Introduction
Myocardial ischemia/reperfusion (I/R) injury,
referring to aggravated tissue injury or irreversible injury in
myocardial ischemic tissues following the reperfusion of blood flow
(1), is associated with serious
clinical manifestations, including myocardial hibernation,
myocardial infarction and acute heart failure (2), and is thus marked as a risk factor for
morbidity and mortality (3). The
application of reperfusion in I/R aims at restoring blood flow,
through which oxygen is provided to ischemic tissues that present
with hypoxia and hypoperfusion caused by the obstruction of
arterial blood flow (4). However,
reperfusion itself can trigger a cascade of pathophysiological
reactions that contribute to the expansion of the infarct area,
accounting for up to 50% of the final infarct size of the ischemic
myocardium (5). Despite the
advances in the therapeutic strategies (6), the onset of I/R injury remains high
during several common clinical conditions, such as coronary bypass
surgery, thrombolytic therapy, cardiopulmonary resuscitation or
organ transplantation (7,8). It has been shown that the pathological
mechanism of I/R is associated with decreased ATP production and
increased reactive oxygen species (ROS) retention, which leads to
profound inflammatory responses that further induce the apoptosis,
necrosis and autophagy of cardiomyocytes (4,9).
Lycium barbarum polysaccharide (LBP), characterized
by the composition of glucose monosaccharides and fructose
monosaccharides at a molar level of 1:2, is an active constituent
extracted from Lycium barbarum, the fruit of which is used in the
practice with traditional Chinese medicine (10). Various studies have reported the
pharmacological and biological effects of LBP, including
antioxidative (11),
immune-regulatory (12),
anti-cancer (13), neuroprotective
(14) and blood
sugar/lipid-lowering properties (15). Notably, LBP is documented to exert
cardio-protective effects against I/R-caused damage in vivo by
reducing the levels of myocardial lactate dehydrogenase (LDH),
increasing sodium-potassium ATPase and calcium ATPase activities,
and repressing the apoptosis of cardiomyocytes (10). Moreover, LBP can attenuate cardiac
hypertrophy, and decrease the levels of certain inflammatory
factors, such as IL-6 and TNF-α, as well as lower the production of
ROS in heart tissues of diabetic rats (16). However, the detailed mechanism via
which LBP protects against I/R-induced injury in cardiomyocytes
remain unknown.
Xiao et al (15) revealed that the autophagic process
modulated by LBP attenuated hepatic inflammatory responses and cell
apoptosis in rats with non-alcoholic steatohepatitis. Autophagy is
an evolutionarily conserved mechanism in charge of controlling
intracellular protein and organelle via lysosome-dependent
degradation (17). The key
regulatory factors of autophagy are Beclin-1, LC3 and sequestosome
1 (SQSTM1 or P62, one of the particular substrate protein of
autophagosome). Beclin 1, a mammalian ortholog of yeast Atg6,
mediates the formation of autophagosomes and, thus, contributes to
autophagy initiation (18). The
conversion of LC3-I to LC3-II is an important autophagy marker.
(19). Furthermore, a decrease in
P62 induces the formation of autolysosomes. Under normal
conditions, autophagy stays at basal level in the heart; however,
dysregulated and enhanced autophagy is observed during the
occurrence of cardiovascular diseases, such as myocardial I/R
injury (20). Accordingly, reducing
excessive I/R injury-induced autophagy via trimetazidine inhibits
myocardial apoptosis and oxidative stress, diminishes myocardial
infarct area and restores cardiac function (21). Additionally, nuclear
factor-erythroid factor 2-related factor 2 (Nrf2) in adenocarcinoma
alveolar basal epithelial (A549) cells is negatively associated
with autophagy induced by particulate matter (PM) 2.5 (22).
Therefore, the present study aimed to investigate
and evaluate the efficacy and mechanism of LBP in myocardial I/R
injury, where the involvement of autophagy and its relationship
with Nrf2 were also elucidated.
Materials and methods
Ethics statement
All animal experiments were performed in accordance
with the Guidelines for the Care and Use of Laboratory Animals
(23). This study was approved by
the Ethic Committee of Experimental Animals of Zhejiang Provincial
Animal Center (approval no. DC201901203; Hangzhou, China). Every
effort was made to minimize pain and discomfort to the animals. The
animals experiments were performed in Zhejiang Provincial Animal
Center.
Establishment of I/R rat models
In total, 24 male Sprague-Dawley rats, aged 7–8
weeks old and weighing 250–280 g, were purchased from Shanghai SLAC
Laboratory Animal Co., Ltd., and housed in controlled conditions
(22–25°C, 55% humidity and a 12:12 h circadian cycle). All rats
were fed ad libitum with commercial food and sterile water in
Zhejiang Provincial Animal Center, and were acclimated for 2 days
prior to the commence of the experiment. Rats were then randomly
assigned into four groups (n=6 for each group): Sham group, LBP
group, I/R group and I/R + LBP group. LBP (purity, 51.87%) was
obtained from Xi'an Natural Field Bio-Technique Co., Ltd. The
specific steps of the extraction of the LBP were as follows: The
deionized water extract of Lycium barbarum was filtered through
filter paper to eliminate dregs. After being concentrated to the
volume under vacuum, the crude extract was diluted to deionized
water, precipitated with 95% ethanol, followed by precipitation and
centrifugation (3,000 × g for 25 min at 4°C). Finally, LBP (the
precipitate) was harvested and minced into powder.
The specific treatment of rats in each group is
listed in Fig. 1A. The rats in the
LBP and I/R + LBP groups were treated with 500 mg/kg/day LBP
(24,25) for 7 days (once a day) via oral
administration (26), before sham
operation and myocardial I/R injury, respectively. Those in the
Sham group and I/R group received 0.9% sodium chloride in the same
manner. On the 8th day, the rats were anesthetized via an
intraperitoneal injection of 1.5% tribromoethanol (250 mg/kg; cat.
no. T48402; Sigma-Aldrich; Merck KGaA). After successful
anesthesia, the rats were placed in the supine position on the
animal operating table. For the induction of ischemia, the chest of
each rat was opened through a left parasternal incision, and the
heart exposed at the left 3rd-4th intercostal space. Then, the left
anterior descending coronary artery 1.5–2 mm below left atrial
appendage was ligated by tying a slipknot with 5/0 silk sutures.
The heart was repositioned into the chest, the blood and gas in the
chest were then squeezed out, and the chest was closed quickly.
After 30 min of induction, the slipknot was untied and the
myocardium underwent the reperfusion for 120 min. The rats in the
sham operation group underwent the same surgical operation, but the
coronary artery was not ligated. After 120 min of reperfusion, each
rat was anesthetized as aforementioned, and transthoracic
echocardiography was performed. Then, all rats were sacrificed via
dislocation under anesthesia. Subsequently, the blood of the rats
was collected from the abdominal aorta and the serum obtained via
centrifugation (at 1,500 × g for 10 min.) was stored at −80°C for
further analysis. The hearts of rats were harvested and rinsed with
ice-cold PBS (cat. no. P4417; Sigma-Aldrich; Merck KGaA), and the
ventricular tissues were immediately stored at −80°C.
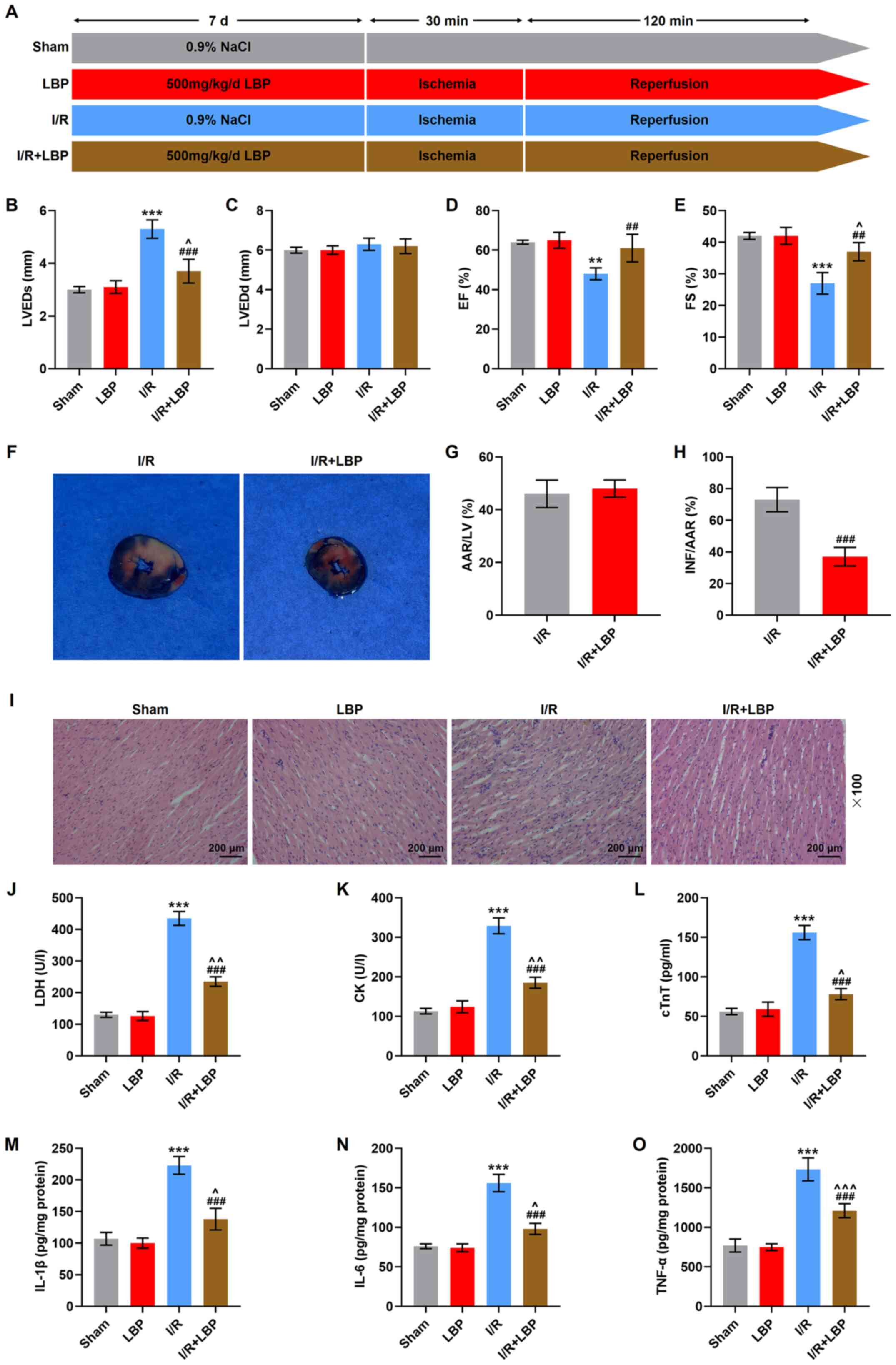 | Figure 1.LBP counteracts I/R-induced heart
malfunction, histopathological abnormalities and inflammation in
rats. (A) A diagram of the establishment of the I/R model and LBP
treatment. (B) LVEDs, (C) LVEDd, (D) EF and (E) FS in I/R- and
LBP-treated rats were determined via transthoracic
echocardiography. (F) Representative images of heart sections of
I/R-injured rats treated with LBP were analyzed via Evan's blue and
2,3,5-triphenyl-tetrazolium chloride double-staining. The
percentages of (G) AAR/LV and (H) INF/AAR were estimated. AAR/LV
reflects the extent of myocardial ischemia, while NF/AAR reflects
the level of dead myocardium. (I) The histopathological changes in
the heart of I/R-injured rats treated with LBP was examined via
H&E staining (scale bar, 200 µm; magnification, ×100). The
levels of (J) LDH, (K) CK, (L) cTnT, (M) IL-1β, (N) IL-6 and (O)
TNF-α in I/R-injured rats treated with LBP were measured using
ELISAs. **P<0.01, ***P<0.001 vs. Sham; ^P<0.05,
^^P<0.01, ^^^P<0.001 vs. LBP;
##P<0.01, ###P<0.001 vs. I/R. Sham,
rats received sham operation; I/R, ischemia/reperfusion; LBP,
Lycium barbarum polysaccharide; LVEDs, left ventricular
end-systolic diameter; LVEDd, left ventricular end-diastolic
diameter; EF, ejection fraction; FS, fractional shortening; LDH,
lactate dehydrogenase; CK, creatine kinase; AAR, area at risk; INF,
infarcted area; LV, left ventricle. |
Evaluation of rat heart function
After 2 h of reperfusion, transthoracic
echocardiography was performed using an ultrasonic imaging system
(Vevo 1100; FUJIFILM VisualSonics, Inc.). The two-dimensional
B-mode and M-mode traces were collected along the short axis of the
left ventricle. Left ventricular end-systolic diameter (LVEDs),
left ventricular end-diastolic diameter (LVEDd), ejection fraction
(EF) and fractional shortening (FS) were analyzed offline using the
advanced cardiovascular measurements package (Vevo 770;
VisualSonics, Inc.) in a blinded manner.
Evan's blue and
2,3,5-triphenyl-tetrazolium chloride (TTC) double-staining
After cannulation of the aorta, the hearts were
perfused with 1% Evan's blue solution (cat. no. G1810; Beijing
Solarbio Science & Technology Co., Ltd.) for 10 min at 37°C.
After being frozen for 20 min, the hearts were transected into 2-mm
thick sections and then stained with 1% TTC (cat. no. 17779;
Sigma-Aldrich; Merck KGaA) at 37°C for 15 min in PBS. The analyzes
of the size of the infarcted area (INF), area at risk (AAR) and the
whole left ventricle (LV) were performed using ImageJ software
(1.52s version; National Institutes of Health). The infarct size
was presented as a ratio of INF to AAR.
H&E staining
After being fixed with 4% paraformaldehyde (cat. no.
16005; Sigma-Aldrich; Merck KGaA) at room temperature for 24 h,
dehydrated with gradient alcohol and transparentized using xylene
(cat. no. 95682; Sigma-Aldrich; Merck KGaA), the ventricular
tissues were paraffinized (cat. no. 1496904; Sigma-Aldrich; Merck
KGaA) and sliced into 5-µm thick sections, which were then dewaxed
using xylene and rehydrated using gradient alcohol. Hematoxylin
(cat. no. H3136; Sigma-Aldrich; Merck KGaA) was used to stain the
sections at room temperature for 12 min. Next, the sections were
differentiated using hydrochloric alcohol and then stained with
eosin (cat. no. E4009; Sigma-Aldrich; Merck KGaA) at room
temperature for 5 min. Lastly, neutral balsam (cat. no. N861409;
Shanghai Macklin Co., Ltd.) was used to seal the sections. After
being dried at 37°C for 4 h, the sections were observed and
pathologically examined under an inverted microscope (ZEISS
Primovert; Carl Zeiss AG) at ×100 magnification.
Cell culture
Rat cardiomyocytes, H9C2, were obtained from the
American Type Culture Collection (ATCC; cat. no. CRL-1446), and
cultured in ATCC-formulated DMEM (cat. no. 30-2002; ATCC)
supplemented with 10% FBS (cat. no. 30-2020; ATCC) at 37°C with 5%
CO2.
Cell transfection
Nrf2 overexpression plasmid was constructed using a
pcDNA3.1 vector (cat. no. V79520; Thermo Fisher Scientific, Inc.),
and its transfection into H9C2 cells was performed using
Lipofectamine® 3000 transfection reagent (cat. no.
L3000015; Thermo Fisher Scientific, Inc.). Briefly, H9C2 cells were
seeded at a density of 1×104 cells/well in 96-well
plates. When the cells reached 80% confluence, Lipofectamine 3000
transfection reagent (0.15 µl) and Nrf2 overexpression plasmid (0.2
µg) were mixed with Opti-MEM (cat. no. 31985062; Thermo Fisher
Scientific, Inc.) supplemented with P3000 reagent (0.4 µg) and
added into the cells, subsequent to which the cells were incubated
with plasmid-lipid complex at 37°C for 48 h. The empty pcDNA3.1
vector was served as a negative control (NC). After 48 h of
transfection, cells were collected for further experiment.
Myocardial I/R cellular model
construction and drug treatment
LBP (purity, 51.87%) was diluted using PBS in order
to prepare the working solutions with different concentrations (15,
30 and 60 µg/ml). To investigate the effects of LBP on myocardial
I/R injury, H9C2 cells with or without transfection of Nrf2
overexpression plasmid underwent hypoxia via incubation with
glucose-free serum in an anaerobic environment (1% O2,
5% CO2 and 94% N2) at 37°C for 24 h. This was
followed by subjection to the reoxygenation process under normoxic
conditions (21% O2, 5% CO2 and 74%
N2) at 37°C for 4 h. LBP (15, 30 and 60 µg/ml) or
rapamycin (RAPA; an autophagy enhancer; 100 µmol/l; cat. no. 37094;
Sigma-Aldrich; Merck KGaA) were added at the start of reoxygenation
at 37°C during the 4 h (27,28).
To determine the interaction between LBP and RAPA, H9C2 cells
received a combined treatment of LBP (15 µg/ml) and RAPA (100
µmol/l) at the start of reoxygenation at 37°C during the 4 h.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA from H9C2 cells was isolated using
TRIzol® reagent (cat. no. 15596026; Thermo Fisher
Scientific, Inc.). The total RNA used as templates for cDNA was
reverse transcribed into cDNA using the SuperScript™ IV
First-Strand Synthesis System (cat. no. 18091050; Thermo Fisher
Scientific, Inc.) at 37°C for 10 min. The amplification of cDNA was
performed using a PCR detection device (CFX Connect; Bio-Rad
Laboratories, Inc.) with PowerUp SYBR Green Master mix (cat. no.
A25742; Thermo Fisher Scientific, Inc.). Primers for Nrf2 (forward,
5′-AAACCCAGTGTGACCAACGT-3′ and reverse, 5′-GCACACGTGTGGTACTCAGA-3′)
and GAPDH (forward, 5′-TGGATAGGGTGGCCGAAGTA-3′ and reverse,
5′-TACAAGGGGAGCAACAGCTG-3′) were used. The thermocycling conditions
were as follows: Initial denaturation at 95°C for 10 min; 40 cycle
of denaturation at 95°C for 15 sec, and annealing and elongation at
60°C for 60 sec without final extension. The relative expression
levels of genes were calculated using the 2−ΔΔCq method
(29), and GAPDH was used as the
internal reference.
Western blotting
Total protein from H9C2 cells was extracted using
RIPA buffer (cat. no. 89900; Thermo Fisher Scientific, Inc.) and
the concentration was determined using a BCA kit (cat. no. A53227;
Thermo Fisher Scientific, Inc.). The protein (40 µg) and marker (4
µl) (cat. no. PR1910; Beijing Solarbio Science & Technology
Co., Ltd.) were separately loaded and electrophoresed via 12%
SDS-PAGE (cat. no. P0672; Beyotime Institute of Biotechnology),
followed by transfer onto PVDF membranes (cat. no. P2438;
Sigma-Aldrich; Merck KGaA), which were blocked using 5% skim milk
in TBS with 1% Tween-20 (TBST; cat. no. TA-125-TT; Thermo Fisher
Scientific, Inc.) at room temperature for 1 h. The membranes were
then incubated at 4°C overnight with the primary antibodies for
Nrf2 (cat. no. ab89443; 68 kDa; 1:1,000; Abcam), Beclin 1 (cat. no.
ab207612; 52 kDa; 1:2,000; Abcam), LC3-II/LC3-I (cat. no. ab48394;
LC3-II, 17 kDa; LC3-I, 19 kDa; 1:1,000; Abcam), LaminA (cat. no.
4777; 74 kDa; 1:2,000; Cell Signaling Technology, Inc.) and GAPDH
(cat. no. ab181602; 36 kDa; 1:10,000; Abcam). After being washed
with TBST, the membranes were incubated with the HRP-conjugated
secondary antibodies, including goat anti-rabbit IgG (cat. no.
A32731; 1:10,000; Thermo Fisher Scientific, Inc.) or goat
anti-mouse IgG (cat. no. A32733; 1:1,000, Thermo Fisher Scientific,
Inc.). The protein bands were visualized using an ECL reagent kit
(cat. no. WP20005; Thermo Fisher Scientific, Inc.) and the grey
value of each band was analyzed with ImageJ software (1.52s
version; National Institutes of Health).
MTT assay
I/R-treated H9C2 cells were seeded at a density of
1×104 cells/well in 96-well plates with transfection, as
well as treatment of LBP and RAPA in combination or alone. Then, 20
µl MTT solution (cat. no. V900888; Sigma-Aldrich; Merck KGaA) was
added and incubated with the cells at 37°C for 4 h. Next, 100 µl
DMSO (cat. no. D2650; Sigma-Aldrich; Merck KGaA) was used to
dissolve the formazan generated during the incubation. The
absorbance at 570 nm was recorded using a microplate reader
(ELx808; BioTek Instruments, Inc.).
Annexin-V/PI staining assay
Cell apoptosis was determined using an Annexin
V-FITC/PI apoptosis detection kit (cat. no. E-CK-A211; Elabscience
Biotechnology, Inc.). Briefly, the I/R-treated H9C2 cells with
transfection, as well as treatment of LBP and RAPA in combination
or alone, were digested using trypsin, centrifuged at 3,000 × g for
5 min at 4°C and then washed with PBS (twice for the processes of
centrifugation and washing, respectively). Then, the cells were
re-suspended using binding buffer and incubated with Annexin V-FITC
solution (5 µl) and PI solution (10 µl) at room temperature for 15
min in the dark. Lastly, the cells were transferred to a flow
cytometer (Cytoflex; Beckman Coulter, Inc.) and the apoptotic rates
of the cells were analyzed using CytExpert software (version
2.2.0.97; Beckman Coulter, Inc.).
ELISA
The ventricular tissues and H9C2 cells were
centrifuged at 1,000 × g for 20 min at 4°C for the collection of
supernatants. The levels of LDH (cat. no. JL13677-96T), creatine
kinase (CK; cat. no. JL34658-96T), sera cardiac troponin T (cTnT;
cat. no. JL18321-96T), IL-1β (cat. no. JL20884-96T), IL-6 (cat. no.
JL20896-96T), TNF-α (cat. no. JL13202-96T), malondialdehyde (MDA;
cat. no. JL13297-96T) and superoxidase dismutase (SOD; cat. no.
JL22893-96T) in the supernatant were calculated using their
corresponding ELISA kits (Jonln; http://www.jonln.com/), according to the
manufacturer's instructions. Briefly, H9C2 cells with transfection,
as well as treatment of LBP and RAPA in combination or alone, were
centrifuged at 1,000 × g for 20 min at 4°C, and the supernatant was
collected, 50 µl of which was transferred to the enzyme-coated
plates. Then, the HRP-labeled antibody (50 µl) was added into the
plates, which were incubated at 37°C for 60 min. After being washed
with the buffer solution (350 µl) five times, the substrate reagent
was added into the plates and the plates were further incubated at
37°C for 15 min in the dark. The reaction was terminated by the
addition of sulfuric acid (in a final volume of 50 µl) and the
optical density value was recorded with a microplate reader
(ELx808; BioTek Instruments, Inc.) at 450 nm.
Statistical analysis
Statistical analysis was performed using SPSS
software (version 20.0; IBM Corp.). All data are expressed as the
mean ± SD. One-way ANOVA along with Tukey's post hoc test was used
for comparison among multiple groups. P<0.05 was considered to
indicate a statistically significant difference. All experiments
were repeated independently in triplicate.
Results
LBP counteracts I/R-induced heart
malfunction, histopathological abnormalities and inflammation in
rats
The transthoracic echocardiography analysis
demonstrated that I/R-treated rats exhibited increased LVEDs and
declined EF and FS (P<0.01, P<0.001; Fig. 1B-E), while LVEDd remained unchanged.
Moreover, LBP reversed the aforementioned changes (P<0.01,
P<0.001; Fig. 1B-E). The results
of Evan's blue and TTC double staining revealed that LBP
significantly abrogated I/R-induced size increases of the infarct
area, as suggested by the decreased INF/AAR (P<0.001; Fig. 1F-H). The histopathological
examination via H&E staining identified that I/R-induced
myocardial distortion and injury were alleviated by LBP (Fig. 1I).
As I/R can induce cardiomyocyte damage along with
the increase on the activities of enzymes, such as LDH and CK,
inflammatory responses and oxidative stress (30–32),
the levels of both enzymes and these corresponding factors were
quantified in I/R-treated rats. As demonstrated in the analyses
with ELISAs, the levels of LDH, CK, cTnT, IL-1β, IL-6 and TNF-α
were increased after the construction of the I/R model
(P<0.001), while LBP treatment reversed these trends in rats
with I/R (P<0.001; Fig. 1J-O).
Thus, LBP may attenuate I/R-induced effects on these factors.
LBP counteracts the I/R-induced
decrease of viability, promotion of apoptosis, cardiomyocyte
damage, inflammation and oxidative stress in H9C2 cells
The effects of LBP on cardiomyocytes were determined
after the construction of cellular I/R model. Since the
cytoprotective effects of LBP at different concentrations (15, 30
and 60 µg/ml) on oxygen glucose deprivation/reoxygenation-induced
hippocampal neurons have been confirmed previously (27), the current study evaluated whether
LBP could also protect H9C2 cells from I/R-induced injury at these
concentrations. The result of MTT assay demonstrated that the
construction of cellular I/R model decreased the viability of H9C2
cells (P<0.001), the trend of which was partially abrogated by
LBP (15, 30 and 60 µg/ml) treatment (P<0.05, P<0.01; Fig. 2A).
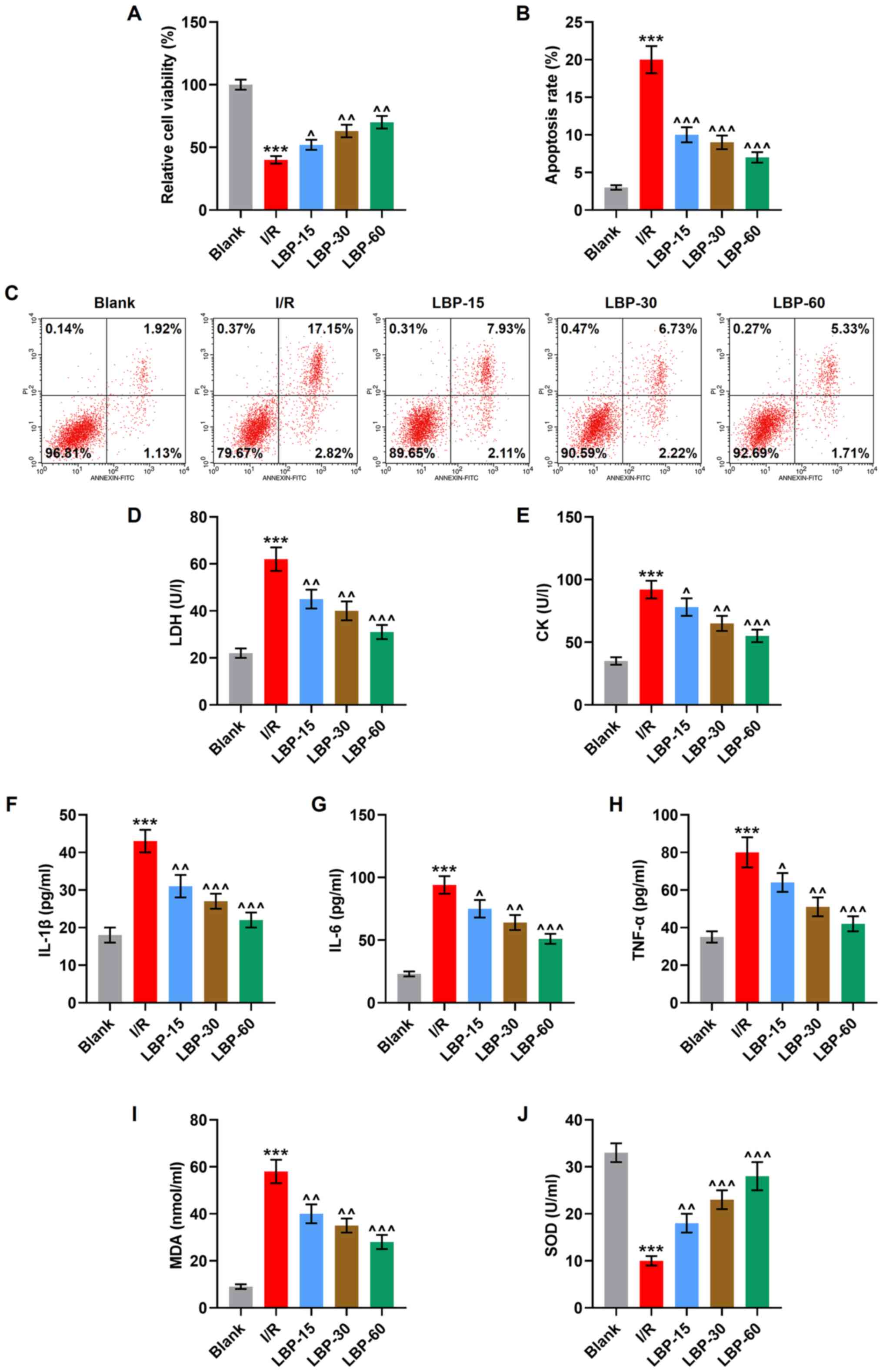 | Figure 2.LBP counteracts I/R-induced decrease
of viability, promotion of apoptosis, cardiomyocyte damage,
inflammation and oxidative stress in H9C2 cells. (A). The viability
of I/R-induced H9C2 cells treated with LBP (15, 30 and 60 µg/ml)
was determined using an MTT assay. (B) The apoptosis of I/R-induced
H9C2 cells treated with LBP (15, 30 and 60 µg/ml) was determined
via (C) flow cytometry. The levels of (D) LDH, (E) CK, (F) IL-1β,
(G) IL-6, (H) TNF-α, (I) MDA and (J) SOD in I/R-induced H9C2 cells
treated with LBP (15, 30 and 60 µg/ml) were measured using ELISAs.
***P<0.001 vs. Blank; ^P<0.05,
^^P<0.01, ^^^P<0.001 vs. I/R. I/R,
ischemia/reperfusion; LBP, Lycium barbarum polysaccharide; LDH,
lactate dehydrogenase; CK, creatine kinase; MDA, malondialdehyde;
SOD, superoxidase dismutase. |
To determine the association between the apoptosis
and the effects of LBP on I/R-treated cardiomyocytes, flow
cytometry was conducted. It was found that a significantly
increased apoptosis occurred after the construction of cellular I/R
model (P<0.001), while LBP (15, 30 and 60 µg/ml) treatment
partially reversed this enhancement induced by I/R (P<0.001;
Fig. 2B and C).
Serum levels of LDH and CK were assayed to assess
cellular damage. The construction of the cellular I/R model was
associated with the increased contents of LDH, CK and MDA, the
enhanced release of IL-1β, IL-6 and TNF-α and decreased SOD
activity (P<0.001), while all of these I/R-induced changes were
partially reversed by the treatment of LBP (15, 30 and 60 µg/ml),
according to the results of ELISAs (P<0.05, P<0.01,
P<0.001; Fig. 2D-J). Taken
together, these results suggested that LBP protects cardiomyocytes
against I/R-induced decrease of viability, promotion of apoptosis,
cardiomyocyte damage, inflammation and oxidative stress.
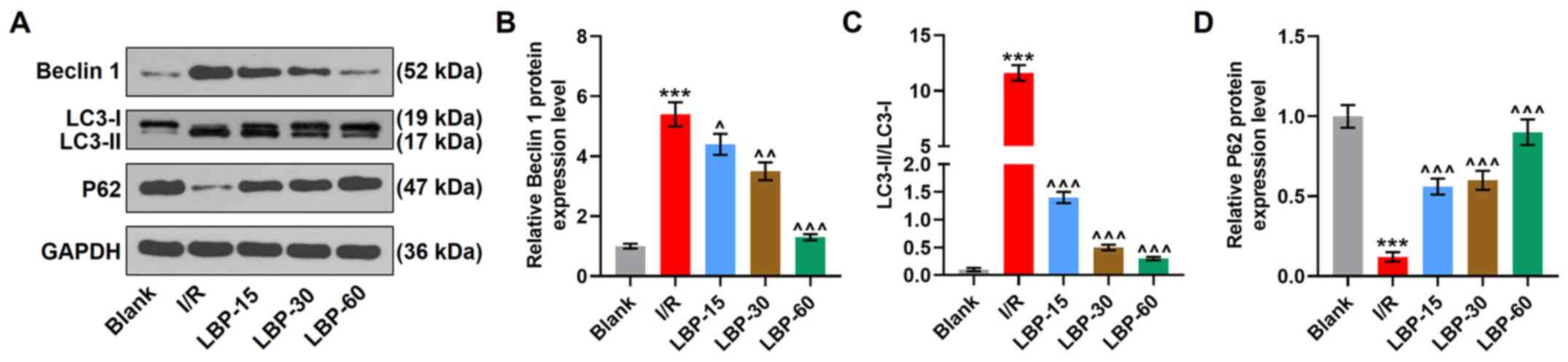
LBP inhibits I/R-induced autophagy in
H9C2 cells
Autophagy in cardiomyocytes is demonstrated to be
promoted in an anoxia duration-dependent manner in response to
anoxia/reperfusion (28).
Therefore, to investigate whether the protective effects of LBP on
cardiomyocytes against I/R-induced injury were associated with the
modulation of autophagy, the current study analyzed the expression
levels of autophagy-related markers. The western blotting results
revealed that the construction of the cellular I/R model increased
the protein expression levels of Beclin 1 and LC3-II/LC3-I, and
decreased those of P62 (P<0.001), whereas treatment of LBP at
60, 30 and 15 µg/ml was able to decrease the protein expression
levels of Beclin 1 and LC3-II/LC3-I and to increase those of P62 in
H9C2 cells after 24-h anoxia and 4-h reperfusion (P<0.05,
P<0.01, P<0.001; Fig. 3A-D),
indicating that LBP inhibits I/R-induced autophagic activity.
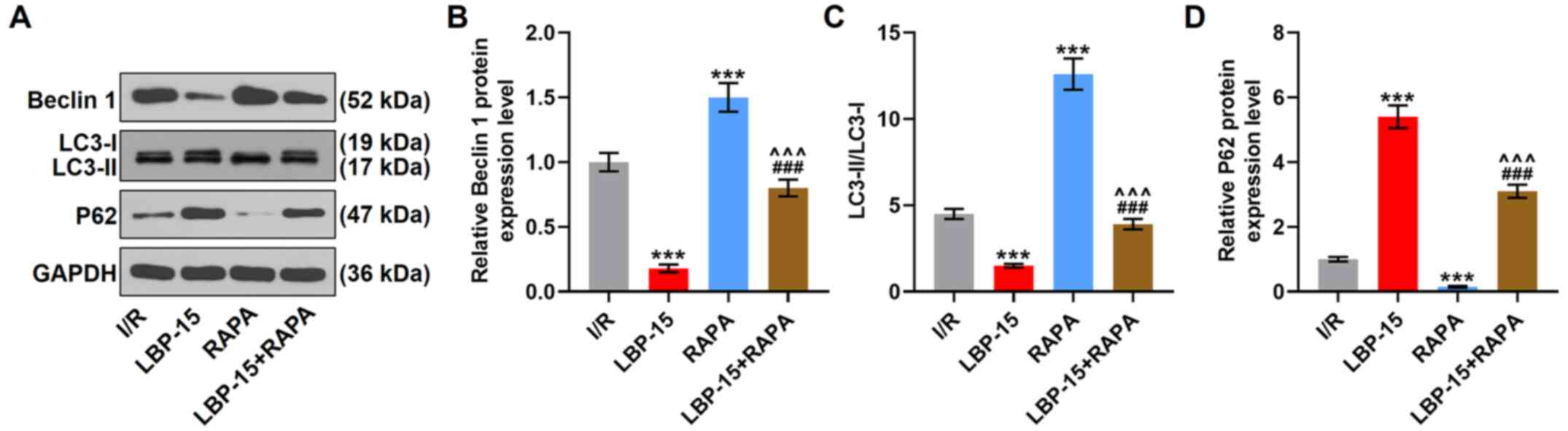
RAPA partially reverses the inhibitory
effect of LBP on autophagy in I/R-treated H9C2 cells
To verify if the inhibition of autophagy was
involved in the cytoprotective effects of LBP on I/R-induced
cardiomyocytes, an autophagy promoter, RAPA, and the lowest
concentration (15 µg/ml) of LBP were used to determine the
interaction between autophagy and LBP. RAPA treatment alone
increased the protein expression levels of Beclin 1 and
LC3-II/LC3-I and decreased those of P62, compared with the I/R
group (P<0.001). Moreover, LBP (15 µg/ml) decreased the
expression levels of Beclin 1 and LC3-II/LC3-I and the increased
expression of P62 in I/R-induced H9C2 cells (P<0.001; Fig. 4A-D). It was also found that LBP (15
µg/ml) treatment reversed RAPA-induced expression changes of these
genes (P<0.001; Fig. 4A-D).
RAPA partially reverses LBP-induced
increase of viability and inhibition of apoptosis, cardiomyocyte
damage, inflammation and oxidative stress in I/R-treated H9C2
cells
Furthermore, based on the results of MTT assay and
flow cytometry, it was observed that RAPA treatment alone decreased
cell viability and increased apoptosis, compared with the I/R group
(P<0.001), and partially reversed the LBP (15 µg/ml)-induced
increase in viability and inhibition of apoptosis in I/R-induced
H9C2 cells (P<0.01, P<0.001; Fig.
5A-C). The results of ELISAs showed that after RAPA treatment,
the levels of LDH, CK, IL-1β, IL-6, TNF-α and MDA were elevated,
while that of SOD was lowered, in comparison with those in the I/R
group (P<0.001). Moreover, the LBP (15 µg/ml)-induced changes on
the levels of these factors were partially reversed (P<0.05,
P<0.01; Fig. 5D-J) by RAPA
treatment.
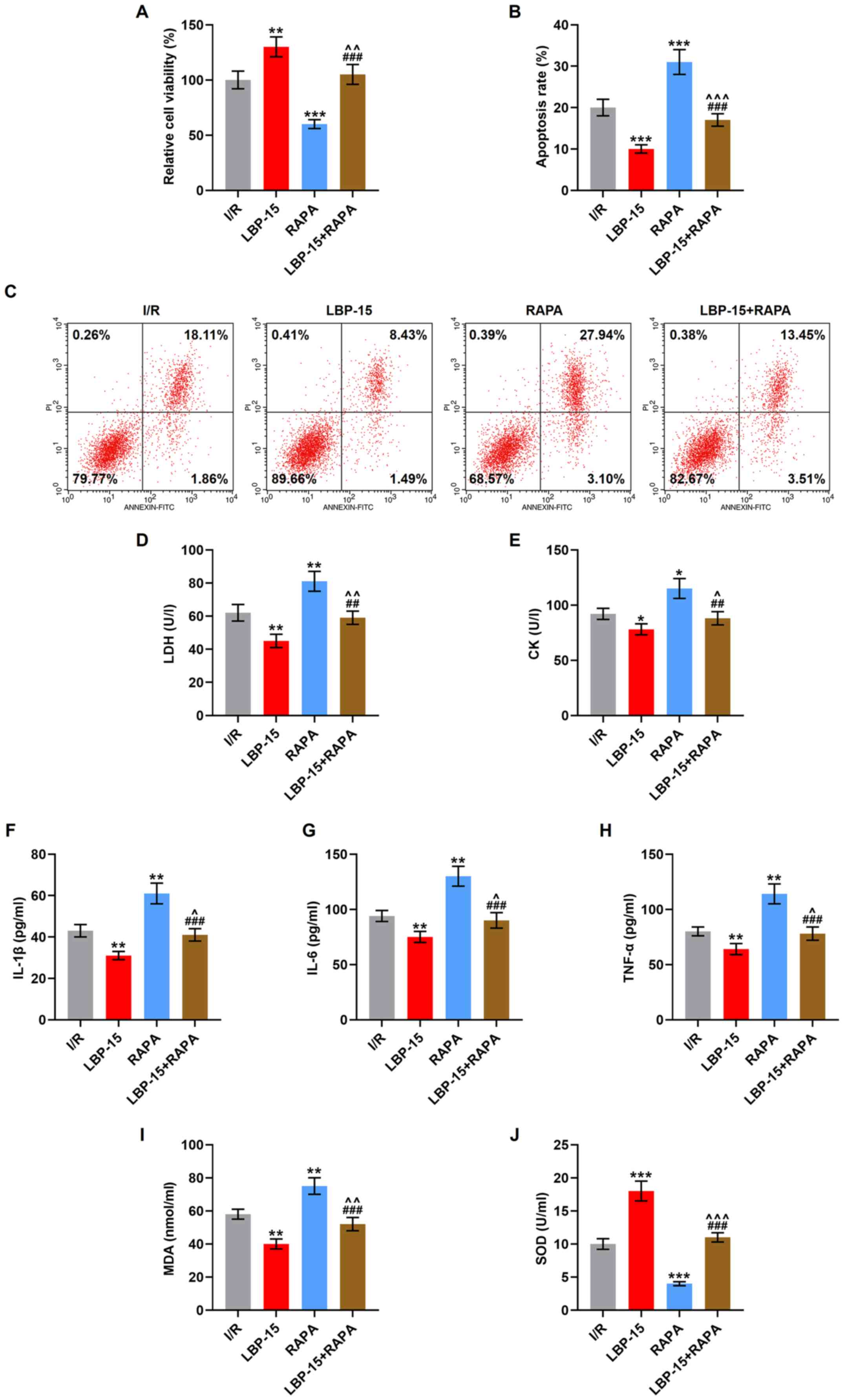 | Figure 5.RAPA partially reverses the
LBP-induced increase of viability and inhibition of apoptosis,
cardiomyocyte damage, inflammation and oxidative stress in
I/R-induced H9C2 cells. (A) The viability of I/R-induced H9C2 cells
with the treatment of LBP (15 µg/ml) and RAPA in combination or
alone was determined using an MTT assay. (B) The apoptosis of
I/R-induced H9C2 cells with the treatment of LBP (15 µg/ml) and
RAPA in combination or alone was detected via (C) flow cytometry.
The levels of (D) LDH, (E) CK, (F) IL-1β, (G) IL-6, (H) TNF-α, (I)
MDA and (J) SOD in I/R-induced H9C2 cells with the treatment of LBP
(15 µg/ml) and RAPA in combination or alone were determined via
ELISAs. *P<0.05, **P<0.01, ***P<0.001 vs. I/R;
^P<0.05, ^^P<0.01,
^^^P<0.001 vs. LBP-15; ##P<0.01,
###P<0.001 vs. RAPA. I/R, ischemia/reperfusion; LBP,
Lycium barbarum polysaccharide; RAPA, rapamycin; LDH, lactate
dehydrogenase; CK, creatine kinase; MDA, malondialdehyde; SOD,
superoxidase dismutase. |
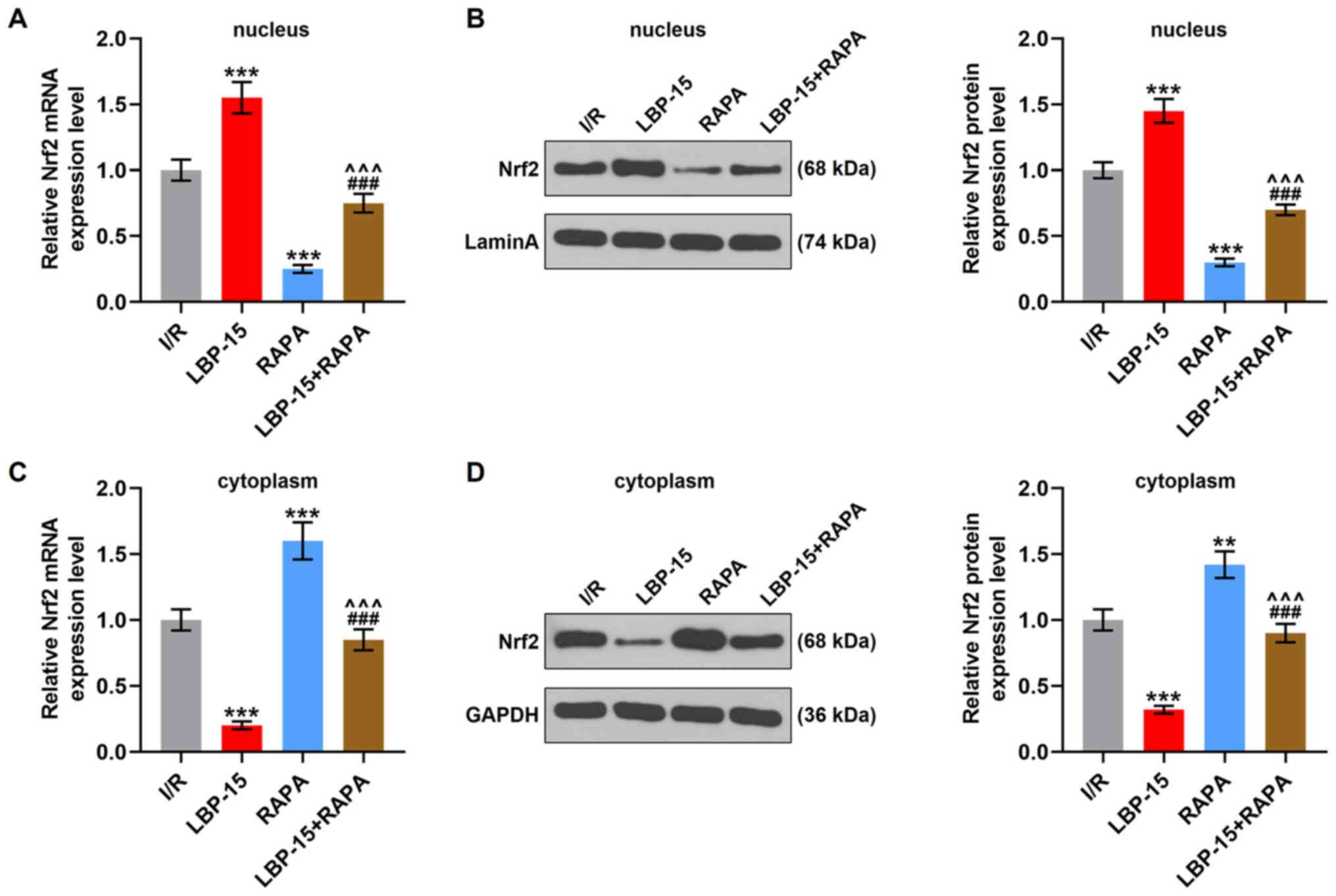
RAPA reverses LBP-induced promotion on
Nrf2 nuclear translocation in I/R-induced H9C2 cells
A previous study reported the increase of Nrf2
expression during autophagic deficiency (22). As shown in Fig. 6A and B, Nrf2 mRNA and protein
expression levels in the nucleus were increased with the treatment
of LBP (15 µg/ml) treatment (P<0.001), but were decreased by
RAPA treatment (P<0.001). On the contrary, LBP treatment (15
µg/ml) was associated with a decreased Nrf2 mRNA and protein
expression levels in the cytoplasm (P<0.001), whilst an opposite
trend was observed after RAPA treatment (P<0.01, P<0.001;
Fig. 6C and D). These results
indicated that LBP facilitated the translocation of Nrf2 into the
nucleus, while RAPA had an opposite effect and partially reversed
the effects of LPB on the expression levels of both nuclear and
cytoplasmic Nrf2 (P<0.001; Fig.
6A-D), suggesting that LPB may inhibit I/R-induced autophagy by
facilitating the nuclear translocation of Nrf2.
Nrf2 partially abrogates RAPA-induced
decrease of viability, promotion of apoptosis, cardiomyocyte
damage, inflammation and oxidative stress in I/R-treated H9C2
cells
The transfection efficiency of Nrf2 overexpression
plasmid in normal H9C2 cells is showed in Fig. 7A. The results demonstrated that the
Nrf2 overexpression plasmid significantly increased Nrf2 expression
in H9C2 cells (P<0.001).
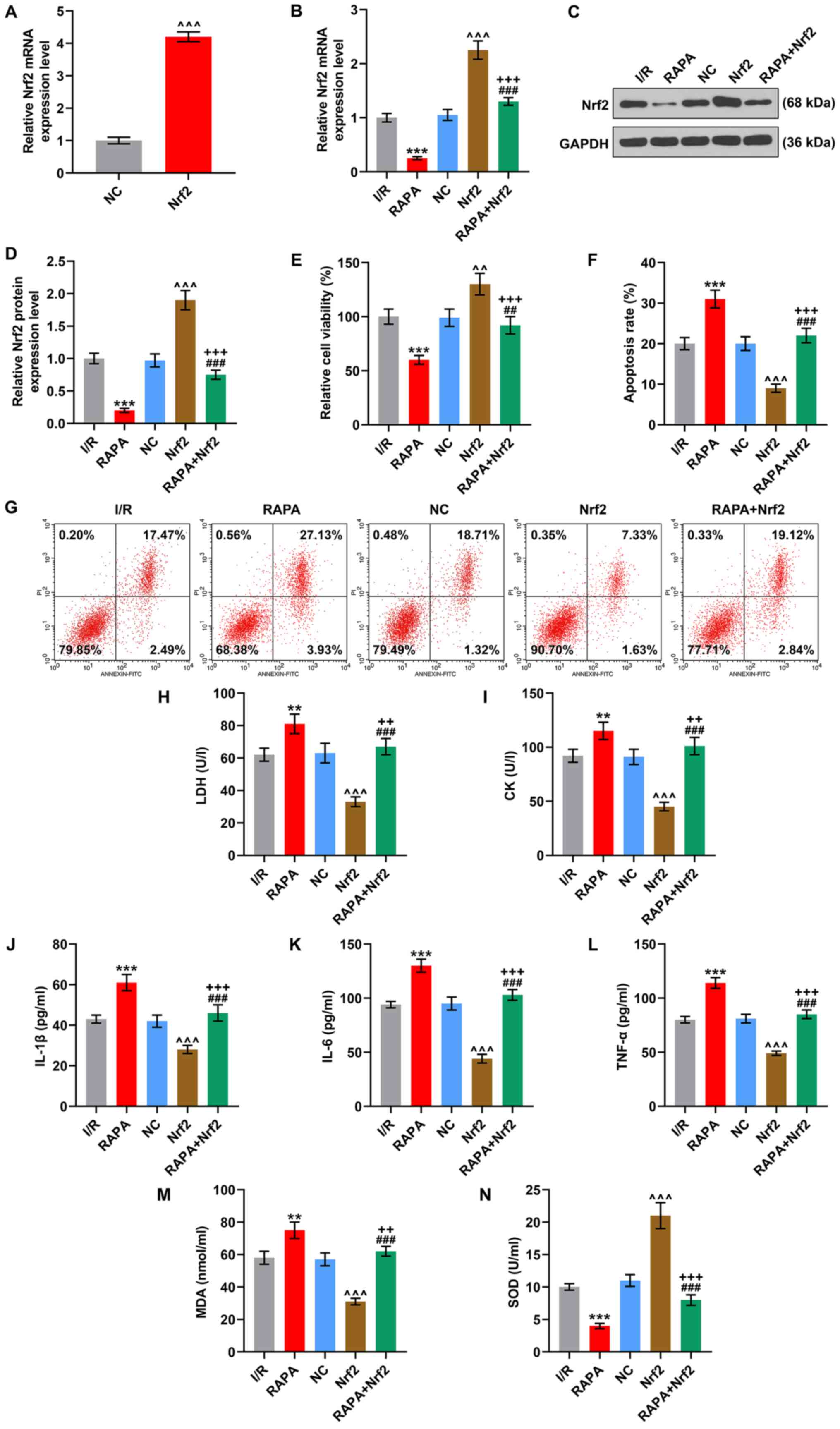 | Figure 7.Nrf2 partially reverses RAPA-induced
decrease of viability, promotion of apoptosis, cardiomyocyte
damage, inflammation and oxidative stress in I/R-induced H9C2
cells. (A) The transfection efficiency of Nrf2 overexpression
plasmid in normal H9C2 cells. The (B) mRNA and (C and D) protein
expression levels of Nrf2 in I/R-induced H9C2 cells with
transfection of Nrf2 overexpression plasmid, RAPA treatment or the
combined management were quantified by reverse
transcription-quantitative PCR and western blotting. GAPDH was used
as internal reference. (E) The viability of I/R-induced H9C2 cells
undergoing transfection of Nrf2 overexpression plasmid, RAPA
treatment or the combined management was determined using an MTT
assay. (F) The apoptosis of I/R-induced H9C2 cells after the
transfection of Nrf2 overexpression plasmid, RAPA treatment or the
combined management was detected via (G) flow cytometry. The levels
of (H) LDH, (I) CK, (J) IL-1β, (K) IL-6, (L) TNF-α, (M) MDA and (N)
SOD in I/R-induced H9C2 cells undergoing transfection of Nrf2
overexpression plasmid, RAPA treatment or the combined management
were calculated using ELISAs. **P<0.01, ***P<0.001 vs. I/R;
^^P<0.01, ^^^P<0.001 vs. NC;
++P<0.01, +++P<0.001 vs. RAPA;
##P<0.01, ###P<0.001 vs. Nrf2. NC,
empty pcDNA3.1 vector as negative control; Nrf2, nuclear
factor-erythroid factor 2-related factor 2; I/R,
ischemia/reperfusion; LBP, Lycium barbarum polysaccharide; RAPA,
rapamycin; LDH, lactate dehydrogenase; CK, creatine kinase; MDA,
malondialdehyde; SOD, superoxidase dismutase. |
Subsequently, the Nrf2 overexpression plasmid was
introduced into I/R-induced and RAPA-treated cardiomyocytes to
determine the interaction between Nrf2 and autophagy. Firstly, in
comparison with the I/R group, the transfection of Nrf2
overexpression plasmid increased the mRNA and protein expression
levels of Nrf2, while RAPA treatment led to a decrease (P<0.001;
Fig. 7B and C). Furthermore, the
effects of Nrf2 overexpression plasmid on Nrf2 expression was
inhibited by RAPA (P<0.001; Fig. 7B
and C). Then, it was observed that Nrf2 overexpression
increased cell viability, inhibited apoptosis, decreased the
contents of LDH, CK and MDA, suppressed the release of IL-1β, IL-6
and TNF-α, and enhanced SOD activity, compared with the I/R group
(P<0.001; Fig. 7D-M). However,
these effects induced by Nrf2 overexpression were partially
reversed by RAPA treatment (P<0.01; Fig. 7E-N).
Discussion
Myocardial I/R injury causes structural and
functional changes at the levels of organs, tissues and cells,
which leads to cellular destruction (33). During the progression of I/R,
ischemia causes the accumulation of H+ and
Ca2+, the collapse of mitochondrial membrane potential
and the depletion of ATP, leading to the formation of ROS, which in
turn activates pro-inflammatory pathways and promotes inflammatory
responses (34). The promoted
inflammatory responses further induce damage to the tissues
surrounding the infarct area by enhancing apoptosis within the
cardiomyocyte (34). LBP is a
cardio-protective agent with functions that counteract I/R-induced
LDH release, inhibition of ATPase activity and promotion of
apoptosis in myocardial I/R injury (10). The present study demonstrated that
LBP modulated autophagy marker expression so as to exert the
protective effects on I/R-treated rats and cardiomyocytes.
Decreased viability and enhanced apoptosis occur in
hypoxia/reperfusion (H/R)-treated cardiomyocytes, which is
experimentally used to construct a model of myocardial I/R injury
at cellular levels (35). A
previous study reported that LBP could partially reverse the
effects of H/R on decreasing the viability and promoting the
apoptosis of H9C2 cells (36). The
current study established I/R models both in vivo and in vitro,
during which LBP treatment at 15, 30 and 60 µg/ml was used. The
results demonstrated that LBP counteracted I/R-induced heart
malfunction and histopathological abnormalities in vivo. Moreover,
in line with a previous finding concerning the effects LBP on the
phenotypic changes of I/R-induced cardiomyocytes (36), the present study observed that LBP
exerted antagonistic effects against the decreased viability and
the increased apoptosis in H/R-induced H9C2 cells.
CK serves a crucial role in regulating ATP
preservation, which is positively correlated with an improved
contractile function in I/R-induced rats (37,38).
During the progression of I/R, the release of CK, as well as LDH
and cTnT, two main inducible factors of myocardial injury, from the
myocardium into the blood are increased. As such, inhibiting the
releases of LDH and CK can exert a cardio-protective effect on
I/R-induced ischemic hearts of rats (32). Furthermore, suppressing inflammatory
cytokines (IL-1β, IL-6 and TNF-α) produced during I/R-induced
inflammatory responses in the myocardium is also contributory to
cardio-protection against I/R injury (39). I/R also induces the production of
excessive ROS, which then oxidizes cellular proteins, membrane
lipids and nucleic acids (40) and
further leads to the aggravation of myocardial I/R injury, with
increased MDA level and decreased SOD (35). Decreased MDA and increased SOD
levels have been indicated to be associated with ameliorated
myocardial I/R injury (41). A
prior study has revealed that LBP, as an oxidative agent, reduces
cardiotoxicity induced by doxorubicin and leads to the decreased CK
and MDA levels and increased those of SOD (42). LBP can also decrease the expression
levels of inflammatory factors (IL-6 and TNF-α) in heart tissues,
based on the results of a study where the effects of LBP on cardiac
hypertrophy in diabetic rats were discussed (16). Similar to the effects of LBP shown
in these findings, the present study identified that LBP
counteracted the effects of I/R both in vivo and in vitro by
abolishing the I/R-induced changes on the levels of all these
aforementioned indexes or factors, suggesting that LBP alleviated
the cell injury, inflammatory responses and oxidative stress in
I/R-induced cardiomyocytes.
Autophagy preserves cardiac structure and function
at baseline by inhibiting misfolded proteins and damaged organelles
in physiological condition, and is further activated during stress,
thereby limiting cardiac injury in numerous pathological conditions
(43,44). For example, autophagy maintains
cardiac function under ischemia and starvation conditions, and
reduces myocardial damage (45).
However, an excessive activation in autophagy under
ischemia/reperfusion has been reported, where autophagy may
facilitate myocardial injury (46–49).
Moreover, an increase in autophagy activity has been observed in
the subacute and chronic stages of cardiac ischemia in a mouse
model with myocardial infarction (50,51).
Autophagy is a mechanism induced by the oxidative stress in
myocardial I/R injury and causes the death of cardiomyocyte
(4). Pharmacological inhibition of
autophagy can help prevent I/R-induced heart failure by improving
cardio-function and alleviating myocardial injury by reversely
regulating I/R-induced increases of LDH, CK, ROS and MDA and
decreases of SOD (21). Thus,
appropriate inhibition of autophagy could be a novel therapeutic
method for I/R injury.
Hence, in the present study, RAPA, an activator of
autophagy, was hypothesized to abolish the cardio-protective
effects of LBP against I/R injury. LBP was previously found to
counteract the oxygen glucose deprivation/reperfusion-induced
upregulated expression of LC3-II/LC3-I and Beclin 1 and
downregulated expression of P62 to suppress autophagic cell death
in primary hippocampal neurons (27). In accordance with this finding in
which the autophagy-repressive effect of LBP has been indicated,
the present study demonstrated that LBP counteracted the effects of
I/R on decreasing the expression of LC3-II/LC3-I and Beclin 1 and
increasing that of P62. More notably, LBP exerted inhibitory
effects on RAPA-potentiated autophagy in I/R-treated H9C2 cells,
while RAPA partially reversed the suppressive effects of LBP on
autophagy, which collectively indicated that the cardio-protective
effects of LBP against I/R-induced injury is, at least partially,
dependent on the inhibition of autophagy.
Inhibited Nrf2 expression is evidenced in
PM2.5-induced autophagy of A549 cells, according to a study by Dai
et al (22). Similar to
Dai's findings, the present study identified that both nuclear
translocation and nuclear expression of Nrf2 were inhibited by
RAPA-induced autophagy in I/R-treated cardiomyocytes, which
suggests that LBP-induced autophagy inhibition can lead to Nrf2
activation. Furthermore, previous studies have shown that the
activation of Nrf2 induced by LBP reduces ROS production and lipid
peroxide level, but elevates SOD levels in human skin fibroblast
cells (52), as well as attenuates
hyperoxic acute lung injury by inhibiting the infiltration of IL-1β
and IL-6 into the lungs of mice (53). Likewise, the present study revealed
that LBP facilitated the nuclear translocation of Nrf2.
Nrf2, a member of the cap ‘n’ collar subfamily of
basic-region leucine zipper (54),
is involved in the regulation of antioxidant responses by inducing
the expression of antioxidative genes, such as heme oxygenase-1,
glutathione peroxidase and SOD (55). The activation of Nrf2 can exert a
cardio-protective effect via both the suppression of oxidization
and inflammation, and the potential regulation of autophagy
(56). Increased Nrf2 expression in
the nucleus is associated with the alleviation of H/R-induced
impaired viability and enhanced apoptosis of cardiomyocytes
(57), as well as improved cardiac
function, reduced myocardial infarction area, decreased levels of
CK, LDH and MDA and increased SOD levels in I/R-treated rats
(58). Moreover, it is associated
with the inhibited production of IL-6 and TNF-α (59). Based on the present results, it was
found that Nrf2 expression inhibited apoptosis and the levels of
CK, LDH, IL-1β, IL-6, TNF-α and MDA, and increased SOD levels; the
effects of which were similar to the aforementioned studies. Taken
together, the current results indicated that LBP inhibited
autophagy to activate Nrf2 so as to exert its cardio-protective
effects against I/R-induced injury.
However, there are several potential limitations to
the present study. For instance, there was lack of more direct
evidence to confirm autophagy using imaging tools, such as scanning
electron microscopy and transmission electron microscopy. Most
polysaccharides cannot be completely digested by the human
digestive system, so the natural polysaccharides mainly exert their
benefits on health by slowing gastric emptying, regulating the gut
microbe structure, influencing microbial fermentation as
substrates, improving bowel function, as well as protecting the
immune system (60). As a natural
polysaccharide, LPB may also not be digested completely by the
human alimentary system. Natural polysaccharides are favorable for
the proliferation of short chain fatty acid-producing bacteria, the
presence of which can improve the intestinal microenvironment
(61). For example, fermented
Yupingfeng polysaccharides can effectively improve the intestinal
flora homeostasis (62).
Additionally, modulating the gut microbiota is a potential
treatment for cardiovascular diseases (63). A previous study reported that LBP
reduced intestinal permeability and inflammatory cytokine levels,
maintained a healthy intestinal microenvironment, and alleviated
myocardial injury 13). Therefore, the mechanism of the protective
effect of LBP on myocardial I/R injury may be associated with its
role in the regulation of intestinal microenvironment
In conclusion, the present study identified that LBP
counteracts H/R-induced decrease of viability, promotion of
apoptosis, inflammation and oxidative stress in cardiomyocytes by
inhibiting autophagy via the activation of Nrf2, thus attenuating
myocardial I/R injury.
Acknowledgements
Not applicable.
Funding
This work was supported by the Zhejiang Province
Science Research Foundation of Traditional Chinese Medicine (grant
no. 2020ZB152).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
HP made substantial contributions to conception and
design, drafted the article and critically revised the manuscript
for important intellectual content. LN, YW, LC, XZ and YZ were
responsible for data acquisition, analysis and interpretation. HP
and YZ confirm the authenticity of all the raw data. All authors
have read and approved the final version of the manuscript. All
authors agree to be accountable for all aspects of the work in
ensuring that questions related to the accuracy or integrity of the
work are appropriately investigated and resolved.
Ethics approval and consent to
participate
All animal experiments were performed in accordance
with the guidelines for the care and use of laboratory animals
(National Institutes of Health). This study was approved by the
Committee of Experimental Animals of Zhejiang Province Animal
Center (approval no. DC201901203). Every effort was made to
minimize pain and discomfort to the animals. The animals
experiments were performed in Zhejiang Province Animal Center.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Yellon DM and Hausenloy DJ: Myocardial
reperfusion injury. N Engl J Med. 357:1121–1135. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lindsey ML, Bolli R, Canty JM Jr, Du XJ,
Frangogiannis NG, Frantz S, Gourdie RG, Holmes JW, Jones SP, Kloner
RA, et al: Guidelines for experimental models of myocardial
ischemia and infarction. Am J Physiol Heart Circ Physiol.
314:H812–H838. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Esposito ML, Zhang Y, Qiao X, Reyelt L,
Paruchuri V, Schnitzler GR, Morine KJ, Annamalai SK, Bogins C,
Natov PS, et al: Left ventricular unloading before reperfusion
promotes functional recovery after acute myocardial infarction. J
Am Coll Cardiol. 72:501–514. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wu MY, Yiang GT, Liao WT, Tsai AP, Cheng
YL, Cheng PW, Li CY and Li CJ: Current mechanistic concepts in
ischemia and reperfusion injury. Cell Physiol Biochem.
46:1650–1667. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kunecki M, Płazak W, Podolec P and Gołba
KS: Effects of endogenous cardioprotective mechanisms on
ischemia-reperfusion injury. Postepy Hig Med Dosw. 71:20–31. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Galagudza MM, Blokhin IO, Shmonin AA and
Mischenko KA: Reduction of myocardial ischemia-reperfusion injury
with pre- and postconditioning: Molecular mechanisms and
therapeutic targets. Cardiovasc Hematol Disord Drug Targets.
8:47–65. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yu P, Zhang J, Yu S, Luo Z, Hua F, Yuan L,
Zhou Z, Liu Q, Du X, Chen S, et al: Protective effect of
sevoflurane postconditioning against cardiac ischemia/reperfusion
injury via ameliorating mitochondrial impairment, oxidative stress
and rescuing autophagic clearance. PLoS One. 10:e01346662015.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Li J, Xiang X, Gong X, Shi Y, Yang J and
Xu Z: Cilostazol protects mice against myocardium
ischemic/reperfusion injury by activating a PPARγ/JAK2/STAT3
pathway. Biomed Pharmacother. 94:995–1001. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Weinreuter M, Kreusser MM, Beckendorf J,
Schreiter FC, Leuschner F, Lehmann LH, Hofmann KP, Rostosky JS,
Diemert N, Xu C, et al: CaM Kinase II mediates maladaptive
post-infarct remodeling and pro-inflammatory chemoattractant
signaling but not acute myocardial ischemia/reperfusion injury.
EMBO Mol Med. 6:1231–1245. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hou YM, Wang J and Zhang XZ: Lycium
barbarum polysaccharide exhibits cardioprotection in an
experimental model of ischemia-reperfusion damage. Mol Med Rep.
15:2653–2658. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Varoni MV, Pasciu V, Gadau SD, Baralla E,
Serra E, Palomba D and Demontis MP: Possible antioxidant effect of
Lycium barbarum polysaccharides on hepatic cadmium-induced
oxidative stress in rats. Environ Sci Pollut Res Int. 24:2946–2955.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bo R, Zheng S, Xing J, Luo L, Niu Y, Huang
Y, Liu Z, Hu Y, Liu J, Wu Y, et al: The immunological activity of
Lycium barbarum polysaccharides liposome in vitro and adjuvanticity
against PCV2 in vivo. Int J Biol Macromol. 85:294–301. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang XJ, Yu HY, Cai YJ and Ke M: Lycium
barbarum polysaccharides inhibit proliferation and migration of
bladder cancer cell lines BIU87 by suppressing Pi3K/AKT pathway.
Oncotarget. 8:5936–5942. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li SY, Yang D, Yeung CM, Yu WY, Chang RC,
So KF, Wong D and Lo AC: Lycium barbarum polysaccharides reduce
neuronal damage, blood-retinal barrier disruption and oxidative
stress in retinal ischemia/reperfusion injury. PLoS One.
6:e163802011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Xiao J, Xing F, Huo J, Fung ML, Liong EC,
Ching YP, Xu A, Chang RC, So KF and Tipoe GL: Lycium barbarum
polysaccharides therapeutically improve hepatic functions in
non-alcoholic steatohepatitis rats and cellular steatosis model.
Sci Rep. 4:55872014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu Q, Han Q, Lu M, Wang H and Tang F:
Lycium barbarum polysaccharide attenuates cardiac hypertrophy,
inhibits calpain-1 expression and inhibits NF-κB activation in
streptozotocin-induced diabetic rats. Exp Ther Med. 18:509–516.
2019.PubMed/NCBI
|
|
17
|
Levine B and Kroemer G: Autophagy in the
pathogenesis of disease. Cell. 132:27–42. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Xie Z and Klionsky DJ: Autophagosome
formation: Core machinery and adaptations. Nat Cell Biol.
9:1102–1109. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tanida I, Ueno T and Kominami E: LC3
conjugation system in mammalian autophagy. Int J Biochem Cell Biol.
36:2503–2518. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ma S, Wang Y, Chen Y and Cao F: The role
of the autophagy in myocardial ischemia/reperfusion injury. Biochim
Biophys Acta. 1852:271–276. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wu S, Chang G, Gao L, Jiang D, Wang L, Li
G, Luo X, Qin S, Guo X and Zhang D: Trimetazidine protects against
myocardial ischemia/reperfusion injury by inhibiting excessive
autophagy. J Mol Med (Berl). 96:791–806. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Dai P, Shen D, Shen J, Tang Q, Xi M, Li Y
and Li C: The roles of Nrf2 and autophagy in modulating
inflammation mediated by TLR4-NFκB in A549 cell exposed to layer
house particulate matter 2.5 (PM2.5). Chemosphere. 235:1134–1145.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
National Research Council. 2011, . Guide
for the Care and Use of Laboratory Animals. Eighth Edition.
Washington, DC: The National Academies Press; https://doi.org/10.17226/12910
|
|
24
|
Liu SY, Chen L, Li XC, Hu QK and He LJ:
Lycium barbarum polysaccharide protects diabetic peripheral
neuropathy by enhancing autophagy via mTOR/p70S6K inhibition in
Streptozotocin-induced diabetic rats. J Chem Neuroanat. 89:37–42.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Du M, Hu X, Kou L, Zhang B and Zhang C:
Lycium barbarum Polysaccharide Mediated the Antidiabetic and
Antinephritic Effects in Diet-Streptozotocin-Induced Diabetic
Sprague Dawley Rats via Regulation of NF-κB. BioMed Res Int.
2016:31402902016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhao Y, Guo R, Li L, Li S, Fan G, Zhao X
and Wang Y: Tongmai formula improves cardiac function via
regulating mitochondrial quality control in the myocardium with
ischemia/reperfusion injury. Biomed Pharmacother. 132:1108972020.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yu Y, Wu X, Pu J, Luo P, Ma W, Wang J, Wei
J, Wang Y and Fei Z: Lycium barbarum polysaccharide protects
against oxygen glucose deprivation/reoxygenation-induced apoptosis
and autophagic cell death via the PI3K/Akt/mTOR signaling pathway
in primary cultured hippocampal neurons. Biochem Biophys Res
Commun. 495:1187–1194. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Xu Q, Li X, Lu Y, Shen L, Zhang J, Cao S,
Huang X, Bin J and Liao Y: Pharmacological modulation of autophagy
to protect cardiomyocytes according to the time windows of
ischaemia/reperfusion. Br J Pharmacol. 172:3072–3085. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wallert M, Ziegler M, Wang X, Maluenda A,
Xu X, Yap ML, Witt R, Giles C, Kluge S, Hortmann M, et al:
α-Tocopherol preserves cardiac function by reducing oxidative
stress and inflammation in ischemia/reperfusion injury. Redox Biol.
26:1012922019. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wu T, Jiang N, Ji Z and Shi G: The IRE1
signaling pathway is involved in the protective effect of low-dose
LPS on myocardial ischemia-reperfusion injury. Life Sci.
231:1165692019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Amani M, Jeddi S, Ahmadiasl N, Usefzade N
and Zaman J: effect of HEMADO on level of CK-MB and LDH enzymes
after ischemia/reperfusion injury in isolated rat heart.
Bioimpacts. 3:101–104. 2013.PubMed/NCBI
|
|
33
|
Lee YR, Chen SH, Lin CY, Chao WY, Lim YP,
Yu HI and Lu CH: In vitro antitumor activity of aloperine on human
thyroid cancer cells through caspase-dependent apoptosis. Int J Mol
Sci. 19:3122018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Frank A, Bonney M, Bonney S, Weitzel L,
Koeppen M and Eckle T: Myocardial ischemia reperfusion injury: From
basic science to clinical bedside. Semin Cardiothorac Vasc Anesth.
16:123–132. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhang W, Li Y and Wang P: Long non-coding
RNA-ROR aggravates myocardial ischemia/reperfusion injury. Braz J
Med Biol Res. 51:e65552018. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Li Q, Zhang Z, Li H, Pan X, Chen S, Cui Z,
Ma J, Zhou Z and Xing B: Lycium barbarum polysaccharides protects
H9c2 cells from hypoxia-induced injury by down-regulation of
miR-122. Biomed Pharmacother. 110:20–28. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Cao F, Zervou S and Lygate CA: The
creatine kinase system as a therapeutic target for myocardial
ischaemia-reperfusion injury. Biochem Soc Trans. 46:1119–1127.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Wang DS, Yan LY, Yang DZ, Lyu Y, Fang LH,
Wang SB and Du GH: Formononetin ameliorates myocardial
ischemia/reperfusion injury in rats by suppressing the
ROS-TXNIP-NLRP3 pathway. Biochem Biophys Res Commun. 525:759–766.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Bai Y, Li Z, Liu W, Gao D, Liu M and Zhang
P: Biochanin A attenuates myocardial ischemia/reperfusion injury
through the TLR4/NF-κB/NLRP3 signaling pathway. Acta Cir Bras.
34:e2019011042019. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Balakumar P and Sharma NK: Healing the
diabetic heart: Does myocardial preconditioning work? Cell Signal.
24:53–59. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Yu L, Gong B, Duan W, Fan C, Zhang J, Li
Z, Xue X, Xu Y, Meng D, Li B, et al: Melatonin ameliorates
myocardial ischemia/reperfusion injury in type 1 diabetic rats by
preserving mitochondrial function: Role of AMPK-PGC-1α-SIRT3
signaling. Sci Rep. 7:413372017. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Xin YF, Wan LL, Peng JL and Guo C:
Alleviation of the acute doxorubicin-induced cardiotoxicity by
Lycium barbarum polysaccharides through the suppression of
oxidative stress. Food Chem Toxicol. 49:259–264. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Nakai A, Yamaguchi O, Takeda T, Higuchi Y,
Hikoso S, Taniike M, Omiya S, Mizote I, Matsumura Y, Asahi M, et
al: The role of autophagy in cardiomyocytes in the basal state and
in response to hemodynamic stress. Nat Med. 13:619–624. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Ikeda Y, Shirakabe A, Maejima Y, Zhai P,
Sciarretta S, Toli J, Nomura M, Mihara K, Egashira K, Ohishi M, et
al: Endogenous Drp1 mediates mitochondrial autophagy and protects
the heart against energy stress. Circ Res. 116:264–278. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Sciarretta S, Zhai P, Shao D, Maejima Y,
Robbins J, Volpe M, Condorelli G and Sadoshima J: Rheb is a
critical regulator of autophagy during myocardial ischemia:
Pathophysiological implications in obesity and metabolic syndrome.
Circulation. 125:1134–1146. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Matsui Y, Takagi H, Qu X, Abdellatif M,
Sakoda H, Asano T, Levine B and Sadoshima J: Distinct roles of
autophagy in the heart during ischemia and reperfusion: Roles of
AMP-activated protein kinase and Beclin 1 in mediating autophagy.
Circ Res. 100:914–922. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Wei C, Li H, Han L, Zhang L and Yang X:
Activation of autophagy in ischemic postconditioning contributes to
cardioprotective effects against ischemia/reperfusion injury in rat
hearts. J Cardiovasc Pharmacol. 61:416–422. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Ma X, Liu H, Foyil SR, Godar RJ,
Weinheimer CJ and Diwan A: Autophagy is impaired in cardiac
ischemia-reperfusion injury. Autophagy. 8:1394–1396. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Huang Z, Han Z, Ye B, Dai Z, Shan P, Lu Z,
Dai K, Wang C and Huang W: Berberine alleviates cardiac
ischemia/reperfusion injury by inhibiting excessive autophagy in
cardiomyocytes. Eur J Pharmacol. 762:1–10. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Kanamori H, Takemura G, Goto K, Maruyama
R, Ono K, Nagao K, Tsujimoto A, Ogino A, Takeyama T, Kawaguchi T,
et al: Autophagy limits acute myocardial infarction induced by
permanent coronary artery occlusion. Am J Physiol Heart Circ
Physiol. 300:H2261–H2271. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Kanamori H, Takemura G, Goto K, Maruyama
R, Tsujimoto A, Ogino A, Takeyama T, Kawaguchi T, Watanabe T,
Fujiwara T, et al: The role of autophagy emerging in postinfarction
cardiac remodelling. Cardiovasc Res. 91:330–339. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Liang B, Peng L, Li R, Li H, Mo Z, Dai X,
Jiang N, Liu Q, Zhang E, Deng H, et al: Lycium barbarum
polysaccharide protects HSF cells against ultraviolet-induced
damage through the activation of Nrf2. Cell Mol Biol Lett.
23:182018. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Zheng G, Ren H, Li H, Li X, Dong T, Xu S,
Yan Y, Sun B, Bai J and Li Y: Lycium barbarum polysaccharide
reduces hyperoxic acute lung injury in mice through Nrf2 pathway.
Biomed Pharmacother. 111:733–739. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Itoh K, Igarashi K, Hayashi N, Nishizawa M
and Yamamoto M: Cloning and characterization of a novel erythroid
cell-derived CNC family transcription factor heterodimerizing with
the small Maf family proteins. Mol Cell Biol. 15:4184–4193. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Magesh S, Chen Y and Hu L: Small molecule
modulators of Keap1-Nrf2-ARE pathway as potential preventive and
therapeutic agents. Med Res Rev. 32:687–726. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Shen Y, Liu X, Shi J and Wu X: Involvement
of Nrf2 in myocardial ischemia and reperfusion injury. Int J Biol
Macromol. 125:496–502. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Dang X, Zhang R, Peng Z, Qin Y, Sun J, Niu
Z and Pei H: HIPK2 overexpression relieves
hypoxia/reoxygenation-induced apoptosis and oxidative damage of
cardiomyocytes through enhancement of the Nrf2/ARE signaling
pathway. Chem Biol Interact. 316:1089222020. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Cheng L, Jin Z, Zhao R, Ren K, Deng C and
Yu S: Resveratrol attenuates inflammation and oxidative stress
induced by myocardial ischemia-reperfusion injury: Role of Nrf2/ARE
pathway. Int J Clin Exp Med. 8:10420–10428. 2015.PubMed/NCBI
|
|
59
|
Yu H, Shi L, Zhao S, Sun Y, Gao Y, Sun Y
and Qi G: Triptolide attenuates myocardial ischemia/reperfusion
injuries in rats by inducing the activation of Nrf2/HO-1 defense
pathway. Cardiovasc Toxicol. 16:325–335. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Zhang T, Yang Y, Liang Y, Jiao X and Zhao
C: Beneficial effect of intestinal fermentation of natural
polysaccharides. Nutrients. 10:10552018. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Tang C, Ding R, Sun J, Liu J, Kan J and
Jin C: The impacts of natural polysaccharides on intestinal
microbiota and immune responses - a review. Food Funct.
10:2290–2312. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Sun H, Ni X, Song X, Wen B, Zhou Y, Zou F,
Yang M, Peng Z, Zhu H, Zeng Y, et al: Fermented Yupingfeng
polysaccharides enhance immunity by improving the foregut
microflora and intestinal barrier in weaning rex rabbits. Appl
Microbiol Biotechnol. 100:8105–8120. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Zhang Z, Liu H, Yu B, Tao H, Li J, Wu Z,
Liu G, Yuan C, Guo L and Cui B: Lycium barbarum polysaccharide
attenuates myocardial injury in high-fat diet-fed mice through
manipulating the gut microbiome and fecal metabolome. Food research
international (Ottawa, Ont). 138:1097782020. View Article : Google Scholar : PubMed/NCBI
|