Introduction
Helicobacter pylori is the major pathogen in
atrophic gastritis, peptic ulcers and gastric cancer (1). The underlying mechanism of H.
pylori-mediated gastric carcinogenesis is the production of
reactive oxygen species (ROS) by neutrophil activation or by H.
pylori itself in the infected gastric epithelial cells
(2). Our previous studies
indicated that H. pylori induced NADPH oxidase activation,
resulting in increased production of ROS in human gastric
epithelial cells (3,4).
ROS are composed of hydroxyl radicals, superoxide
anion and hydrogen peroxide (5).
The normal range of ROS levels can be strictly controlled by
antioxidant defense systems such as superoxide dismutase (SOD),
catalase and glutathione peroxidase in mammalian cells (6). However, excessive ROS generation
potentially causes oxidative stress, which leads to tumor
initiation, growth and progression, resulting in cancer development
(6–8). ROS activate Janus kinase (JAK)/STAT
signaling and phosphorylate STAT3 in stem cells and a human
salivary gland cell line (9,10).
Inflammatory stimuli such as cytokines potentially activate JAKs
via phosphorylation; thereafter, activated JAKs recruit STATs
(11). Activated/phosphorylated
(p-)STATs form dimers that translocate from the cytoplasm to the
nucleus, regulating the expression of pro-inflammatory genes such
as interleukin-6 by binding to specific promoter sequences
(12). Activated STAT3 mediates
the expression of cancer-inducing genes related to tumor cell
survival, migration and metastasis (13,14)
such as integrin β6 (15).
Integrins are heterodimeric, transmembrane proteins
that comprise α and β subunits (16). Mammals possess 18 α and eight β
integrin subunits that combine to form 24 types of αβ integrin
heterodimers (17). Different
integrin heterodimers bind to different extracellular ligands and
distinct integrin heterodimers are present in various tissues
(18). Among these integrins,
integrin α5β1 is a heterodimer with α5 and β1 subunits. Integrin
α5β1 is related to tumor metastasis and carcinogenesis (17). Integrin-mediated cell adhesion
serves an important role in cancer metastasis by inducing further
cell migration and invasion (19,20).
Integrin α5 promotes migration and invasion through STAT3 signaling
in non-small cell lung cancer cells (21).
Our previous study showed that an H. pylori
strain originally isolated from a Korean patient (HP99) increased
integrin α5/β1 expression, which was inhibited by treatment with
diphenyleneiodonium chloride, an inhibitor of NADPH oxidase in AGS
cells (22). The results clearly
demonstrated the involvement of NADPH oxidase in H.
pylori-induced integrin expression. In addition, HP99 induces
the expression of proteinase-activated receptor-2, which mediates
α5/β1 expression and cell adhesion to fibronectin in gastric
epithelial AGS cells (23). A
study using a cDNA microarray of 352 cancer-related genes
demonstrated that H. pylori (strain NCTC11637) increased
integrin α5 expression in AGS cells (24). Since H. pylori increases
ROS-mediated activation of the JAK1/STAT3 signaling pathway in AGS
cells (25), JAK1/STAT3 may be
involved in the upregulation of integrin expression in H.
pylori-stimulated AGS cells.
Astaxanthin (ASX) is a red-orange and lipid-soluble
oxycarotenoid pigment (26). It is
found in seafood such as shrimp, lobster, trout, krill and salmon
(26). ASX can scavenge ROS in
both the inner and outer membrane of the cells (27). Therefore, ASX may inhibit
development of oxidative stress-induced diseases, including
inflammatory diseases (28).
Furthermore, ASX has anticancer effects, inducing
anti-proliferative activity and suppressing cell migration and
invasion in colorectal cancer, breast cancer, melanoma, gastric
cancer and oral cancer cells (29–31).
By inhibiting matrix metalloproteinase-1, −2 and −9 expression, ASX
suppresses melanoma cancer cell migration and metastasis (30). ASX inhibits cancer development and
progression by inhibiting STAT3 signaling in oral squamous
carcinoma cells (31). Another
study demonstrated that, in H. pylori-stimulated cells, ASX
treatment activated peroxisome proliferator-activated receptor γ
(PPAR-γ) to induce the expression of its downstream antioxidant
gene catalase, and suppressed intracellular and mitochondrial ROS
production (32). Furthermore, ASX
prevents H. pylori-induced decreases in SOD2 levels and SOD
activity, and reduces mitochondrial ROS in gastric epithelial cells
(33). Therefore, ASX may suppress
H. pylori-induced cell adhesion and migration by reducing
ROS-mediated activation of STAT3 and expression of integrins in
gastric epithelial cells.
The present study aimed to determine whether ASX
suppresses integrin α5/β1 expression, cell adhesion and migration
by inhibiting the JAK1/STAT3 signaling pathway in H.
pylori-stimulated gastric epithelial cells. In addition,
integrin α5/β1 expression, cell adhesion and migration were
determined after treatment with a JAK/STAT inhibitor, AG490, and an
integrin α5β1 antagonist, K34C, in H. pylori-stimulated
cells.
Materials and methods
Cell line and culture conditions
The AGS human gastric epithelial cell line (CRL
1739) was obtained from American Type Culture Collection. Cells
were cultured in complete medium comprising RPMI 1640 medium
(Gibco; Thermo Fisher Scientific, Inc.) supplemented with 100 U/ml
penicillin, 2 mM glutamine, 10% FBS (Gibco; Thermo Fisher
Scientific, Inc.) and 100 µg/ml streptomycin. Subsequently, the
cells were cultured in a cell incubator at 37°C in a humidified
atmosphere with 95% air and 5% CO2.
Bacterial strain
H. pylori, strain NCTC11637
[cytotoxin-associated gene (cag) A+, vacuolating
cytotoxin gene (vac) A+ strain], was obtained from
American Type Culture Collection. The bacterium was inoculated on
chocolate agar plates (Becton, Dickinson and Company) at 37°C under
microaerophilic conditions (low levels of dioxygen but
CO2-enriched environment) using an anaerobic chamber
(BBL Campy Pouch® System; Becton, Dickinson and
Company).
Reagents
ASX (Sigma-Aldrich; Merck KGaA) was dissolved in
DMSO at 4 mM and stored in nitrogen gas at −80°C. Before use for
treatment, the ASX stock solution was thawed at 21–23°C and diluted
in 100% FBS to the desired concentrations (final concentration of 1
and 5 µM). The JAK/STAT inhibitor AG490 (cat. no. T3434; R&D
Systems, Inc.) and integrin α5β1 antagonist K34C
(N-(2,6-Dimethylbenzoyl)-O-[3-(2-pyridinylamino)propyl]-L-tyrosine;
cat. no. 5114; R&D Systems, Inc.) were dissolved in DMSO at 50
mM.
Culture of AGS cells with H.
pylori
AGS cells (8×104/2 ml or
4×105/10 ml) were seeded and cultured overnight to
achieve 80% confluency. Whole H. pylori was harvested from
the chocolate agar plates, suspended in antibiotic-free RPMI 1640
medium supplemented with 10% FBS, and subsequently added to the
cells. The cells were culture with H. pylori in a cell
incubator at 37°C in a humidified atmosphere with 95% air and 5%
CO2 at a ratio of 1:50 for 30 min (to determine the
levels of intracellular ROS, p-JAK1, JAK1, p-STAT3 and STAT3), 6 h
(to determine the expression levels of integrin α5 and β1), 20 h
(for the migration assay) and 24 h (for the adhesion assay).
Experimental protocol
AGS cells [1.5×105/2 ml (for measurement
of intracellular ROS levels) or 7.5×105/10 ml (for
Western blot analysis)] were treated at 37°C with ASX (1 or 5 µM)
for 3 h before stimulation with H. pylori. Thereafter, cells
were stimulated with H. pylori at 37°C for 30 min (to
determine the levels of intracellular ROS, p-JAK1, JAK1, p-STAT3
and STAT3), 6 h (to determine the expression levels of integrin α5
and β1), 20 h (for the migration assay) and 24 h (for the adhesion
assay). To determine the involvement of JAK/STAT and integrin in
cell adhesion and migration, the cells were pretreated with AG490
(40 µM) and K34C (20 µM) at 37°C for 2 h and stimulated with H.
pylori at 37°C for 20 h (for the migration assay) and 24 h (for
the adhesion assay). For each experiment, the untreated cells were
cultured with H. pylori for 30 min (to determine the levels
of intracellular ROS, p-JAK1, JAK1, p-STAT3 and STAT3), 6 h (to
determine the expression levels of integrin α5 and β1), 20 h (for
the migration assay) and 24 h (for the adhesion assay) with the
vehicle DMSO (0.1%) alone instead of ASX, AG490 or K34C.
To determine whether AG490 and K34C have an effect
on the inhibitory activity of ASX on integrin α5 and β1 expression
in H. pylori-stimulated cells, cells were pretreated with
ASX (5 µM), AG490 (40 µM) and K34C (20 µM) at 37°C, and then
stimulated with H. pylori at 37°C for 6 h. The pretreatment
period for ASX was 3 h, while that of AG490 or K34C was 2 h.
Protein expression levels of integrin α5 and β1 were determined
using western blot analysis as described below.
To determine the appropriate incubation time for ROS
level, activation of JAK1/STAT3, integrin expression and adhesion
assays, time-course experiments were performed after stimulation of
AGS cells with H. pylori. AGS cells (1.5×105/2
ml/well in 6-well plates) were stimulated with H. pylori at
37°C for 20, 30 and 60 min (for intracellular ROS levels), 15, 30
and 60 min (for total and phospho-specific levels of JAK/STAT
proteins), 2, 4, 6 and 8 h (for integrin expression), or 6, 12, 18
and 24 h (for the adhesion assay).
Measurement of intracellular ROS
levels
The cells were treated with 10 µg/ml
dichlorofluorescein diacetate (Sigma-Aldrich; Merck KGaA) at 37°C
for 30 min. Dichlorofluorescein fluorescence (excitation, 495 nm;
emission, 535 nm) was detected using a Victor3 Multi-label Counter
(PerkinElmer, Inc.) [https://resources.perkinelmer.com/lab-solutions/resources/docs/bro_victor3.pdf].
Intracellular ROS levels were normalized to the cell numbers and
expressed as the relative percentage of control cells.
Normalization was achieved by dividing intracellular ROS levels by
the cell numbers.
Preparation of cell extracts
The cells were harvested using trypsin-EDTA and
pelleted by centrifugation at 1,000 × g at 4°C for 5 min. The cell
pellets were resuspended with lysis buffer containing 10 mM Tris
(pH 7.4), 15 mM NaCl, 1% NP-40 and commercial protease inhibitor
complex (Complete; Roche Diagnostics GmbH), and lysed by drawing
the cells through a 1-ml syringe with several rapid strokes. The
mixture was then incubated at 0°C on ice for 30 min and centrifuged
at 13,000 × g at 4°C for 15 min. The supernatant was collected and
used as whole-cell extracts. The protein concentration was
determined by using the Bradford assay (Bio-Rad Laboratories,
Inc.).
Western blot analysis
Whole-cell extracts (6–40 µg protein/lane) were
loaded per lane, separated using 8–10% SDS-PAGE under reductive
conditions and transferred to nitrocellulose membranes (Amersham;
Cytiva) by electroblotting. Protein transfer was verified using
reversible Ponceau S staining. Membranes were blocked using 3%
non-fat, dry milk in TBS with 0.2% Tween 20 (TBS-T) at 21–23°C for
1 h. The proteins were detected by incubation with antibodies
against integrin α5 (1:500 dilution) (cat. no. 610633; BD
Biosciences), integrin β1 (1:500 dilution) (cat. no. 610467; BD
Biosciences), p-JAK1 (1:300 dilution) (cat. no. 3331S; Cell
Signaling Technology, Inc.), JAK1 (1:500 dilution) (cat. no. 3332S;
Cell Signaling Technology, Inc.), p-STAT3 (1:200 dilution) (cat.
no. 9131S; Cell Signaling Technology, Inc.), STAT3 (1:500 dilution)
(cat. no. 4904S; Cell Signaling Technology, Inc.) and β-actin
(1:2,000 dilution) (cat. no. sc-47778; Santa Cruz Biotechnology,
Inc.) in TBS-T containing 3% dry milk overnight at 4°C. After
washing with TBS-T, the primary antibodies were detected using
HRP-conjugated secondary antibodies [anti-mouse (1:2,000 dilution)
(cat. no. sc-2005 Santa Cruz Biotechnology, Inc.) or anti-rabbit
(1:2,000 dilution) (cat. no. sc-2357 Santa Cruz Biotechnology,
Inc.)] and visualized using the enhanced chemiluminescence
detection system (Santa Cruz Biotechnology, Inc.) based on exposure
to BioMax MR film (Kodak). The enhanced chemiluminescence detection
system is comprised of a two-component kit containing a bottle of
luminol substrate and a bottle of peroxide solution. The solutions
are equilibrated to 21–23°C and then mixed in equal volumes
directly before use. The levels of integrins were compared with
those of β-actin. The levels of p-JAK1 or p-STAT3 were compared
with those of total JAK1 or total STAT3, respectively. Protein band
density was semi-quantified using Image J software (version 1.46;
National Institutes of Health). The data are presented as the mean
± standard error from three immunoblots and are shown as the
relative density of the protein band normalized to the indicated
protein (β-actin or total JAK1 or STAT3).
Adhesion assay
The adhesive potential of AGS cells stimulated with
H. pylori was assessed using human fibronectin-coated
96-well strips (cat. no. ECM101; Sigma-Aldrich; Merck KGaA). Cells
were pretreated with ASX at 37°C for 3 h and then stimulated with
H. pylori at 37°C for 24 h. The cells were detached by
incubation with 6.6 mM EDTA at 37°C for 5 min, washed with PBS and
suspended in serum-free RPMI 1640 containing 0.02% BSA
(Sigma-Aldrich; Merck KGaA) at a density of 1.5×105
cells/ml. The cell suspension (100 µl) was added to each well of
the fibronectin-coated plates and cells were incubated at 37°C for
1 h. The medium was removed from the wells and the cells were
gently washed twice with PBS containing 0.9 mM Ca2+ and
0.5 mM Mg2+. The adherent cells were stained with 0.2%
crystal violet in 10% ethanol for 5 min at 21–23°C and subsequently
washed three times with PBS. After washing, cells were solubilized
using solubilization buffer (a 50/50 mixture of 0.1 M
NaH2PO4, pH 4.5 and 50% ethanol) at 21–23°C
for 10 min, and the absorbance at 570 nm was quantified using a
microplate reader. The percentage of adherent cells in the 0 h or
‘None’ (uninfected cells without any treatment) groups was set as
100%.
Wound-healing assay
Cells were seeded in 12-well plates and allowed to
grow in a complete medium until 90–100% confluency. Subsequently,
individual wells were scratched using a 200-µl micropipette tip to
create a denuded zone (the ‘wound space’ of an artificially
ruptured monolayer of AGS cells) of constant width (1 mm). Complete
media were removed from these cells and replaced with RPMI-1640
containing 4% FBS. The cells were pretreated with 5 or 10 µM ASX at
37°C for 3 h and then stimulated with H. pylori at 37°C for
20 h. To determine cell migration, cell images were captured at
baseline (0 h) and after 20 h of H. pylori stimulation using
phase-contrast light microscopy. The final gap width of the scratch
was measured and compared with the initial gap. The wound area was
quantified using ImageJ software (version 1.46; National Institutes
of Health).
Statistical analysis
Statistical analysis was performed using SPSS
software, version 14 (SPSS, Inc). All values are presented as the
mean ± standard error of three independent experiments. For each
experiment, four samples were used for each group (total number for
each group was 12). One-way ANOVA, followed by Tukey's post-hoc
test, was used for statistical analysis. P<0.05 was considered
to indicate a statistically significant difference.
Results
ASX inhibits integrin α5 expression in
H. pylori-stimulated AGS cells
To determine whether H. pylori induces
integrin α5 and β1 expression, AGS cells were stimulated with H.
pylori for the indicated time periods. As shown in Fig. 1A, H. pylori increased the
expression levels of integrin α5 at 2 h, which continued to 6 h.
Therefore, to measure the effect of ASX on H. pylori-induced
integrin α5 expression, a 6 h-culture period was used. However,
integrin β1 expression was not altered following H. pylori
stimulation. H. pylori-induced integrin α5 expression was
suppressed by ASX treatment (final concentration, 1 and 5 µM) at 6
h (Fig. 1B).
ASX inhibits H. pylori-induced cell
adhesion and migration
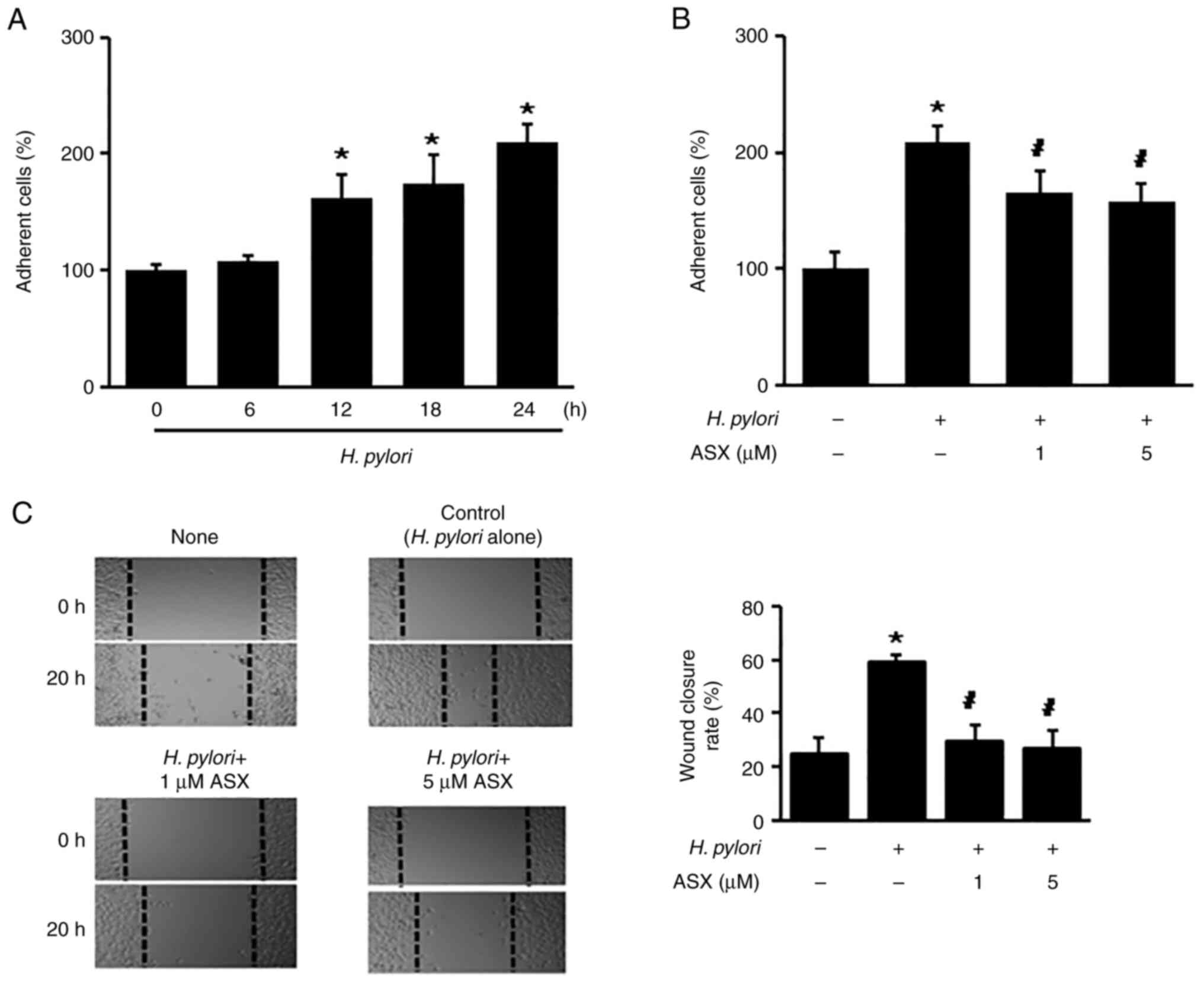
To determine whether H. pylori increases the
number of adherent cells, the cells were stimulated with H.
pylori up to 24 h. H. pylori stimulation significantly
increased the number of adherent cells at 12 h, which continued to
24 h (Fig. 2A). At 24 h of
culture, the number of adherent cells in the H.
pylori-stimulated group was two times higher compared with that
of the unstimulated group (‘H. pylori -, ASX -’) (Fig. 2B). The H. pylori-stimulated
increase in adherent cells was reduced by ASX treatment (both 1 and
5 µM) at 24 h (Fig. 2B).
Since the percentage of wound closure reflects the
migration rate, images were captured at 0 and 20 h after wounding.
As shown in Fig. 2C, H.
pylori increased the rate of wound closure, which was
suppressed by ASX at 20 h.
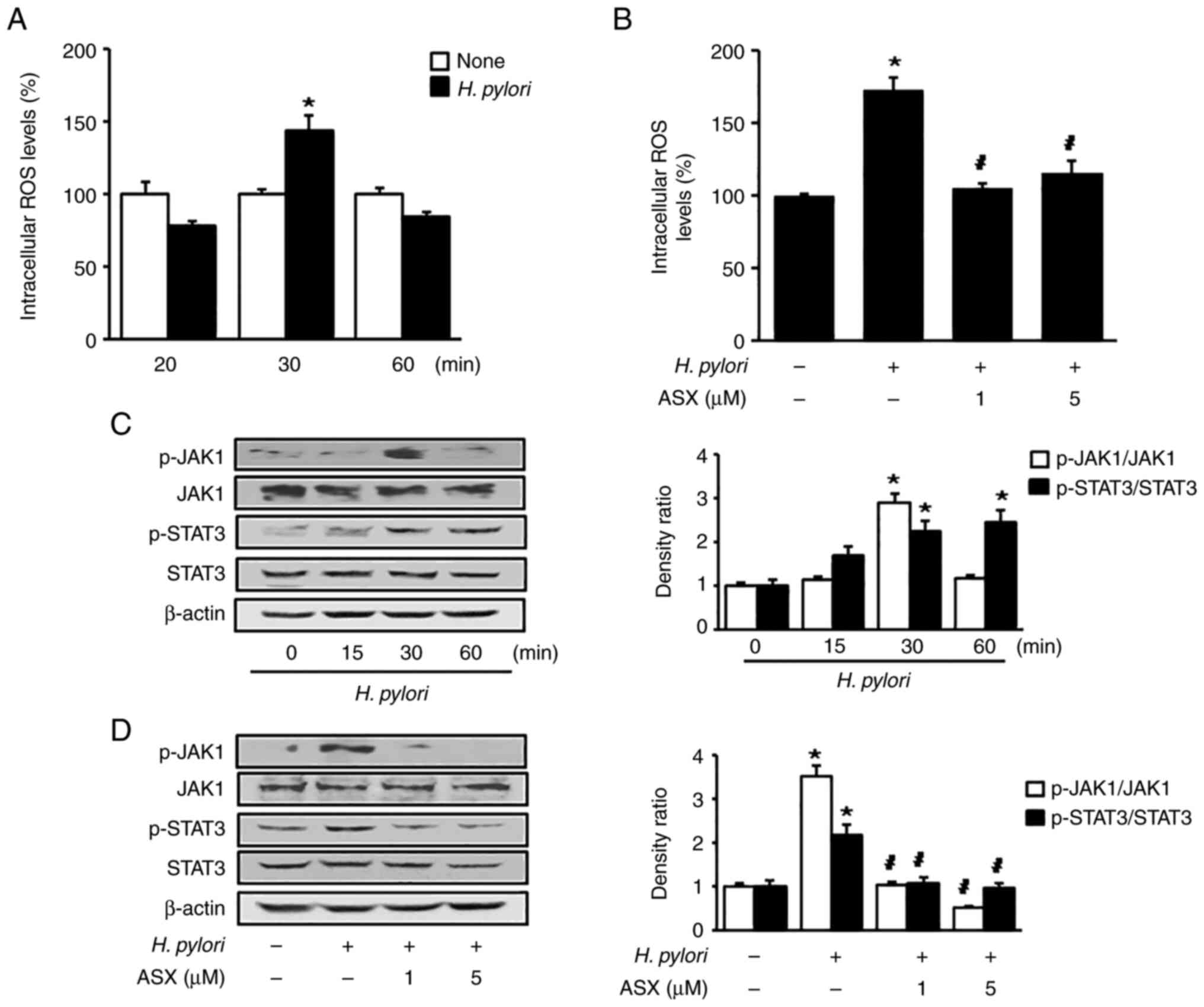
ASX reduces ROS levels and suppresses
activation of JAK1/STAT3 in H. pylori-stimulated cells
As shown in Fig.
3A, H. pylori increased intracellular ROS levels at 30
min in AGS cells. Therefore, a 30-min culture period was used to
examine the effect of ASX on ROS levels in H.
pylori-stimulated cells (Fig.
3B). Both 1 and 5 µM ASX reduced the ROS levels, which were
increased by H. pylori stimulation at 30 min.
Stimulation with H. pylori increased the
p-JAK1/total-JAK1 ratio at 30 min, and this decreased at 60 min
(Fig. 3C). The p-STAT3/total-STAT3
ratio was significantly increased by H. pylori stimulation
up to 60 min. However, the total levels of JAK1 and STAT3 were not
changed by H. pylori. ASX markedly reduced the increased
ratio of p-JAK1/total-JAK1 and p-STAT3/total-STAT3 in H.
pylori-stimulated cells (Fig.
3D).
AG490 or K34C suppress H.
pylori-induced integrin α5 expression, cell adhesion and
migration
To determine the effect of JAK/STAT on integrin α5
expression in H. pylori-stimulated AGS cells, a JAK/STAT
inhibitor, AG490, was used to treat the H. pylori-stimulated
cells. AG490 reduced the H. pylori-induced increase in
integrin α5 levels (Fig. 4A). To
assess whether H. pylori-induced cell adhesion and migration
are mediated by JAK/STAT activation and integrin α5β1, AG490 and
K34C, an integrin α5β1 antagonist, were used. AG490 and K34C
reduced the H. pylori-induced increase in adherent cells and
wound closure levels (Fig. 4B and
C). Therefore, H. pylori may increase cell adhesion and
migration via JAK1/STAT3 activation and integrin α5 expression in
AGS cells.
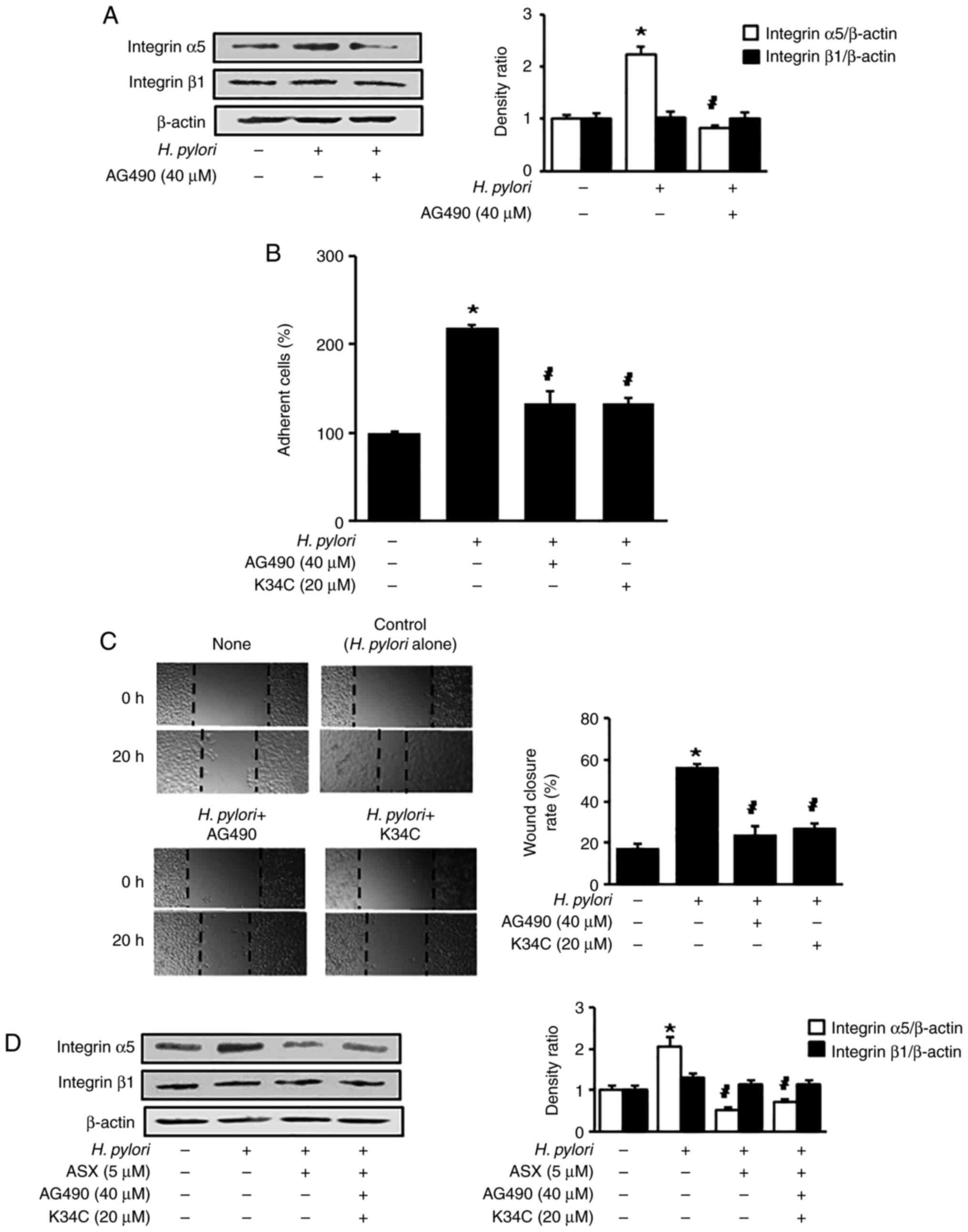 | Figure 4.AG490 and K34C suppress
Helicobacter pylori-induced integrin α5 expression, cell
adhesion and migration. (A) Cells were pretreated with 40 µM AG490
for 2 h, and subsequently stimulated with H. pylori for 6 h.
Protein levels of integrin α5 and β1 were determined using western
blot analysis, with β-actin as the loading control. (B) Cells were
pretreated with 40 µM AG490 or 20 µM K34C for 2 h, and subsequently
stimulated with H. pylori for 24 h. Adherent cells were
stained and absorbance was detected at 570 nm. Results are
presented as the mean ± standard error (the total number for each
group was 12). The percentage of adherent cells in the ‘None’ group
was set as 100%. (C) Cells were pretreated with 40 µM AG490 or 20
µM K34C for 2 h, and subsequently stimulated with H. pylori
for 20 h. The wound healing assay was performed to detect cell
migration. Representative images of wounds in AGS cells were
captured at baseline (0 h) and at 20 h (magnification, ×100). Wound
closure was evaluated by measuring the remaining cell-free area and
expressed as a percentage of the initial cell-free area. The bar
graph indicates wound closure rate (%). Results are presented as
the mean ± SE (the total number for each group was 12). (D) Cells
were pretreated with ASX (5 µM), AG490 (40 µM) and K34C (20 µM),
and then stimulated with H. pylori for 6 h. The pretreatment
period for ASX was 3 h, while that of AG490 or K34C was 2 h.
Protein levels of integrin α5 and β1 were determined using western
blot analysis, with β-actin as the loading control. The
densitometry data are presented as the mean ± standard error from
three immunoblots, and are shown as the relative density of protein
bands normalized to β-actin. *P<0.05 vs. ‘None’ (unstimulated
cells without treatment of AG490, K34C or ASX).
#P<0.05 vs. ‘Control’ (H. pylori-stimulated
cells without treatment of AG490, K34C or ASX). ASX,
astaxanthin. |
To determine whether AG490 and K34C have an effect
on the inhibitory activity of ASX on the expression of integrin α5
in H. pylori-stimulated cells, the cells were pretreated
with ASX, AG490 and K34C, followed by H. pylori stimulation
for 6 h. The inhibitory activity of ASX on expression of integrin
α5 was not changed by addition of AG490 in H.
pylori-stimulated cells (Fig.
4D). As shown in Figs. 1B and
3D, the levels of integrin α5
expression, p-JAK1/total-JAK1 ratio, and p-STAT3/total-STAT3 ratio
in H. pylori-stimulated cells treated with ASX (5 µM) were
lower than those of unstimulated cells (H. pylori -, ASX -).
As shown in Fig. 4A, the level of
integrin α5 expression in H. pylori-stimulated cells treated
with a JAK/STAT inhibitor AG490 was similar to unstimulated cells.
The results demonstrate that ASX act as an inhibitor of JAK1/STAT3
like AG490, but the inhibitory effect of ASX (5 µM) for integrin α5
expression was higher than the effect of AG490 (40 µM). Thus, the
addition of AG490 to ASX did not have synergistic effect on
inhibition by ASX on integrin α5 expression in H.
pylori-stimulated cells. K34C is a selective ligand for
integrin α5β1 (34), which
inhibits functions of integrin α5β1 including migration in glioma
cells (35). As shown in Fig. 4B and C, cell adhesion and migration
in H. pylori-stimulated cells treated with of K34C (20 µM)
were similar to the levels in unstimulated cells. Similarly, ASX
treatment reduced H. pylori-induced increases in adherent
cells and migration (Fig. 1B and
C). These results indicate that ASX may inhibit functions of
integrin α5β1 since ASX reduced the expression of integrin α5 in
H. pylori-stimulated cells. Even though K34C is a selective
functional inhibitor of integrin α5β1, further study is necessary
to determine whether K34C reduces expression of integrin α5 in
H. pylori-stimulated cells.
Discussion
In the present study, ASX inhibited H.
pylori-induced integrin α5 expression, cell adhesion and
migration by suppressing ROS-mediated activation of JAK1/STAT3 in
gastric epithelial AGS cells. In addition, both a JAK/STAT
inhibitor (AG490) and an integrin α5β1 antagonist (K34C) suppressed
cell adhesion and migration in H. pylori-stimulated AGS
cells.
For the virulence factor of H. pylori, cag A
is a unique genomic fragment containing ~30 genes and is required
for the secretion of the cag A protein (36). Vac A induces cytoplasmic
vacuolation in gastric epithelial cells (37) and is responsible for apoptosis in
gastric epithelial cells (38).
There are different allelic variants in the vac A sequence; three
different families of vacA signal sequences (s1a, s1b and s2) and
two different families of middle-region alleles (m1 and m2)
(39). These variants are related
to the differences in cytotoxin production and clinical outcomes of
the H. pylori infection (40–42).
Regarding integrin expression in H.
pylori-stimulated gastric epithelial cells, H. pylori in
a Korean isolate (HP99) stimulated integrin α5 and β1 expression in
gastric epithelial cells (22,23).
However, H. pylori (NCTC11637) upregulated integrin α5, but
not integrin β1, in gastric epithelial cells (24), which supports the present findings
using NCTC11637. In a previous study by our group, HP99 was
identified as H. pylori cag A+, vac A+ (s1, m2) strain
(43), while NCTC11637 is cag A+,
vac A+ (s1, m1) strain (44).
These different allelic variants in the vac A sequence may affect
the expression of integrin α5 and β1 expression in gastric
epithelial cells.
In regard to integrin expression, cell adhesion and
the virulence factor of H. pylori, Su et al (45) revealed that antibodies against α5-
and β1-integrin decreased adherence in AGS cells. Miyata et
al (46) also demonstrated the
relation of integrin α5β1 expression and adhesion to fibronectin in
gastric cancer cells. In H. pylori-stimulated cells, the
virulence factor cag A mediates integrin α5β1 expression in
infected gastric epithelial cells (47). These studies showed the close
relation of cag A, integrin α5 expression and cell adhesion in
gastric epithelial cells.
To investigate the levels of ROS and JAK/STAT3, a 30
min-culture period was used, since increased ROS levels and
activation of JAK1/STAT3 were observed after 30 min of stimulation
with H. pylori. As aforementioned, cag A phosphorylation is
important for integrin-mediated cell adhesion in gastric epithelial
cells (46). Backert et al
(48) found that tyrosine
phosphorylation of the cag A protein occurred at 15 min and
increased until 180 min in H. pylori-stimulated gastric
epithelial cells. Tsugawa et al (49) demonstrated that phosphorylated cag
A was dephosphorylated by the H. pylori vac A, which exerted
its effect by reducing intracellular glutathione and increasing ROS
in gastric epithelial cells. Further studies should be performed to
determine whether H. pylori cag A is translocated into AGS
cells after 30 min of stimulation with H. pylori.
ROS contribute to the development of gastric cancer
and ROS levels are elevated in patients with gastric cancer
(50,51). H. pylori stimulation
increases ROS levels in gastric epithelial cells (52–54),
which activates the JAK1/STAT3 signaling pathway to induce cytokine
IL-8 expression in gastric epithelial cells (55).
JAK/STAT signaling serves a ciritcal role in the
pathogenesis of solid tumour development (11). Furthermore, constitutively
activated STAT3 increases integrin β6 expression, which mediates
tumorigenesis and enhances cell motility of prostate epithelial
cells (15). JAK2-STAT5 signaling
mediates the integrin-dependent migration of mesenchymal stem cells
in ischemic cerebral lesions in vivo (56). These findings suggest a potential
association among JAK/STAT, integrins and adhesion/migration in
gastric cancer development (11,15,56).
In the present study, integrin α5 expression was decreased by
treatment with AG490, a JAK/STAT inhibitor. These results suggest
that the JAK1/STAT3 signaling pathway may regulate integrin α5
expression. AG490 also inhibited integrin-mediated cell adhesion
and migration of H. pylori-stimulated AGS cells. The present
findings demonstrated the activation of JAK1/STAT3, as examined by
detecting phosphorylation of JAK1/STAT3, in H.
pylori-stimulated cells. Further studies are necessary to
determine the translocation of STAT3 to the nucleus to induce
integrin α5 expression in H. pylori-stimulated AGS
cells.
ASX suppressed JAK1/STAT-3 activation to inhibit
invasion and angiogenesis in an oral cancer model (57). Another study reported the
protective effect of ASX against breast cancer cells in
vitro through the suppression of proliferation and migration
(58). To the best of our
knowledge, the novel finding of the present study is that the
antioxidant effect of ASX may attenuate JAK1/STAT3 activation,
integrin α5 protein expression, cell adhesion and migration in
H. pylori-stimulated gastric epithelial cells.
Regarding the dosage of ASX, 5 µM ASX inhibits
oxidative stress-induced cell death by reducing ROS levels and
inhibiting the degradation of DNA repair protein Ku in gastric
epithelial cells (59).
Furthermore, 5 µM ASX inhibits H. pylori-induced IL-8
expression by activating PPAR-γ and its target gene catalase in AGS
cells (32). Our previous study
demonstrated that 5 µM ASX did not induce cell death, while 10 and
20 µM ASX decreased the viable cell numbers in gastric epithelial
AGS cells (60). Therefore, 1 and
5 µM ASX were used to determine the effect of ASX on integrin
expression in H. pylori-stimulated gastric epithelial cells
in the present study.
In the present study, AGS cells were cultured with
H. pylori. JAK1/STAT3 signaling was activated in H.
pylori-stimulated cells. Backert et al (48) and Tsugawa et al (49) determined the presence of cag A or
vac A in gastric epithelial cells after stimulation with H.
pylori. Therefore, further studies should be performed to
determine the presence of H. pylori virulence factors in
H. pylori-stimulated AGS cells to confirm the infection of
H. pylori in AGS cells.
In conclusion, ASX suppressed H.
pylori-induced cell adhesion and migration by suppressing
integrin α5 expression via the ROS-mediated JAK1/STAT3 signaling
pathway in human gastric epithelial cells.
Acknowledgements
Not applicable.
Funding
The present study was supported by the BK21 FOUR project, Yonsei
University, Republic of Korea.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
HK conceived and designed the experiments. JWL
assisted in the experimental design. JW performed the experiments.
JW and JWL analyzed the data. JW and JWL confirmed the authenticity
of all raw data. JW wrote the manuscript. JWL and HK reviewed and
edited the manuscript. JWL designed the additional experiments for
the revised manuscript. All authors have read and approved the
final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Kusters JG, van Vliet AH and Kuipers EJ:
Pathogenesis of Helicobacter pylori infection. Clin
Microbiol Rev. 19:449–490. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Handa O, Naito Y and Yoshikawa T:
Helicobacter pylori: A ROS-inducing bacterial species in the
stomach. Inflamm Res. 59:997–1003. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Cha B, Lim JW, Kim KH and Kim H:
15-deoxy-D12,14-prostaglandin J2 suppresses RANTES expression by
inhibiting NADPH oxidase activation in Helicobacter
pylori-infected gastric epithelial cells. J Physiol Pharmacol.
62:167–174. 2011.PubMed/NCBI
|
|
4
|
Cha B, Lim JW, Kim KH and Kim H: HSP90beta
interacts with Rac1 to activate NADPH oxidase in Helicobacter
pylori-infected gastric epithelial cells. Int J Biochem Cell
Biol. 42:1455–1461. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Halliwell B: Reactive oxygen species in
living systems: Source, biochemistry, and role in human disease. Am
J Med. 91:14S–22S. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Snezhkina AV, Kudryavtseva AV, Kardymon
OL, Savvateeva MV, Melnikova NV, Krasnov GS and Dmitriev AA: ROS
generation and antioxidant defense systems in normal and malignant
cells. Oxid Med Cell Longev. 2019:61758042019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Fiedor J and Burda K: Potential role of
carotenoids as antioxidants in human health and disease. Nutrients.
6:466–488. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sies H and Jones DP: Reactive oxygen
species (ROS) as pleiotropic physiological signalling agents. Nat
Rev Mol Cell Biol. 21:363–383. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Venkatabalasubramanian S: The complex
interplay between JAK-STAT pathway and ROS in regulating stem cells
during inflammation and cancer. Chakraborti S: Handbook of
Oxidative Stress in Cancer: Therapeutic Aspects. Springer;
Singapore: pp. 1–12. 2022
|
|
10
|
Charras A, Arvaniti P, Le Dantec C,
Dalekos GN, Zachou K, Bordron A and Renaudineau Y: JAK inhibitors
and oxidative stress control. Front Immunol. 10:28142019.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Thomas SJ, Snowden JA, Zeidler MP and
Danson SJ: The role of JAK/STAT signalling in the pathogenesis,
prognosis and treatment of solid tumours. Br J Cancer. 113:365–371.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Pfitzner E, Kliem S, Baus D and Litterst
CM: The role of STATs in inflammation and inflammatory diseases.
Curr Pharm Des. 10:2839–2850. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yu H, Kortylewski M and Pardoll D:
Crosstalk between cancer and immune cells: Role of STAT3 in the
tumour microenvironment. Nat Rev Immunol. 7:41–51. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kamran MZ, Patil P and Gude RP: Role of
STAT3 in cancer metastasis and translational advances. Biomed Res
Int. 2013:4218212013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Azare J, Leslie K, Al-Ahmadie H, Gerald W,
Weinreb PH, Violette SM and Bromberg J: Constitutively activated
Stat3 induces tumorigenesis and enhances cell motility of prostate
epithelial cells through integrin beta 6. Mol Cell Biol.
27:4444–4453. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Park EJ, Myint PK, Ito A, Appiah MG,
Darkwah S, Kawamoto E and Shimaoka M: Integrin-ligand interactions
in inflammation, cancer, and metabolic disease: Insights into the
multifaceted roles of an emerging ligand irisin. Front Cell Dev
Biol. 8:5880662020. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hou J, Yan D, Liu Y, Huang P and Cui H:
The roles of integrin α5β1 in human cancer. Onco Targets Ther.
13:13329–13344. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hynes RO: Integrins: Bidirectional,
allosteric signaling machines. Cell. 110:673–687. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Gallant ND, Michael KE and García AJ: Cell
adhesion strengthening: Contributions of adhesive area, integrin
binding, and focal adhesion assembly. Mol Biol Cell. 16:4329–4340.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Bendas G and Borsig L: Cancer cell
adhesion and metastasis: Selectins, integrins, and the inhibitory
potential of heparins. Int J Cell Biol. 2012:6767312012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yang Y, Wang Y, Che X, Hou K, Wu J, Zheng
C, Cheng Y, Liu Y, Hu X and Zhang J: Integrin α5 promotes migration
and invasion through the FAK/STAT3/AKT signaling pathway in
icotinib-resistant non-small cell lung cancer cells. Oncol Lett.
22:5562021. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Cho SO, Kim KH, Yoon JH and Kim H:
Signaling for integrin alpha5/beta1 expression in Helicobacter
pylori-infected gastric epithelial AGS cells. Ann N Y Acad Sci.
1090:298–304. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Seo JH, Lim JW, Yoon JH and Kim H:
Proteinase-activated receptor-2 mediates the expression of integrin
alpha5 and beta1 in Helicobacter pylori-infected gastric
epithelial AGS cells. Digestion. 80:40–49. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lim JW, Kim H and Kim KH: Cell
adhesion-related gene expression by Helicobacter pylori in
gastric epithelial AGS cells. Int J Biochem Cell Biol.
35:1284–1296. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Cha B, Lim JW and Kim H: Jak1/Stat3 is an
upstream signaling of NF-κB activation in Helicobacter
pylori-induced IL-8 production in gastric epithelial AGS cells.
Yonsei Med J. 56:862–866. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Higuera-Ciapara I, Félix-Valenzuela L and
Goycoolea FM: Astaxanthin: A review of its chemistry and
applications. Crit Rev Food Sci Nutr. 46:185–196. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hussein G, Sankawa U, Goto H, Matsumoto K
and Watanabe H: Astaxanthin, a carotenoid with potential in human
health and nutrition. J Nat Prod. 69:443–449. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Sztretye M, Dienes B, Gönczi M, Czirják T,
Csernoch L, Dux L, Szentesi P and Keller-Pintér A: Astaxanthin: A
potential mitochondrial-targeted antioxidant treatment in diseases
and with aging. Oxid Med Cell Longev. 2019:38496922019. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Faraone I, Sinisgalli C, Ostuni A,
Armentano MF, Carmosino M, Milella L, Russo D, Labanca F and Khan
H: Astaxanthin anticancer effects are mediated through multiple
molecular mechanisms: A systematic review. Pharmacol Res.
155:1046892020. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Chen YT, Kao CJ, Huang HY, Huang SY, Chen
CY, Lin YS, Wen ZH and Wang HMD: Astaxanthin reduces MMP
expressions, suppresses cancer cell migrations, and triggers
apoptotic caspases of in vitro and in vivo models in melanoma. J
Func Foods. 31:20–31. 2017. View Article : Google Scholar
|
|
31
|
Kowshik J, Nivetha R, Ranjani S,
Venkatesan P, Selvamuthukumar S, Veeravarmal V and Nagini S:
Astaxanthin inhibits hallmarks of cancer by targeting the
PI3K/NF-κΒ/STAT3 signalling axis in oral squamous cell carcinoma
models. IUBMB Life. 71:1595–1610. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kim SH, Lim JW and Kim H: Astaxanthin
inhibits mitochondrial dysfunction and interleukin-8 expression in
Helicobacter pylori-infected gastric epithelial cells.
Nutrients. 10:13202018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kim SH, Lim JW and Kim H: Astaxanthin
prevents decreases in superoxide dismutase 2 level and superoxide
dismutase activity in Helicobacter pylori-infected gastric
epithelial cells. J Cancer Preven. 24:54–58. 2019. View Article : Google Scholar
|
|
34
|
Martinkova E, Maglott A, Leger DA, Bonnet
D, Stiborova M, Takeda K, Martin S and Dontenwill M: alpha5beta1
integrin antagonists reduce chemotherapy-induced premature
senescence and facilitate apoptosis in human glioblastoma cells.
Int J Cancer. 127:1240–1248. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Renner G, Noulet F, Mercier MC, Choulier
L, Etienne-Selloum N, Gies JP, Lehmann M, Lelong-Rebel I, Martin S
and Dontenwill M: Expression/activation of α5β1 integrin is linked
to the β-catenin signaling pathway to drive migration in glioma
cells. Oncotarget. 7:62194–62207. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Diaz MI, Valdivia A, Martinez P, Palacios
JL, Harris P, Novales J, Garrido E, Valderrama D, Shilling C,
Kirberg A, et al: Helicobacter pylori vacA s1a and s1b
alleles from clinical isolates from different regions of Chile show
a distinct geographic distribution. World J Gastroenterol.
11:6366–6372. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Leunk RD, Johnson PT, David BC, Kraft WG
and Morgan DR: Cytotoxic activity in broth-culture filtrates of
Campylobacter pylori. J Med Microbiol. 26:93–99. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Chiozzi V, Mazzini G, Oldani A, Sciullo A,
Ventura U, Romano M, Boquet P and Ricci V: Relationship between Vac
A toxin and ammonia in Helicobacter pylori-induced apoptosis
in human gastric epithelial cells. J Physiol Pharmacol. 60:23–30.
2009.PubMed/NCBI
|
|
39
|
Atherton JC, Cao P, Peek RM Jr, Tummuru
MK, Blaser MJ and Cover TL: Mosaicism in vacuolating cytotoxin
alleles of Helicobacter pylori: Association of specific vacA
types with cytotoxin production and peptic ulceration. J Biol Chem.
270:17771–17777. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Atherton JC, Peek RM Jr, Tham KT, Cover TL
and Blaser MJ: Clinical and pathological importance of
heterogeneity in vacA, the vacuolating cytotoxin gene of
Helicobacter pylori. Gastroenterology. 112:92–99. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
McClain MS, Cao P, Iwamoto H,
Vinion-Dubiel AD, Szabo G, Shao Z and Cover TL: A 12-amino-acid
segment, present in type s2 but not type s1 Helicobacter
pylori VacA proteins, abolishes cytotoxin activity and alters
membrane channel formation. J Bacteriol. 183:6499–6508. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Telford JL, Ghiara P, Dell'Orco M,
Comanducci M, Burroni D, Bugnoli M, Tecce MF, Censini S, Covacci A,
Xiang Z, et al: Gene structure of the Helicobacter pylori
cytotoxin and evidence of its key role in gastric disease. J Exp
Med. 179:1653–1658. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Seo JH, Lim JW, Kim H and Kim KH:
Helicobacter pylori in a Korean isolate activates
mitogen-activated protein kinases, AP-1, and NF-kappaB and induces
chemokine expression in gastric epithelial AGS cells. Lab Invest.
84:49–62. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Xerry J and Owen RJ: Conservation and
microdiversity of the phospholipase A (pldA) gene of
Helicobacter pylori infecting dyspeptics from different
countries. FEMS Immunol Med Microbiol. 32:17–25. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Su B, Johansson S, Fällman M, Patarroyo M,
Granström M and Normark S: Signal transduction-mediated adherence
and entry of Helicobacter pylori into cultured cells.
Gastroenterology. 117:595–604. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Miyata S, Koshikawa N, Yasumitsu H and
Miyazaki K: Trypsin stimulates integrin alpha(5)beta(1)-dependent
adhesion to fibronectin and proliferation of human gastric
carcinoma cells through activation of proteinase-activated
receptor-2. J Biol Chem. 275:4592–4598. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Yeh YC, Cheng HC, Yang HB, Chang WL and
Sheu BS: H. pylori CagL-Y58/E59 Prime Higher Integrin α5β1 in
Adverse pH Condition to Enhance Hypochlorhydria Vicious Cycle for
Gastric Carcinogenesis. PLoS One. 8:e727352013. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Backert S, Ziska E, Brinkmann V,
Zimny-Arndt U, Fauconnier A, Jungblut PR, Naumann M and Meyer TF:
Translocation of the Helicobacter pylori CagA protein in
gastric epithelial cells by a type IV secretion apparatus. Cell
Microbiol. 2:155–164. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Tsugawa H, Suzuki H, Saya H, Hatakeyama M,
Hirayama T, Hirata K, Nagano O, Matsuzaki J and Hibi T: Reactive
oxygen species-induced autophagic degradation of Helicobacter
pylori CagA is specifically suppressed in cancer stem-like
cells. Cell Host Microbe. 12:764–777. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Gu H, Huang T, Shen Y, Liu Y, Zhou F, Jin
Y, Sattar H and Wei Y: Reactive oxygen species-mediated tumor
microenvironment transformation: The mechanism of radioresistant
gastric cancer. Oxid Med Cell Longev. 2018:58012092018. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Liou GY and Storz P: Reactive oxygen
species in cancer. Free Radic Res. 44:479–496. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Ding SZ, Minohara Y, Fan XJ, Wang J, Reyes
VE, Patel J, Dirden-Kramer B, Boldogh I, Ernst PB and Crowe SE:
Helicobacter pylori infection induces oxidative stress and
programmed cell death in human gastric epithelial cells. Infect
Immun. 75:4030–4039. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Smoot DT, Elliott TB, Verspaget HW, Jones
D, Allen CR, Vernon KG, Bremner T, Kidd LCR, Kim KS, Groupman JD
and Ashktorab H: Influence of Helicobacter pylori on
reactive oxygen-induced gastric epithelial cell injury.
Carcinogenesis. 21:2091–2095. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Shimoyama T, Fukuda S, Liu Q, Nakaji S,
Fukuda Y and Sugawara K: Production of chemokines and reactive
oxygen species by human neutrophils stimulated by Helicobacter
pylori. Helicobacter. 7:170–174. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Choi JH, Cho SO and Kim H: α-Lipoic acid
inhibits expression of IL-8 by suppressing activation of MAPK,
Jak/Stat, and NF-κB in H. pylori-infected gastric epithelial
AGS cells. Yonsei Med J. 57:260–264. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Zhang Y, Zheng J, Zhou Z, Zhou H, Wang Y,
Gong Z and Zhu J: Fractalkine promotes chemotaxis of bone
marrow-derived mesenchymal stem cells towards ischemic brain
lesions through Jak2 signaling and cytoskeletal reorganization.
FEBS J. 282:891–903. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Kowshik J, Baba AB, Giri H, Deepak Reddy
G, Dixit M and Nagini S: Astaxanthin inhibits JAK/STAT-3 signaling
to abrogate cell proliferation, invasion and angiogenesis in a
hamster model of oral cancer. PLoS One. 9:e1091142014. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
McCall B, McPartland CK, Moore R,
Kamenetskii AF and Booth BW: Effects of astaxanthin on the
proliferation and migration of breast cancer cells in vitro.
Antioxidants (Basel). 7:1352018. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Lee J, Lim JW and Kim H: Astaxanthin
inhibits oxidative stress-induced Ku protein degradation and
apoptosis in gastric epithelial cells. Nutrients. 14:39392022.
View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Kim S, Lee H, Lim JW and Kim H:
Astaxanthin induces NADPH oxidase activation and
receptor-interacting protein kinase 1-mediated necroptosis in
gastric cancer AGS cells. Mol Med Rep. 24:8372021. View Article : Google Scholar : PubMed/NCBI
|