Introduction
Acute kidney injury (AKI) is known to be associated
with a decline in kidney function within 48 h (1). It is often associated with a high
mortality rate in patients, specifically in 1% of the affected
population (2).
Ischemia-reperfusion injury, sepsis and nephrotoxic insults are the
major risk factors for AKI (3–5).
Meanwhile, renal tubular epithelial cells are the main cell type in
kidney tissue; therefore, the dysfunction of these cells is the
main pathophysiological process of AKI (6). Studies have shown that the injury of
renal tubular epithelial cells plays an important role in the
pathogenesis of AKI (7,8). Moreover, the main features of AKI
include inflammatory cell infiltration and the massive secretion of
inflammatory factors, necrosis, apoptosis and delayed renal
resident cell proliferation (9,10).
Although significant efforts have been made in the treatment of
AKI, patient outcomes remain dismal. Thus, there is an urgent need
for the development of novel strategies for the treatment of
AKI.
MicroRNAs (miRNAs/miRs) are a novel class of
non-coding small ribonucleic acids, which can regulate gene
expression by suppression of mRNA translation or degradation of
mRNAs (11,12). Moreover, miRNAs participate in
cellular process, including the growth of renal tubular epithelial
cells (13,14). Meanwhile, miRNAs have been reported
to be associated with AKI progression (15–17).
For instance, miR-21 has been shown to inhibit the progression of
AKI (18); Jiang et al
(19) found that miR-500a-3p
alleviated kidney injury by targeting mixed lineage kinase domain
like pseudokinase. However, other miRNAs associated with the
progression of AKI warrant further investigation.
Exosomes are microvesicles ranging from 70 to 120 nm
in diameter and are derived from multivesicular bodies (20). In addition, exosomes participate in
cell communication by transferring proteins and nucleic acids, and
this function can lead to the mediation of intercellular
communication (21,22). In recent studies, a number of
exosomal proteins, miRNAs and lncRNAs have been reported to promote
the progression of AKI. For example, Cao et al (14) found that exosomal miR-125b-5p
deriving from mesenchymal stem cells (MSCs) could promote tubular
repair by suppression of p53 in ischemic AKI; Zhang et al
(23) indicated that endothelial
progenitor cells-derived exosomal miR-21-5p could alleviate
sepsis-induced AKI by inhibiting runt-related transcription factor
1 expression. However, the function of exosomes in AKI needs to be
further explored. On the other hand, among the potential targets of
miR-1184, forkhead box O4 (FOXO4) has been found to be associated
with cell growth (24). Thus, the
present study focused on the relationship between miR-1184 and the
FOXO signaling pathway.
On the other hand, MSCs, characterized by the
abilities of self-renewal, differentiation, immunomodulation and
trophic support, are essential in regenerative medicine owing to
the capacity to create a microenvironment conducive to the repair
of injured tissues (14). Previous
studies indicated that exosomes derived from MSCs have been
proposed as an alternative to MSC-based therapy for several
diseases (14,25). In addition, MSC-derived exosomes are
known to be involved in AKI progression. For instance, Cao et
al (26) found that exosomes
derived from MSCs (MSC-exos) could significantly attenuate
cisplatin-induced murine AKI through inhibiting inflammation; Ji
et al (25) demonstrated
that platelet-rich plasma could promote MSC-derived exosome
paracrine signaling to repair AKI via the AKT/Rab27 pathway. Thus,
exosomes derived from MSCs can play a vital role in AKI
progression.
Based on this background, the present study aimed to
detect the differentially expressed miRNAs closely associated with
the progression of AKI. The findings presented herein may provide
new insight on the role of miRNAs in AKI and may aid in the
development of novel treatment methods for AKI.
Materials and methods
Cell culture and treatment
HK-2 cell lines (American Type Culture Collection)
and MSCs (American Type Culture Collection) were maintained in
RPMI-1640 medium (Thermo Fisher Scientific, Inc.), supplemented
with 10% FBS (Gibco; Thermo Fisher Scientific, Inc.), 1% penicillin
(Thermo Fisher Scientific, Inc.) and 1% streptomycin (Thermo Fisher
Scientific, Inc.) in a condition with 5% CO2 and 37°C.
To mimic AKI in vitro, HK-2 cells were treated with 20 µM
cisplatin (MedChemExpress) for 48 h according to previous refs.
(8,27).
Cell transfection
HK-2 cells or MSCs (5×103 cells/well)
were transfected with miR-1184 agomir or agomir-negative control
(NC) using Lipofectamine® 2000 (Thermo Fisher
Scientific, Inc.) at 37°C for 48 h. miR-1184 agomir (50 nM) and
agomir-NC (50 nM) were obtained from Shanghai GenePharma Co., Ltd.
The sequences were as follows: miR-1184 agomir,
5′-CCUUCGGUAGUUCAGCGACGUCC-3′ and agomir-NC,
5′-UUCUCCGAACGUGUCACGUUU-3′. After 48 h of transfection, cells were
used in subsequent experiments.
For FOXO4 overexpression, MSCs (5×103
cells/well) were transfected with pcDNA3.1 (1 µg/µl; Shanghai
GenePharma Co., Ltd.) or pcDNA3.1-FOXO4 (1 µg/µl; Shanghai
GenePharma Co., Ltd.) using Lipofectamine 2000 for 48 h at 37°C,
according to the manufacturer's instructions. After 48 h of
transfection, cells were used in subsequent experiments.
Bioinformatics analysis
The differentially expressed miRNAs were presented
in a volcano plot and a heatmap using the GSE53771 dataset
(28) from the Gene Expression
Omnibus (GEO) database (https://www.ncbi.nlm.nih.gov/geo). In addition, the
differentially expressed miRNAs were screened using R analysis
(29). Using a t-test, P<0.05
and fold-change >1.5 or <0.667 were considered as
differentially expressed miRNAs. On the other hand, the functions
of miRNA-targeted mRNAs in terms of ‘Cellular Components’ and
‘Biological Processes’ were investigated by Gene Ontology (GO)
analysis (http://www.geneontology.org). Pathway analysis was
performed to define biological pathways by using the KEGG
orthology-based annotation system (KOBAS; version 3.0; http://kobas.cbi.pku.edu.cn/index.php).
Prediction of miRNA downstream
target
The downstream target of miR-1184 was predicted
using TargetScan (version 7.2; http://www.targetscan.org/vert_72/) and miRDB (version
2.0; http://www.mirdb.org/).
Exosome extraction and
identification
Briefly, MSCs were maintained in RPMI-1640 medium
until they reached 80% confluence. The medium was then replaced
with the serum-free medium. The supernatants were centrifuged for 1
h (300 × g for 15 min at 4°C, 2,000 × g for 15 min at 4°C and
10,000 × g for 30 min at °C) following 48 h of culture.
Subsequently, the supernatants were filtrated and collected to
extract exosomes using ultracentrifugation (2,000 × g for 15 min at
4°C). Western blot analysis was used to detect the exosome
isolation, and the detailed protocol was in accordance with a
previous study (25). The particle
sizes were detected by Nanoparticle tracking analysis (NTA).
NTA
A total of ~0.3 ml supernatant was loaded into the
sample chamber of an LM10 NanoSight unit (NanoSight, Ltd.) and
three videos of either 30 or 60 sec were recorded of each sample.
Data analysis was performed using NTA 2.1 software (NanoSight,
Ltd.). In NTA, the paths of unlabeled particles acting as point
scatterers, undergoing Brownian motion in a 0.25 ml chamber through
which a 635-nm laser beam was passed, was determined from a video
recording, with the mean squared displacement determined for each
possible particle. The diffusion coefficient and sphere-equivalent
hydrodynamic radius were subsequently determined using the
Stokes-Einstein equation (25).
Fluorescence staining
MSCs (5×104 per well) were seeded
overnight. Subsequently, cells were labeled for 24 h at 4°C with
phalloidin (1:1,000; Abcam; cat. no. ab176753) or PKH26 red
membrane dye (1:1,000; Biolab Co., Ltd.; cat. no. HR9070). The
nuclei were stained with 5 µl/ml DAPI (Beyotime Institute of
Biotechnology). The results were observed under a fluorescence
microscope (magnification, ×200; Olympus Corporation).
Western blot analysis
RIPA lysis buffer (Beyotime Institute of
Biotechnology) was used to extract total protein from the HK-2
cells. A BCA protein kit (Thermo Fisher Scientific, Inc.) was used
to quantify the total protein. SDS-PAGE (10%) was used to separate
the protein (40 µg per lane), and the protein was then transferred
to PVDF membranes (Thermo Fisher Scientific, Inc.). After blocking
with 5% skimmed milk at room temperature for 1 h, the membranes
were incubated overnight at 4°C with anti-CD63 (cat. no. ab134045;
1:1,000), anti-CD81 (cat. no. ab109201; 1:1,000), anti-Bax (cat.
no. ab32503; 1:1,000), anti-Bcl-2 (cat. no. ab32124; 1:1,000),
anti-cleaved caspase-3 (cat. no. ab32042; 1:1,000), anti-TSG101
(cat. no. ab125011; 1:1,000), anti-FOXO4 (cat. no. ab128908;
1:1,000), anti-p27 Kip1 (cat. no. ab32034; 1:1,000), anti-CDK2
(cat. no. ab32147; 1:1,000) and anti-β-actin (cat. no. ab8227;
1:1,000) primary antibodies (all Abcam). Subsequently, the
membranes were incubated with secondary anti-rabbit antibodies
(HRP-conjugated; Abcam; cat. no. ab7090; 1:5,000) for 1 h at room
temperature. Protein bands were visualized using an ECL kit (Thermo
Fisher Scientific, Inc.). β-actin was used as the loading control.
All antibodies were purchased from Abcam. ImageJ software (version
6.0; National Institutes of Health) was used for densitometry.
Reverse transcription-quantitative PCR
(RT-qPCR)
TRIzol® reagent (Invitrogen; Thermo
Fisher Scientific, Inc.) was used to isolate total RNA from HK-2
cells or MSCs. The PrimeScript RT Reagent Kit (Takara Bio, Inc.)
was used to reverse transcribe total RNA into cDNA, according to
the manufacturer's protocol. Subsequently, qPCR was performed using
the SYBR Premix Ex Taq II kit (ELK Biotechnology). RT-qPCR
reactions were performed under the following protocol: Initial
denaturation for 2 min at 94°C, followed by 35 cycles (30 sec at
94°C and 45 sec at 55°C). The following primer pairs were used for
RT-qPCR: miR-1184 forward, 5′-CTGGACTGAGCCGTGCTAC-3′ and reverse,
5′-CTCAACTGGTGTCGTGGAGTC-3′; and U6 forward,
5′-CTCGCTTCGGCAGCACAT-3′ and reverse, 5′-AACGCTTCACGAATTTGCGT-3′.
The 2−ΔΔCq method (30)
was used to quantify the data. U6 was used as an internal
control.
Cell counting kit-8 (CCK-8) assay
HK-2 cells (5×103 cells/well) were seeded
and treated with cisplatin (20 µM), MSC-ExomiR-1184
agomir or cisplatin + MSC-ExomiR-1184 agomir for
72 h at 37°C. Subsequently, CCK-8 reagent (10 µl; Beyotime
Institute of Biotechnology) was added to the cells for 2 h at 37°C.
The absorbance (450 nm) was measured using a microplate reader
(Thermo Fisher Scientific, Inc.).
Cell apoptosis analysis
HK-2 cells were trypsinized, washed with PBS,
resuspended in Annexin V Binding Buffer, and stained with 5 µl FITC
and 5 µl PI for 15 min in the dark. The cells were the analyzed
using a flow cytometer (FACSLyric™; BD Biosciences) to assess the
incidence of cell apoptosis (early + late apoptosis). FlowJo
(version 10.6.2; FlowJo LLC) was used to analyze the data.
Dual-luciferase reporter assay
The FOXO4 3′-untranslated region (UTR) containing
putative miR-1184 binding sites were obtained from Sangon Biotech
Co., Ltd., and cloned into pmirGLO Dual-Luciferase miRNA Target
Expression Vectors (Promega Corporation) to construct FOXO4
wild-type (WT) or mutant (MUT) reporter vectors. The mutated 3′-UTR
was generated using a site directed mutagenesis kit (Sangon Biotech
Co., Ltd.). FOXO4 (WT) or FOXO4 (MUT) were transfected into HK-2
cells (5×103) together with NC or miR-1184 agomir using
Lipofectamine 2000. After 48 h of transfection, relative luciferase
activities were then analyzed using a Dual-Glo Luciferase Assay
System (Promega Corporation). Renilla luciferase activity
was used for normalization.
ELISA
HK-2 cell supernatants were collected by
centrifugation (500 × g, 10 min, 4°C). Subsequently, the levels of
TNF-α (cat. no. H052-1) and IL-1β (cat. no. H002) were investigated
using ELISA kits (Nanjing Jiancheng Bioengineering Institute),
according to the manufacturers' protocols.
Cell cycle analysis
In brief, HK-2 cells were harvested, fixed with 75%
ethanol on ice for 20 min, permeabilized with 0.25% Triton X-100
and stained with PI/RNase (BD Pharmingen; BD Biosciences).
Following incubation at 4°C for 15 min, cells were analyzed using a
flow cytometer (BD FACSAria III; BD Biosciences). The data were
quantified using FlowJo software (version 3.0; FlowJo LLC).
Statistical analysis
Each group was examined in three independent
experiments and all data are expressed as the mean ± SD. Western
blot analysis, RT-qPCR, flow cytometry, CCK-8 assay and
immunofluorescence staining were repeated three times. An unpaired
Student's t-test was used to analyze the differences between two
groups, and one-way ANOVA followed by Tukey's test were used to
analyze differences among multiple groups (>2 groups, using
GraphPad Prism 7; GraphPad Software, Inc.). P<0.05 was
considered to indicate a statistically significant difference.
Results
Differentially expressed miRNAs in
AKI
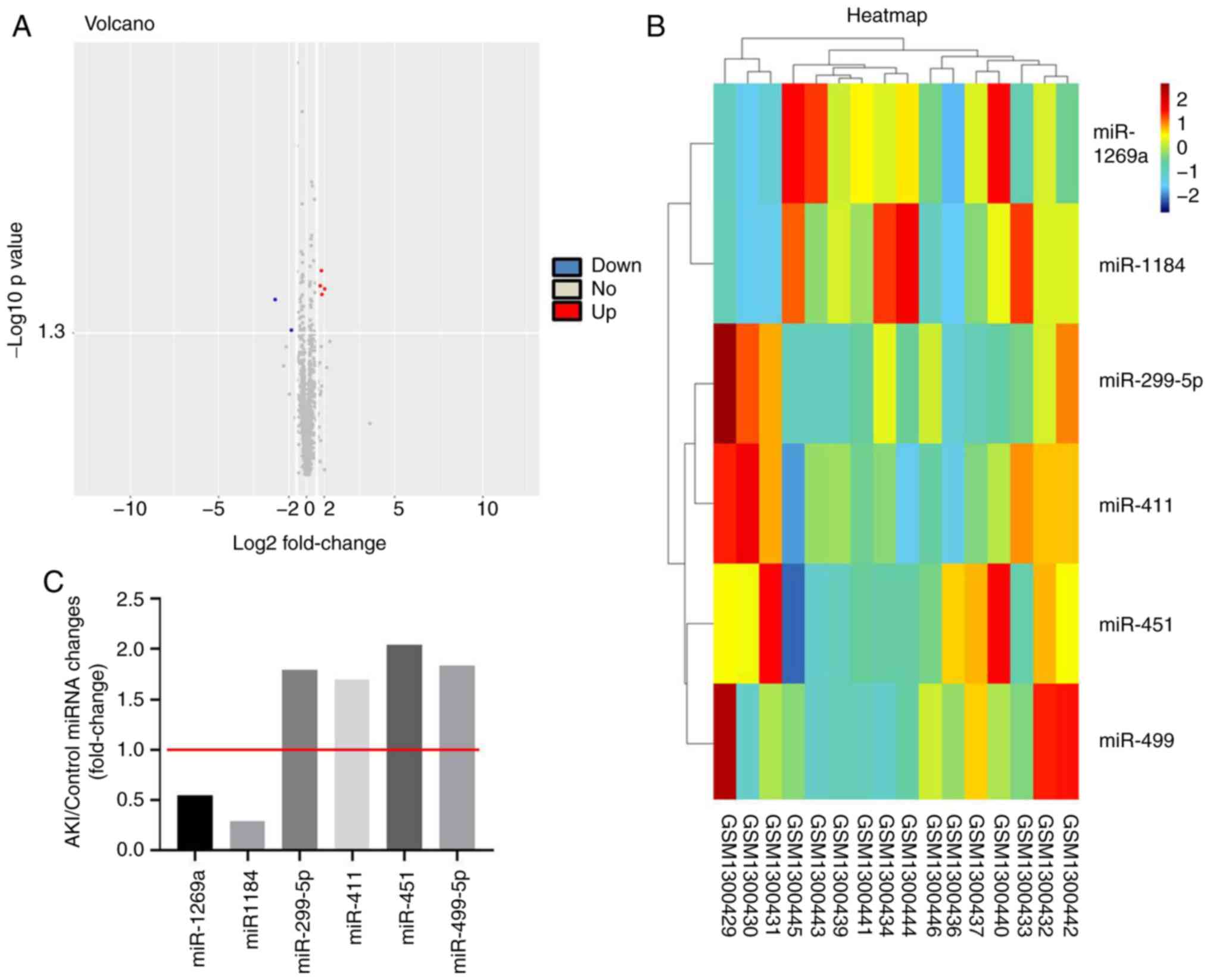
To detect differentially expressed miRNAs in AKI,
bioinformatics analysis was performed. As indicated in Fig. 1A and B, differentially expressed
miRNAs (miR-1269a, miR-1184, miR-299-5p, miR-411, miR-451 and
miR-499) are presented using a volcano plot and heatmap. In
addition, the six differentially expressed miRNAs (miR-1269a,
miR-1184, miR-299-5p, miR-411, miR-451 and miR-499; miR-1269a and
miR-1184 were downregulated, while the other four miRNAs were
upregulated) in AKI are presented (Fig.
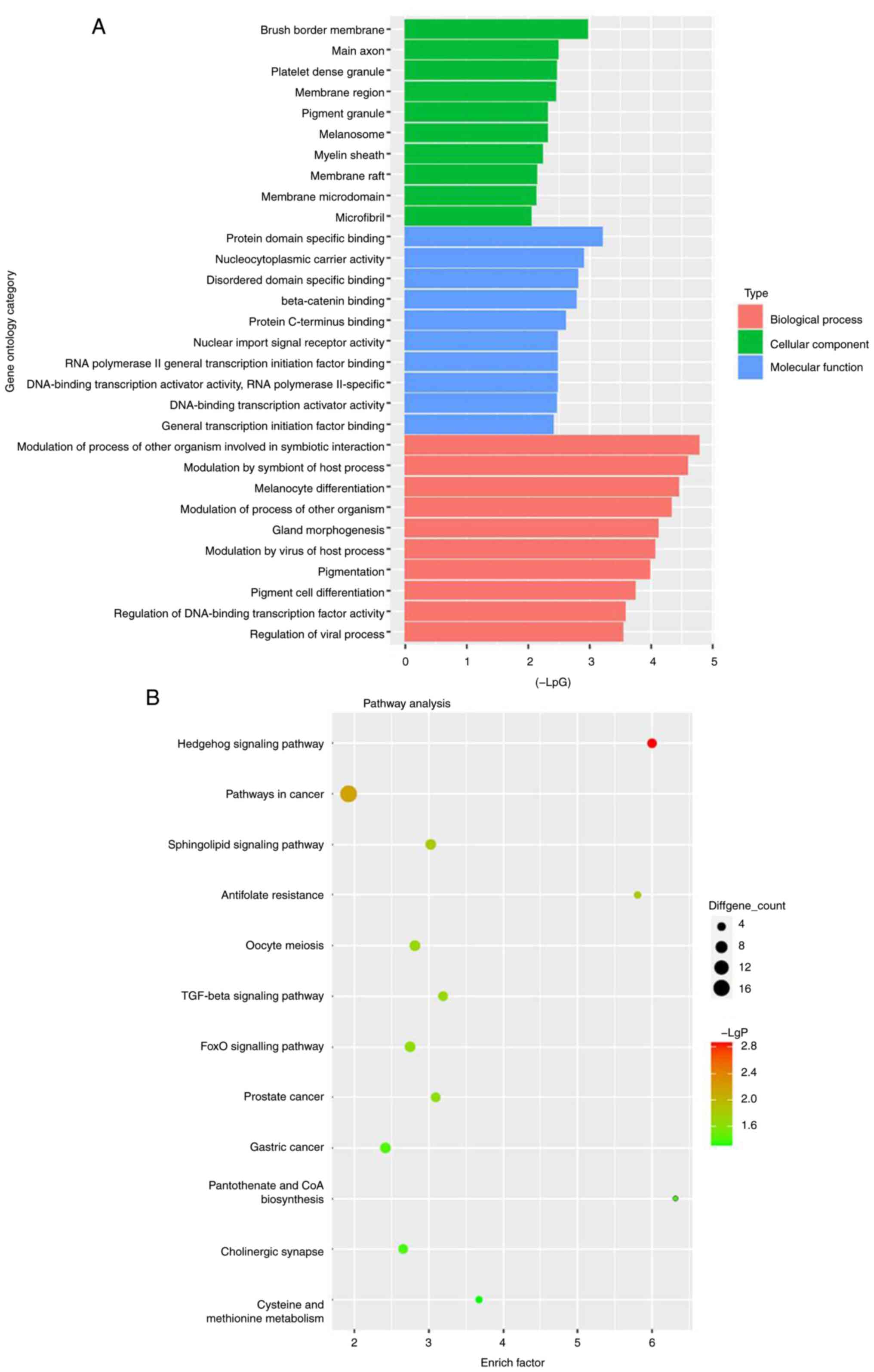
1C). Moreover, GO and pathway analyses were performed to
determine the most common ‘Biological Process’ of miR-1184. The
results indicated that miR-1184 was enriched in the following
‘Cellular Component’ terms: ‘Brush border membrane’, ‘main axon’,
‘platelet dense granule’, ‘membrane region’, ‘pigment granule’,
‘melanosome’, ‘myelin sheath’, ‘membrane raft’, ‘membrane
microdomain’ and ‘microfibril’ (Fig.
2A). miR-1184 was enriched in the following ‘Molecular
Function’ terms: ‘Protein domain specific binding’,
‘nucleocytoplasmic carrier activity’, ‘disordered domain specific
binding’, ‘β-catenin binding’, ‘protein C-terminus binding’,
‘nuclear import signal receptor activity’, ‘RNA polymerase II
general transcription initiation factor binding’, ‘DNA-binding
transcription activator activity’ and ‘RNA polymerase II-specific’
(Fig. 2A). miR-1184 was enriched in
the following ‘Biological Process’ terms: ‘Modulation of process of
other organism involved in symbiotic interaction’, ‘modulation by
symbiont of host process’, ‘melanocyte differentiation’,
‘modulation of process of other organism’, ‘gland morphogenesis’,
‘modulation by virus of host process’, ‘pigmentation’, ‘pigment
cell differentiation’, ‘regulation of DNA-binding transcription
factor activity’ and ‘regulation of viral process’ (Fig. 2A). Notably, the most enriched
‘Cellular Component’ was ‘brush border membrane’, the most enriched
‘Molecular Function’ was ‘protein domain specific binding’ and the
most enriched ‘Biological Process’ was ‘modulation of process of
other organism involved in symbiotic interaction’ (Fig. 2A). Pathway analysis revealed that
the ‘Hedgehog signaling pathway’, ‘pathways in cancer’,
‘sphingolipid signaling pathway’, ‘antifolate resistance’, ‘oocyte
meiosis’, ‘TGF-beta signaling pathway’, ‘FOXO signaling pathway’,
‘prostate cancer’, ‘gastric cancer’, ‘pantothenate and CoA
biosynthesis’, ‘cholinergic synapse’ and ‘cysteine and methionine
metabolism’ were associated with the development of AKI (Fig. 2B). Based on the aforementioned data,
miR-1184 was selected for further analysis.
Successful isolation of exosomes from
MSCs
In order to detect the efficiency of cell
transfection and exosome isolation, RT-qPCR and NTA was performed.
As shown in Fig. 3A, the expression
of miR-1184 in MSCs was significantly upregulated by miR-1184
agomir, and NTA revealed a similar size distribution of exosomes
(Fig. 3B). Moreover, exosomal
proteins (TSG101, CD63 and CD81) were highly expressed in exosomes
derived from MSCs, while they were expressed at low levels in MSCs
(Fig. 3C). Moreover, the expression
of miR-1184 in exosomes derived from MSCs was significantly
upregulated by miR-1184 agomir (Fig.
3D), and MSC-derived exosomes labeled with fluorescent PKH26
were internalized by unstained MSCs (Fig. 3E). Furthermore, the expression level
of miR-1184 in MSCs was significantly increased by exosomes derived
from miR-1184 agomir-treated MSCs (Fig.
3F). Taken together, the data demonstrated that exosomes were
successfully isolated from MSCs.
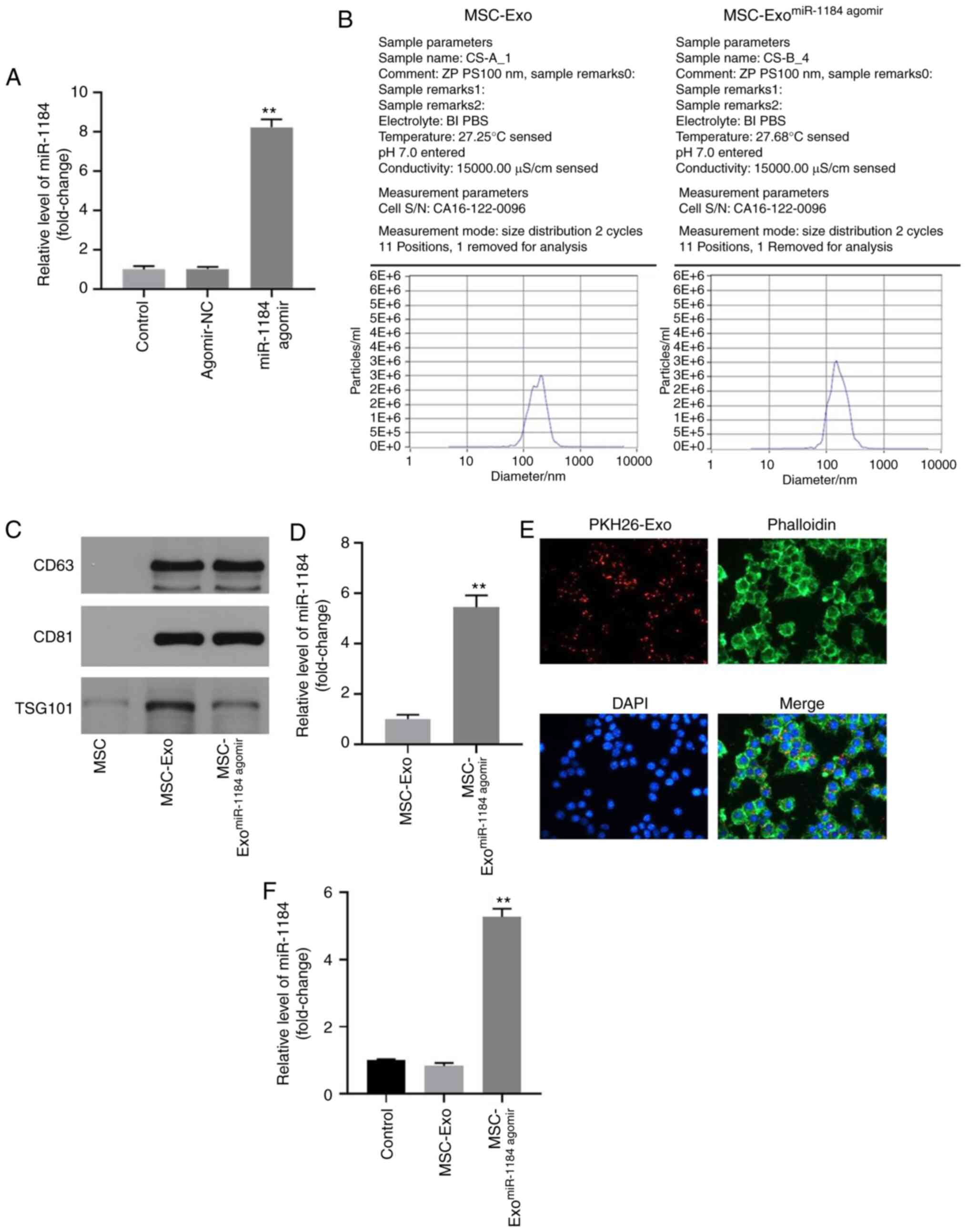 | Figure 3.Exosomes were successfully isolated
from MSCs. (A) MSCs were transfected with miR-1184 agomir or
agomir-NC. Then, the expression of miR-1184 in MSCs was detected by
RT-qPCR. **P<0.01 vs. control. (B) The particle sizes of
exosomes were measured by Nanoparticle tracking analysis. (C) The
expression levels of CD63, CD81 and TSG101 in MSCs, MSC-Exo or
MSC-ExomiR-1184 agomir were detected by western
blotting. (D) The expression of miR-1184 in MSC-Exo or
MSC-ExomiR-1184 agomir was detected by RT-qPCR.
**P<0.01 vs. MSC-Exo. (E) The location of exosomes was observed
by immunofluorescence staining. (F) MSCs were co-cultured with
MSC-Exo or MSC-ExomiR-1184 agomir. Then, the expression
of miR-1184 in MSCs was tested by RT-qPCR. **P<0.01 vs. control.
MSCs, mesenchymal stem cells; miR, microRNA; NC, negative control;
RT-qPCR, reverse transcription-quantitative PCR; MSC-Exo, exosomes
derived from MSCs; MSC-ExomiR-1184 agomir, exosomes
derived from miR-1184 agomir-treated MSCs. |
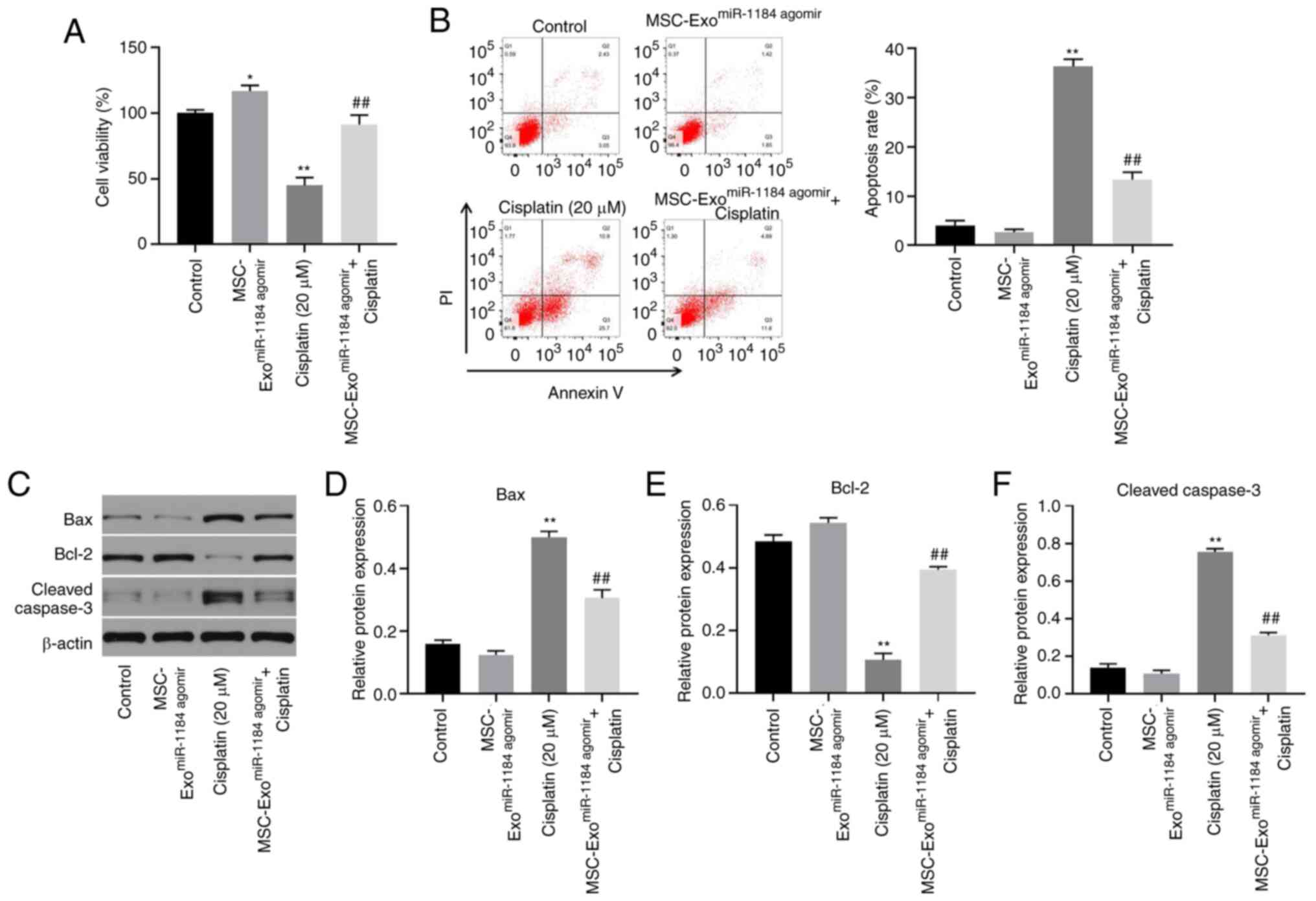
Exosomes derived from miR-1184
agomir-treated MSCs significantly reverse cisplatin-induced HK-2
cell growth inhibition
In order to detect the effects of exosomes on AKI
in vitro, a CCK-8 assay was used. As illustrated in Fig. 4A, the viability of the HK-2 cells
was significantly increased when the cells were co-cultured with
exosomes derived from miR-1184 agomir-treated MSCs, and exosomes
expressing miR-1184 significantly reversed the cisplatin-induced
inhibition of cell viability. In addition, cisplatin significantly
induced HK-2 cell apoptosis, while this phenomenon was reversed in
the presence of exosomes derived from miR-1184 agomir-treated MSCs
(Fig. 4B). Moreover, cisplatin
significantly upregulated the expression levels of Bax and cleaved
caspase-3 and downregulated the protein expression level of Bcl-2,
while this phenomenon was reversed by exosomes derived from
miR-1184 agomir-treated MSCs (Fig.
4C-F). Taken together, the results demonstrated that exosomes
derived from miR-1184 agomir-treated MSCs significantly reversed
cisplatin-induced HK-2 cell growth inhibition.
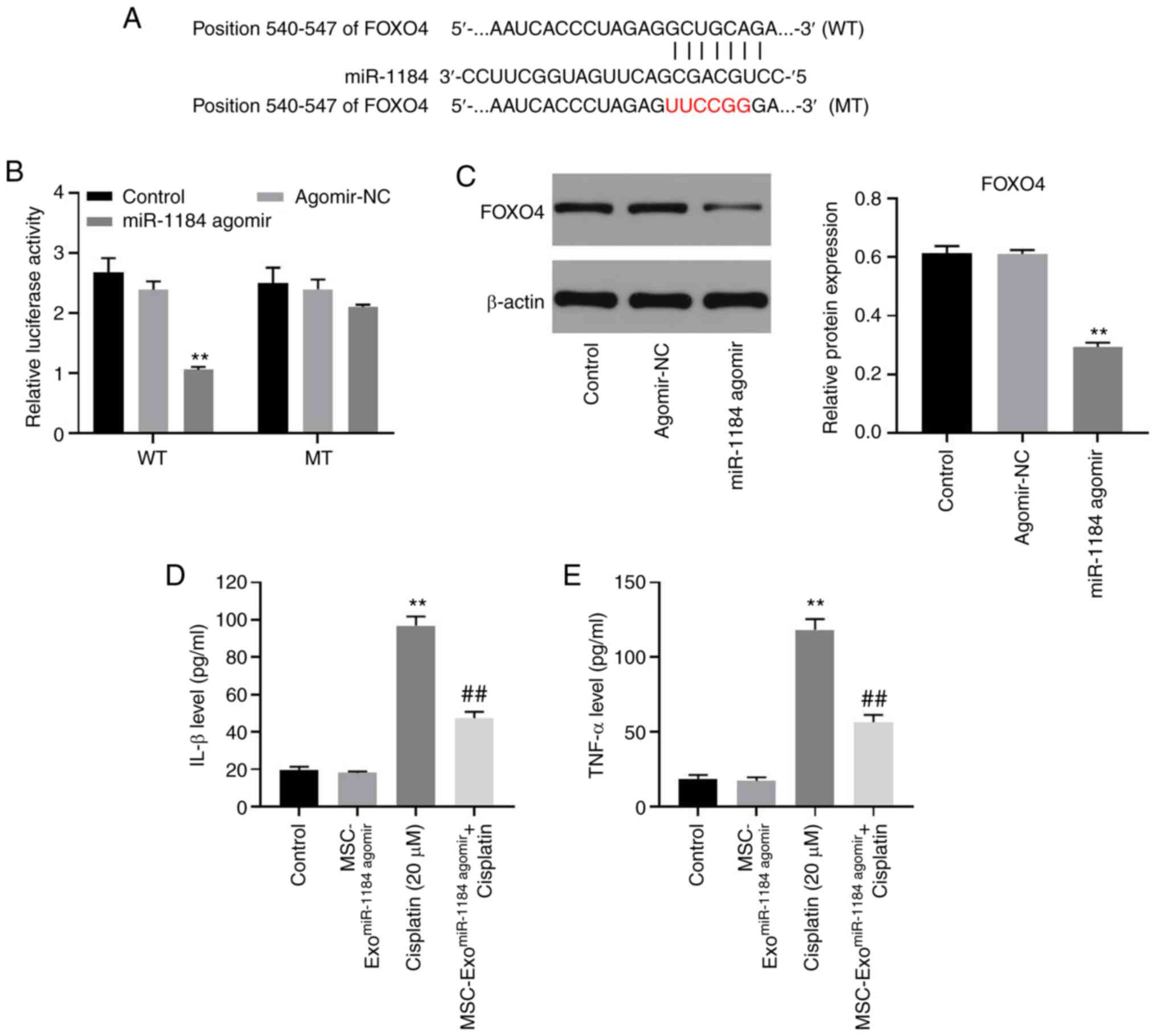
miR-1184 directly targets FOXO4 in
HK-2 cells
To explore the potential target of miR-1184,
TargetScan and miRDB databases were used. As shown in Fig. 5A, FOXO4 may be the target of
miR-1184, and the relative luciferase activity in the WT-FOXO4
group was significantly decreased by miR-1184 agomir (Fig. 5B). Moreover, the expression level of
FOXO4 in HK-2 cells was significantly inhibited by the
overexpression of miR-1184 (Fig.
5C). Furthermore, the levels of IL-1β and TNF-α in the
supernatants of HK-2 cells were significantly upregulated by
cisplatin, while this phenomenon was partially reversed by
MSC-ExomiR-1184 agomir (Fig.
5D and E). Therefore, miR-1184 directly targeted FOXO4 in HK-2
cells.
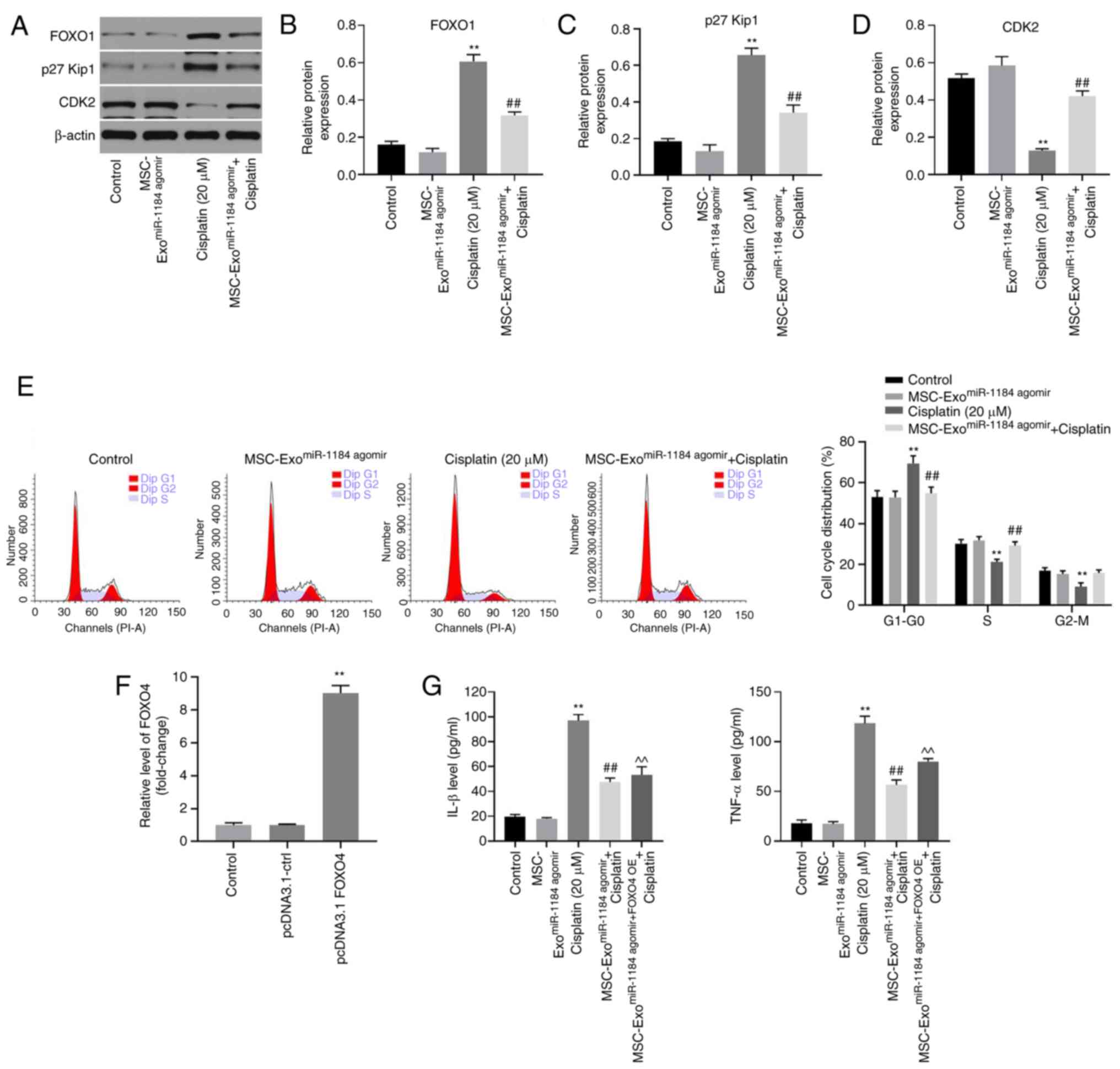
Exosomes derived from miR-1184
agomir-treated MSCs significantly induce G1 arrest in
HK-2 cells via the mediation of FOXO1, p27 Kip1 and CDK2
In order to further explore the underlying
mechanisms through which exosomes or miR-1184 agomir regulate
cisplatin-induced HK-2 cell injury, western blot analysis was
performed. As indicated in Figs.
6A-C and S1A-C the protein
expression levels of FOXO1 and p27 Kip1 in HK-2 cells were
significantly upregulated by cisplatin. By contrast, cisplatin
significantly inhibited the expression level of CDK2 (Figs. 6D and S1D). Furthermore, the effect of cisplatin
on these three proteins was reversed by MSC-ExomiR-1184
agomir (Figs. 6A-D and
S1A-D). In addition,
cisplatin-induced G1 arrest was reversed by
MSC-ExomiR-1184 agomir (Fig.
6E). The expression of FOXO4 in MSCs was significantly
upregulated by transfection with the FOXO4-overexpression vector
(Fig. 6F). Meanwhile, the
anti-inflammatory effect of exosomes derived from miR-1184
agomir-treated MSCs on cisplatin-treated HK-2 cells was rescued in
the presence of exosomes derived from MSCs co-treated with miR-1184
agomir and FOXO4 overexpression (Fig.
6G). Collectively, the results demonstrated that exosomes
derived from miR-1184 agomir-treated MSCs significantly induced
G1 arrest in HK-2 cells via the mediation of FOXO1, p27
Kip1 and CDK2.
Discussion
It has been reported that miRNAs are involved in the
progression of AKI (19,31,32).
In the present study, it was found that miR-1184 was downregulated
in cisplatin-treated HK-2 cells, and that exosomes derived from
miR-1184 agomir-treated HK-2 cells significantly reversed
cisplatin-induced HK-2 cell injury. Thus, the present study firstly
explored the function of miR-1184 in AKI, suggesting that miR-1184
may function as an inhibitor in AKI. Moreover, miR-1184 is known to
be involved in other diseases. For example, Wang et al
(33) indicated that
hsa_circ_0128846 promoted the tumorigenesis of colorectal cancer by
binding to miR-1184 and releasing ajuba LIM protein, and
inactivating Hippo/Yes1 associated transcriptional regulator
signaling. Chen et al (34)
found that miR-1184 mediated the proliferation of colon cancer
cells by targeting casein kinase 2 α 1. Thus, other functions of
miR-1184 (for example its role in renal cancer) also need to be
explored in the future.
It has been reported that exosomes derived from MSCs
play important roles in the progression of AKI. For example,
Alzahrani (35) found that exosomes
derived from MSCs could promote the progression of AKI; Cao et
al (14) indicated that
exosomal miR-125b-5p derived from MSCs could promote tubular repair
by suppression of p53 in ischemic AKI. In the present study, it was
found that exosomal miR-1184 derived from MSCs could inhibit
cisplatin-induced HK-2 cell injury. Thus, this research first
explored the relationship between miR-1184 and exosomes derived
from MSCs.
It has been confirmed that miRNAs exert their
biological functions due to their target genes (36). In the present study, the results of
the dual-luciferase reporter assay indicated that FOXO4 was a
downstream target of miR-1184 in AKI. FOXO4 is a member of the FOXO
family, and is a crucial mediator of cell proliferation (37). Consistently, the data of the present
study revealed that miR-1184 may function as a mediator in AKI by
targeting FOXO4. On the other hand, it has been reported that p27
Kip1 is a cell cycle regulator, firstly regarded as a
cyclin-dependent kinase antagonist (38). It has been reported that p27 Kip1
expression is upregulated during the progression of AKI (39,40).
Taken together with the findings of the present study, it can be
concluded that p27 Kip1 is a negative mediator in AKI.
Additionally, CDK2 is a mediator of cell cycle distribution and is
a downstream protein of p27 Kip1 (41). In the present study, exosomes
derived from miR-1184 agomir-treated HK-2 cells reversed
cisplatin-induced cell growth inhibition via the mediation of
FOXO1, p27 Kip1 and CDK2. Thus, these findings are consistent with
those of previous studies. Taken together, the findings presented
herein revealed that miR-1184 exerted an inhibitory effect on AKI
by targeting FOXO1.
There are some limitations of the present study,
which are as follows: i) Other mRNAs targeted by miR-1184 in AKI
need to be explored in the future; ii) rescue experiments should be
performed in order to further verify the function of exosomes in
AKI; iii) electron microscopy analysis is needed to further
identify the exosomes; iv) the function of miR-1184 antagomir
should be further confirmed; and v) the function of miR-184 in
renal cancer needs to be explored. Therefore, further
investigations are required in the future.
In conclusion, the present study demonstrated that
exosomal-miR-1184 derived from MSCs alleviated cisplatin-associated
AKI. Thus, these findings may lead to the development of novel
strategies for the treatment of AKI.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
This research was supported by the Construction of
Key Projects by Zhejiang Provincial Ministry (grant nos.
WKJ-ZJ-1915 and WKJ-ZJ-2017), the Zhejiang Province Chinese
Medicine Modernization Program (grant no. 2020ZX001) and the
General Project of Zhejiang Education Department (grant no.
Y201942823).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
MT and JJ conceived and supervised the present
study. JZ and WH designed the experiments and reviewed the results.
DZ and QH performed the experiments. All authors have read and
approved the final manuscript. MT and JJ confirm the authenticity
of all the raw data.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Stephen Inbaraj B and Chen BH: An overview
on recent in vivo biological application of cerium oxide
nanoparticles. Asian J Pharm Sci. 15:558–575. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zhang J, Healy HG, Baboolal K, Wang Z,
Venuthurupalli SK, Tan KS, Cameron A and Hoy WE; CKD.QLD
Collaborative, : Frequency and consequences of acute kidney injury
in patients with CKD: A registry study in queensland australia.
Kidney Med. 1:180–190. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Guillemet L, Jamme M, Bougouin W, Geri G,
Deye N, Vivien B, Varenne O, Pène F, Mira JP, Barat F, et al:
Effects of early high-dose erythropoietin on acute kidney injury
following cardiac arrest: Exploratory post hoc analyses from an
open-label randomized trial. Clin Kidney J. 13:413–420.
2020.PubMed/NCBI
|
|
4
|
Güzel C, Yeşiltaş S, Daşkaya H, Uysal H,
Sümer I and Türkay M: The effect of gender on acute kidney injury
developing in the intensive care unit. Hippokratia. 23:126–130.
2019.PubMed/NCBI
|
|
5
|
Xiong J, Zhang M, Guo X, Pu L, Xiong H,
Xiang P, Liu J and Li A: Acute kidney injury in critically ill
cirrhotic patients with spontaneous bacterial peritonitis: A
comparison of KDIGO and ICA criteria. Arch Med Sci. 16:569–576.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Deng J, Tan W, Luo Q, Lin L, Zheng L and
Yang J: Long non-coding RNA MEG3 promotes renal tubular epithelial
cell pyroptosis by regulating the miR-18a-3p/GSDMD pathway in
lipopolysaccharide-induced acute kidney injury. Front Physiol.
12:6632162021. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Han D, Fang R, Shi R, Jin Y and Wang Q:
LncRNA NKILA knockdown promotes cell viability and represses cell
apoptosis, autophagy and inflammation in lipopolysaccharide-induced
sepsis model by regulating miR-140-5p/CLDN2 axis. Biochem Biophys
Res Commun. 559:8–14. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yang J, Wu L, Liu S, Hu X, Wang Q and Fang
L: Long non-coding RNA NEAT1 promotes lipopolysaccharide-induced
injury in human tubule epithelial cells by regulating
miR-93-5p/TXNIP axis. Med Microbiol Immunol. 210:121–132. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Xu Y, Qin S, Niu Y, Gong T, Zhang Z and Fu
Y: Effect of fluid shear stress on the internalization of
kidney-targeted delivery systems in renal tubular epithelial cells.
Acta Pharm Sin B. 10:680–692. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Guthrie GD and Bell S: Deprivation and
kidney disease-a predictor of poor outcomes. Clin Kidney J.
13:128–132. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Jia X, Wei L and Zhang Z: NEAT1
Overexpression indicates a poor prognosis and induces chemotherapy
resistance via the miR-491-5p/SOX3 signaling pathway in ovarian
cancer. Front Genet. 12:6162202021. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhang F, Sang Y, Chen D, Wu X, Wang X,
Yang W and Chen Y: M2 macrophage-derived exosomal long non-coding
RNA AGAP2-AS1 enhances radiotherapy immunity in lung cancer by
reducing microRNA-296 and elevating NOTCH2. Cell Death Dis.
12:4672021. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang Q, Tao Y, Xie H, Liu C and Liu P:
MicroRNA-101 inhibits renal tubular epithelial-to-mesenchymal
transition by targeting TGF-β1 type I receptor. Int J Mol Med.
47:1192021. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Cao JY, Wang B, Tang TT, Wen Y, Li ZL,
Feng ST, Wu M, Liu D, Yin D, Ma KL, et al: Exosomal miR-125b-5p
deriving from mesenchymal stem cells promotes tubular repair by
suppression of p53 in ischemic acute kidney injury. Theranostics.
11:5248–5266. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Scullion KM, Vliegenthart ADB, Rivoli L,
Oosthuyzen W, Farrah TE, Czopek A, Webb DJ, Hunter RW, Bailey MA,
Dhaun N and Dear JW: Circulating argonaute-bound microRNA-126
reports vascular dysfunction and treatment response in acute and
chronic kidney disease. iScience. 24:1019372020. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sun B, Qu Z, Cheng GL, Yang YW, Miao YF,
Chen XG, Zhou XB and Li B: Urinary microRNAs miR-15b and miR-30a as
novel noninvasive biomarkers for gentamicin-induced acute kidney
injury. Toxicol Lett. 338:105–113. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Jeon BS, Lee SH, Hwang SR, Yi H, Bang JH,
Tham NTT, Lee HK, Woo GH, Kang HG and Ku HO: Identification of
urinary microRNA biomarkers for in vivo gentamicin-induced
nephrotoxicity models. J Vet Sci. 21:e812020. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Pan T, Jia P, Chen N, Fang Y, Liang Y, Guo
M and Ding X: Delayed remote ischemic preconditioning
confersrenoprotection against septic acute kidney injury via
exosomal miR-21. Theranostics. 9:405–423. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Jiang L, Liu XQ, Ma Q, Yang Q, Gao L, Li
HD, Wang JN, Wei B, Wen J, Li J, et al: hsa-miR-500a-3P alleviates
kidney injury by targeting MLKL-mediated necroptosis in renal
epithelial cells. FASEB J. 33:3523–3535. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang X, Wang N, Huang Y, Li Y, Li G, Lin
Y, Atala AJ, Hou J and Zhao W: Extracellular vesicles from three
dimensional culture of human placental mesenchymal stem cells
ameliorated renal ischemia/reperfusion injury. Int J Artif Organs.
Jan 19–2021.(Epub ahead of print). doi: 10.1177/0391398820986809.
View Article : Google Scholar
|
|
21
|
Jiang Z, Hou Z, Li L, Liu W, Yu Z and Chen
S: Exosomal circEPB41L2 serves as a sponge for miR-21-5p and
miR-942-5p to suppress colorectal cancer progression by regulating
the PTEN/AKT signalling pathway. Eur J Clin Invest. 51:e135812021.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Pineles B, Mani A, Sura L, Rossignol C,
Albayram M, Weiss MD and Goetzl L: Neuronal exosome proteins: Novel
biomarkers for predicting neonatal response to therapeutic
hypothermia. Arch Dis Child Fetal Neonatal Ed. May 21–2021.(Epub
ahead of print). doi: 10.1136/archdischild-2020-321096. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhang Y, Huang H, Liu W, Liu S, Wang XY,
Diao ZL, Zhang AH, Guo W, Han X, Dong X and Katilov O: Endothelial
progenitor cells-derived exosomal microRNA-21-5p alleviates
sepsis-induced acute kidney injury by inhibiting RUNX1 expression.
Cell Death Dis. 12:3352021. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Huang Y, He Y, Makarcyzk MJ and Lin H:
Senolytic peptide FOXO4-DRI selectively removes senescent cells
from in vitro expanded human chondrocytes. Front Bioeng Biotechnol.
9:6775762021. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ji C, Zhang J, Zhou Z, Shi H, Liu W, Sun
F, Zhang C, Zhang L, Sun Z and Qian H: Platelet-rich plasma
promotes MSCs exosomes paracrine to repair acute kidney injury via
AKT/Rab27 pathway. Am J Transl Res. 13:1445–1457. 2021.PubMed/NCBI
|
|
26
|
Cao J, Wang B, Tang T, Lv L, Ding Z, Li Z,
Hu R, Wei Q, Shen A, Fu Y and Liu B: Three-dimensional culture of
MSCs produces exosomes with improved yield and enhanced therapeutic
efficacy for cisplatin-induced acute kidney injury. Stem Cell Res
Ther. 11:2062020. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
McSweeney KR, Gadanec LK, Qaradakhi T, Ali
BA, Zulli A and Apostolopoulos V: Mechanisms of cisplatin-induced
acute kidney injury: Pathological mechanisms, pharmacological
interventions, and genetic mitigations. Cancers (Basel).
13:15722021. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Cheng Q and Wang L: LncRNA XIST serves as
a ceRNA to regulate the expression of ASF1A, BRWD1M, and PFKFB2 in
kidney transplant acute kidney injury via sponging hsa-miR-212-3p
and hsa-miR-122-5p. Cell Cycle. 19:290–299. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Varet H, Brillet-Guéguen L, Coppée JY and
Dillies MA: SARTools: A DESeq2- and EdgeR-based R pipeline for
comprehensive differential analysis of RNA-Seq data. PLoS One.
11:e01570222016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ren GL, Zhu J, Li J and Meng XM: Noncoding
RNAs in acute kidney injury. J Cell Physiol. 234:2266–2276. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhang W, Zhou X, Zhang H, Yao Q, Liu Y and
Dong Z: Extracellular vesicles in diagnosis and therapy of kidney
diseases. Am J Physiol Renal Physiol. 311:F844–F851. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wang X, Chen Y, Liu W, Liu T and Sun D:
Hsa_circ_0128846 promotes tumorigenesis of colorectal cancer by
sponging hsa-miR-1184 and releasing AJUBA and inactivating
Hippo/YAP signalling. J Cell Mol Med. 24:9908–9924. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chen S, Wang Y, Xu M, Zhang L, Su Y, Wang
B and Zhang X: miR-1184 regulates the proliferation and apoptosis
of colon cancer cells via targeting CSNK2A1. Mol Cell Probes.
53:1016252020. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Alzahrani FA: Melatonin improves
therapeutic potential of mesenchymal stem cells-derived exosomes
against renal ischemia-reperfusion injury in rats. Am J Transl Res.
11:2887–2907. 2019.PubMed/NCBI
|
|
36
|
Shin JH, Shin DH and Kim JS: Let-7 miRNA
and CDK4 siRNA co-encapsulated in Herceptin-conjugated liposome for
breast cancer stem cells. Asian J Pharm Sci. 15:472–481. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhang D, Liu K, Hu W, Lu X, Li L, Zhang Q,
Huang H and Wang H: Prenatal dexamethasone exposure caused fetal
rats liver dysplasia by inhibiting autophagy-mediated cell
proliferation. Toxicology. 449:1526642021. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Henriques J and Lindorff-Larsen K: Protein
dynamics enables phosphorylation of buried residues in
Cdk2/cyclin-A-bound p27. Biophys J. 119:2010–2018. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Park JW, Cho JW, Joo SY, Kim CS, Choi JS,
Bae EH, Ma SK, Kim SH, Lee J and Kim SW: Paricalcitol prevents
cisplatin-induced renal injury by suppressing apoptosis and
proliferation. Eur J Pharmacol. 683:301–309. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Coombes JD, Mreich E, Liddle C and Rangan
GK: Rapamycin worsens renal function and intratubular cast
formation in protein overload nephropathy. Kidney Int.
68:2599–2607. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Guerra B, Dembic M, Siddiqui MA, Dominguez
I, Ceppi P and Andresen BS: Down-regulation of CK2α leads to
up-regulation of the cyclin-dependent kinase inhibitor
p27KIP1 in conditions unfavorable for the growth of
myoblast cells. Cell Physiol Biochem. 54:1177–1198. 2020.
View Article : Google Scholar : PubMed/NCBI
|