added to each well and incubated for 2 h at
37°C. The optical density (OD) value was measured at a wavelength
of 450 nm using a microplate reader (Thermo Fisher Scientific). The
experiment was repeated in triplicate.
Wound healing assay
A549 and H446 cells were digested using 0.25%
trypsin solution (Gibco; Thermo Fisher Scientific, Inc.) and
cultured in a 6-well plate at a density of 5×105
cells/well overnight at 37°C. Upon cells reaching 90% confluence, a
single vertical scratch was made in the cell monolayer with a 10-µl
micropipette tip. The cells were washed with PBS thrice to remove
the detached cells. Subsequently, A549 and H446 cells were cultured
in serum-free DMEM with/without shikonin (0, 10, 20 or 50 µM) at
37°C or transfected A549 cells were cultured in serum-free DMEM
with 50 µM shikonin at 37°C with 5% CO2 in an incubator.
At 0 h and following 24 h of incubation, the wound area was
visualized and photographed using a light microscope
(magnification, ×100) and the migration was analyzed using ImageJ
software (version 1.8.0; National Institutes of Health).
Colony formation assay
After transfection for 48 h, A549 and H446 cells
were plated into 6-well plates at a density of 0.5–1×103
cells/well; three wells were plated for each experimental
condition. The cells were cultured in complete medium supplemented
with 30% FBS in an incubator with 37°C for 14 days; the medium was
changed every 3 days. During the incubation, the cell state and
colony size were observed under a light microscope (magnification,
×100). After culture, cells were washed with PBS, fixed with 4%
paraformaldehyde for 20 min at room temperature and stained with
0.2% crystal violet for 5 min at room temperature. The cells were
then washed with water, dried, photographed and counted using
ImageJ software; >50 cells counted as a colony.
Transwell assay
Transwell assay was used to analyze cell invasion
and migration. Transwell chambers (Corning, Inc.) were precoated
with (invasion) or without (migration) Matrigel (Becton, Dickinson
and Company) at 37°C for 30 min. Following cell digestion using
0.25% trypsin solution, the culture medium was discarded by
centrifugation (300 × g for 5 min at 37°C) and the cell pellet was
washed with PBS 1–2 times. A549 cells were treated with shikonin
(0, 10, 20 or 50 µM) for 24 h at 37°C. Next, cells were incubated
in serum-free DMEM at a density of 5×105 cells/ml for 24
h at 37°C, then 100 µl cell suspension was added to the upper
chamber of the Transwell plate. A total of 600 µl medium
supplemented with 20% FBS was plated into the lower chambers.
Following incubation for 24 h at 37°C, the cells were fixed with 4%
paraformaldehyde for 10 min at room temperature and stained with
0.1% crystal violet for 10 min at room temperature. Finally, the
cell numbers were counted under a light microscope (magnification,
×200) using ImageJ software.
Flow cytometric analysis of
apoptosis
Cell apoptosis was analyzed using an Annexin
V-FITC/propidium iodide (PI) staining kit (Invitrogen; Thermo
Fisher Scientific, Inc.). Briefly, A549 cells were treated with
shikonin (0, 10, 20 or 50 µM) for 24 h at 37°C or transfected A549
cells were treated with 50 µM shikonin for 24 h at 37°C and
collected by centrifugation (300 × g for 5 min at 37°C). Cells
(1×105 cells/well) were then washed with cold (4°C) PBS
twice and resuspended in 1X binding buffer (200 µl). Cells were
subsequently incubated with 5 µl Annexin V-FITC/PI at room
temperature in the dark for 15 min. Apoptotic cells (early and late
apoptotic cells) were visualized using a flow cytometer (Accuri™ C6
Plus; BD Biosciences).
Measurement of lactic acid production,
glucose uptake and cellular ATP levels
A549 and H446 cells (1×104 cells/well)
were seeded into 6-well plates and cultured in DMEM. A549 and H446
cells were treated with shikonin (0, 10, 20 or 50 µM) for 24 h at
37°C or transfected A549 cells were treated with 50 µM shikonin for
24 h at 37°C, then cell culture medium was subsequently collected
and centrifuged at 8,000 × g for 5 min at room temperature. The
supernatant was used to measure lactate or glucose concentrations,
or cellular ATP levels. Extracellular lactate levels were measured
using a lactate analysis kit (cat. no. E4341; BioVision, Inc.)
according to the manufacturer's protocol. Cell lysates were used to
measure glucose levels using a glucose assay kit (cat. no. K686;
BioVision, Inc.) according to the manufacturer's protocol. ATP
levels were measured using the CellTiter-Glo®
luminescent cell activity assay kit (cat. no. G7572; Promega
Corporation) according to the manufacturer's protocol. The relative
value of lactate, glucose uptake and ATP levels were normalized to
the control group (set to 1).
Statistical analysis
Statistical analysis was performed using SPSS 21.0
software (IBM Corp.) and data are expressed as the mean ± SEM of at
least three independent experiments. Statistical differences
between groups were determined using a one-way ANOVA followed by a
Tukey's post hoc test. A paired Student's t-test was used to
analyze the mRNA expression levels of PFKFB2 between NSCLC and
adjacent normal tissues. P<0.05 was considered to indicate a
statistically significant difference.
Results
Shikonin inhibits the proliferation,
invasion and migration of A549 cells
To investigate the antitumor effects of shikonin,
the inhibitory effect of shikonin was determined using cellular
functional experiments. The results of the cell proliferation
experiment revealed that shikonin (10, 20 or 50 µM) could inhibit
the proliferation of A549 cells in a dose-dependent manner compared
with the control group (all P<0.05; Fig. 1A). The results from the wound
healing assay demonstrated that 10, 20 or 50 µM shikonin could
significantly inhibit the migration of A549 cells compared with the
control group (all P<0.05; Fig.
1B). The results of the Transwell assays showed that 10, 20 or
50 µM shikonin could significantly inhibit the migration and
invasion of A549 cells in a dose-dependent manner compared with the
control group (all P<0.05; Fig.
1C). Analysis of cell apoptosis revealed that 10, 20 or 50 µM
shikonin increased the apoptosis of A549 cells compared with the
control group (all P<0.05; Fig.
1D).
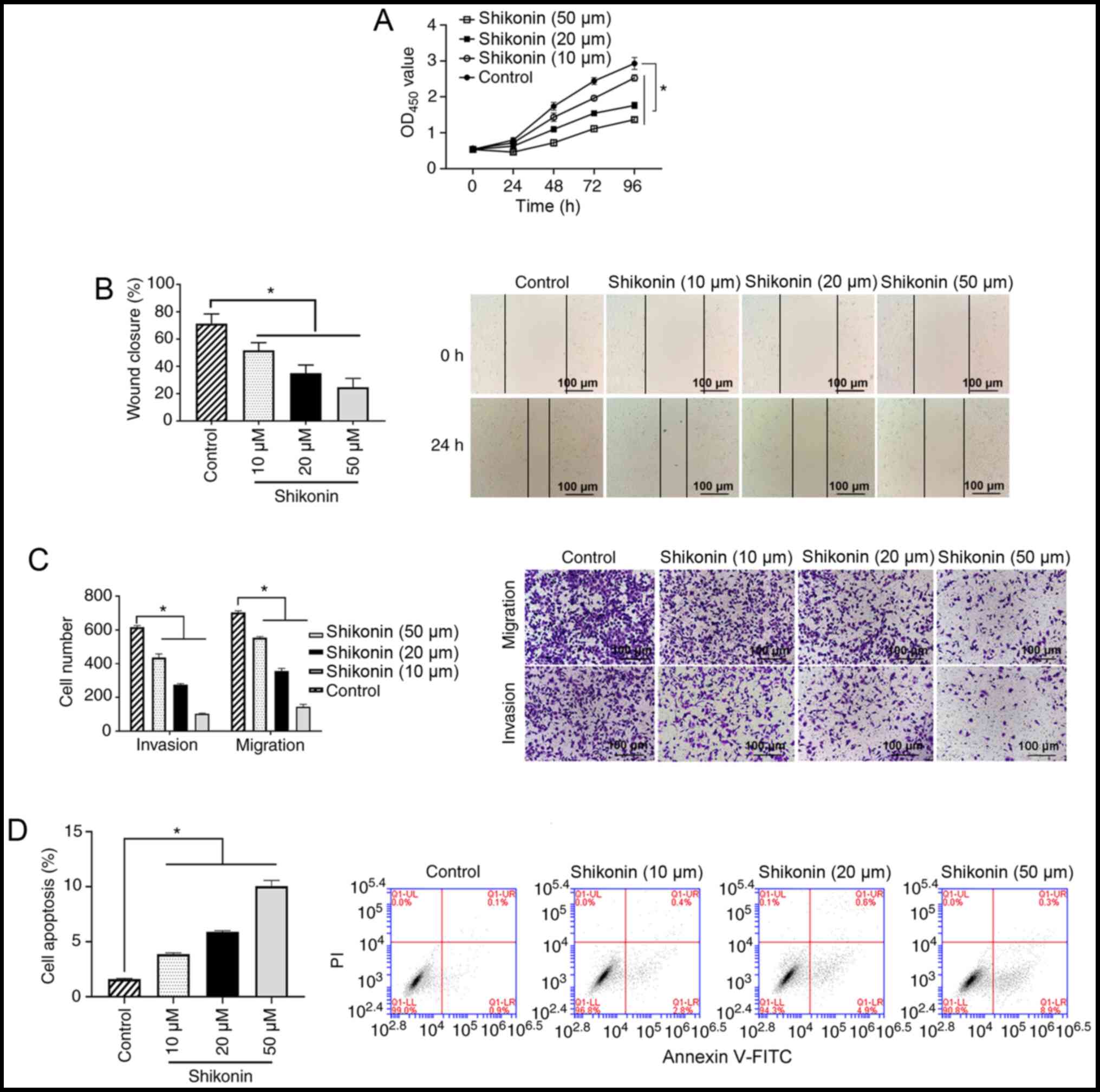 | Figure 1.Effect of shikonin on A549 cells. (A)
A549 cells were treated with 0, 10, 20 or 50 µM shikonin for 24,
48, 72 or 96 h, and cell proliferation was measured using a Cell
Counting Kit-8 assay. *P<0.05. (B) A549 cells were treated with
0, 10, 20 or 50 µM shikonin for 24 h, and the cell migration
ability was analyzed using a wound healing assay. Scale bar,
100-µm. (C) A549 cells were treated with 0, 10, 20 or 50 µM
shikonin for 24 h, and the cell invasion and migration abilities
were measured using Transwell assays. Scale bar, 100-µm. (D) A549
cells were treated with 0, 10, 20 or 50 µM shikonin for 24 h, and
cell apoptosis was analyzed using flow cytometry. *P<0.05. OD,
optical density; PI, propidium iodide. |
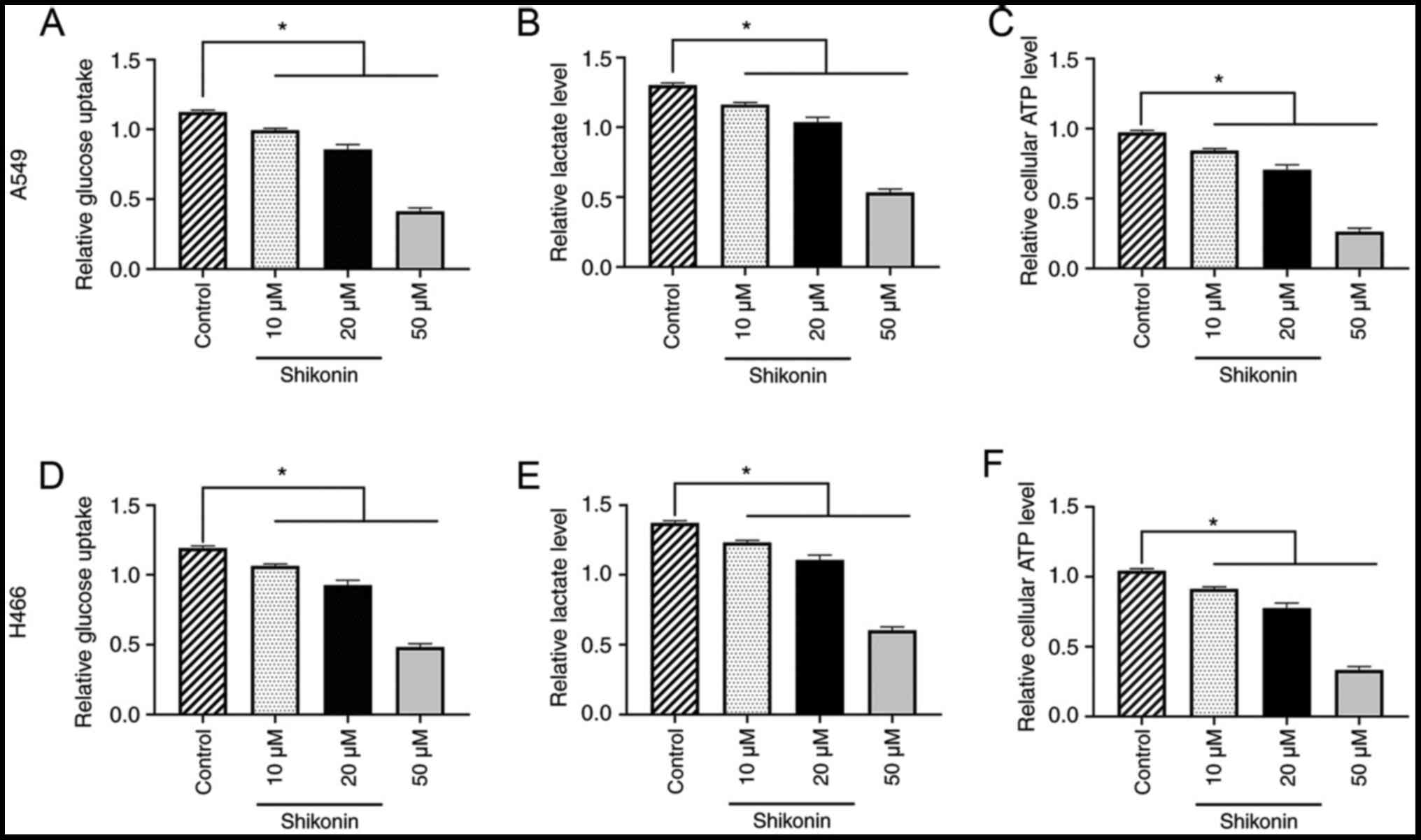
Shikonin inhibits the Warburg effect
in a dose-dependent manner and regulates glucose metabolism in A549
and H446 cells
To further determine the underlying mechanism of the
effect of shikonin on lung cancer cells, the effect of shikonin on
the Warburg effect and glucose metabolism was investigated in A549
and H446 cells. The results found that 10, 20 or 50 µM shikonin
could dose-dependently reduce glucose uptake, lactate and ATP
levels in A549 cells compared with the control group (all
P<0.05; Fig. 2A-C). Similar
results were obtained for H446 cells (all P<0.05; Fig. 2D-F). These data indicated that
shikonin may inhibit the Warburg effect and regulate glucose
metabolism in lung cancer cells.
PFKFB2 expression is regulated by
shikonin
To determine the mechanism through which shikonin
inhibits glycolysis and the migration, proliferation and invasion
of tumor cells, the protein expression levels of key enzymes
involved in aerobic glucose metabolism were analyzed in A549 and
H446 cells treated with shikonin. As shown in Fig. 3A, western blotting analysis revealed
that shikonin downregulated the protein expression levels of PFKFB2
in a dose-dependent manner in A549 and H446 cells, while the
expression levels of the other proteins (PDK1, GLUT1, PGK2, LDHA,
PKM2, GLUT3, PDH and p-PDH) were not altered by shikonin treatment.
These findings indicated that PFKFB2 expression may be regulated by
shikonin. In addition, RT-qPCR analysis was performed to analyze
the mRNA expression levels of PFKFB2 in A549 and H446 cells treated
with 50 µM shikonin. The results revealed that treatment with
shikonin significantly downregulated the mRNA expression levels of
PFKFB2 compared with the control group (both P<0.05; Fig. 3B).
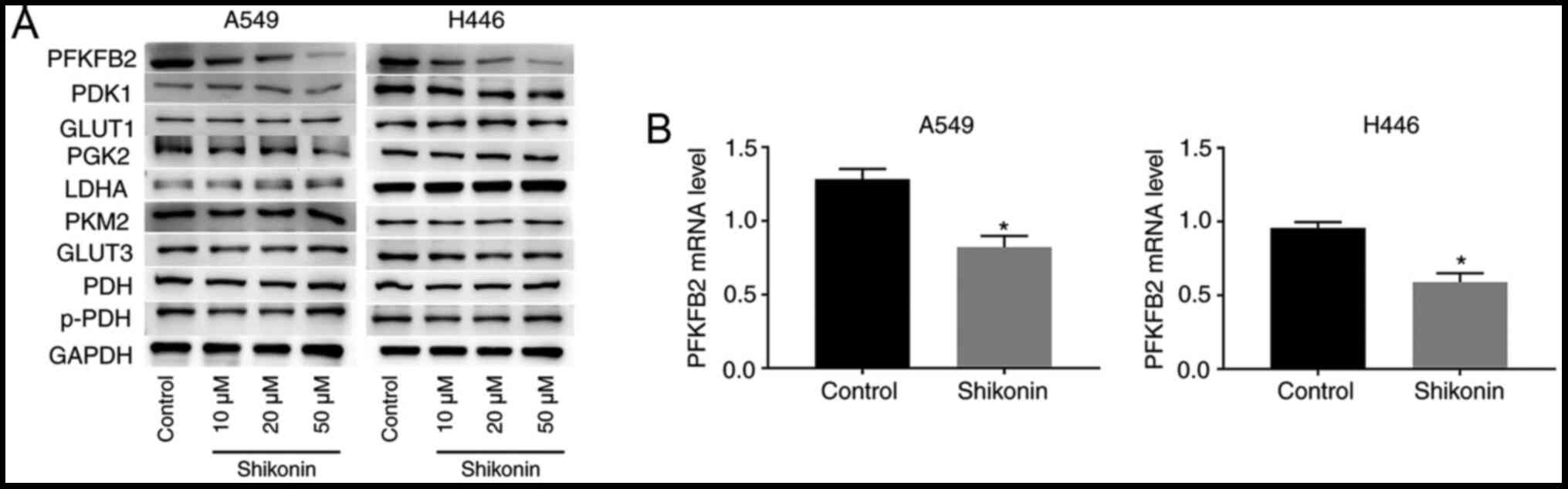 | Figure 3.Shikonin downregulates PFKFB2
expression levels in A549 and H446 cells. (A) A549 and H446 cells
were incubated with 0, 10, 20 or 50 µM shikonin for 24 h.
Expression levels of PFKFB2, PDK1, GLUT1, PGK2, LDHA, PKM2, GLUT3,
PDH and p-PDH in A549 and H446 cells were analyzed using western
blotting. (B) mRNA expression levels of PFKFB2 were analyzed using
reverse transcription-quantitative PCR in A549 and H446 cells
treated with 50 µM shikonin. *P<0.05 vs. control. PFKFB2,
6-phosphofructo-2-kinase/fructose-2,6-biphosphatase 2; PDK1,
pyruvate dehydrogenase kinase 1; GLUT, glucose transporter; PGK2,
phosphoglycerate kinase 2; LDHA, lactate dehydrogenase A; PKM2,
pyruvate kinase M1/2; PDH, pyruvate dehydrogenase phosphatase
catalytic subunit 1; p-, phosphorylated. |
Expression levels of PFKFB2 are
upregulated in lung cancer tissues, and the overexpression of
PFKFB2 promotes proliferation, migration and the Warburg effect in
lung cancer cells
To investigate the role of PFKFB2, the expression
levels of PFKFB2 were first analyzed in 20 NSCLC and adjacent
normal tissues. The results demonstrated that PFKFB2 expression
levels were significantly upregulated in NSCLC tissues compared
with the adjacent normal tissues (P<0.05; Fig. 4A). The expression levels of PFKFB2
were also significantly upregulated in lung cancer cells (A549 and
H446) compared with Beas-2B normal lung epithelial cells (both
P<0.05; Fig. 4B). In addition,
RT-qPCR analysis found that PFKFB2 mRNA expression levels were
significantly downregulated in A549 and H446 cells transfected with
siRNA-PFKFB2 compared with siRNA-NC (both P<0.05) and
significantly upregulated in A549 and H446 cells transfected with
pcDNA3.1-PFKFB2 compared with pcDNA3.1-NC (both P<0.05)
(Fig. 4C), which indicated that the
cell transfections were successful. The results of the CCK-8 and
colony formation assays revealed that the overexpression of PFKFB2
significantly increased the proliferation of A549 and H446 cells
compared with the pcDNA3.1-NC group, while the knockdown of PFKFB2
significantly inhibited the proliferation of A549 and H446 cells
compared with the siRNA-NC group (all P<0.05; Fig. 4D and E). The results of the wound
healing assay demonstrated that the overexpression of PFKFB2
significantly promoted the migration of A549 and H446 cells
compared with the pcDNA3.1-NC group, while the knockdown of PFKFB2
significantly inhibited the migration of A549 and H446 cells
compared with the siRNA-NC group (all P<0.05; Fig. 4F).
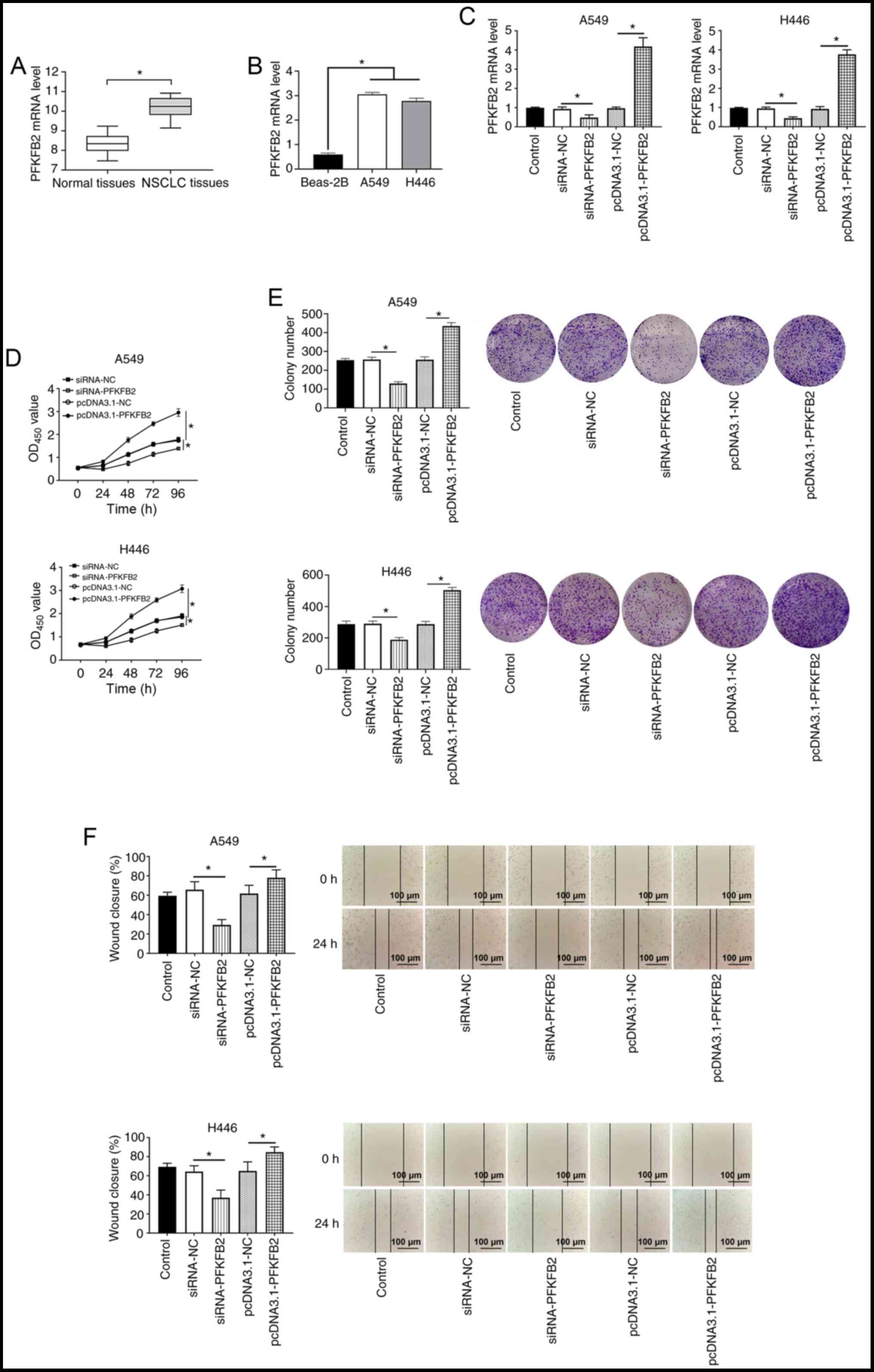 | Figure 4.PFKFB2 expression levels are
upregulated in lung cancer, and the overexpression of PFKFB2
promotes proliferation, migration and the Warburg effect in A549
and H446 cells. (A) mRNA expression levels of PFKFB2 in NSCLC and
adjacent normal tissues were analyzed using RT-qPCR. (B) mRNA
expression levels of PFKFB2 were analyzed in Beas-2B, A549 and H446
cells using RT-qPCR. (C) mRNA expression levels of PFKFB2 in A549
and H446 cells following transfections were analyzed using RT-qPCR.
(D) Proliferation of A549 and H446 cells was detected using a Cell
Counting Kit-8 assay. (E) Colony formation assay was used to
measure the colony number in A549 and H446 cells. (F) Migratory
ability of A549 and H446 cells was analyzed using a wound healing
assay. Scale bar, 100-µm. *P<0.05. PFKFB2,
6-phosphofructo-2-kinase/fructose-2,6-biphosphatase 2; RT-qPCR,
reverse transcription-quantitative PCR; NSCLC, non-small cell lung
cancer; siRNA, small interfering RNA; NC, negative control; OD,
optical density. |
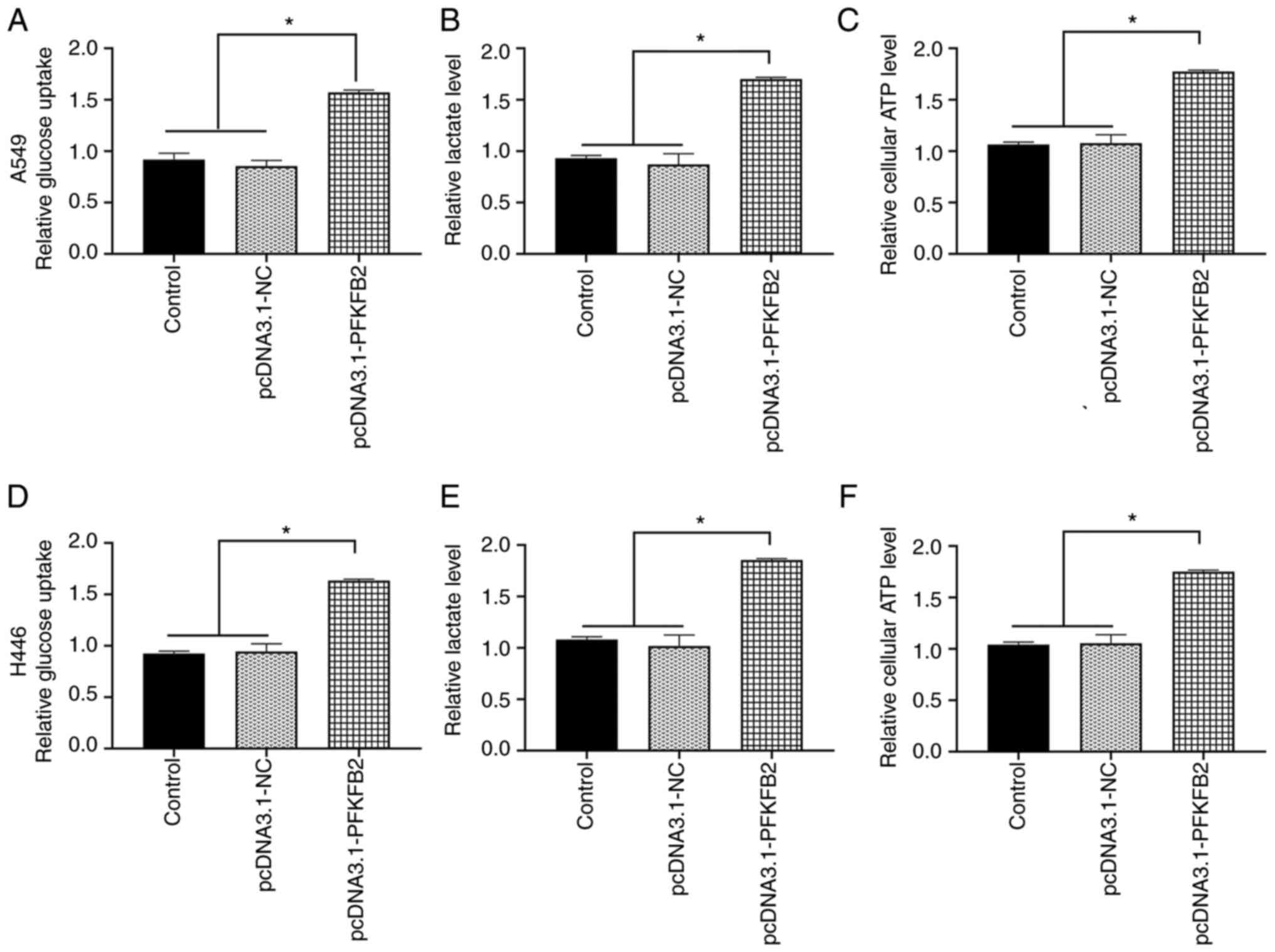
Subsequently, whether PFKFB2 regulated the
glycometabolic activity in A549 and H446 cells was investigated.
The overexpression of PFKFB2 in A549 cells significantly increased
glucose uptake, lactate and ATP levels compared with the control
and pcDNA3.1-NC groups (all P<0.05; Fig. 5A-C). Similar results were obtained
for H446 cells (all P<0.05; Fig.
5D-F).
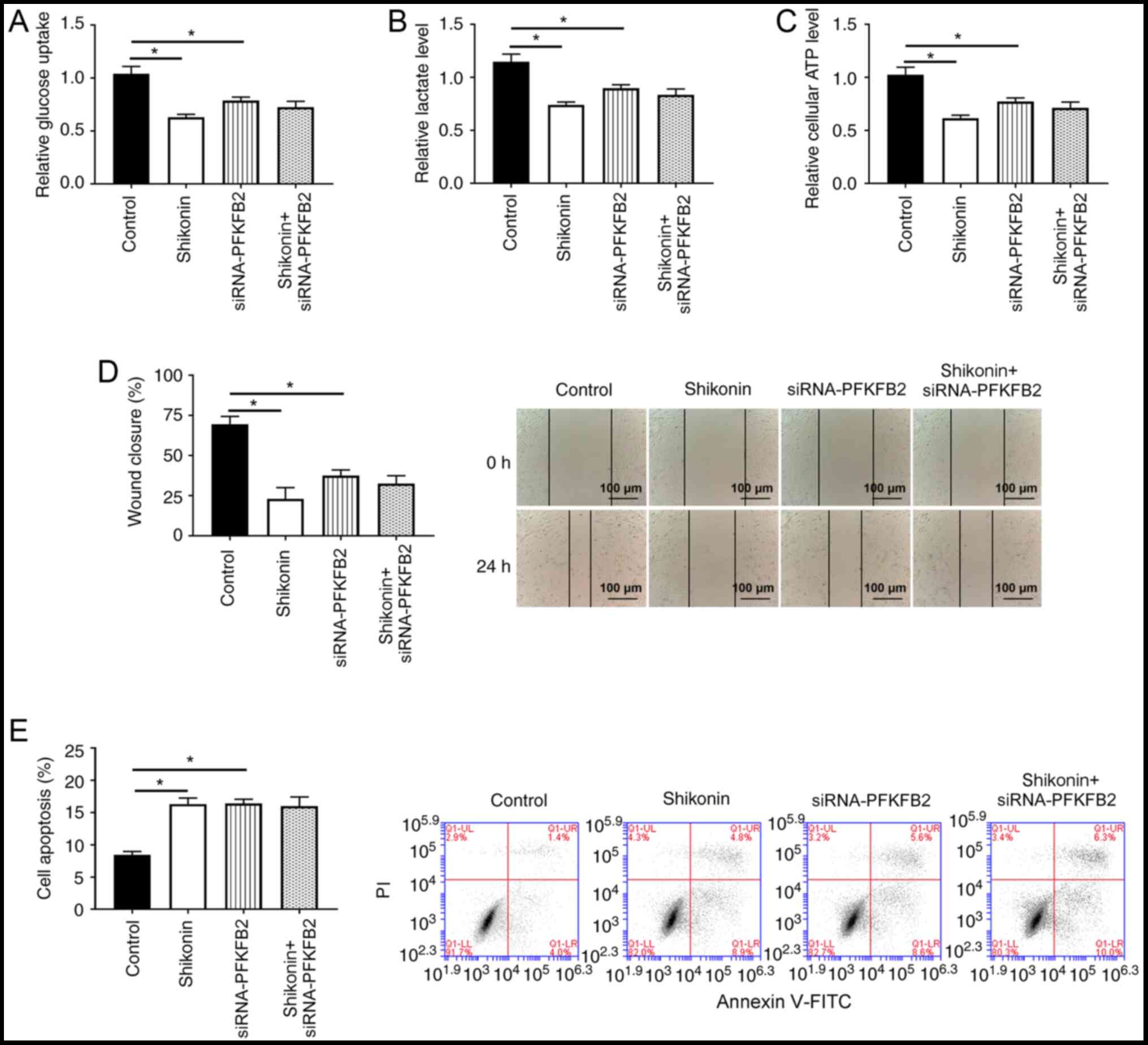
PFKFB2 participates in
shikonin-induced glycolysis, apoptosis and migration in lung cancer
cells
Whether PFKFB2 participated in shikonin-induced
aerobic glycolysis, cell apoptosis and migration was subsequently
investigated. As shown in Fig.
6A-D, the knockdown of PFKFB2 or treatment of shikonin
significantly inhibited glucose uptake, lactic acid production, ATP
levels and migration in A549 cells compared with the control group
(all P<0.05). Notably, there were no significant differences in
glucose uptake, lactate levels, ATP production and migration
between A549 cells with knocked down PFKFB2 or treated with
shikonin and the knockdown of PFKFB2 in A549 cells treated with
shikonin, suggesting that shikonin-mediated glycolysis and
migration may be regulated by PFKFB2 in lung cancer cells. In
addition, the knockdown of PFKFB2 or treatment of shikonin
significantly increased cell apoptosis compared with the control
group (P<0.05), while the knockdown of PFKFB2 in
shikonin-treated cells did not significantly alter the apoptosis
compared with treatment of shikonin or the knockdown of PFKFB2
(Fig. 6E).
Discussion
Lung cancer has become the most commonly diagnosed
cancer worldwide, accounting for 11.6% of total cancers diagnosed
(1). Currently, due to the lack of
effective tumor molecular markers available to guide clinical
diagnosis and treatment, the rate of successful treatment in
patients with lung cancer remains poor; therefore, it remains an
urgent priority to develop novel therapeutic drugs (28). Traditional Chinese medicine has long
been used for the treatment of lung cancer and has demonstrated
considerable clinical value, such as inhibiting metastasis,
enhancing the host immune response and reducing the adverse effects
of chemotherapy (29–31). The results of the present study
revealed that shikonin inhibited cell proliferation, invasion, and
migration, reduced glucose uptake, lactate and ATP levels,
increased apoptosis and downregulated the expression levels of
PFKFB2 in lung cancer cells.
Shikonin, a natural naphthoquinone compound, is
extracted and separated from the dry roots of Lithospermum
erythrorhizon and has been found to play roles in numerous
biological activities, including exerting anti-inflammatory,
antiviral and antitumor effects, immune system regulation,
promoting wound healing and protecting against multi-drug
resistance (32–34). Among these biological activities,
the anticancer effect of shikonin has been well reported. For
example, a previous study reported that shikonin reduced the tumor
diameter by 25% and increased the 1-year survival rate by 47.3% in
patients with lung cancer (35). In
addition, Zhang et al (36)
demonstrated that shikonin inhibited the migration and invasion of
thyroid cancer cells. Guo et al (37) also found that shikonin inhibited
cell proliferation and induced apoptosis in glioma. Wang et
al (38) reported that shikonin
inhibited the proliferation and promoted apoptosis of breast cancer
cells. The results of the present study demonstrated that shikonin
could inhibit proliferation, migration and invasion, and increase
the apoptosis of A549 cells in a dose-dependent manner, which were
consistent with the findings of previous studies (36–38).
However, the results of the present study showed a lower apoptosis
rate, which is different to the results obtained in the
aforementioned studies. Taken together, these results indicated
that shikonin might exert antitumor effects in lung cancer.
Cancer cells do not use mitochondrial oxidative
phosphorylation, even in the presence of oxygen, but instead use
aerobic glycolysis, a phenomenon termed the Warburg effect
(39,40). The Warburg effect has been reported
to permit tumor cells to overcome metabolic stress and promote
energy replenishment, which is vital for the proliferation and
survival of cancer cells (41,42).
In addition, the Warburg effect was suggested to impact the
microenvironment and promote chemoresistance, angiogenesis and
metastasis (43,44). Hence, investigating the Warburg
effect may be crucial for determining the underlying mechanisms of
the development and progression of NSCLC. The results of the
present study found that shikonin could dose-dependently reduce
glucose uptake, lactate and ATP levels in lung cancer cells. These
data indicated that shikonin may inhibit the malignant evolution of
lung cancer by inhibiting the Warburg effect.
PFKFB is an important regulator of glycolysis in
cancer, and it was previously reported that the PFKFB family served
an important role in the development and progression of numerous
tumor types, such as lung, pancreatic and gastric cancer (45–47).
Previous studies demonstrated that the expression levels of PFKFB2
were upregulated in melanoma, osteosarcoma and thyroid carcinoma
(48–50). Furthermore, Liu et al
(51) reported that microRNA-613
inhibited cell proliferation, invasion and the Warburg effect by
regulating PFKFB2 expression in gastric cancer. Ozcan et al
(22) also found that PFKFB2
inhibited glycolysis and proliferation in pancreatic cancer cells.
The present study results showed that the expression levels of
PFKFB2 were significantly upregulated in human NSCLC tissues and
lung cancer cell lines, and that the overexpression of PFKFB2
increased cell proliferation, migration, glucose uptake, lactate
and ATP levels in lung cancer cells. In addition, shikonin
treatment downregulated the expression levels of PFKFB2.
Furthermore, there were no significant differences in glucose
uptake, lactate levels, ATP production, apoptosis and migration
between the knockdown of PFKFB2 or treatment of shikonin and the
knockdown of PFKFB2 in cells treated with shikonin. These results
indicated that PFKFB2 may play an important role in the effects of
shikonin treatment in lung cancer.
To the best of our knowledge, the present study was
the first to demonstrate that shikonin inhibited lung cancer cell
migration and the Warburg effect by regulating PFKFB2 expression,
which may provide a novel insight into the mechanism underlying the
anticancer effects of shikonin. However, it is important to note
the limitations of the present study. For example, the proportion
of apoptotic cells was too low to ascertain that shikonin induced
apoptosis. In addition, the current study did not block the Warburg
effect to further validate the effect of shikonin on cell
proliferation, apoptosis and migration. The above limitations may
weaken the conclusions of the present study; therefore, koningic
acid, an irreversible and selective inhibitor of GAPDH (a
rate-controlling glycolytic enzyme during the Warburg effect) will
be used in future studies to block the Warburg effect to further
validate the mechanism of shikonin in lung cancer (52).
In conclusion, the results of the present study
suggested that shikonin inhibited the metabolism, migration and
proliferation of lung cancer cells. In lung cancer tissues, PFKFB2
expression levels were upregulated and promoted the progression of
lung cancer. In addition, PFKFB2 was found to participate in the
shikonin-induced effects on the glycolysis and migration of lung
cancer cells. These data suggested that treatments targeting PFKFB2
may benefit patients with lung cancer. In addition, shikonin may
represent a potential novel compound for the treatment of lung
cancer.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
HZ conceived and designed the study; LS, ZL and XS
performed the experiments; TW, YL and YZ analyzed and interpreted
the data; and all authors wrote the manuscript. All authors read
and approved the final manuscript. LS and HZ confirm the
authenticity of all the raw data.
Ethics approval and consent to
participate
The study protocols were approved by the Ethics
Committee of The Affiliated Hospital of Qingdao University, and
written informed consent was obtained from all patients prior to
participation.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Blandin Knight S, Crosbie PA, Balata H,
Chudziak J, Hussell T and Dive C: Progress and prospects of early
detection in lung cancer. Open Biol. 7:1700702017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Rossi A, Tay R, Chiramel J, Prelaj A and
Califano R: Current and future therapeutic approaches for the
treatment of small cell lung cancer. Expert Rev Anticancer Ther.
18:473–486. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
King-Kallimanis BL, Kanapuru B, Blumenthal
GM, Theoret MR and Kluetz PG: Age-related differences in
patient-reported outcomes in patients with advanced lung cancer
receiving anti-PD-1/PD-L1 therapy. Semin Oncol. 45:201–209. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Shukuya T and Carbone DP: Predictive
markers for the efficacy of anti-PD-1/PD-L1 antibodies in lung
cancer. J Thorac Oncol. 11:976–988. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Festino L, Botti G, Lorigan P, Masucci GV,
Hipp JD, Horak CE, Melero I and Ascierto PA: Cancer treatment with
Anti-PD-1/PD-L1 agents: Is PD-L1 expression a biomarker for patient
selection? Drugs. 76:925–945. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhong W, Chen S, Qin Y, Zhang H, Wang H,
Meng J, Huai L, Zhang Q, Yin T, Lei Y, et al: Doxycycline inhibits
breast cancer EMT and metastasis through
PAR-1/NF-κB/miR-17/E-cadherin pathway. Oncotarget. 8:104855–104866.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhong W, Chen S, Zhang Q, Xiao T, Qin Y,
Gu J, Sun B, Liu Y, Jing X, Hu X, et al: Doxycycline directly
targets PAR1 to suppress tumor progression. Oncotarget.
8:16829–16842. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Liberti MV and Locasale JW: The Warburg
effect: How does it benefit cancer cells? Trends Biochem Sci.
41:211–218. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Bi YH, Han WQ, Li RF, Wang YJ, Du ZS, Wang
XJ and Jiang Y: Signal transducer and activator of transcription 3
promotes the Warburg effect possibly by inducing pyruvate kinase M2
phosphorylation in liver precancerous lesions. World J
Gastroenterol. 25:1936–1949. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Luo Y, Medina Bengtsson L, Wang X, Huang
T, Liu G, Murphy S, Wang C, Koren J III, Schafer Z and Lu X: UQCRH
downregulation promotes Warburg effect in renal cell carcinoma
cells. Sci Rep. 10:150212020. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Martinez-Outschoorn UE, Lin Z, Trimmer C,
Flomenberg N, Wang C, Pavlides S, Pestell RG, Howell A, Sotgia F
and Lisanti MP: Cancer cells metabolically ‘fertilize’ the tumor
microenvironment with hydrogen peroxide, driving the Warburg
effect: Implications for PET imaging of human tumors. Cell Cycle.
10:2504–2520. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Gregory CD and Paterson M: An
apoptosis-driven ‘onco-regenerative niche’: Roles of
tumour-associated macrophages and extracellular vesicles. Philos
Trans R Soc Lond B Biol Sci. 373:201700032018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chen Z, Zuo X, Yao Z, Han G, Zhang L, Wu J
and Wang X: MiR-3662 suppresses hepatocellular carcinoma growth
through inhibition of HIF-1α-mediated Warburg effect. Cell Death
Dis. 9:5492018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang S, Zhang Y, Cai Q, Ma M, Jin LY, Weng
M, Zhou D, Tang Z, Wang JD and Quan Z: Circular RNA FOXP1 promotes
tumor progression and Warburg effect in gallbladder cancer by
regulating PKLR expression. Mol Cancer. 18:1452019. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li J, Zhang J, Xie F, Peng J and Wu X:
Macrophage migration inhibitory factor promotes Warburg effect via
activation of the NF-κB/HIF-1α pathway in lung cancer. Int J Mol
Med. 41:1062–1068. 2018.PubMed/NCBI
|
|
17
|
Lu J, Tan M and Cai Q: The Warburg effect
in tumor progression: Mitochondrial oxidative metabolism as an
anti-metastasis mechanism. Cancer Lett. 356:156–164. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Feng Y, Liu J, Guo W, Guan Y, Xu H, Guo Q,
Song X, Yi F, Liu T, Zhang W, et al: Atg7 inhibits Warburg effect
by suppressing PKM2 phosphorylation resulting reduced
epithelial-mesenchymal transition. Int J Biol Sci. 14:775–783.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Xu L, Li Y, Zhou L, Dorfman RG, Liu L, Cai
R, Jiang C, Tang D, Wang Y, Zou X, et al: SIRT3 elicited an
anti-Warburg effect through HIF1α/PDK1/PDHA1 to inhibit
cholangiocarcinoma tumorigenesis. Cancer Med. 8:2380–2391. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhu W, Huang Y, Pan Q, Xiang P, Xie N and
Yu H: MicroRNA-98 suppress Warburg effect by targeting HK2 in colon
cancer cells. Dig Dis Sci. 62:660–668. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Khan MA, Zubair H, Anand S, Srivastava SK,
Singh S and Singh AP: Dysregulation of metabolic enzymes in tumor
and stromal cells: Role in oncogenesis and therapeutic
opportunities. Cancer Lett. 473:176–185. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ozcan SC, Sarioglu A, Altunok TH, Akkoc A,
Guzel S, Guler S, Imbert-Fernandez Y, Muchut RJ, Iglesias AA,
Gurpinar Y, et al: PFKFB2 regulates glycolysis and proliferation in
pancreatic cancer cells. Mol Cell Biochem. 470:115–129. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yang H, Shu Z, Jiang Y, Mao W, Pang L,
Redwood A, Jeter-Jones SL, Jennings NB, Ornelas A, Zhou J, et al:
6-Phosphofructo-2-kinase/fructose-2,6-biphosphatase-2 regulates
TP53-dependent paclitaxel sensitivity in ovarian and breast
cancers. Clin Cancer Res. 25:5702–5716. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Pape MR, Prokop C and Hedrich HJ:
Localization of the Pfkfb 2 gene on rat Chromosome 13. Mamm Genome.
7:5591996. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Novellasdemunt L, Tato I, Navarro-Sabate
A, Ruiz-Meana M, Méndez-Lucas A, Perales JC, Garcia-Dorado D,
Ventura F, Bartrons R and Rosa JL: Akt-dependent activation of the
heart 6-phosphofructo-2-kinase/fructose-2,6-bisphosphatase (PFKFB2)
isoenzyme by amino acids. J Biol Chem. 288:10640–10651. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Jeung YJ, Kim HG, Ahn J, Lee HJ, Lee SB,
Won M, Jung CR, Im JY, Kim BK, Park SK, et al: Shikonin induces
apoptosis of lung cancer cells via activation of FOXO3a/EGR1/SIRT1
signaling antagonized by p300. Biochim Biophys Acta.
1863:2584–2593. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Barta JA, Powell CA and Wisnivesky JP:
Global epidemiology of lung cancer. Ann Glob Health. 85:82019.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li L, Wang S, Yang X, Long S, Xiao S, Wu W
and Hann SS: Traditional Chinese medicine, Fuzheng Kang-Ai
decoction, inhibits metastasis of lung cancer cells through the
STAT3/MMP9 pathway. Mol Med Rep. 16:2461–2468. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Liao YH, Li CI, Lin CC, Lin JG, Chiang JH
and Li TC: Traditional Chinese medicine as adjunctive therapy
improves the long-term survival of lung cancer patients. J Cancer
Res Clin Oncol. 143:2425–2435. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yang J, Zhu X, Yuan P, Liu J, Wang B and
Wang G: Efficacy of traditional Chinese medicine combined with
chemotherapy in patients with non-small cell lung cancer (NSCLC): A
meta-analysis of randomized clinical trials. Support Care Cancer.
28:3571–3579. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Andújar I, Ríos JL, Giner RM and Recio MC:
Pharmacological properties of shikonin-a review of literature since
2002. Planta Med. 79:1685–1697. 2013. View Article : Google Scholar
|
|
33
|
Wang F, Yao X, Zhang Y and Tang J:
Synthesis, biological function and evaluation of shikonin in cancer
therapy. Fitoterapia. 134:329–339. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wei PL, Tu CC, Chen CH, Ho YS, Wu CT, Su
HY, Chen WY, Liu JJ and Chang YJ: Shikonin suppresses the migratory
ability of hepatocellular carcinoma cells. J Agric Food Chem.
61:8191–8197. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Guo XP, Zhang XY and Zhang SD: Clinical
trial on the effects of shikonin mixture on later stage lung
cancer. Zhong Xi Yi Jie He Za Zhi. 11:598–599, 580. 1991.(In
Chinese). PubMed/NCBI
|
|
36
|
Zhang Y, Sun B, Huang Z, Zhao DW and Zeng
Q: Shikonin inhibites migration and invasion of thyroid cancer
cells by downregulating DNMT1. Med Sci Monit. 24:661–670. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Guo N, Miao R, Gao X, Huang D, Hu Z, Ji N,
Nan Y, Jiang F and Gou X: Shikonin inhibits proliferation and
induces apoptosis in glioma cells via downregulation of CD147. Mol
Med Rep. 19:4335–4343. 2019.PubMed/NCBI
|
|
38
|
Wang W, Wu Y, Chen S, Liu X, He J, Wang S,
Lu W, Tang Y and Huang J: Shikonin is a novel and selective IMPDH2
inhibitor that target triple-negative breast cancer. Phytother Res.
35:463–476. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Yu Z, Huang L, Qiao P, Jiang A, Wang L,
Yang T, Tang S, Zhang W and Ren C: PKM2 Thr454 phosphorylation
increases its nuclear translocation and promotes xenograft tumor
growth in A549 human lung cancer cells. Biochem Biophys Res Commun.
473:953–958. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Rong F, Tian X, Fang Z, Sun Y, Li F, Gao
Y, Feng Y, Li L, Wang Y, Liu X, et al: MicroRNA-143 (miR-143)
regulates cancer glycolysis via targeting hexokinase 2 gene. J Biol
Chem. 287:23227–23235. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
DeBerardinis RJ, Lum JJ, Hatzivassiliou G
and Thompson CB: The biology of cancer: Metabolic reprogramming
fuels cell growth and proliferation. Cell Metab. 7:11–20. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Schild T, Low V, Blenis J and Gomes AP:
Unique metabolic adaptations dictate distal organ-specific
metastatic colonization. Cancer Cell. 33:347–354. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Shamsi M, Saghafian M, Dejam M and
Sanati-Nezhad A: Mathematical modeling of the function of Warburg
effect in tumor microenvironment. Sci Rep. 8:89032018. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Hussain A, Qazi AK, Mupparapu N, Guru SK,
Kumar A, Sharma PR, Singh SK, Singh P, Dar MJ, Bharate SB, et al:
Modulation of glycolysis and lipogenesis by novel PI3K selective
molecule represses tumor angiogenesis and decreases colorectal
cancer growth. Cancer Lett. 374:250–260. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Yalcin A, Telang S, Clem B and Chesney J:
Regulation of glucose metabolism by
6-phosphofructo-2-kinase/fructose-2,6-bisphosphatases in cancer.
Exp Mol Pathol. 86:174–179. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Minchenko OH, Ogura T, Opentanova IL,
Minchenko DO, Ochiai A, Caro J, Komisarenko SV and Esumi H:
6-Phosphofructo-2-kinase/fructose-2,6-bisphosphatase gene family
overexpression in human lung tumor. Ukr Biokhim Zh. 77:46–50.
1999.PubMed/NCBI
|
|
47
|
Minchenko OH, Tsuchihara K, Minchenko DO,
Bikfalvi A and Esumi H: Mechanisms of regulation of PFKFB
expression in pancreatic and gastric cancer cells. World J
Gastroenterol. 20:13705–13717. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Houles T, Gravel SP, Lavoie G, Shin S,
Savall M, Méant A, Grondin B, Gaboury L, Yoon SO, St-Pierre J and
Roux PP: RSK regulates PFK-2 activity to promote metabolic rewiring
in melanoma. Cancer Res. 78:2191–2204. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Zhao SJ, Shen YF, Li Q, He YJ, Zhang YK,
Hu LP, Jiang YQ, Xu NW, Wang YJ, Li J, et al: SLIT2/ROBO1 axis
contributes to the Warburg effect in osteosarcoma through
activation of SRC/ERK/c-MYC/PFKFB2 pathway. Cell Death Dis.
9:3902018. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Camargo Barros-Filho M, Barreto Menezes de
Lima L, Bisarro Dos Reis M, Bette Homem de Mello J, Moraes Beltrami
C, Lopes Pinto CA, Kowalski LP and Rogatto SR: PFKFB2 promoter
hypomethylation as recurrence predictive marker in
well-differentiated thyroid carcinomas. Int J Mol Sci. 20:13342019.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Liu H, Chen K, Wang L, Zeng X, Huang Z, Li
M, Dong P and Chen X: miR-613 inhibits Warburg effect in gastric
cancer by targeting PFKFB2. Biochem Biophys Res Commun. 515:37–43.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Liberti MV, Allen AE, Ramesh V, Dai Z,
Singleton KR, Guo Z, Liu JO, Wood KC and Locasale JW: Evolved
resistance to partial GAPDH inhibition results in loss of the
Warburg effect and in a different state of glycolysis. J Biol Chem.
295:111–124. 2020. View Article : Google Scholar : PubMed/NCBI
|