Introduction
Diabetes mellitus (DM), one of the major risk
factors for the initiation and progression of nephropathy, can
cause kidney damage by inducing the production of reactive oxygen
metabolites and attenuating antioxidative mechanisms (1). Diabetic nephropathy (DN) is
characterized by both ultrastructural and morphological
alternations in the kidney, including glomerular mesangial
expansion, basement membrane thickening and tubular hypertrophy
(2). The pathogenesis of DN is
complex and oxidative stress is widely considered to contribute to
its progression (3), as excessive
reactive oxygen species (ROS) production, including mitochondrial
ROS, causes oxidative stress, and can ultimately lead to
mitochondrial damage and cellular death (4). Therefore, the effective prevention
and treatment of DN is important.
An increasing body of evidence has suggested that
autophagy is a potential therapeutic target for DN (5,6).
Autophagy is an evolutionarily conserved biological process in
cells that degrades damaged organelles and excess proteins to
maintain normal balance in response to stress and injury (7). In this process, the unimpeded
autophagic flux is important for autophagy to degrade organelles
and proteins. Mitophagy is a selective type of autophagy, in which
impaired mitochondria are degraded by lysosomes. However,
dysfunction of autophagy can lead to certain diseases, including
Alzheimer's disease (8),
cardiovascular disease and inflammatory bowel disease (9). Cinacalcet-induced autophagy has
previously been reported to protect against kidney damage in a DN
model, acting through the CaMKKβ-LKB1-AMPK signaling pathway
(10). In addition, overexpression
of STAMP2 can attenuate renal injury by upregulating autophagy in
DN (11). Lian et al
(12) reported that inhibiting
autophagy can aggravate lead-induced kidney injury. These previous
studies suggested that autophagy serves an important role in
DN.
Melatonin (MLT; 5-methoxy-N-acetyltryptamine)
is secreted by the pineal gland and is a natural antioxidant that
inhibits oxidative stress by scavenging ROS (13). Once synthesized, MLT binds to two
specific G protein-coupled receptors: MLT receptor type 1 (MTI) and
MLT receptor type 2 (MT2) (14).
In diabetic kidney injury, MLT can mediate the anti-apoptotic
effect of MT2 and can also regulate mitochondrial fission/fusion
homeostasis, leading to protective effects (15,16).
In addition, MLT activates Wnt/β-catenin and TGF-β1-Smad2/3
signaling pathways to alleviate oxidative stress, which inhibits
TLR4 or NF-κB and TGF-β1/Smad3 signaling pathways to improve renal
inflammation and fibrosis (17–19).
It has also been reported that MLT serves an essential role in
anti-inflammatory processes, such as rheumatoid arthritis (20,21).
Yu et al (22) and Zhang
et al (23) previously
reported that MLT can augment mitochondrial function via modulating
AMPK activity and autophagy. Siddhi et al (24) also reported that MLT prevents DN by
modulating the AMPK/SIRT1 signal pathway via recovering the
autophagy and mitochondrial function. In addition, Qiu et al
(25) reported that MLT suppresses
ferroptosis via activation of the Nrf2/HO-1 signaling pathway in a
mouse model of sepsis-induced acute kidney injury. Therefore, the
protective mechanisms underlying the effects of MLT on DM-induced
kidney injury involve glucose homeostasis, oxidative stress,
inflammation and apoptosis. However, the pathogenesis of DM-induced
kidney injury is complex, and the mechanisms underlying the
protective effects of MLT against this disease are not yet fully
understood.
In the present study, the effects of MLT treatment
on fibrosis, oxidative damage and autophagy levels in the kidneys
of rats with DM were assessed.
Materials and methods
Experimental animal model
A total of 30 healthy clean-grade male 6-week-old
Sprague-Dawley (SD) rats (weight, 80±15 g) were purchased from
Liaoning Changsheng Biotechnology [animal license no. SCXK (Liao)
2020–0001]. The experimental procedures were approved by the Ethics
Committee of Yangzhou University [Yangzhou, China; approval no.
SYXK (Su) 2022–0044]. High-sugar and high-fat feed was purchased
from Jiangsu Synergy Pharmaceutical & Biological Engineering
Co., Ltd. Rats were maintained in a specific-pathogen-free housing
facility at a constant temperature of 25°C and 40–70% humidity
under a 12-h dark/light cycle, with free access to water and
standard clean-grade foods, and were acclimated for 1 week before
being fed high-sugar and high-fat feed for 4 weeks. A total of 20
rats were randomly divided into control and modeling groups after
being fed a high-fat, high-sugar diet for 5 weeks. The modeling
group was intraperitoneally injected with 35 mg/kg streptozotocin
[STZ; dissolved in 0.01 mol/l citrate buffer (pH 4.3)] after 12 h
of fasting without water to produce the DM rat model. The control
group of rats was intraperitoneally injected with an equal volume
of PBS. Fasting blood glucose (FBG) was measured using small drops
of blood collected from the tail for 3 consecutive days. Rats with
a stable FBG ≥16.7 mmol/l were considered a successful model of DM.
The total duration of the experiment was 17 weeks.
The health and behavior of rats were monitored daily
and no animals died prior to sacrifice. All rats were sacrificed by
cervical dislocation and death was verified by the cessation of
respiratory movements. Diethyl ether was used for anesthesia prior
to sacrifice. This method was approved by the Ethics Committee of
Yangzhou University [approval no. (SYXK (Su) 2022–0044)]. Briefly,
animals were placed in a plexiglass anesthesia box and a beaker
containing a cotton ball of diethyl ether was placed inside the
box. Respiratory rate was monitored to ensure that the animals were
fully anesthetized. For blood glucose (BG) detection, small drops
of blood were obtained from the tail using a lancet; the blood was
then placed onto BG test strips, which were placed into a BG meter
to obtain the BG value.
Detection of lipid metabolism
Blood lipids were detected using an automatic
biochemical analyzer (AU5800; Beckman Coulter, Inc.).
Serum preparation
Blood samples of rats were taken from the heart and
centrifuged at 1,000 × g at 4°C for 15 min. Serum was collected and
filtered through a 0.22-µm filter and stored at −80°C until
use.
Cell culture
The NRK-52E cell line was purchased from
MilliporeSigma. Cells were cultured in DMEM (Gibco; Thermo Fisher
Scientific, Inc.) supplemented with 10% fetal bovine serum (Gibco;
Thermo Fisher Scientific, Inc.) at 37°C in a humidified 5%
CO2 atmosphere. Cells were treated with 30 mM glucose
and 60 µM MLT at 37°C for 24 h.
Experimental groupings
The in vivo portion of the present study was
divided into three experimental groups: i) Control; ii) DM group;
and iii) DM + MLT group (n=10 per group). The concentration of 10
mg/l MLT in water was freely available every night. The in
vitro portion of the present study was also divided into three
experimental groups: i) Control; ii) high glucose (HG) group; and
iii) HG + MLT (60 µM) group.
Cell transfection
The plasmids were obtained from Professor Lin Wang
(Shandong Agricultural University, College of Veterinary Medicine,
Taian, Shangdong, China). After treatment with glucose and MLT, and
once NRK-52E cells reached 60–70% confluence, the RFP-GFP-LC3
plasmid was transfected into cells using Lipofectamine®
2000 transfection reagent (Invitrogen; Thermo Fisher Scientific,
Inc.) for the detection of autophagic flux. 200 µl transfection
system with 1 µg plasmid at 37°C for 24 h. The time interval
between transfection and subsequent experimentation was 24 h.
Subsequently the cells were treated with glucose and MLT. In the
present study, every group of cells was transiently transfected;
therefore, there were no negative or positive transfection
controls. When the autophagosome and lysosome did not fuse, both
RFP and GFP fluorescence were observed; however, when the
autophagosome and lysosome fused, resulting in GFP quenching, only
RFP fluorescence could be observed.
Reagents
The following reagents and materials were purchased
for use in the present study: STZ (MilliporeSigma); MLT (purity
>98%; MilliporeSigma); anhydrous glucose (Macklin, Inc.);
insulin (Novo Nordisk A/S); BG meter and test strips (Roche
Diagnostics); β-actin (cat. no. 93473SF) antibodies (Cell Signaling
Technology, Inc.); anti-collagen IV (COL-IV; cat. no. AF0510)
antibodies (Affinity Biosciences); anti-LC3 (cat. no. 4599T)
antibodies (Cell Signaling Technology, Inc.); anti-P62 (cat. no.
23214S) antibodies (Cell Signaling Technology, Inc.); ECL
Chemiluminescence Kit and BCA Protein Concentration Assay Kit
(Beyotime Institute of Biotechnology); and malondialdehyde (MDA,
cat. no. A003-1-2), superoxide dismutase (SOD, A001-3-2), catalase
(CAT, A007-1-1), glutathione peroxidase (GSH-Px, A005-1-2) and GSH
(A006-2-1) ELISA assay kits (Nanjing Jiancheng Bioengineering
Institute).
Oral glucose tolerance test (OGTT) and
intraperitoneal insulin tolerance test (IPITT)
OGTT was performed 1 week prior to euthanasia.
Animals were fasted for 12 h and were then orally administered 50%
glucose solution (2 g/kg). BG levels were determined from tail
blood samples at 0, 30, 60, 90 and 120 min. IPITT was performed 1
week prior to euthanasia. Animals were fasted for 4 h, after which,
1.5 U/kg insulin was injected intraperitoneally. BG was measured
from tail blood samples at 0, 30, 60, 90 and 120 min. As
aforementioned, BG was detected from small drops of blood collected
from the tail using a lancet; the blood was then placed onto BG
test strips, which were placed into a BG meter to obtain the BG
value. The area under the curve (AUC) was calculated as
follows=[0.5 × (BG at 0 min + BG at 30 min)/2] + [0.5 × (BG at 30
min + BG at 60 min)/2] + [1 × (BG at 60 min + BG at 120
min)/2].
Renal tissue oxidative damage index
test
Renal tissue was collected in 1.5 ml centrifuge
tube.
To prepare 10% tissue homogenate, kidney tissues
were placed in saline and centrifuged at 1,000 × g for 10 min at
4°C. The supernatant was collected to evaluate MDA, GSH-Px and GSH
levels and SOD and CAT activity levels by ELISA kits according to
manufacturer's protocols. Protein concentrations were measured
using a BCA assay. All results were normalized to protein
concentrations and expressed as U/mg protein or nmol/mg protein, as
appropriate.
Hematoxylin and eosin (H&E),
Masson's trichome and Periodic acid-Schiff (PAS) staining
Right kidney cortical tissues were fixed with 4%
formalin to prepare tissue sections at room temperature for 24 h
and the tissues embedded in paraffin. Sections (1×1×1 cm) were
dewaxed and stained with H&E (1% hematoxylin was stained for 5
min, dye with 0.5% eosin solution for 1–3 min), Masson's trichome
(Masson Ponceau acidic complexing liquid: Ponceau 0.7 g, acidic
complexing 0.3 g, distilled water 99 ml, glacial acetic acid 1 ml,
for 5–10 min) and PAS staining for 10 min at room temperature. The
morphological changes of kidney tissues in each group were observed
using a light microscope (Nikon Corporation).
Transmission electron microscopy
(TEM)
Fresh kidney tissues were cut into tissue blocks
~1×1×1 mm and were fixed with 2.5% glutaraldehyde at 4°C overnight.
Subsequently, the tissues were rinsed with PBS. NRK-52E cells were
collected in a 1.5-ml centrifuge tube and fixed with 2.5%
glutaraldehyde at 4°C for 24 h. Samples were dehydrated using an
ethanol gradient (45–95%), soaked and embedded in epoxy resin
overnight at room temperature, then ultra-thin sections (<100
nm) were cut. Uranium acetate was placed on the wax plate or on the
soaked filter paper, the carrier mesh with the sections placed in
the dye solution (cut side in contact with the stain) for 15–30
min, the sections were rinsed with double distilled water, blotted
with filter paper, and then the carrier mesh covered with 30 g/l
lead citrate stain (to prevent the formation of lead carbonate
precipitation, solid sodium hydroxide was placed in the petri
dish). After staining for 5–10 min, the sections were rinsed with
double steaming water, blotted dry, placed in a clean petri dish
for later use and imaged using TEM (Philips Healthcare).
Immunohistochemistry and
immunofluorescence
The kidney tissue embedded in paraffin wax was
sectioned at ~5 µm thick. After the paraffin sections were dewaxed,
the sections were washed with clean water after dewaxing solution I
(10 min), Dewaxing solution II (10 min), Dewaxing solution III (10
min), anhydrous ethanol I (5 min), anhydrous ethanol II (5 min) and
anhydrous ethanol III (5 min) in sequence. For antigen retrieval
(EDTA pH 9.0) was added to a microwaveable vessel into which the
slides were placed. The vessel was placed inside the 1,200 W
microwave. The EDTA was boiled for 8 min at 1,200 W, then under
medium heat before a pause for 8 min, followed by low and medium
heat for 7 min. Following heat recovery of antigens, endogenous
peroxidase activity was blocked using 0.3% hydrogen peroxide for 10
min at room temperature. After preincubation with 10% BSA
(MilliporeSigma) for 10 min at room temperature to prevent
nonspecific binding of antibodies, renal tissues were incubated
with primary antibodies against COL-IV (1:500) overnight at 4°C.
After incubation with the appropriate secondary antibodies,
sections were developed with 3,3′-diaminobenzidine and then
counterstained with hematoxylin for 5 min (all at room
temperature). Images were captured using a fluorescence microscope
(Leica Microsystems GmbH).
Kidney tissues were sectioned, dewaxed, rehydrated
and subjected to microwave antigen recovery in citrate buffer for
15 min as aforementioned in immunohistochemistry. After cooling,
slides were blocked with 4% hydrogen peroxide for 10 min at room
temperature, 5% BSA (MilliporeSigma) for 1 h at room temperature,
then incubated with primary antibodies LC3 (cat. no. 4599T) and P62
(cat. no. 23214S), both at 1:100 overnight at 4°C. Slides were
washed with PBS and incubated with secondary antibodies
(FITC-labeled mouse IgG; cat. no. A0568, Beyotime Institute of
Biotechnology; 1:500) for 1 h at room temperature. Slides were
washed and stained with DAPI at room temperature for 3 min to
visualize the nuclei and images were captured using a fluorescence
microscope (Leica Microsystems GmbH).
NRK-52E cells were transfected with the RFP-GFP-LC3
plasmid, then fixed with 4% paraformaldehyde for 15 min at room
temperature. Cells were imaged using a confocal microscope.
Western blotting
Kidney tissues and NRK-52E cells were homogenized in
RIPA (NCM Biotech) buffer containing a protease inhibitor mixture
(cat. no. P001; NCM Biotech) and centrifuged at 13,000 × g at 4°C
for 10 min. The supernatant was collected and protein
concentrations were quantified using the BCA assay. Protein
concentration was made uniform with the addition of lysate, 5X SDS
Loading Buffer was added to samples by volume and mixed, and the
samples were boiled at 100°C for 10 min. Equivalent amounts of
protein (30 µg) were separated by SDS-PAGE on 8–12% gels and
transferred to polyvinylidene difluoride membranes with transfer
buffer at room temperature for 1.5 h. Membranes were then blocked
in 5% skim milk powder dissolved in TBST (20% Tween) for 2 h at
room temperature. The following target antibodies were then added
to the membranes: LC3 (cat. no. 4108S; 1:1,000; Cell Signaling
Technology, Inc.), P62 (cat. no. P0067; 1:1,000, MilliporeSigma)
and COL-IV (cat. no. AF0510; 1:1,000, Affinity Biosciences), α-SMA
(cat. no. 19245T; 1:1,000; CST), ATG5 (cat. no. 12994T; 1:1,000;
CST), ATG7 (cat. no. 8558T; 1:1,000; CST), TGF-1β (cat. no. 3709S;
1:1,000; SCT), AMPK (cat. no. 4150T; 1:1,000; CST), P-AMPK (cat.
no. 50081S, 1:1,000, CST), PI3K (cat. no. 4249T, 1:1,000, CST),
phosphorylated (p-)PI3K (cat. no. 17366, 1:1,000, CST), AKT (cat.
no. 4691, 1:1,000, CST), p-AKT (cat. no. 4060, 1:1,000, CST), m-TOR
(cat. no. 2983; 1:1,000; CST), p-mTOR (cat. no. 5536T; 1:1,000;
CST), MT1 (cat. no. sc-390328; 1:500; Santa Cruz Biotechnology,
Inc), MT2 (cat. no. c-398788; 1:500; Santa Cruz Biotechnology,
Inc.) and β-actin (cat. no. 4970T; 1:1,000; CST). The secondary
antibody (cat. no. 7074P2; 1:5,000; CST) was incubated for 2 h at
room temperature. Protein bands were visualized using a
chemiluminescence kit (NCM Biotech). Semi-quantification of protein
expression levels was performed using ImageJ 8.0 software (National
Institutes of Health).
Statistical analysis
SPSS software (version 26; IBM Corp.) was used for
data analysis. One-way ANOVA followed by Tukey's or Tamhane's test,
or unpaired Student's t-test, were used to determine statistical
significance between groups. Data are expressed as the mean ±
standard deviation. Each experiment was repeated three times.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Effects of MLT on glucose and insulin
tolerance, renal weight index (RI), lipid metabolism and oxidative
damage in diabetic rats
The potential protective effects of MLT against DN
in a rat model of DM were examined (Fig. 1A). To evaluate glucose and insulin
tolerance, BG levels were measured at different time points (0, 30,
60, 90 and 120 min) following treatment with glucose and insulin in
DM rats, As shown in Fig. 1B, the
glucose tolerance-AUC (G-AUC) and insulin tolerance-AUC (I-AUC)
values were significantly higher in rats in the DM group compared
with those in the control group. After treatment with MLT, G-AUC
was significantly lower in rats compared with that in the DM group,
whereas I-AUC was not significantly different in MLT-treated rats
compared with that in DM rats. Serum creatinine and urine
albumin-creatinine ratio levels were significantly increased in the
DM group compared with those in the control group, whereas MLT
treatment significantly decreased these levels compared with those
in the DM group. The relative RI was calculated as follows: Kidney
weight/body weight. The relative RI of DM rats was significantly
increased compared with that in the control group, whereas MLT
treatment inhibited the increase in relative RI. Therefore, these
results suggested that MLT improved glucose tolerance in DM rats
but had no significant effect on insulin tolerance.
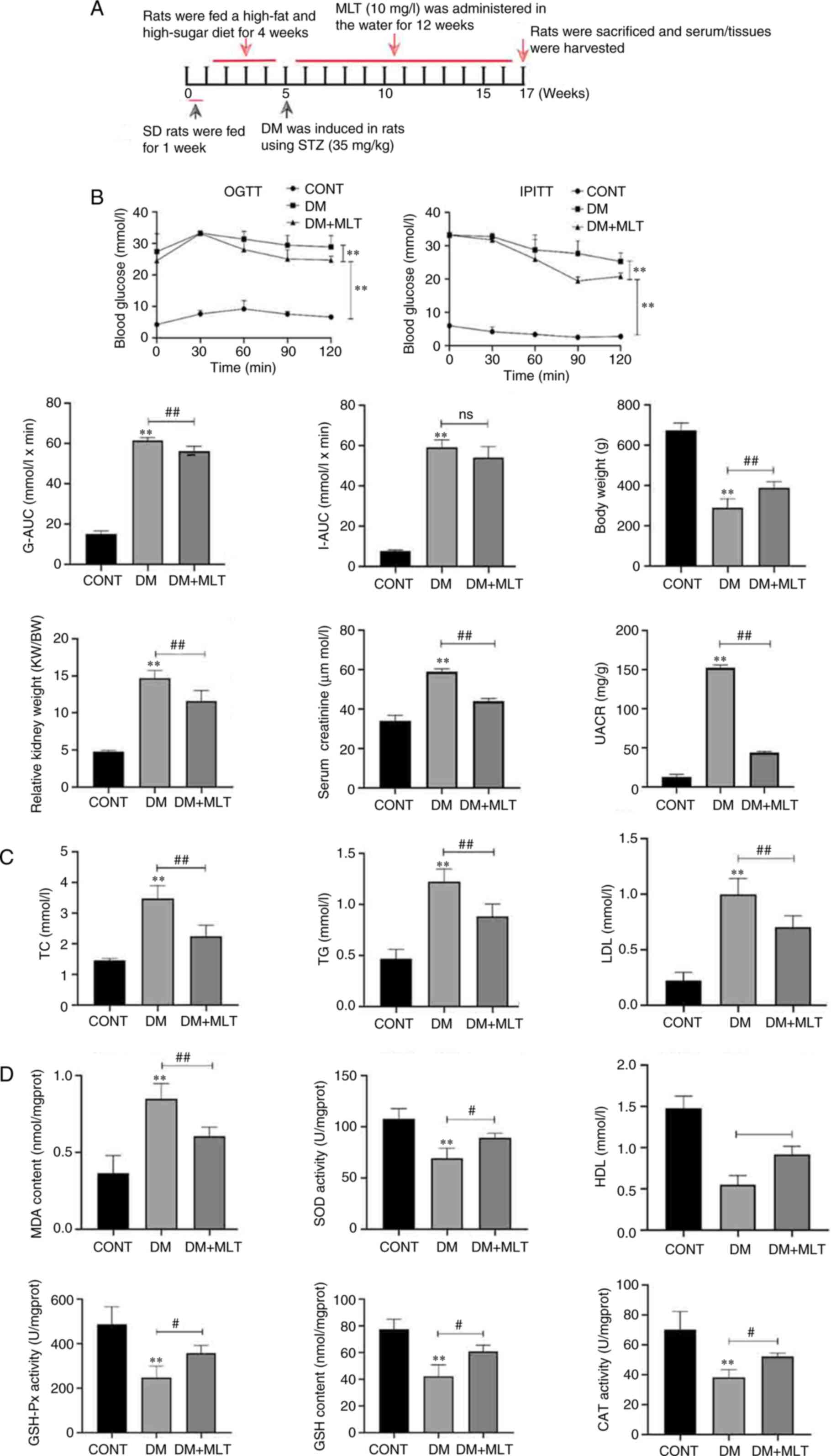 | Figure 1.Effects of MLT on glucose tolerance,
insulin tolerance, relative renal weight index, lipid metabolism
and oxidative damage in DM rats. (A) Treatment protocol to evaluate
potential protective effects of MLT in DM rats. (B) G-AUC, I-AUC
and relative renal weight index of rats in each group. OGTT, IPITT,
serum creatinine and UACR levels were analyzed. (C and D) ELISA
kits were used to measure kidney injury index. Data are presented
as the mean ± standard deviation (n=5). **P<0.01 vs. CONT;
#P<0.05 ##P<0.01 vs. DM, Sprague
Dawley; DM, diabetes mellitus; STZ, streptozotocin; CONT, control;
MLT, melatonin; OGTT, oral glucose tolerance test; IPITT,
intraperitoneal insulin tolerance test; G-AUC, glucose
tolerance-area under the curve; I-AUC, insulin tolerance-area under
the curve; UACR, urine albumin-creatinine ratio; TC, total
cholesterol; TG, triglycerides; LDL, low-density lipoprotein; MDA,
malondialdehyde; SOD, superoxide dismutase; HDL, high-density
lipoprotein; GSH-Px, glutathione peroxidase; CAT, catalase; ns, not
significant; KW, kidney weight; BW, body weight. |
The effects of MLT on lipid metabolism were also
examined. Compared with those in the control group, the DM group of
rats demonstrated significantly increased total cholesterol (TC),
triglycerides (TG) and low-density lipoprotein (LDL) levels
(Fig. 1C). After 12 weeks of MLT
treatment, TC, TG and LDL levels were significantly decreased
compared with those in the DM group. However, compared with those
in the control group, high-density lipoprotein (HDL) levels were
reduced in the DM group and after treatment with MLT, HDL levels
were significantly increased compared with those in the DM group
(Fig. 1D). These results suggested
that MLT treatment partially recovered the lipid metabolism levels
in diabetic rats.
The effects of MLT on oxidative damage were also
examined. Compared with in the control group, the DM group of rats
demonstrated a significant increase in MDA content, and a
significant decrease in SOD, CAT, GSH-PX and GSH activities
(Fig. 1D). After intervention with
MLT, MDA content was significantly lower, and levels of SOD, CAT,
GSH-PX and GSH were significantly higher compared with those in the
DM group. These data suggested that MLT significantly reduced
oxidative damage in DM rats.
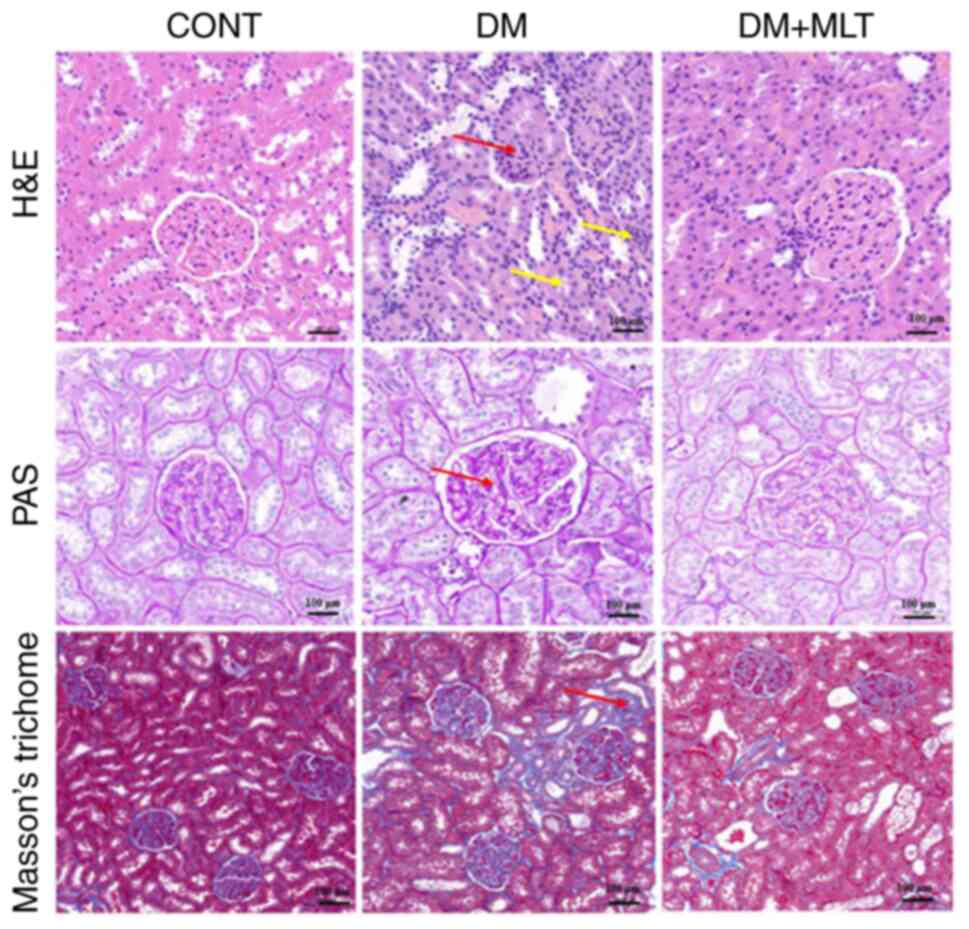
Effects of MLT on histopathological
changes in kidney tissue in DM rats
The effects of MLT treatment on histopathological
changes in the kidneys of DM rats were evaluated. H&E staining
demonstrated that the DM group exhibited thickening of the
glomerular basement membrane and shrinkage of the glomeruli (red
arrow). In addition, some tubular epithelial cells were vacuolated,
necrotic, atrophied and detached, and infiltration of interstitial
inflammatory cells was observed (yellow arrow; Fig. 2). However, in the control group,
the renal tubule structure was clearly visible, epithelial cells
were neatly arranged, the basement membrane was more intact and
there was no major collagen fiber deposition. PAS staining further
demonstrated that the glomerular basement membrane in DM rats was
thickened (red arrow). Masson's trichome staining demonstrated blue
collagen fiber deposition in the interstitial glomeruli and tubules
of DM rats (red arrow). Following intervention with MLT, glomerular
and tubular lesions, interstitial inflammatory cell infiltration,
congestion and collagen fiber deposition were reduced. These
results suggested that MLT protected against kidney injury.
Effects of MLT on renal fibrosis and
MLT receptors
The expression levels of COL-IV in kidney tissue
were examined and it was demonstrated that the protein expression
levels of COL-IV were markedly increased in the kidney tissue of DM
rats compared with those in the control rats (Fig. 3A). MLT reduced the protein
expression levels of COL-IV compared with those in the DM group. In
NRK-52E cells, HG treatment caused an increase in the protein
expression levels expression of α-SMA, COL-IV and TGF-β1 compared
with those in the control group (Fig.
3B). MLT treatment markedly reduced the expression levels of
these proteins.
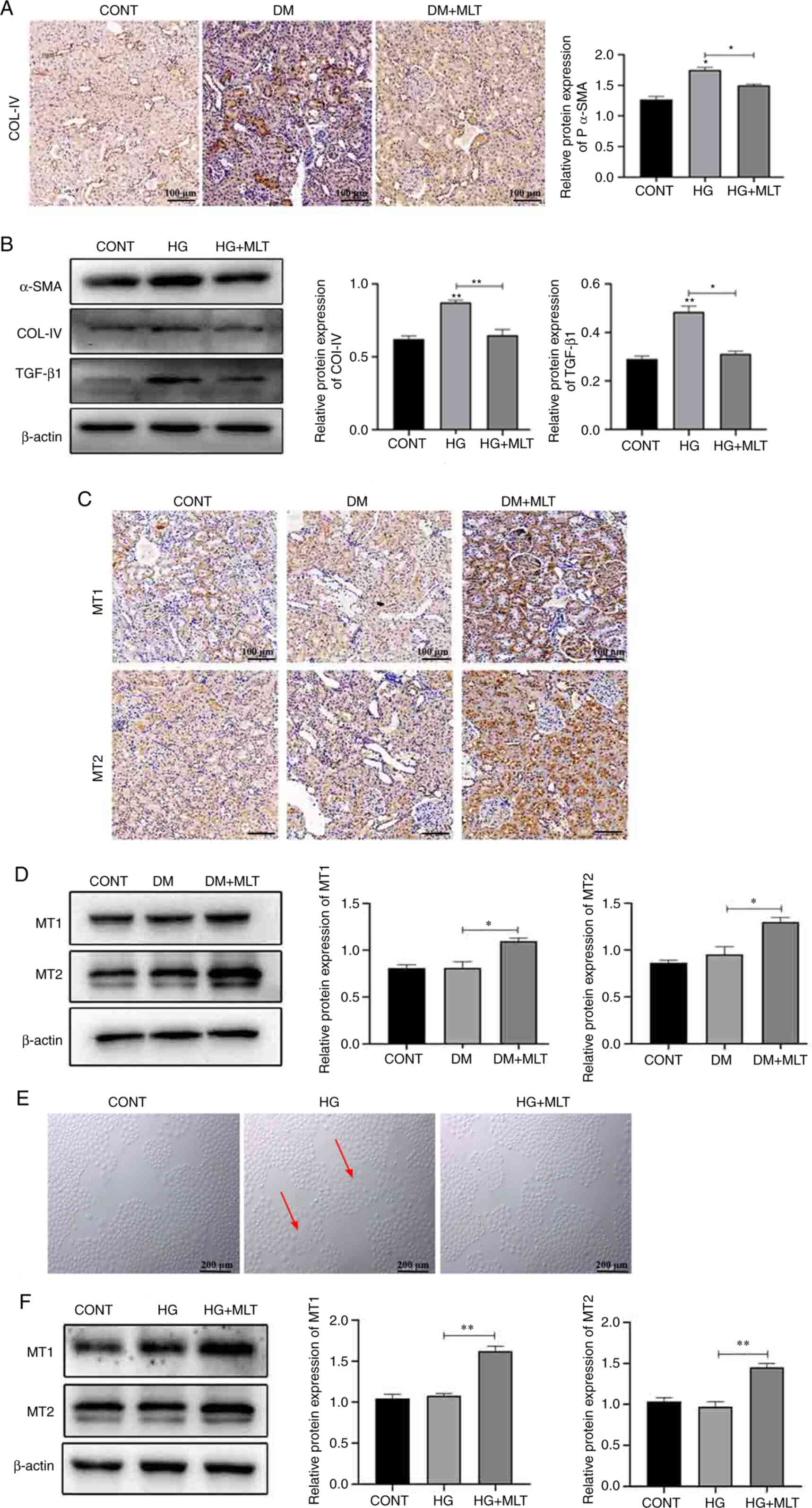 | Figure 3.Effects of MLT on renal fibrosis and
MLT receptors. (A) Immunohistochemical staining of COL-IV in kidney
tissue of DM rats. Scale bar, 100 µm. (B) Protein expression levels
of α-SMA, COL-IV, TGF-β1 and β-actin in HG- and MLT-treated cells.
(C) Immunohistochemical staining of MT1 and MT2 in kidney tissue of
DM rats. (D) Protein expression levels of MT1 and MT2 in kidney
tissue of DM rats treated with MLT. (E) Morphological changes of
NRK-52E cells with HG and MLT treatment (red arrows indicate
damaged cells). Scale bar, 200 µm. (F) Protein expression levels of
MT1 and MT2 in NRK-52E cells treated with HG and MLT. Data are
presented as the mean ± standard deviation (n=3). *P<0.05,
**P<0.01. CONT, control; DM, diabetes mellitus; MLT, melatonin;
COL-IV, type IV collagen; HG, high glucose; MT1, melatonin receptor
1; MT2, melatonin receptor 2. |
The effects of MLT were also assessed on the
expression of MT1 and MT2 in both kidney tissue and NRK-52E cells.
No marked change was demonstrated in MT1 and MT2 expression in DM
rats compared with that in the control group. Following treatment
with MLT, the expression levels of MT1 and MT2 were significantly
increased compared with those in the DM group (Fig. 3C and D). In NRK-52E cells, HG
induced cellular injury (red arrows); however, after treatment with
MLT, cell injury was reduced (Fig.
3E). Next, the effect of MLT on MT1 and MT2 expression levels
in NRK-52E cells was examined (Fig.
3F). MLT treatment increased the expression levels of MT1 and
MT2 in HG-induced NRK-52E cells compared with those in the HG
group. These results suggested that MLT exerts a protective effect
against DM-induced kidney damage and HG-induced NRK-52E cell
injury.
Effects of MLT on mitochondrial
structure and autophagy in kidney tissue of DM rats and NRK-52E
cells
TEM was used to examine the structure of
mitochondria and autophagosomes in DM rat kidney tissue. Compared
with those in the control group, the mitochondria of kidney cells
in the DM group were dilated and swollen, cristae were reduced or
absent (yellow arrow) and a large number of mitochondria were
surrounded by a double-membrane structure (red arrow), forming
autophagosomes (Fig. 4A). After
treatment with MLT for 12 weeks, a reduction in mitochondrial
damage in rat kidney tissues was observed, in addition to a
reduction in the number of damaged mitochondria wrapped in bilayer
membranes. Immunofluorescence was used to measure the expression of
LC3 and P62 in DM rat kidney tissue. Compared with that in the
control group, the fluorescence intensity of LC3 and P62 were
increased in the DM group (Fig.
4B). MLT treatment reduced the fluorescence intensity of LC3
and P62 compared with that in the DM group. Immunohistochemistry
was used to analyze the expression levels of autophagy protein
(ATG)5, ATG7 and LC3. The expression levels of ATG5, ATG7 and LC3
were increased in the kidneys of DM rats compared with those in the
control rats and MLT treatment reduced the expression levels of
these proteins (Fig. 4C). Western
blotting was used to semi-quantify the protein expression levels of
autophagy-related proteins. In DM rats, the protein expression
levels of ATG5, ATG7, P62 and LC3II/I were significantly increased,
whereas MLT treatment significantly reduced the expression of these
proteins (Fig. 4D).
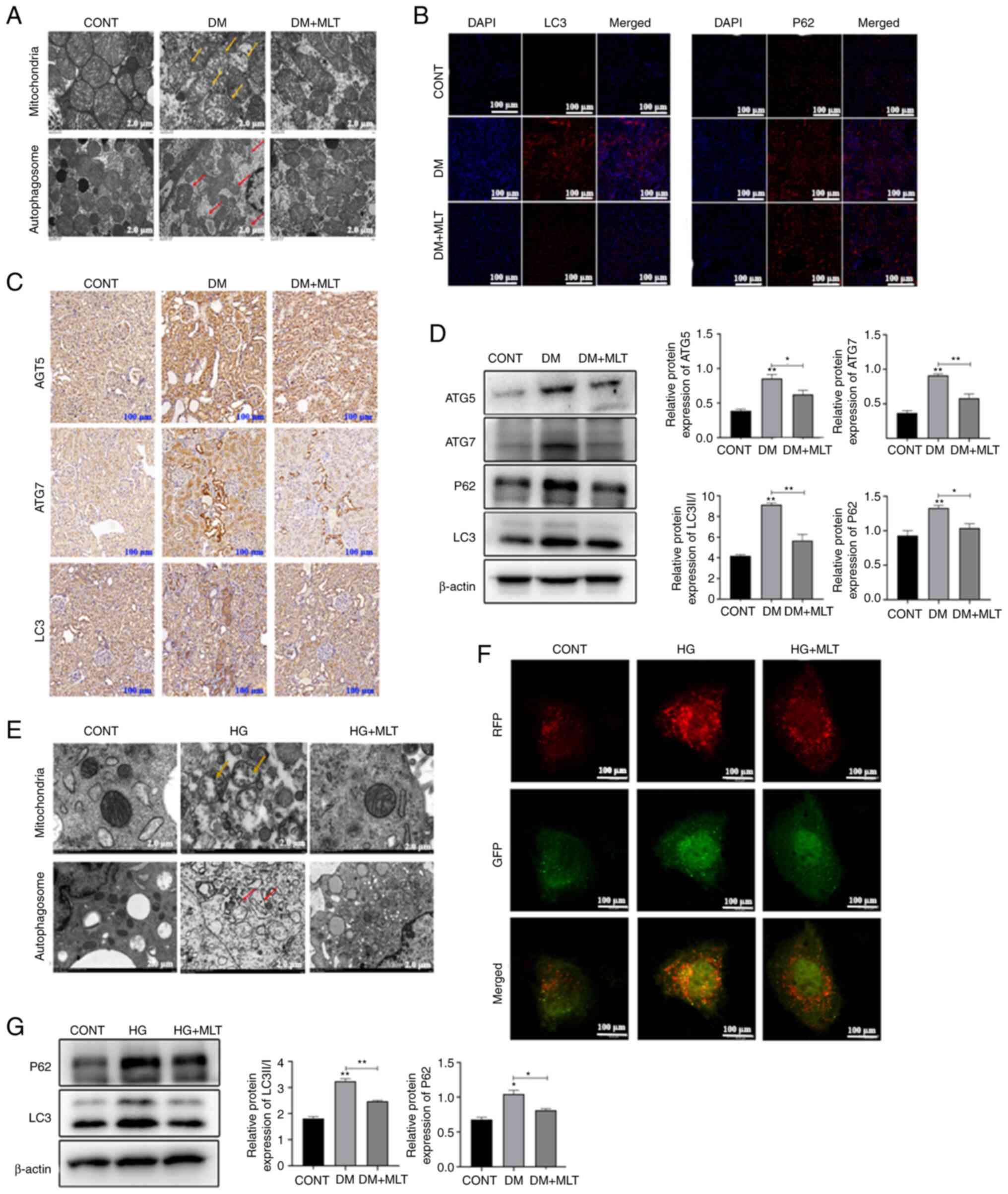 | Figure 4.Effects of MLT on mitochondria
structure and autophagy in kidney tissue of DM rats and NRK-52E
cells. (A) Ultrastructure of mitochondria and autophagosomes of
kidney cells of DM rats treated with MLT (yellow arrow indicates
damaged mitochondria; red arrow indicates autophagosomes). Scale
bar, 2 µm. (B) LC3 and P62 immunofluorescence staining of kidney
tissues of DM rats treated with MLT. Scale bar, 100 µm. (C)
Immunohistochemical staining of ATG5, ATG7 and LC3 in kidneys of DM
rats treated with MLT. Scale bar, 100 µm. (D) Protein expression
levels of ATG5, ATG7, P62, LC3 and β-actin in kidney tissue of DM
rats. (E) Ultrastructure of mitochondria and autophagosomes of
NRK-52E cells treated with HG and MLT (yellow arrow represents
indicates mitochondria; red arrow indicates autophagosomes). Scale
bar, 2 µm. (F) RFP and GFP immunofluorescence staining of NRK-52E
cells treated with HG and MLT. Scale bar, 100 µm. (G) Protein
expression levels of P62, LC3 and β-actin in NRK-52E cells treated
with HG and MLT. Data are presented as the mean ± standard
deviation (n=3). *P<0.05, **P<0.01. CONT, control; DM,
diabetes mellitus; MLT, melatonin; HG, high glucose; ATG,
autophagy-related protein. |
To further examine the effect of MLT on DM-induced
autophagy, NRK-52E cell model was induced by HG. TEM was used to
detect the mitochondrial structure in HG-induced NRK-52E cells and
demonstrated that the mitochondrial ridge disappeared, mitochondria
were swollen (yellow arrow) and numerous mitophagosomes (red arrow)
were observed (Fig. 4E). After MLT
treatment, mitochondrial structures were generally intact and
mitophagosome numbers were reduced. To determine the mechanism by
which this occurs, western blotting and immunofluorescence were
used to analyze changes in autophagy. HG inhibited autophagy
degradation by mediating an increase in the protein expression
levels of P62, whereas MLT treatment reduced the protein expression
levels of P62 and LC3II and decreased the yellow fluorescence
signal (Fig. 4F and G). Therefore,
these data suggested that MLT protected mitochondria structure in
DM rat tissues and HG-induced NRK-52E cells and could also regulate
autophagy levels.
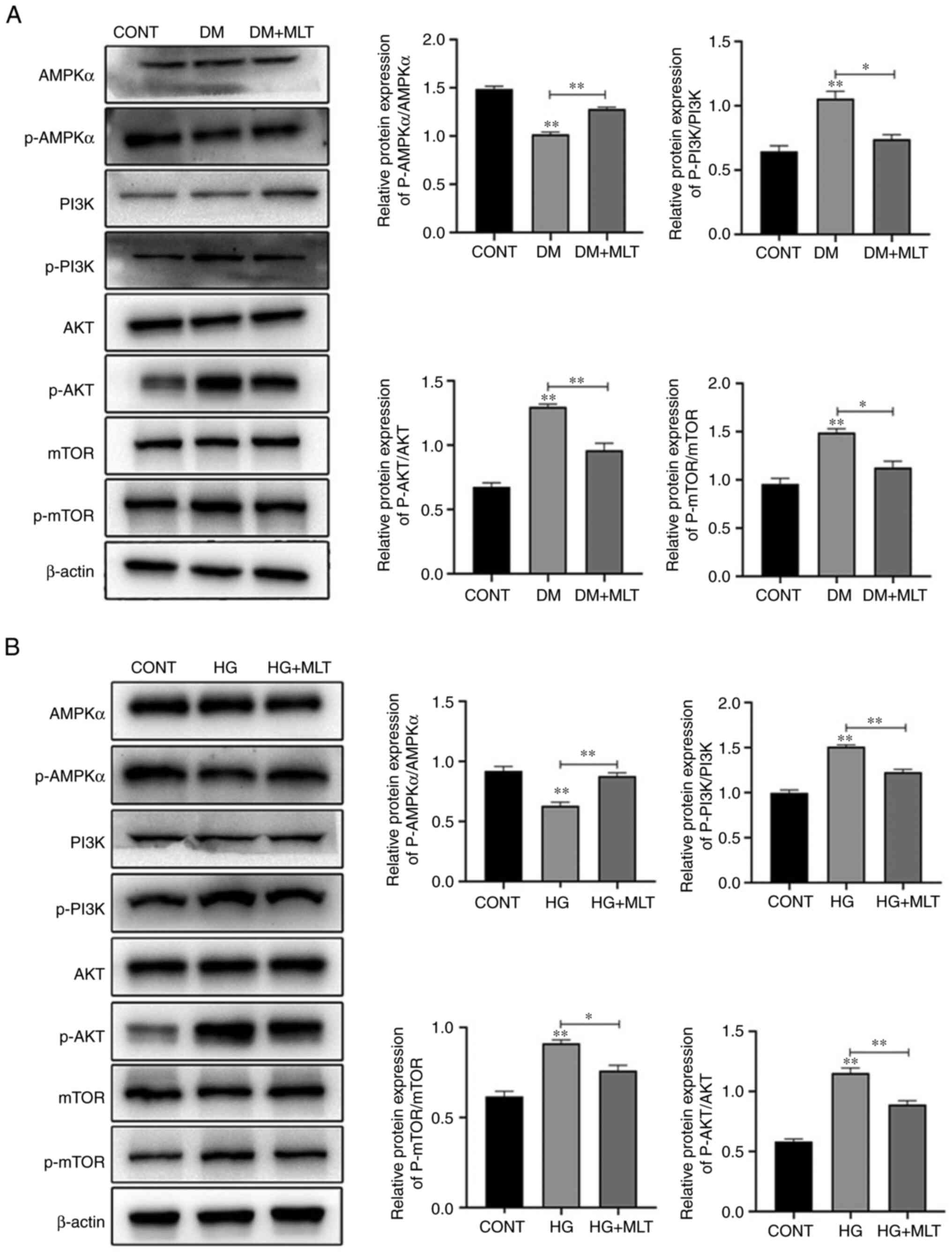
Effects of MLT on AMPKα/PI3K/AKT/mTOR
signaling in DM rats and NRK-52E cells
To further examine the effect of MLT on the
regulation of the autophagy pathway, western blotting was used to
examine the expression of autophagy-related signaling pathway
proteins. In DM rat kidney tissues and NRK-52E cells, the
phosphorylation levels of the key autophagy protein AMPKα were
significantly decreased, whereas the phosphorylation levels of
PI3K, AKT and mTOR were significantly increased compared with those
in the control groups (Fig. 5A and
B). After treatment with MLT, the phosphorylation levels of
AMPKα were significantly increased, whereas the phosphorylation
levels of PI3K, AKT and mTOR were significantly decreased. These
results suggested that MLT affected the AMPKα/PI3K/AKT/mTOR
signaling pathway, which is important for autophagy formation.
Discussion
Diabetes mellitus (DM) is one of the chronic
metabolic noncommunicable diseases that has become a worldwide
epidemic (26). DN is one of the
most common microvascular complications associated with DM and is a
major cause of end-stage renal disease (ESRD) (27). The onset of DM causes certain
pathological changes in kidneys, including the deposition of
extracellular matrix mainly in the glomerular membrane, thickening
of the glomerular basement membrane, proliferative changes and
tubular atrophy, ultimately leading to interstitial fibrosis and
glomerulosclerosis (28). In the
present study, the aforementioned features of kidney tissue damage
were demonstrated in rats with DM, such as mitochondrial expansion
and swelling, and reduction or disappearance of cristae, indicating
that DM rats were a suitable model for DN. In addition,
hyperglycemia also increases ROS production, and excessive ROS can
lead to increased expression of extracellular matrix genes by
regulating the activation of PKC, mitogen-activated protein kinase
and certain cytokines (IL-18) and transcription factors (NF-κB),
which can lead to the development of fibrosis and ESRD (29).
In the present study, the MDA content in DM rat
kidney tissues was significantly increased, and the levels of SOD,
CAT, GSH-Px and GSH were significantly decreased. This result
suggested the presence of oxidative damage in kidney tissues of DM
rats. DN involves multiple pathologies, including abnormal glucose
metabolism, oxidative stress, endoplasmic reticulum stress and
inflammatory responses. Novel therapeutic treatments, in addition
to identifying cellular processes underlying the pathogenesis of
DN, may establish future successful treatments for DN.
Autophagy is a process that involves the removal of
damaged proteins and organelles to maintain cellular homeostasis
under various stress conditions and includes macroautophagy,
microautophagy and chaperone-mediated autophagy (30). Autophagy can regulate a variety of
stress responses, and disruptions in this regulatory mechanism may
be related to age and the pathogenesis of DM-related diseases
(31,32). It has previously been reported that
in STZ-induced type 1 DM or HG conditions, the autophagic activity
of podocytes is reduced, accompanied by a decrease in expression
levels of autophagy-related proteins, Beclin-1, ATG12-5 and LC3
(33,34). Li and Siragy (35) also reported defective autophagy,
and decreased LC3II and P62 accumulation in podocytes cultured with
HG, suggesting that hyperglycemia reduces autophagic activity in
podocytes. However, in other previous studies, the expression of
the autophagy-related proteins LC3II and Beclin-1, and the number
of autophagosomes, were found to increase in podocytes during HG
treatment (36,37). This is consistent with studies
conducted by Lenoir et al (38) and Dong et al (39), which reported similar results in
primary podocytes and podocyte cell lines, suggesting that HG
promotes autophagy in podocytes. Katsuragi et al (40) reported that the protein expression
levels of LC3II and P62 were significantly increased in HK-2 cells
after exposure to late glycosylation end products, suggesting that
the accumulation of autophagosomes is caused by reduced lysosomal
degradation in renal tubules. In the present study, it was
demonstrated that the expression levels of LC3II and P62 were
significantly increased in the kidney tissue of DM rats. In the
late stage of DM, glucose utilization is deficient and other
substrates used for energy are used preferentially, which can
result in nutritional deficiencies. In this situation, LC3I is
rapidly converted to LC3II, which results in the formation of
autophagosomes to degrade proteins and damaged organelles to
provide energy (41). P62 is a
multifunctional protein that can link ubiquitinated proteins to
LC3/ATG8 separately and undergoes self-oligomerization, entering
the autophagolysosome to degrade ubiquitinated substrates (42). However, when the expression of P62
is upregulated, the degradation of autophagic lysosomes is
impaired, resulting in poor autophagic flux and the accumulation of
large amounts of autophagosomes, which may also be a potential
reason for the occurrence of ESRD. In the present study, it was
further demonstrated that HG prevented the degradation of
autophagic lysosomes in NRK-52E cells as P62 was increased, which
inhibited the autophagic flux.
AMPK is a key enzyme in the protein kinase signaling
transduction chain and is also an important regulator of
intracellular energy and metabolic balance (43). Activation of AMPK promotes the
early inhibition of mTOR and induces autophagy (44). However, in the present study, AMPK
phosphorylation was inhibited in the kidneys of DM rats and the
protein expression levels of P62 were increased, demonstrating
inhibition of the autophagic flux in DN, which could lead to kidney
injury. AMPK can also phosphorylate the Raptor subunit of the mTOR
complex 1 (mTORC1), which inhibits activation of mTORC1 (45). In the present study, it was
demonstrated that phosphorylation of PI3K/AKT/mTOR was increased in
the kidneys of DM rats and NRK-52E cells, which further suggested
that HG prevented the autophagic flux. Therefore, renal injury
could be prevented by promoting autophagy degradation.
MLT is an indoleamine that is synthesized by the
pineal gland and produced by peripheral organs, such as the retina,
Harderian glands, intestines and skin (46). The secretion of MLT follows a
circadian rhythm, which is suppressed during the day and active at
night (47,48). Therefore, in the present study, MLT
treatment was administered in the night. MLT has numerous
biological activities, including antioxidant, anti-inflammatory and
anti-apoptotic properties, and the mechanisms by which MLT protects
against DN include the regulation of ROS and AMPK signaling
(49). In the present study, it
was demonstrated that oxidative damage, mitochondrial damage and
fibrosis in renal tissue was alleviated in DM rats after receiving
MLT treatment. Meanwhile, the expression levels of LC3II and P62
were downregulated in vitro and in vivo. These
findings indicated that MLT promoted autophagy degradation, which
facilitated the occurrence of autophagy. Previous studies also
indicated that MLT inhibits cell death by mediating autophagy
(50,51). In addition, MLT also inhibited the
phosphorylation of PI3K/AKT/mTOR in the kidneys of DM rats and
NRK-52E cells, which further demonstrated the protective effect of
MLT through the regulation of autophagy. A previous study reported
that MLT can effectively alleviate nickel-induced brain injury by
activating the PI3K/AKT/mTOR signaling pathway (52). In the present study, it was also
demonstrated that MLT treatment promoted the protein expression
levels of MT1 and MT2 in vivo and in vitro, which may
be beneficial to renal injury. In a previous study, MLT inhibited
apoptosis by binding to surface membrane receptors MT1 and MT2 in
chondrocytes (53), which further
indicated the importance of MT1 and MT2 in renal injury. Taken
together, these data suggested that MLT may serve a role in
protecting against renal injury in DM and NRK-52E cell injury by
alleviating the impairment of autophagy, promoting the degradation
of misfolded proteins and damaged organelles, and attenuating renal
fibrosis.
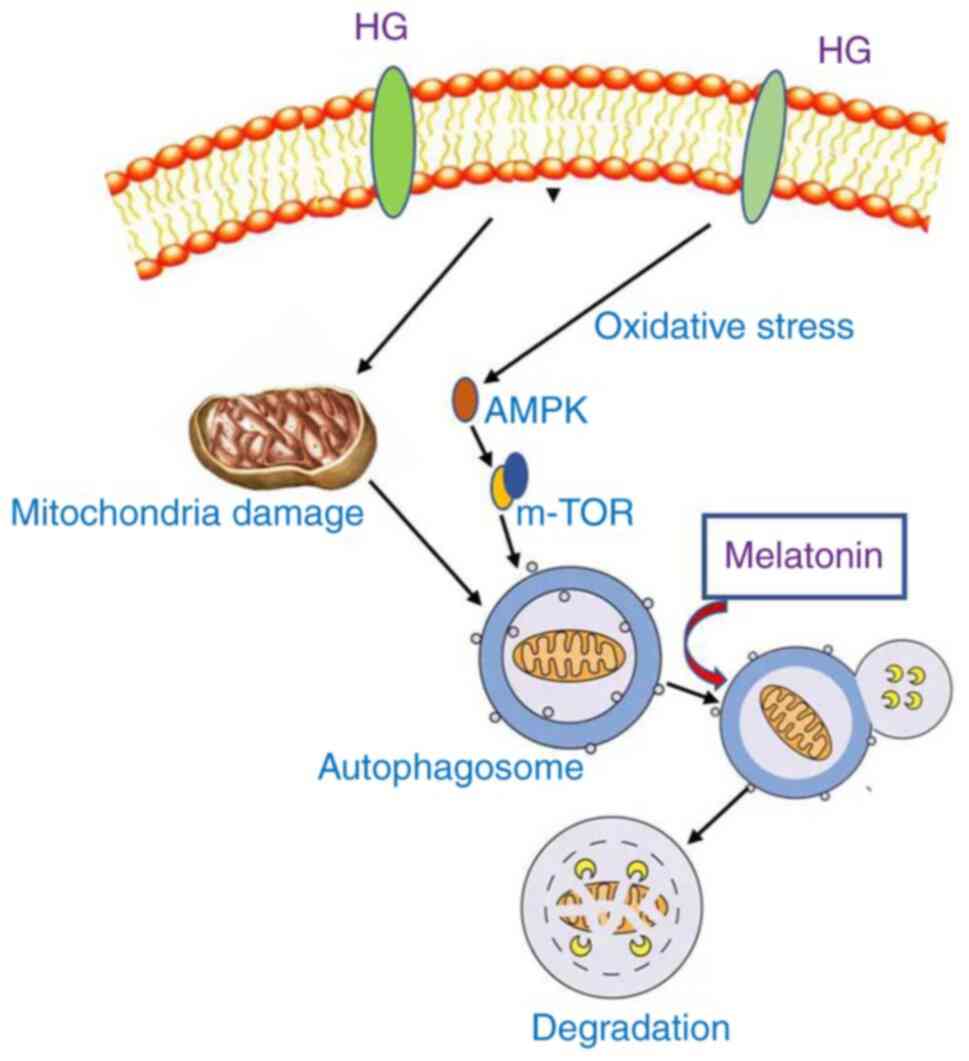
In conclusion, MLT exerts a protective effect
against renal injury in DM rats and NRK-52E cells by alleviating
the impairment of autophagy (Fig.
6). The present study provided novel evidence for the
relationship between MLT and autophagy in the context of HG
treatment. MLT promoted autophagy degradation by regulating the
autophagic flux, which further alleviated kidney damage. In future
studies, specific components of the AMPK/PI3K/AKT/mTOR signaling
pathway and the process of autophagy should be investigated to
further confirm the conclusions reached in the present study.
Acknowledgements
Professor Lin Wang (Shandong Agricultural
University, College of Veterinary Medicine, Taian, Shangdong,
China) provided the GFP-RFP-LC3 plasmid.
Funding
This work was supported by the Priority Academic Program
Development of Jiangsu Higher Education Institutions.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
ZL and YL conceived and designed the study. NL
performed the experiment, figure preparation and manuscript draft.
NL and YM developed the methods and performed data analysis. NL and
YW acquired and interpreted the data. NL wrote and revised the
paper. ZL and YL confirm the authenticity of all the raw data. All
authors agree to be accountable for all aspects of the work in
ensuring that questions related to the accuracy or integrity of any
part of the work are appropriately investigated and resolved. All
authors have read and approved the final manuscript.
Ethics approval and consent to
participate
The experimental procedures were approved by the
Ethics Committee of Yangzhou University [Yangzhou, China; approval
no. SYXK (Su) 2022-0044].
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
CAT
|
catalase
|
|
DN
|
diabetic nephropathy
|
|
DM
|
diabetes mellitus
|
|
GSH-Px
|
glutathione peroxidase
|
|
AUC
|
area under the curve
|
|
G-AUC
|
glucose tolerance-AUC
|
|
I-AUC
|
insulin tolerance-AUC
|
|
MLT
|
melatonin
|
|
MDA
|
malondialdehyde
|
|
SOD
|
superoxide dismutase
|
References
|
1
|
Sha J, Sui B, Su X, Meng Q and Zhang C:
Alteration of oxidative stress and inflammatory cytokines induces
apoptosis in diabetic nephropathy. Mol Med Rep. 16:7715–7723. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Thibodeau JF, Holterman CE, Burger D, Read
NC, Reudelhuber TL and Kennedy CR: A novel mouse model of advanced
diabetic kidney disease. PLoS One. 9:e1134592014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Gao Y, Ma Y, Xie D and Jiang H: ManNAc
protects against podocyte pyroptosis via inhibiting mitochondrial
damage and ROS/NLRP3 signaling pathway in diabetic kidney injury
model. Int Immunopharmacol. 107:1087112022. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zhong Y, Liu J, Sun D, Guo T, Yao Y, Xia
X, Shi C and Peng X: Dioscin relieves diabetic nephropathy via
suppressing oxidative stress and apoptosis, and improving
mitochondrial quality and quantity control. Food Funct.
13:3660–3673. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Teh YM, Mualif SA and Lim SK: A
comprehensive insight into autophagy and its potential signaling
pathways as a therapeutic target in podocyte injury. Int J Biochem
Cell Biol. 143:1061532022. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Medras ZJH, Mostafa YM, Ahmed AAM and
EI-Sayed NM: Arctigenin improves neuropathy via ameliorating
apoptosis and modulating autophagy in streptozotocin-induced
diabetic mice. CNS Neurosci Ther. May 11–2023.(Epub ahead of
print). View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wang LH, Wang YY, Liu L and Gong Q: From
diabetes to diabetic complications: Role of autophagy. Curr Med
Sci. 43:434–444. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Liu L, Dai WZ, Zhu XC and Ma T: A review
of autophagy mechanism of statins in the potential therapy of
Alzheimer's disease. J Integr Neurosci. 21:462022. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Feng H, Wang N, Zhang N and Liao HH:
Alternative autophagy: Mechanisms and roles in different diseases.
Cell Commun Signal. 20:432022. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lim JH, Kim HW, Kim MY, Kim TW, Kim EN,
Kim Y, Chung S, Kim YS, Choi BS, Kim YS, et al: Cinacalcet-mediated
activation of the CaMKKβ-LKB1-AMPK pathway attenuates diabetic
nephropathy in db/db mice by modulation of apoptosis and autophagy.
Cell Death Dis. 9:2702018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Song FQ, Song M, Ma WX, Gao Z, Ti Y, Zhang
X, Hu BA, Zhong M, Zhang W and Yu Y: Overexpressing STAMP2
attenuates diabetic renal injuries via upregulating autophagy in
diabetic rats. Biochem Biophys Res Commun. 579:47–53. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lian CY, Chu BI, Xia WH, Wang ZY, Fan RF
and Wang L: Persistent activation of Nrf2 in a p62-dependent
non-canonical manner aggravates lead-induced kidney injury by
promoting apoptosis and inhibiting autophagy. J Adv Res. 46:87–100.
2023. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hsiao CC, Lin CC, Hou YS, Ko JY and Wang
CJ: Low-energy extracorporeal shock wave ameliorates streptozotocin
induced diabetes and promotes pancreatic beta cells regeneration in
a rat model. Int J Mol Sci. 20:49342019. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Owino S, Contreras-Alcantara S, Baba K and
Tosini G: Melatonin signaling controls the daily rhythm in blood
glucose levels independent of peripheral clocks. PLoS One.
11:e01482142016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Agil A, Chayah M, Visiedo L,
Navarro-Alarcon M, Rodríguez Ferrer JM, Tassi M, Reiter RJ and
Fernández-Vázquez G: Melatonin improves mitochondrial dynamics and
function in the kidney of Zücker diabetic fatty rats. J Clin Med.
9:29162020. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yapislar H, Haciosmanoglu E, Sarioglu T
and Ekmekcioglu C: The melatonin MT2 receptor is
involved in the anti-apoptotic effects of melatonin in rats with
type 2 diabetes mellitus. Tissue Cell. 76:1017632022. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang W, Zhang J, Wang X, Wang H, Ren Q and
Li Y: Effects of melatonin on diabetic nephropathy rats via
Wnt/β-catenin signaling pathway and TGF-β-Smad signaling pathway.
Int J Clin Exp Pathol. 11:2488–2496. 2018.PubMed/NCBI
|
|
18
|
Fan Z, Qi X, Yang W and Wu Y: Melatonin
ameliorates renal fibrosis through the inhibition of NF-κB and
TGF-β1/Smad3 pathways in db/db diabetic mice. Arch Med Res.
51:524–534. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wei J, Wang Y, Qi X, Fan Z and Wu Y:
Melatonin ameliorates hyperglycaemia-induced renal inflammation by
inhibiting the activation of TLR4 and TGF-β1/Smad3 signalling
pathway. Am J Transl Res. 12:1584–1599. 2020.PubMed/NCBI
|
|
20
|
Chen YT, Yang CC, Sun CK, Chiang HJ, Chen
YL, Sung PH, Zhen YY, Huang TH, Chang CL, Chen HH, et al:
Extracorporeal shock wave therapy ameliorates
cyclophosphamide-induced rat acute interstitial cystitis though
inhibiting inflammation and oxidative stress-in vitro and in vivo
experiment studies. Am J Transl Res. 6:631–648. 2014.PubMed/NCBI
|
|
21
|
Xu Q and Cheung RTF: Melatonin mitigates
type 1 diabetes-aggravated cerebral ischemia-reperfusion injury
through anti-inflammatory and anti-apoptotic effects. Brain Behav.
e31182023.(Epub ahead of print). View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yu L, Gong B, Duan W, Fan C, Zhang J, Li
Z, Xue X, Xu Y, Meng D, Li B, et al: Melatonin ameliorates
myocardial ischemia/reperfusion injury in type 1 diabetic rats by
preserving mitochondrial function: Role of AMPK-PGC-1α-SIRT3
signaling. Sci Rep. 7:413372017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhang Y, Wang Y, Xu J, Tian F, Hu S, Chen
Y and Fu Z: Melatonin attenuates myocardial ischemia-reperfusion
injury via improving mitochondrial fusion/mitophagy and activating
the AMPK-OPA1 signaling pathways. J Pineal Res. 66:e125422019.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Siddhi J, Sherkhane B, Kalavala AK, Arruri
V, Velayutham R and Kumar A: Melatonin prevents diabetes-induced
nephropathy by modulating the AMPK/SIRT1 axis: Focus on autophagy
and mitochondrial dysfunction. Cell Biol Int. 46:2142–2157. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Qiu WH, An S, Wang T, Li J, Yu B, Zeng Z,
Chen Z, Lin B, Lin X and Gao YG: Melatonin suppresses ferroptosis
via activation of the Nrf2/HO-1 signaling pathway in the mouse
model of sepsis-induced acute kidney injury. Int Immunopharmacol.
112:1091622022. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Morya AK, Ramesh PV, Kaur K, Gurnani B,
Heda A, Bhatia K and Sinha A: Diabetes more than retinopathy, it's
effect on the anterior segment of eye. World J Clin Cases.
11:3736–3749. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Cooper ME: Pathogenesis, prevention, and
treatment of diabetic nephropathy. Lancet. 352:213–219. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lu Q, Ji XJ, Zhou YX, Yao XQ, Liu YQ and
Yin XX: Quercetin inhibits the mTORC1/p70S6K signaling-mediated
renal tubular epithelial-mesenchymal transition and renal fibrosis
in diabetic nephropathy. Pharmacol Res. 99:237–247. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kashihara N, Haruna Y, Kondeti VK and
Kanwar YS: Oxidative stress in diabetic nephropathy. Curr Med Chem.
17:4256–4269. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ichimiya T, Yamakawa T, Hirano T, Yokoyama
Y, Hayashi Y, Hirayama D, Wagatsuma K, Itoi T and Nakase H:
Autophagy and autophagy-related diseases: A review. Int J Mol Sci.
21:89742020. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Gonzalez CD, Lee MS, Marchetti P,
Pietropaolo M, Towns R, Vaccaro MI, Watada H and Wiley JW: The
emerging role of autophagy in the pathophysiology of diabetes
mellitus. Autophagy. 7:2–11. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Rubinsztein DC, Mariño G and Kroemer G:
Autophagy and aging. Cell. 146:682–695. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ding DF, You N, Wu XM, Xu JR, Hu AP, Ye
XL, Zhu Q, Jiang XQ, Miao H, Liu C and Lu YB: Resveratrol
attenuates renal hypertrophy in early-stage diabetes by activating
AMPK. Am J Nephrol. 31:363–374. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Xu XH, Ding DF, Yong HJ, Dong CL, You N,
Ye XL, Pan ML, Ma JH, You Q and Lu YB: Resveratrol
transcriptionally regulates miRNA-18a-5p expression ameliorating
diabetic nephropathy via increasing autophagy. Eur Rev Med
Pharmacol Sci. 21:4952–4965. 2017.PubMed/NCBI
|
|
35
|
Li C and Siragy HM: (Pro)renin receptor
regulates autophagy and apoptosis in podocytes exposed to high
glucose. Am J Physiol Endocrinol Metab. 309:E302–E310. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ma T, Zhu J, Chen X, Zha D, Singhal PC and
Ding G: High glucose induces autophagy in podocytes. Exp Cell Res.
319:779–789. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wei M, Li Z and Yang Z: Crosstalk between
protective autophagy and NF-κB signal in high glucose-induced
podocytes. Mol Cell Biochem. 394:261–273. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Lenoir O, Jasiek M, Hénique C, Guyonnet L,
Hartleben B, Bork T, Chipont A, Flosseau K, Bensaada I, Schmitt A,
et al: Endothelial cell and podocyte autophagy synergistically
protect from diabetes-induced glomerulosclerosis. Autophagy.
11:1130–1145. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Dong C, Zheng H, Huang S, You N, Xu J, Ye
X, Zhu Q, Feng Y, You Q, Miao H, et al: Heme oxygenase-1 enhances
autophagy in podocytes as a protective mechanism against high
glucose-induced apoptosis. Exp Cell Res. 337:146–159. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Katsuragi Y, Ichimura Y and Komatsu M:
p62/SQSTM1 functions as a signaling hub and an autophagy adaptor.
FEBS J. 282:4672–4678. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Han YP, Liu LJ, Yan JL, Chen MY, Meng XF,
Zhou XR and Qian LB: Autophagy and its therapeutic potential in
diabetic nephropathy. Front Endocrinol (Lausanne). 14:11394442023.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Komatsu M, Kageyama S and Ichimura Y:
p62/SQSTM1/A170: Physiology and pathology. Pharmacol Res.
66:457–462. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Herzig S and Shaw RJ: AMPK: Guardian of
metabolism and mitochondrial homeostasis. Nat Rev Mol Cell Biol.
19:121–135. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Kma L and Baruah TJ: The interplay of ROS
and the PI3K/Akt pathway in autophagy regulation. Biotechnol Appl
Biochem. 69:248–264. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Gwinn DM, Shackelford DB, Egan DF,
Mihaylova MM, Mery A, Vasquez DS, Turk BE and Shaw RJ: AMPK
phosphorylation of raptor mediates a metabolic checkpoint. Mol
Cell. 30:214–226. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Tan DX, Hardeland R, Back K, Manchester
LC, Alatorre-Jimenez MA and Reiter RJ: On the significance of an
alternate pathway of melatonin synthesis via 5-methoxytryptamine:
Comparisons across species. J Pineal Res. 61:27–40. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Linowiecka K, Slominski AT, Reiter RJ,
Böhm M, Steinbrink K, Paus R and Kleszczyński K: Moletanin: A
potential regulator of DNA methylation. Antioxidants (Basel).
12:11552023. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Shao R, Wang Y, He C and Chen L: Melatonin
and its emerging physiological role in reproduction: A review and
update. Curr Mol Med. Apr 17–2023.(Epub ahead of print). PubMed/NCBI
|
|
49
|
Zhang H, Zhang HM, Wu LP, Tan DX, Kamat A,
Li YQ, Katz MS, Abboud HE, Reiter RJ and Zhang BX: Impaired
mitochondrial complex III and melatonin responsive reactive oxygen
species generation in kidney mitochondria of db/db mice. J Pineal
Res. 51:338–344. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Dun RL, Lan TY, Tsai J, Mao JM, Shao YQ,
Hu XH, Zhu WJ, Qi GC and Peng Y: Protective effect of melatonin for
renal ischemia-reperfusion injury: A systematic review and
meta-analysis. Front Physiol. 12:7910362022. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Song D, Liu Y, Yao Y, Liu F, Tao W, Zhou
X, Li R, Zhang X and Li X: Melatonin improves bisphenol A-induced
cell apoptosis, oxidative stress and autophagy impairment via
inhibition of the p38 MAPK signaling pathway in FLK-BLV cells.
Environ Toxicol. 37:1551–1562. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Qiao S, Sun Y, Jiang Y, Chen X, Cai J, Liu
Q and Zhang Z: Melatonin ameliorates nickel induced autophagy in
mouse brain: Diminution of oxidative stress. Toxicology.
473:1532072022. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Lim HD, Kim YS, Ko SH, Yoon IJ, Cho SG,
Chun YH, Choi BJ and Kim EC: Cytoprotective and anti-inflammatory
effects of melatonin in hydrogen peroxide-stimulated CHON-001 human
chondrocyte cell line and rabbit model of osteoarthritis via the
SIRT1 pathway. J Pineal Res. 53:225–237. 2012. View Article : Google Scholar : PubMed/NCBI
|