Introduction
In China, the morbidity and mortality rates of
patients with diabetes are increasing on a yearly basis, and
diabetes has become a major threat to the health of the Chinese
populace (1). Diabetic liver
injury (DLI) is a major complication of type 2 diabetes mellitus
(T2DM) caused by a variety of factors, such as hyperglycemia,
insulin resistance, dyslipidemia, oxidative stress, and
inflammation (2). During the
development of diabetes, DLI can progress from simple fatty liver
to non-alcoholic steatohepatitis, liver fibrosis, cirrhosis, and
even liver cancer (3).
Ionizing radiation (IR) is one of the most important
environmental factors. High-dose ionizing radiation (HDIR) may
cause severe tissue and organ damage and induce apoptosis. HDIR may
also cause genomic instability and promote tumorigenesis (4). Low-dose ionizing radiation (LDIR) is
typically defined as ≤0.2 Gy at low linear energy transfer (LET) or
≤0.05 Gy at high LET (5). LDIR has
been shown to exhibit a hormetic and adoptive effect (6–8).
Studies have shown that LDIR is associated with cell growth and
development, immunoregulation, and disease prevention (9). Our previous study also indicated that
LDIR stimulated the proliferation of normal cells, such as rat
mouse bone marrow hematopoietic progenitor cells, mesenchymal stem
cells, and several normal human cell lines (9,10),
instead of promoting tumorigenesis (10).
MicroRNA-155 (miRNA/miR-155) resides within the
B-cell integration cluster gene on human chromosome 21 (11). miR-155 is one of the most
multifunctional miRNAs, and its sequence is highly conserved in
evolution. It has been shown that miR-155 is abnormally expressed
in several physiological and pathological processes, including
hematopoietic lineage differentiation, immune response,
inflammation, and tumorigenesis (12). miR-155 is also closely related to
the proliferation, differentiation, invasion, and prognosis of
tumor cells (13). A large number
of studies have shown that abnormal expression of miR-155 is
closely associated with viral hepatitis, alcoholic liver disease,
non-alcoholic steatohepatitis, liver fibrosis, hepatocellular
carcinoma, and other liver diseases (14–17).
Substantial evidence indicates that miR-155 plays a
crucial role in the pathogenesis of diabetes and its complications
(18). For example, El Samaloty
et al (19) reported a
higher expression level of miR-155 in patients with chronic
hepatitis C virus (HCV) infection, but a significant decrease in
diabetic HCV patients; Bai et al (20) reported that miR-155 expression was
increased in patients with diabetic nephropathy compared to T2DM
patients without diabetic nephropathy. However, there is limited
research that focuses on the impact of LDIR on diabetes and its
associated complications (21–23).
In particular, whether LDIR may modulate the expression of miR-155
during the treatment of diabetes and its complications should be
assessed. Therefore, in the present study, the effect of HG on
hepatocytes and how the miR-155-SOCS1 axis impacted the behavior of
hepatocytes during this process was assessed. Additionally, the
protective effect of LDIR on hepatocytes via the regulation of
miR-155-SOCS1 axis was assessed. The aim of the present was to
explore a novel potential strategy for protection against and
treatment of DLI.
Material and methods
Cell lines and cell culture
Mouse AML12 hepatocytes were obtained from Procell
Life Science and Technology Co., Ltd. Cells were grown in DMEM/F12
supplemented with 10% FBS (Procell Life Science and Technology Co.,
Ltd.) and penicillin (100 U/ml)-streptomycin (100 µg/ml)
(Life-ilab). To construct a high glucose-induced hepatocyte injury
model, AML12 cells were cultured in normal glucose (NG) (5.6 mM +
19.4 mM mannitol to adjust for osmotic pressure) or HG (30 mM) to
120 h (24,25). All cells were maintained in a
humidified 37°C incubator with 5% CO2.
Low-dose ionizing radiation
AML12 cells received a total dose of 75 mGy X-ray at
a dose rate of 25 mGy/min using an X-RAD 320 (Precision X-RAD;
Precision X-Ray). Following LDIR, the tissue culture medium was
replaced and the cells were further cultured under the same
conditions until cells were required for subsequent
experiments.
Apoptosis assay
Apoptosis was analyzed using an Annexin V-FITC
Apoptosis Detection Kit (BD Biosciences). According to the
manufacturer's instructions, 1×105 AML12 cells were
washed with binding buffer and stained with 50 µl staining solution
containing 5 µl Annexin V-FITC and 5 µl PI at room temperature for
15 min in the dark. Apoptosis was measured using a FACScan flow
cytometer (BD Biosciences) and analyzed using FlowJo software
(version 10.0; FlowJo LLC).
Transfection of miR-155 mimics,
miR-155 inhibitor, and siRNA targeting SOCS1
AML12 cells were transfected with a 21 (siSOCS1#1)
or 19 (siSOCS1#2) base-pair siRNA targeting SOCS1
[5′-CTACCTGAGTTCCTTCCCCTT-3′ (26)
and 5′-ACACTCACTTCCGCACCTT-3′ (27)] using X-tremeGENE siRNA transfection
reagent (MilliporeSigma), according to the manufacturer's protocol.
At 48 h-post siRNA transfection, total protein was collected from
the transfected cells and the expression levels of SOCS1 were
western blotting. The siRNA sequence with the better interference
result was chosen for subsequent experiments. miR-155 mimics,
NC-mimics, miR-155 inhibitor, and NC-inhibitor were synthesized by
Guangzhou RiboBio Co., Ltd. and transfected into cells at a
concentration of 10 nmol/ml with X-tremeGENE siRNA transfection
reagent. The subsequent experiments were performed at 48 h
post-transfection.
RNA extraction and reverse
transcription quantitative-PCR (RT-qPCR)
According to the manufacturer's instructions, total
RNA was extracted from cultured AML12 cells using an Eastep Super
Total RNA Extraction kit (Promega Corporation), and 1 µg total RNA
was used for cDNA synthesis using a TransScript miRNA RT Enzyme Mix
(TransGen Biotech Co., Ltd.).
qPCR was used to detect the enrichment of miRNAs
using an Eastep qPCR MasterMix (Promega Corporation) according to
the manufacturer's instructions. Data were analyzed using the
2−ΔΔCq method (28),
and U6 snRNA expression was used as the endogenous control. The
forward sequences of the primers were: miR-155,
5′-ttaatgctaattgtgatagg-3′ and for U6, 5′-CTCGCTTCGGCAGCACATA-3′.
The reverse primers were included in the cDNA synthesis kit.
Western blot analysis
AML12 cells were washed twice with ice-cold PBS and
total protein was collected using RIPA lysis buffer (Beyotime
Institute of Biotechnology). Protein concentration was determined
with an enhanced bicinchoninic acid protein assay kit (Beyotime
Institute of Biotechnology). A total of 25–50 µg protein was loaded
on a 5–12% SDS gel and resolved SDS-PAGE (Epizyme) and transferred
to a PVDF membrane (MilliporeSigma). Subsequently, the membranes
were blocked in TBST containing 5% BSA at room temperature for 1 h
after which the blots were probed with primary antibodies against
SOCS1 (cat. no. 3950; 1:1,000; all from Cell Signaling Technology,
Inc.) and β-actin (cat. no. 81115; 1:1,000; ProteinTech Group,
Inc.) at 4°C overnight. Subsequently, the membranes were incubated
with horseradish peroxidase-conjugated secondary antibodies (cat.
no. PR30012; 1:3,000; ProteinTech Group, Inc.), and the peroxidase
activity was visualized using an enhanced chemiluminescence reagent
(Life-ilab). The mean band intensity was determined using
densitometry analysis in Quantity One software (version 4.6;
Bio-Rad Laboratories, Inc.).
Dual-luciferase reporter assay
Luciferase assays were implemented by adopting the
Dual-Luciferase Reporter Assay System (Promega Corporation) in
accordance with the manufacturer's instructions. The wild type (WT)
or mutant (MT) SOCS1 3′-UTR sequences including the miR-155
targeting site were inserted into the pGL3 vector (Invitrogen;
Thermo Fisher Scientific, Inc.) to construct pGL3-luc-SOCS1-WT and
pGL3-luc-SOCS1-MT. AML12 cells were transfected with 50 ng
pGL3-luc-SOCS1-WT/MT, 5 pmol miR-155 mimic or NC mimics (control,)
and 5 ng Renilla luciferase using adopting Lipofectamine 2000
(Invitrogen; Thermo Fisher Scientific, Inc.) and seeded in a
96-well plate. The luciferase activities were detected 48 h
post-transfection on a GloMax system (Promega Corporation).
ELISA
Mouse TNF-α (cat. no. MTA00B), IL-1β (cat. no.
MLB00C), IL-6 (cat. no. M6000B), IL-10 (cat. no. M1000B), and IFN-γ
(cat. no. MIF00) were detected according to the manufacturer's
protocol of the ELISA kits (R&D Systems China Co., Ltd.).
Statistical analysis
Data are presented as the mean ± SD of at least
three repeats. SPSS 19.0 (IBM Corp.) was used to analyze the
experimental data. Differences between groups were compared using a
Student's t-test or a one-way ANOVA followed by a post hoc LSD or
Tukey's test. P<0.05 was considered to indicate a statistically
significant difference.
Results
LDIR attenuates HG-induced miR-155
upregulation
Previous studies indicated that the levels of
miR-155 were upregulated in T1DM and downregulated in T2DM patients
(19,29–31).
However, whether miR-155 levels are altered in DLI remains to be
determined. To determine whether miR-155 could be induced by HG in
hepatocytes, AML12 cells were treated with 30 mM glucose for 24,
48, 72, 96, and 120 h, and the expression levels of miR-155 were
measured. As shown in Fig. 1A, the
levels of miR-155 were upregulated following HG treatment, with
expression peaking at 48 h post-HG treatment, where it was 2×
higher than that in the NG group, P<0.001.
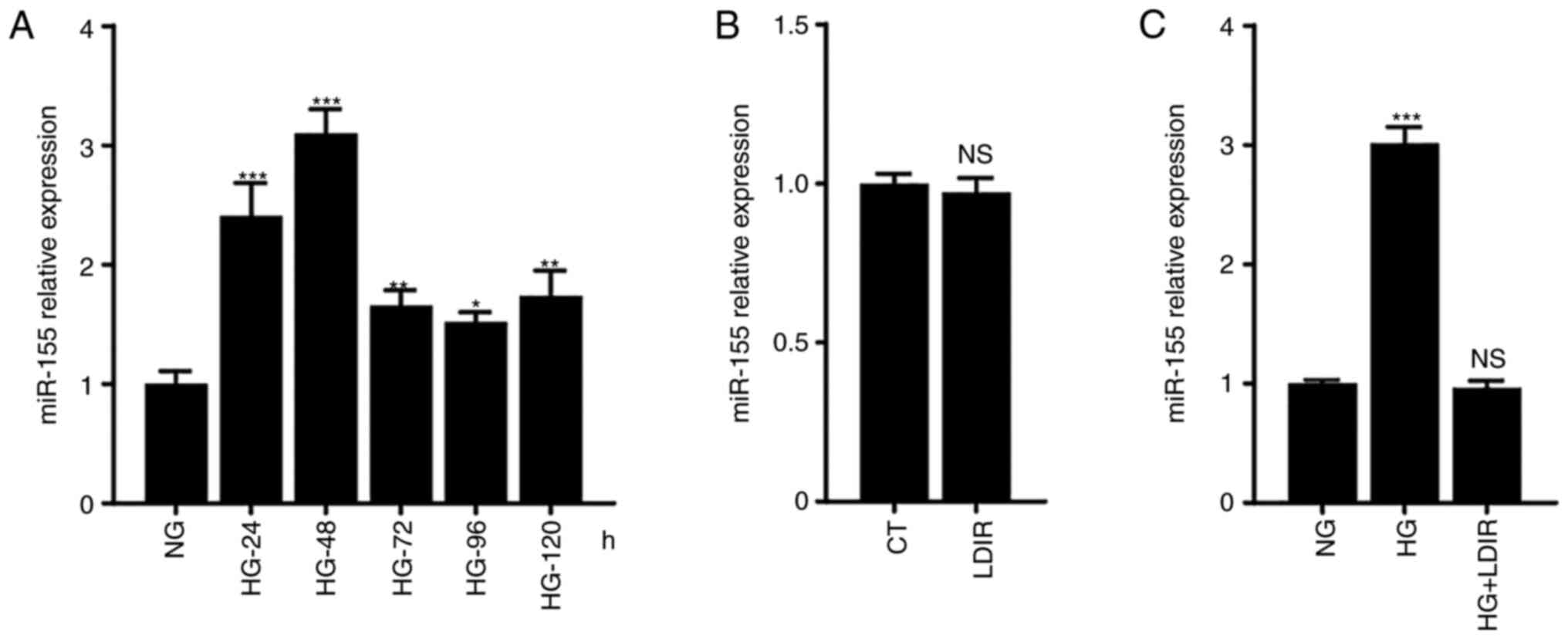 | Figure 1.miR-155 expression levels following
the treatment with HG and/or LDIR. AML12 cells were treated with HG
for 24, 48, 72, 96, or 120 h, with 75 mGy LDIR, or with HG for 48 h
followed by LDIR, after which the miR-155 expression levels were
determined. (A) HG treatment increased the expression levels of
miR-155, which peak at 48 h post-HG treatment. (B) LDIR alone did
not affect the expression levels of miR-155. (C) LDIR suppressed
the HG-induced miR-155 upregulation. *P<0.05, **P<0.01,
***P<0.001. NS, not significant; HG, high glucose; NG, normal
glucose; LDIR, low-dose ionizing radiation; miR, microRNA. |
To determine whether LDIR affected the HG-induced
miR-155 upregulation, AML12 cells were irradiated with 75 mGy
X-ray, and it was found that single LDIR treatment did not affect
the levels of miR-155 in AML12 cells (Fig. 1B). Next, the cells were treated
with HG for 48 h, after which they were irradiated with 75 mGy
X-ray, and the expression of miR-155 was determined. Interestingly,
it was found that LDIR significantly suppressed the HG-induced
miR-155 upregulation in AML-12 cells (Fig. 1C).
LDIR alleviates HG-induced AML12 cell
apoptosis
Diabetes may induce apoptosis in multiple types of
cells, including podocytes, cardiomyocytes, Schwann cells, and
hepatocytes (32–35). In the present study, AML12 cell
apoptosis following HG treatment was assessed. After AML12 cells
were cultured in medium containing 30 mM glucose for 48 h, cell
apoptosis was determined by flow cytometry. As shown in Fig. 2A, compared with the NG group, HG
treatment significantly induced cell apoptosis (42.6 vs. 0.88%,
P<0.001). Next, whether LDIR could alleviate the HG-induced cell
apoptosis was assessed. AML12 cells treated with HG for 48 h were
irradiated with 75 mGy X-ray. The flow cytometry results showed
that LDIR did not induce cell apoptosis, but significantly
decreased the HG-induced cell apoptosis (Fig. 2A, 42.6 vs. 13.84%, P<0.001).
Transfection of miR-155 mimics induced apoptosis in AML12 cells and
LDIR decreased the miR-155-induced cell apoptosis (Fig. 2B, 25.09 vs. 14.24%, P<0.001).
Moreover, it was found that miR-155 inhibitor decreased the
HG-induced cell apoptosis (Fig.
2C, 38.68 vs. 13.11%, P<0.001).
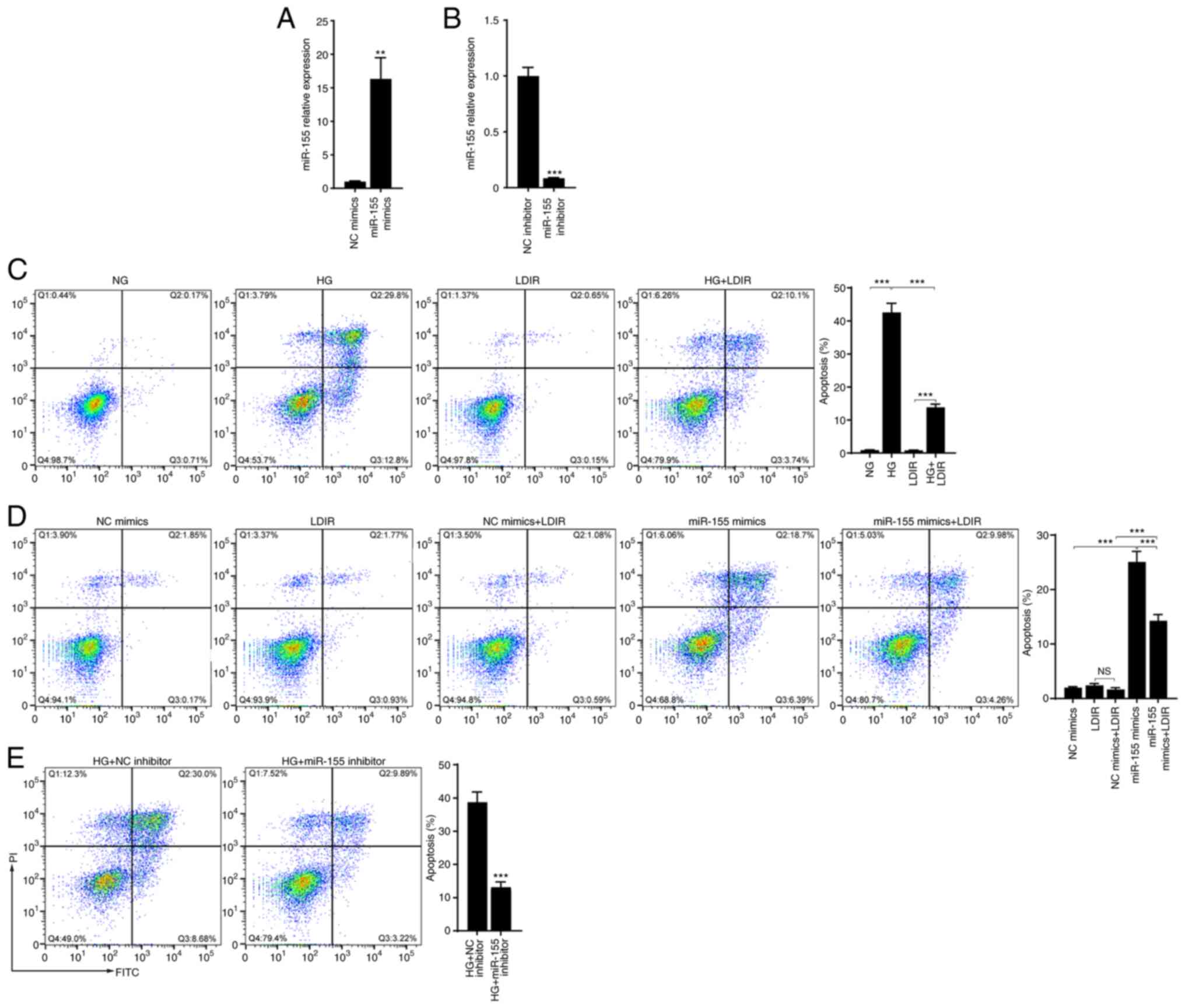 | Figure 2.LDIR attenuated HG or miR-155-induced
cell apoptosis. AML12 cells were treated with HG for 48 h, with 75
mGy LDIR, or transfected with miR-155 mimics or miR-155 inhibitor,
and cell apoptosis was determined by flow cytometry. (A and B)
Relative miR-155 expression levels following transfection with
miR-155 mimics and miR-155 inhibitor. (C) LDIR significantly
attenuated the HG-induced cell apoptosis. (D) Transfection of
miR-155 mimics induced cell apoptosis, which was attenuated by
LDIR. (E) Transfection of miR-155 attenuated the HG-induced cell
apoptosis. **P<0.01, ***P<0.001. NS, not significant; HG,
high glucose; LDIR, low-dose ionizing radiation; miR, microRNA; NC,
negative control. |
LDIR attenuates HG- or miR-155-induced
SOCS1 suppression
SOCS1 has been reported to be a regulatory target
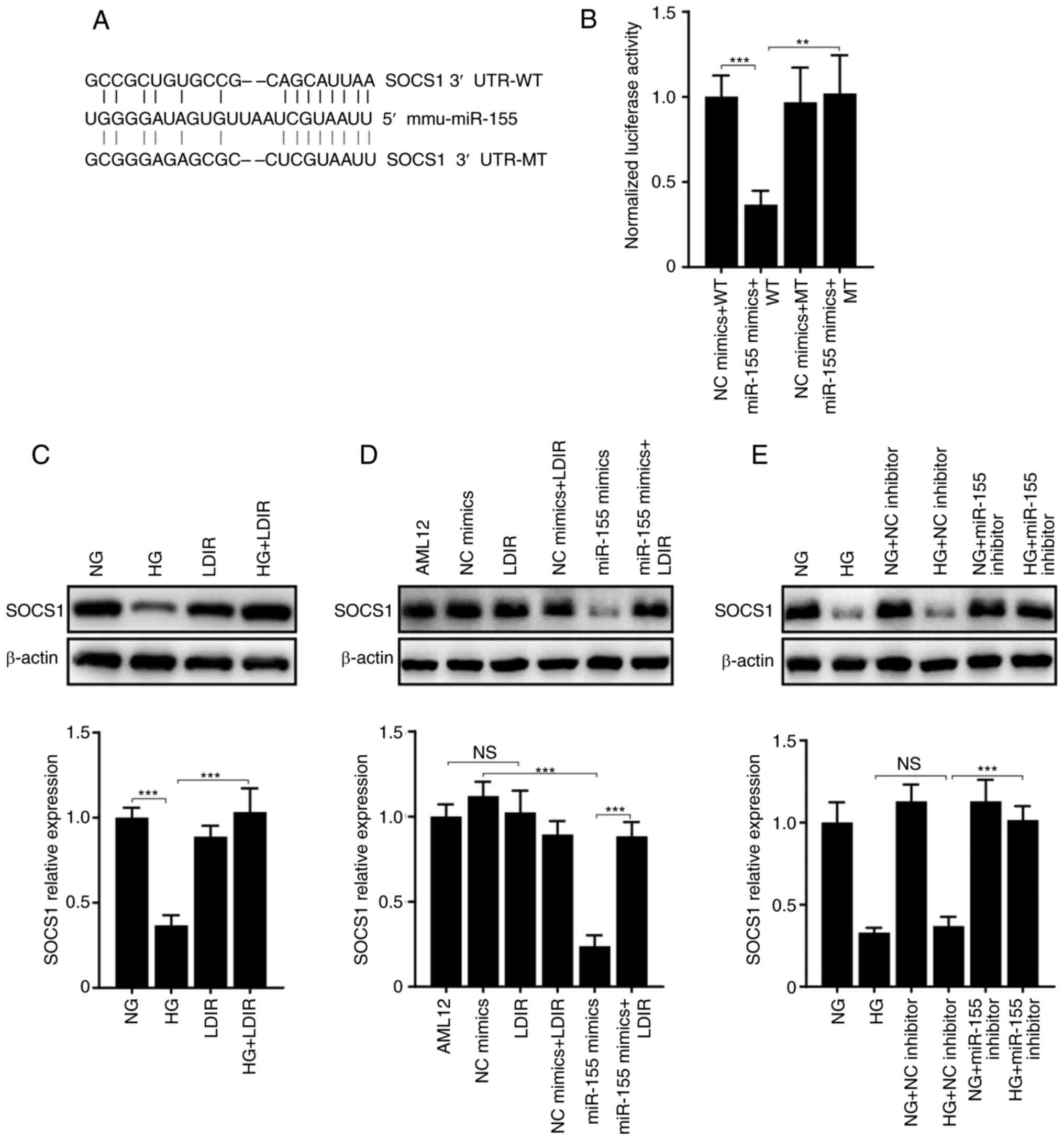
protein of miR-155 (36); Fig. 3A shows the predicted miR-155
binding site on the 3′ UTR of SOCS1. Next, luciferase reporter
assays were performed to determine the relation between miR-155 and
SOCS1. Luciferase reporter plasmids including WT or MT SOCS1 3′-UTR
were constructed (Fig. 3A). As
shown in Fig. 3B, the normalized
luciferase activity of the WT SOCS1 3′-UTR was significantly
reduced when co-transfected with miR-155 mimics. However, the
luciferase activity of the MT 3′-UTR was not affected. These
outcomes suggest SOCS1 is a direct target of miR-155 in AML12
cells.
Since it was demonstrated that miR-155 inhibitor may
disturb HG-induced cell apoptosis, it was speculated that LDIR may
alleviate HG-induced cell apoptosis through a miR-155-SOCS1 axis.
First, the expression levels of SOCS1 in AML12 cells treated with
HG and LDIR were determined. The western blotting results showed
that HG significantly suppressed the expression of SOCS1, whereas
LDIR attenuated this process (Fig.
3C, P<0.001). As a target of miR-155, the expression levels
of SOCS1 were suppressed by the transfection of miR-155 mimics.
However, LDIR disturbed the communication between miR-155 and
SOCS1, and this resulted in the recovery of SOCS1 expression levels
(Fig. 3D). Similarly, miR-155
inhibitor may have also disturbed the suppression of SOCS1 that was
induced by HG (Fig. 3E).
LDIR suppresses the HG-induced release
of inflammatory factors
It has been reported that HG treatment may
significantly elevate the expression levels of inflammatory factors
in endothelial cells (37,38). In the present study, the effects of
LDIR on HG-induced inflammation in hepatocytes were detected using
ELISA kits. The levels of TNF-α, IL-1β, IL-6, and IL-10 were
significantly elevated by HG treatment (Fig. 4A; P<0.001, P<0.001,
P<0.001, and P<0.05, respectively). However, the levels of
IFN-γ were not affected by HG treatment (P>0.05). LDIR reduced
the levels of TNF-α, IL-1β, IL-6, and IL-10, compared with cells
treated with HG alone (Fig. 4A,
P<0.001, P<0.01, P<0.01, and P<0.05, respectively).
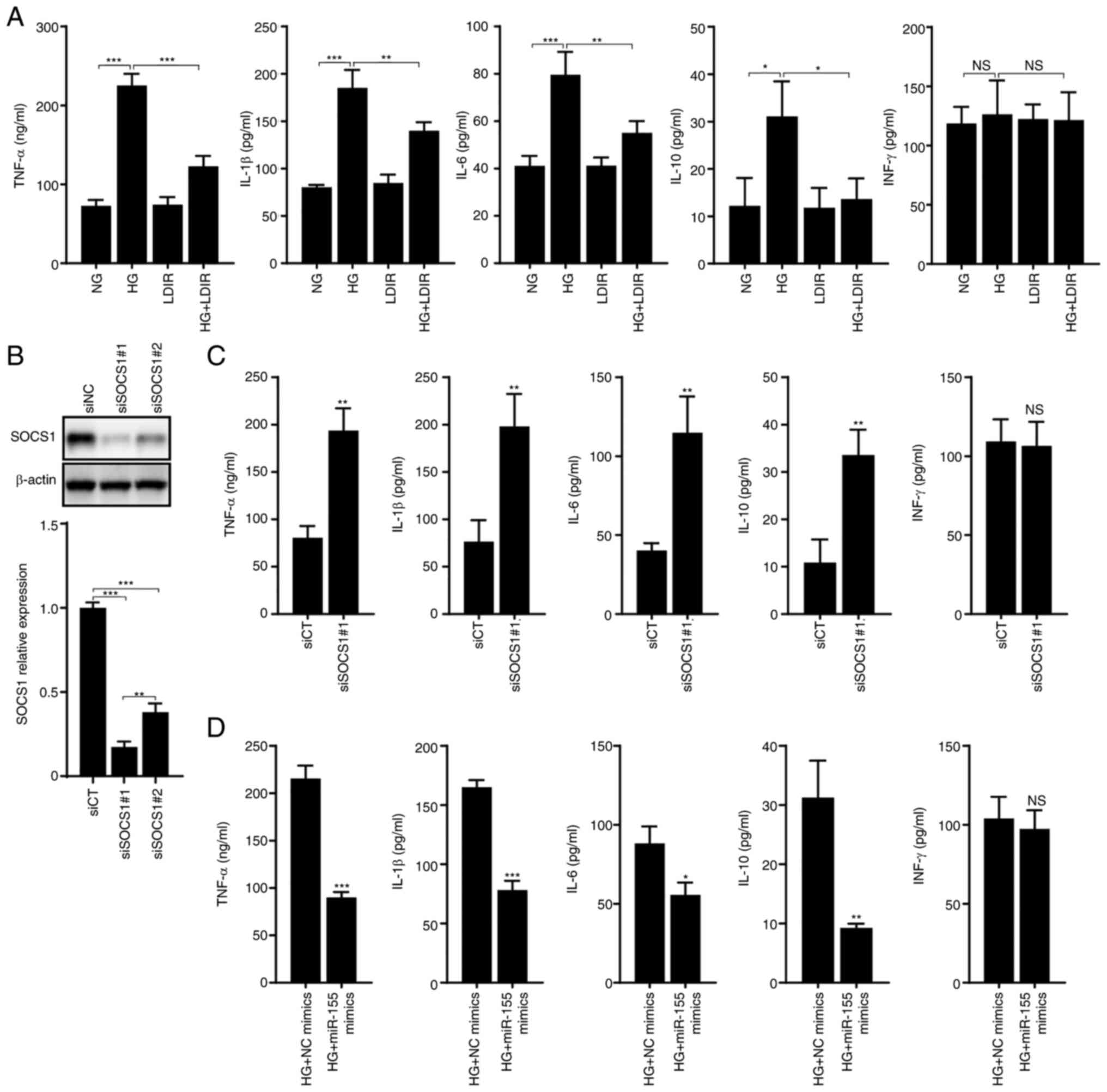 | Figure 4.LDIR attenuated the HG-induced
release of inflammatory factors through regulation of a
miR-155-SOCS1 axis. AML12 cells were treated by HG for 48 h, 75 mGy
LDIR, transfected with siSOCS1, or transfected with miR-155 mimics,
after which, the release of five inflammatory factors, including
TNF-α, IL-1β, IL-6, IL-10, and IFN-γ was detected by ELISA. (A)
LDIR suppresses the HG-induced release of TNF-α, IL-1β, IL-6, and
IL-10, but did not affect the release of IFN-γ. (B) SOCS1
expression was knocked down by transfection of siSOCS1 in AML12
cells. (C) Knockdown of SOCS1 expression upregulated the release of
TNF-α, IL-1β, IL-6, and IL-10. (D) Transfection of miR-155 mimics
suppressed the HG-induced release of TNF-α, IL-1β, IL-6, and IL-10.
*P<0.05, **P<0.01, ***P<0.001. HG, high glucose; NG,
normal glucose; zLDIR, low-dose ionizing radiation; miR, microRNA;
SOCS1, suppressor of cytokine signaling 1; si, small interfering;
CT, control. |
SOCS1 has been reported to negatively regulate the
expression of several inflammatory factors (39,40).
In the present study, two siRNA sequences were used to knockdown
SOCS1 expression, independently, and siSOCS1#1, which exhibited the
better knockdown efficiency, was used for subsequent experiments
(Fig. 4B). The release of
inflammatory factors in SOCS1 knockdown cells was measured, and it
was found that the levels of TNF-α, IL-1β, IL-6, and IL-10 were
significantly increased (Fig. 4C,
P<0.01). Since miR-155 is a negative regulator of SOCS1, here,
it was demonstrated that in HG-treated AML12 cells, the
overexpression of miR-155 mimics significantly reduced the levels
of TNF-α, IL-1β, IL-6, and IL-10 (Fig.
4D, P<0.001, P<0.001, P<0.05, and P<0.01,
respectively).
Discussion
The liver is the largest gland in the body and is
pivotal for substance metabolism. The liver is one of the organs
that is involved in diabetes injury. In patients with T2DM, the
incidence of non-alcoholic fatty liver disease ranges from 50–70%
(41), and approximately 20–30% of
these patients show abnormal liver function (42). In addition, the rate of cirrhosis
in diabetics is 1–3× higher than that in healthy individuals
(42). Several drugs, such as
liraglutide, exenatide, and lixisenatide, have been synthesized
with the aim of reducing triacylglycerol, total cholesterol, and
low-density lipoprotein cholesterol levels in the blood of T2DM
patients. These drugs have also been shown to exert a protective
effect against T2DM-induced hepatic steatosis and liver damage by
inhibiting oxidative stress and various inflammatory responses
(43–45). Compared with the development of
drugs, there are no studies that have focused on the treatment of
DLI patients using physical methods, to the best of our knowledge.
Several studies have tried to use LDIR to treat diabetes, where it
has been shown to exert a favorable effect in animal experiments
(21,22,46);
however, using LDIR to treat DLI has not been previously reported
on, to the best of our knowledge.
Epidemiological, preclinical, and clinical studies
highlight the beneficial effects of LDIR in healthy and diseased
conditions (47,48). Several studies have reported the
hormetic effects of LDIR, which increases the longevity of
organisms (49), reduces tumor
metastasis and mortalities (50,51),
improves neuronal function (52),
improves the condition of patients with T2DM (53), and even improves the health of
patients with COVID-19 (54).
Takehara et al (21)
reported that in a diabetic mouse model, pre-treatment with LDIR
inhibited alloxan-induced diabetes and delayed the onset of
hyperglycemia by elevating antioxidant levels and protecting
pancreatic cells. Zhang et al (46) reported that in a diabetic
nephropathy (DN) mouse model, treatment with LDIR cured the DN by
upregulating the expression of the renal antioxidant enzyme
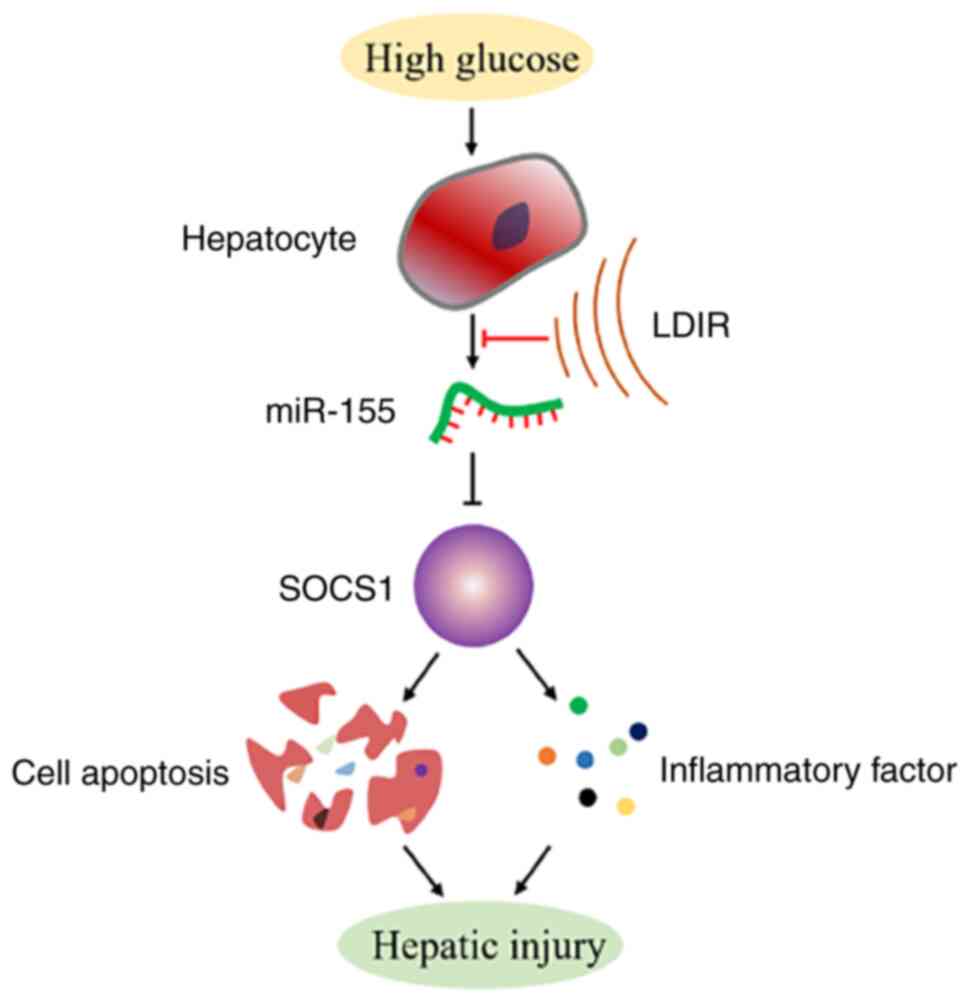
superoxide dismutase-1 (SOD-1). In the present study, it was first
demonstrated that LDIR may protect against DLI through suppression
of a miR-155/SOCS1 axis. HG may induce hepatic apoptosis by
upregulating the levels of miR-155 and downregulating the levels of
SOCS1 protein. HG also stimulated the secretion of TNF-α, IL-1β,
IL-6, and IL-10. However, LDIR blocked the HG-induced activation of
miR-155/SOCS1 axis and suppressed the release of inflammatory
factors (Fig. 5).
miR-155 exerts different biological functions in
different cell types and disease models. In certain tumors, it acts
as an oncogene to promote cell proliferation, invasion, and
metastasis (55–58), whereas, in other tumors, it acts as
a tumor suppressor to inhibit cell proliferation and promote
apoptosis (59,60). Li et al (61) reported that triptolide could
significantly induce the expression of miR-155 both in normal human
hepatocytes and in rodent liver tissues, and inhibition of miR-155
mitigated the hepatic damage caused by triptolide. According to the
results of the present study, HG-induced miR-155 could also damage
AML12 hepatocytes, and inhibition of miR-155 mitigated this damage,
consistent with the findings of Li et al (61).
Additionally, miR-155 is an important regulator of
inflammation and immunity. Preclinical studies and clinical trials
have indicated that miR-155 levels are altered in several types of
liver disease, such as alcoholic liver injury, infectious liver
injury, and viral hepatitis (62).
Sarkar et al (63) reported
that miR-155 expression was downregulated in HBV-infected serum
samples and liver biopsy; Bala et al (64) demonstrated that chronic alcohol
treatment induced a time-dependent increase in the levels of
miR-155 in macrophages in vivo and in vitro; and Dai
et al (65) demonstrated
that the expression of miR-155 was decreased in serum and liver
tissue samples from patients with cirrhosis. SOCS is an important
inhibitor of cytokine signaling pathways and also a key
physiological regulator of natural and acquired immunity systems
(66). SOCS1 is one of the most
frequent modulatory targets of miR-155. The miR-155/SOCS1 axis,
which is known to inhibit the JAK-STAT pathway, plays a critical
role in the regulation of cell proliferation, inflammatory
responses, and viral replication (15,36,67,68).
The miR-155/SOCS1 axis is also reported to play a role in diabetes.
Lin et al (69) reported
that the levels of miR-155 are lower in T2DM patients. They
indicated that miR-155 positively modulated glucose uptake through
coordination with SOCS1, and overexpression of miR-155 in
transgenic mice resulted in hypoglycemia, and improved glucose
tolerance and insulin sensitivity. Several transcription factors,
including NF-κB, AP-1, and STAT3, have been identified as
positive regulators of miR-155; however, the exact signaling
pathways remain unclear (18).
In conclusion, in the present study, it was shown
that LDIR may exert a hepatoprotective effect by regulating a
miR-155-SOCS1 axis. Pretreatment with LDIR may abrogate the
HG-induced activation of the miR-155-SOCS1 axis and suppress the
release of inflammatory factors. Considering that there are many
diabetic patients in China and there is still a lack of effective
preventative measures for diabetic liver disease, the present study
provides a potential therapeutic strategy for the treatment of DLI.
However, due to the lack of direct human studies, the therapeutic
efficacy of LDIR remains contested. Moreover, the present study was
a preliminary in vitro study, and to obtain a more accurate
conclusion, further in vivo experiments are still
required.
Acknowledgements
Not applicable.
Funding
The present study was supported in part by grants from the
Science and Technology Department of Jilin Province (grant no.
20200201309JC) and the National Natural Science Foundation of China
(grant no. 81903250).
Availability of data and materials
The datasets used and analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XL conceived and designed the study. HF, SL, and BJ
acquired, analyzed, and interpreted the data. HF drafted the
manuscript. All authors read and approved the final manuscript. HF
and XL confirm the authenticity of all the raw data.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Wang J, Wu W, Dong G, Huang K and Fu J:
Pediatric diabetes in China: Challenges and actions. Pediatr
Diabetes. 23:545–550. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bedi O, Aggarwal S, Trehanpati N,
Ramakrishna G and Krishan P: Molecular and pathological events
involved in the pathogenesis of diabetes-associated nonalcoholic
fatty liver disease. J Clin Exp Hepatol. 9:607–618. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Assuncao SNF, Sorte NCB, Alves CD, Mendes
PSA, Alves CRB and Silva LR: Nonalcoholic fatty liver disease
(NAFLD) pathophysiology in obese children and adolescents: Update.
Nutr Hosp. 34:727–730. 2017. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Farhood B, Goradel NH, Mortezaee K,
Khanlarkhani N, Najafi M and Sahebkar A: Melatonin and cancer: From
the promotion of genomic stability to use in cancer treatment. J
Cell Physiol. 234:5613–5627. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Mettler FA, Sinclair WK, Anspaugh L,
Edington C, Harley JH, Ricks RC, Selby PB, Webster EW and Wyckoff
HO: The 1986 and 1988 UNSCEAR (United Nations Scientific Committee
on the Effects of Atomic Radiation) reports: Findings and
implications. Health Phys. 58:241–250. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Luckey TD: Physiological benefits from low
levels of ionizing radiation. Health Phys. 43:771–789. 1982.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Feinendegen LE: Evidence for beneficial
low level radiation effects and radiation hormesis. Br J Radiol.
78:3–7. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Olivieri G, Bodycote J and Wolff S:
Adaptive response of human lymphocytes to low concentrations of
radioactive thymidine. Science. 223:594–597. 1984. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Averbeck D, Salomaa S, Bouffler S,
Ottolenghi A, Smyth V and Sabatier L: Progress in low dose health
risk research: Novel effects and new concepts in low dose
radiobiology. Mutat Res Rev Mutat Res. 776:46–69. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Jiang H, Xu Y, Li W, Ma K, Cai L and Wang
G: Low-dose radiation does not induce proliferation in tumor cells
in vitro and in vivo. Radiat Res. 170:477–487. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bedewy AML, Elmaghraby SM, Shehata AA and
Kandil NS: Prognostic value of miRNA-155 expression in B-cell
Non-Hodgkin Lymphoma. Turk J Haematol. 34:207–212. 2017.PubMed/NCBI
|
|
12
|
Bayraktar R and Van Roosbroeck K: miR-155
in cancer drug resistance and as target for miRNA-based
therapeutics. Cancer Metastasis Rev. 37:33–44. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Mashima R: Physiological roles of miR-155.
Immunology. 145:323–333. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Bala S, Csak T, Saha B, Zatsiorsky J,
Kodys K, Catalano D, Satishchandran A and Szabo G: The
pro-inflammatory effects of miR-155 promote liver fibrosis and
alcohol-induced steatohepatitis. J Hepatol. 64:1378–1387. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chen L, Ming X, Li W, Bi M, Yan B, Wang X,
Yang P and Yang B: The microRNA-155 mediates hepatitis B virus
replication by reinforcing SOCS1 signalling-induced autophagy. Cell
Biochem Funct. 38:436–442. 2020. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Babuta M, Furi I, Bala S, Bukong TN, Lowe
P, Catalano D, Calenda C, Kodys K and Szabo G: Dysregulated
autophagy and lysosome function are linked to exosome production by
Micro-RNA 155 in alcoholic liver disease. Hepatology. 70:2123–2141.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang J and Che J: CircTP63 promotes
hepatocellular carcinoma progression by sponging miR-155-5p and
upregulating ZBTB18. Cancer Cell Int. 21:1562021. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Jankauskas SS, Gambardella J, Sardu C,
Lombardi A and Santulli G: Functional role of miR-155 in the
pathogenesis of diabetes mellitus and its complications. Noncoding
RNA. 7:392021.PubMed/NCBI
|
|
19
|
El Samaloty NM, Hassan ZA, Hefny ZM and
Abdelaziz DHA: Circulating microRNA-155 is associated with insulin
resistance in chronic hepatitis C patients. Arab J Gastroenterol.
20:1–7. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Bai X, Luo Q, Tan K and Guo L: Diagnostic
value of VDBP and miR-155-5p in diabetic nephropathy and the
correlation with urinary microalbumin. Exp Ther Med. 20:862020.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Takehara Y, Yamaoka K, Hiraki Y, Yoshioka
T and Utsumi K: Protection against alloxan diabetes by low-dose
60Co gamma irradiation before alloxan administration. Physiol Chem
Phys Med NMR. 27:149–159. 1995.PubMed/NCBI
|
|
22
|
Zhang C, Tan Y, Guo W, Li C, Ji S, Li X
and Cai L: Attenuation of diabetes-induced renal dysfunction by
multiple exposures to low-dose radiation is associated with the
suppression of systemic and renal inflammation. Am J Physiol
Endocrinol Metab. 297:E1366–E1377. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhao Y, Kong C, Chen X, Wang Z, Wan Z, Jia
L, Liu Q, Wang Y, Li W, Cui J, et al: Repetitive exposure to
low-dose X-irradiation attenuates testicular apoptosis in type 2
diabetic rats, likely via Akt-mediated Nrf2 activation. Mol Cell
Endocrinol. 422:203–210. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang Y, Li Y, Zhao J, Wang C, Deng B,
Zhang Q and Shi C: Protective effects and mechanisms of
Polyethylene Glycol Loxenatide Against Hyperglycemia and liver
injury in db/db diabetic mice. Front Pharmacol. 12:7818562021.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lizotte F, Denhez B, Guay A, Gevry N, Cote
AM and Geraldes P: Persistent insulin resistance in podocytes
caused by epigenetic changes of SHP-1 in diabetes. Diabetes.
65:3705–3717. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Shen L, Evel-Kabler K, Strube R and Chen
SY: Silencing of SOCS1 enhances antigen presentation by dendritic
cells and antigen-specific anti-tumor immunity. Nat Biotechnol.
22:1546–1553. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
27
|
Shi D, Li D, Yin Q, Qiu Y, Yan H, Shen Y,
Lu G and Liu W: Silenced suppressor of cytokine signaling 1 (SOCS1)
enhances the maturation and antifungal immunity of dendritic cells
in response to Candida albicans in vitro. Immunol Res. 61:206–218.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Polina ER, Oliveira FM, Sbruzzi RC,
Crispim D, Canani LH and Santos KG: Gene polymorphism and plasma
levels of miR-155 in diabetic retinopathy. Endocr Connect.
8:1591–1599. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Garcia-Diaz DF, Pizarro C, Camacho-Guillen
P, Codner E, Soto N and Perez-Bravo F: Expression of miR-155,
miR-146a, and miR-326 in T1D patients from Chile: Relationship with
autoimmunity and inflammatory markers. Arch Endocrinol Metab.
62:34–40. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Mostahfezian M, Azhir Z, Dehghanian F and
Hojati Z: Expression pattern of microRNAs, miR-21, miR-155 and
miR-338 in patients with type 1 diabetes. Arch Med Res. 50:79–85.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chen X, Wang J, Lin Y, Liu Y and Zhou T:
Signaling pathways of podocyte injury in diabetic kidney disease
and the effect of sodium-glucose cotransporter 2 inhibitors. Cells.
11:39132022. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ouyang C, You J and Xie Z: The interplay
between autophagy and apoptosis in the diabetic heart. J Mol Cell
Cardiol. 71:71–80. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Liu YP, Shao SJ and Guo HD: Schwann cells
apoptosis is induced by high glucose in diabetic peripheral
neuropathy. Life Sci. 248:1174592020. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Schattenberg JM and Schuchmann M: Diabetes
and apoptosis: Liver. Apoptosis. 14:1459–1471. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Lin X, Chen L, Li H, Liu Y, Guan Y, Li X,
Jia Z, Lin X, Jia J, Sun Y and Xiao D: miR-155 accelerates
proliferation of mouse hepatocytes during liver regeneration by
directly targeting SOCS1. Am J Physiol Gastrointest Liver Physiol.
315:G443–G453. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhou X, Wang L, Zhang Z, Liu J, Qu Q, Zu Y
and Shi D: Fluorometholone inhibits high glucose-induced cellular
senescence in human retinal endothelial cells. Hum Exp Toxicol.
41:96032712210761072022. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Li B, Li H, Dai L, Liu C, Wang L, Li Q and
Gu C: NIK-SIX1 signalling axis regulates high glucose-induced
endothelial cell dysfunction and inflammation. Autoimmunity.
55:86–94. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
He Y, Zhang W, Zhang R, Zhang H and Min W:
SOCS1 inhibits tumor necrosis factor-induced activation of ASK1-JNK
inflammatory signaling by mediating ASK1 degradation. J Biol Chem.
281:5559–5566. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wu XY, Yu J and Tian HM: Effect of SOCS1
on diabetic renal injury through regulating TLR signaling pathway.
Eur Rev Med Pharmacol Sci. 23:8068–8074. 2019.PubMed/NCBI
|
|
41
|
Lee YH, Cho Y, Lee BW, Park CY, Lee DH,
Cha BS and Rhee EJ: Nonalcoholic fatty liver disease in diabetes.
Part I: Epidemiology and diagnosis. Diabetes Metab J. 43:31–45.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Cusi K, Sanyal AJ, Zhang S, Hartman ML,
Bue-Valleskey JM, Hoogwerf BJ and Haupt A: Non-alcoholic fatty
liver disease (NAFLD) prevalence and its metabolic associations in
patients with type 1 diabetes and type 2 diabetes. Diabetes Obes
Metab. 19:1630–1634. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Voukali M, Kastrinelli I, Stragalinou S,
Tasiopoulou D, Paraskevopoulou P, Katsilambros N, Kokkinos A,
Tentolouris N and Ioannidis I: Study of postprandial lipaemia in
type 2 diabetes mellitus: Exenatide versus liraglutide. J Diabetes
Res. 2014:3040322014. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Sun F, Wu S, Wang J, Guo S, Chai S, Yang
Z, Li L, Zhang Y, Ji L and Zhan S: Effect of glucagon-like
peptide-1 receptor agonists on lipid profiles among type 2
diabetes: A systematic review and network meta-analysis. Clin Ther.
37:225–241. e2282015. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Roca-Rodriguez MM, Muros de Fuentes MT,
Piedrola-Maroto G, Quesada-Charneco M, Maraver-Selfa S, Tinahones
FJ and Mancha-Doblas I: Lixisenatide in patients with type 2
diabetes and obesity: Beyond glycaemic control. Aten Primaria.
49:294–299. 2017.(In Spanish). PubMed/NCBI
|
|
46
|
Zhang C, Xing X, Zhang F, Shao M, Jin S,
Yang H, Wang G, Cui J, Cai L, Li W and Lu X: Low-dose radiation
induces renal SOD1 expression and activity in type 1 diabetic mice.
Int J Radiat Biol. 90:224–230. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Shibamoto Y and Nakamura H: Overview of
biological, epidemiological, and clinical evidence of radiation
hormesis. Int J Mol Sci. 19:23872018. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Scott BR: Radiation-hormesis phenotypes,
the related mechanisms and implications for disease prevention and
therapy. J Cell Commun Signal. 8:341–352. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Lopez-Martinez G and Hahn DA: Early life
hormetic treatments decrease irradiation-induced oxidative damage,
increase longevity, and enhance sexual performance during old age
in the Caribbean fruit fly. PLoS One. 9:e881282014. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Cheda A, Wrembel-Wargocka J, Lisiak E,
Nowosielska EM, Marciniak M and Janiak MK: Single low doses of X
rays inhibit the development of experimental tumor metastases and
trigger the activities of NK cells in mice. Radiat Res.
161:335–340. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
51
|
Doss M: Evidence supporting radiation
hormesis in atomic bomb survivor cancer mortality data. Dose
Response. 10:584–592. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Cuttler JM, Moore ER, Hosfeld VD and
Nadolski DL: Update on a patient with Alzheimer disease treated
with CT scans. Dose Response. 15:15593258176931672017. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Kojima S, Cuttler JM, Shimura N, Koga H,
Murata A and Kawashima A: Radon therapy for autoimmune diseases
pemphigus and diabetes: 2 case reports. Dose Response.
17:15593258198509842019. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Dhawan G, Kapoor R, Dhawan R, Singh R,
Monga B, Giordano J and Calabrese EJ: Low dose radiation therapy as
a potential life saving treatment for COVID-19-induced acute
respiratory distress syndrome (ARDS). Radiother Oncol. 147:212–216.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Qu YL, Wang HF, Sun ZQ, Tang Y, Han XN, Yu
XB and Liu K: Up-regulated miR-155-5p promotes cell proliferation,
invasion and metastasis in colorectal carcinoma. Int J Clin Exp
Pathol. 8:6988–6994. 2015.PubMed/NCBI
|
|
56
|
Bhattacharya S, Chalk AM, Ng AJ, Martin
TJ, Zannettino AC, Purton LE, Lu J, Baker EK and Walkley CR:
Increased miR-155-5p and reduced miR-148a-3p contribute to the
suppression of osteosarcoma cell death. Oncogene. 35:5282–5294.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
McDonald SJ, Cranford TL, VanderVeen BN,
Cardaci TD, Velázquez KT, Enos RT, Chatzistamou I, Fan D and Murphy
EA: miR155 deficiency reduces breast tumor burden in the MMTV-PyMT
mouse model. Physiol Genomics. 54:433–442. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Su N, Li L, Zhou E, Li H, Wu S and Cao Z:
Resveratrol downregulates miR-155-5p to block the malignant
behavior of gastric cancer cells. Biomed Res Int. 2022:69686412022.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Li S, Zhang T, Zhou X, Du Z, Chen F, Luo J
and Liu Q: The tumor suppressor role of miR-155-5p in gastric
cancer. Oncol Lett. 16:2709–2714. 2018.PubMed/NCBI
|
|
60
|
Yao LY, Ma J, Zeng XM and Ou-Yang J:
MicroRNA-155-5p inhibits the invasion and migration of prostate
cancer cells by targeting SPOCK1. Oncol Lett. 20:3532020.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Li Y, Guo L, Hou Z, Gong H, Yan M and
Zhang B: Role of MicroRNA-155 in Triptolide-induced hepatotoxicity
via the Nrf2-dependent pathway. J Ethnopharmacol. 281:1144892021.
View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Xue X, Wang J, Fu K, Dai S, Wu R, Peng C
and Li Y: The role of miR-155 on liver diseases by modulating
immunity, inflammation and tumorigenesis. Int Immunopharmacol.
116:1097752023. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Sarkar N, Panigrahi R, Pal A, Biswas A,
Singh SP, Kar SK, Bandopadhyay M, Das D, Saha D, Kanda T, et al:
Expression of microRNA-155 correlates positively with the
expression of Toll-like receptor 7 and modulates hepatitis B virus
via C/EBP-β in hepatocytes. J Viral Hepat. 22:817–827. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Bala S and Szabo G: MicroRNA signature in
alcoholic liver disease. Int J Hepatol. 2012:4982322012. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Dai W, Zhao J, Tang N, Zeng X, Wu K, Ye C,
Shi J, Lu C, Ning B, Zhang J and Lin Y: MicroRNA-155 attenuates
activation of hepatic stellate cell by simultaneously preventing
EMT process and ERK1 signalling pathway. Liver Int. 35:1234–1243.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Liang YB, Tang H, Chen ZB, Zeng LJ, Wu JG,
Yang W, Li ZY and Ma ZF: Downregulated SOCS1 expression activates
the JAK1/STAT1 pathway and promotes polarization of macrophages
into M1 type. Mol Med Rep. 16:6405–6411. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Tan L, Jiang W, Lu A, Cai H and Kong L:
miR-155 aggravates liver Ischemia/reperfusion injury by suppressing
SOCS1 in mice. Transplant Proc. 50:3831–3839. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Ren Y, Cui Y, Xiong X, Wang C and Zhang Y:
Inhibition of microRNA-155 alleviates lipopolysaccharide-induced
kidney injury in mice. Int J Clin Exp Pathol. 10:9362–9371.
2017.PubMed/NCBI
|
|
69
|
Lin X, Qin Y, Jia J, Lin T, Lin X, Chen L,
Zeng H, Han Y, Wu L, Huang S, et al: MiR-155 enhances insulin
sensitivity by coordinated regulation of multiple genes in mice.
PLoS Genet. 12:e10063082016. View Article : Google Scholar : PubMed/NCBI
|