Introduction
Colorectal cancer (CRC) is a common type of
intestinal tumour with ~2 million new cases and 1 million estimated
associated deaths reported in 2020, representing 10.7 and 9.5% of
all new cancer cases and deaths globally, respectively (1). Moreover, cancer statistics from 2018
show that CRC resulted in the deaths of ~27,390 male and 23,240
female patients in the United States, which is the highest death
toll among digestive tract tumours (2). However, despite the advances in the
treatment of CRC, such as surgical removal and systemic
chemotherapy, the mortality rate remains high due to recurrence and
distant organ metastases (3).
Distant metastasis to the liver is responsible for CRC-related
death, with a 5-year survival rate of <10% (4). Therefore, exploring new biomarkers
that indicate the promotion of tumour initiation and progression,
and assessing the underlying molecular mechanisms, are of foremost
importance to develop targeted treatments of CRC malignancies.
N6-methyladenosine (m6A), a common RNA
modification in epigenetic regulation, affects multiple aspects of
RNA metabolism, such as pre-mRNA processing, translation
efficiency, transcript stability and microRNA biogenesis (5–8).
Methyltransferase-like 3 (METTL3), also called MT-A70, belongs to
the class I methyltransferase (MTase) family and is an important
catalytic enzyme of m6A MTase systems (9). METTL3 is widely involved in numerous
cancer types, such as gastric and bladder cancer, by acting as an
oncogene or as a tumour suppressor (10). Yue et al (11) reported that METTL3 was upregulated
in gastric cancer and enhanced the stability of zinc finger
MYM-type containing 1 mRNA, thereby facilitating the
epithelial-mesenchymal transition (EMT) program and metastasis. In
another study, Han et al (12) reported that METTL3 was upregulated
in bladder cancer and reduced the expression of phosphatase and
tensin homolog by accelerating the maturation of pri-miR221/222,
which ultimately promoted the proliferation of bladder cancer. Li
et al (13) reported that,
as an oncogene, METTL3 promoted CRC progression by maintaining sex
determining region Y-Box 2 (SOX2) expression in a m6A-insulin-like
growth factor 2 mRNA binding protein 2-dependent manner. However,
it remains unclear as to whether METTL3-mediated m6A modification
affects any long noncoding (lnc) RNAs in CRC.
Small nucleolar RNA host gene 1 (SNHG1) is an
oncogenic lncRNA that serves a key role in CRC progression, such as
in proliferation, metastasis, EMT and oncogenesis (14–18).
In the present study, the association between METTL3-mediated m6A
and SNHG1 in CRC was evaluated and the function of the METTL3/SNHG1
axis in CRC was assessed.
Materials and methods
Patients and tissue samples
Pairs of CRC and adjacent tissue specimens (n=74)
were obtained from Liaoning Cancer Hospital and Institute
(Shenyang, China) during surgical resection from January 2015 to
September 2016. All tissue specimens were preserved in liquid
nitrogen. The Medical Ethics Committee of Liaoning Cancer Hospital
Research Institute approved the study (approval no.
LNCANHOS-2018-012) and all patients signed written informed consent
forms.
Cell culture
The normal human colonic epithelial NCM460 cell line
and four human CRC cell lines (LOVO, RKO, SW480 and HT29) were
purchased from The Cell Bank of Type Culture Collection of The
Chinese Academy of Sciences. All cells were cultured using RPMI
1640 medium (Gibco; Thermo Fisher Scientific, Inc.) and
supplemented with 10% foetal bovine serum (FBS; Gibco; Thermo
Fisher Scientific, Inc.), 100 IU/ml penicillin (Shanghai Baoman
Biotechnology Co., Ltd.) and 100 mg/ml streptomycin (Shanghai
Baoman Biotechnology Co., Ltd.). All cells were maintained in a
cell incubator with 5% CO2 at 37°C.
Bioinformatics analysis of data from
the Gene Expression Omnibus (GEO) and The Cancer Genome Atlas
(TCGA) databases
Data from two CR dsxdC-related GEO datasets,
GSE41258 (19) and GSE41328
(20) were analysed using GEO2R
(https://www.ncbi.nlm.nih.gov/geo/geo2r) to analyse the
expression of METTL3 in CRC (21).
Probe nos. 209265_s_at and 213653_at represent METTL3 in datasets
GSE41258 and GSE41328, respectively. The expression and
relationship of METTL3 and SNHG1 using data from TCGA (https://portal.gdc.cancer.gov/) were analysed
using the University of ALabama at Birmingham CANcer data analysis
portal (UALCAN; http://ualcan.path.uab.edu/index.html), according to
the website's instructions (22).
Potential m6A modification sites of SNHG1 were predicted using
RMBase (V2.025; http://rna.sysu.edu.cn/rmbase/index.php), according to
the website's instructions (23).
Immunohistochemistry (IHC)
staining
IHC staining was performed according to a previously
described method (24). CRC tissue
were fixed with 10% formalin at room temperature for 48 h,
dehydrated via gradient alcohol, paraffin-embedded, sliced (4-µm
thick), dewaxed in xylene (for 10 min, repeated three times) and
then rehydrated using a descending alcohol series, followed by
antigen retrieval using Target Retrieval Solution (Dako; Agilent
Technologies, Inc.), according to the manufacturer's instructions.
Hydrogen peroxide (3%) was applied for 15 min to block endogenous
peroxidase activity at room temperature. Sections were sealed using
10% Rangoat serum (Wuhan Servicebio Technology Co., Ltd.) for 5 min
at room temperature. Incubation with anti-METTL3 rabbit monoclonal
primary antibodies (1:500; cat. no. ab195352; Abcam) was performed
overnight at 4°C in the refrigerator and then the next day with
biotinylated HRP-conjugated goat anti-rabbit immunoglobulin G
secondary antibodies at 37°C for 30 min (1:2,000; cat. no.
ab205718; Abcam). This was followed by incubation with 2 µg/ml
streptavidin maleate peroxidase for 30 min at 37°C (LSAB kit; Dako;
Agilent Technologies, Inc.), staining with 3,3-diaminobenzidine
color development kit for 20 min in the dark and then
counterstaining with haematoxylin at room temperature for 2 min.
The sections were then dehydrated and finally mounted. Control
sections of tissue were processed under the same conditions but did
not contain primary antibodies. Images were observed using a
fluorescence microscope.
RNA extraction and reverse
transcription (RT)-quantitative PCR (qPCR)
Total RNA was extracted from CRC tissues and
5×105 cells of each cell line using TRIzol®
reagent according to the manufacturer's instructions (Invitrogen;
Thermo Fisher Scientific, Inc.). Reverse transcription was
performed using PrimeScript RT Master Mix at 37°C for 15 min and
85°C for 5 sec (Takara Biotechnology Co., Ltd.). Takara
Biotechnology TB green premix Ex Taq™ II (cat. no. RR820A) was used
for the qPCR, and the thermocycling conditions were as follows:
95°C for 30 sec, followed by 40 cycles of 95°C for 5 sec and 60°C
for 30 sec, and then 95°C for 15 sec and 60°C for 60 sec. The
relative expression of METTL3 or SNHG1 was quantified using the
2−ΔΔCq method (25) and
β-actin was used as an internal control (Takara Biotechnology Co.,
Ltd.). Primer sequences are listed in Table IA.
 | Table I.Primer and oligonucleotide sequences
used in the present study. |
Table I.
Primer and oligonucleotide sequences
used in the present study.
| A, Primer sequences
used in the present study |
|---|
|
|---|
| Name | Sequence
(5′-3′) |
|---|
| METTL3 | F:
TTGTCTCCAACCTTCCGTAGT |
|
| R:
CCAGATCAGAGAGGTGGTGTAG |
| SNHG1 | F:
GCACGTTGGAACCGAAGAGA |
|
| R:
GCAGCTGAATTCCCCAGGATA |
| β-actin | F:
CTTCTACAATGAGCTGCGTG |
|
| R:
TCATGAGGTAGTCAGTCAGG |
|
| B,
Oligonucleotide sequences used in the present study |
|
| Name | Sequence
(5′-3′) |
|
| METTL3 | F:
CTTGGTACCGAGCTCGGATCCATGTCGGACACGTGGAGCTC |
|
| R:
TGCTGGATATCTGCAGAATTCGCTCTGTAAGGAAGTGCTTC |
| shMETTL3#1 | F:
CCGGGCAAGAATTCTGTGACTATGGCTCGAGCCATAGTCACAGAATTCTTGCTTTTTG |
|
| R:
AATTCAAAAAGCAAGAATTCTGTGACTATGGCTCGAGCCATAGTCACAGAATTCTTGC |
| shMETTL3#2 | F:
CCGGGCTGCACTTCAGACGAATTATCTCGAGATAATTCGTCTGAAGTGCAGCTTTTTG |
|
| R:
AATTCAAAAAGCTGCACTTCAGACGAATTATCTCGAGATAATTCGTCTGAAGTGCAGC |
| shNC |
TTCTCCGAACGTGTCACGT |
| siSNHG1-1 |
GAAACAGCAGTTGAGGGTTTG |
| siSNHG1-2 |
GGTTTGCTGTGTATCACATTT |
| siSNHG1-3 |
GCCAATTGTTGATTGAACTTC |
| siNC |
UUCUCCGAACGUGUCACGUTT |
Western blotting
Western blotting was conducted as described
previously (26). Total proteins
were extracted from NCM460, HT29, LOVO and PKO cell lines using
RIPA lysis buffer (Sigma-Aldrich; Merck KGaA) and protein
concentrations were then semi-quantified using a BCA protein assay
kit (Santa Cruz Biotechnology, Inc.) Proteins (10 µl/lane) were
separated by 10% SDS-PAGE and then transferred to PVDF membranes
(Amresco, LLC). The membranes were blocked with 5% BSA
(Sigma-Aldrich; Merck KGaA) for 1 h at room temperature, after
which the blocking solution was removed and anti-METTL3 rabbit
monoclonal (1:1,000; cat. no. ab195352; Abcam) and anti-GAPDH mouse
monoclonal (1:500; cat. no. ab8245; Abcam) primary antibodies were
refrigerated at 4°C and incubated with the membranes overnight.
Primary antibodies were then washed away with TBST (0.1% Tween) and
goat anti-mouse immunoglobulin G HRP-conjugated secondary
antibodies (1:2,000; cat. no. ab205719; Abcam) were added at room
temperature for 1 h. The ECL Western Blotting substrate kit (cat.
no. ab65623; Abcam) was utilized to visualise the target proteins
and ImageJ software (v2; National Institutes of Health) was used to
analyse the protein bands.
Cell Counting Kit-8 (CCK-8) assay
HT29 and LOVO cells were seeded in 96-well plates at
a density of 2×103 cells/well and incubated at 37°C with
5% CO2. On days 1–5, 10 µl CCK-8 solution (Dojindo
Molecular Technologies, Inc.) was added to each well and incubated
at 37°C. After 2 h, the 96-well plates were removed and a
microplate reader (Bio-Rad Laboratories, Inc.) was used to measure
the absorbance at 450 nm. Experiments were performed in
triplicate.
Transwell assay
The assay was performed as previously published
(27). LOVO and HT29 cells
(5×104 for migration assays and 1×105 for
invasion assays) were seeded into uncoated or Matrigel-precoated
(BD Biosciences; 4 h at 37°C) upper chambers (Corning, Inc.).
Serum-free medium was added to both upper chambers and medium
containing 10% FBS was added to the lower cell chambers. The cells
were incubated at 37°C with 5% CO2 for 12 h, after which
the cells remaining in the upper chamber were removed and those in
the lower chamber were fixed using anhydrous ethanol at room
temperature for 30 min, stained with 1% crystal violet for 1 h and
counted using an inverted microscope (Leica Microsystems GmbH).
Plasmid and oligonucleotide
transfection
Specific lentivirus short hairpin (sh) RNA targeting
METTL3, negative control shRNA (shNC) and METTL3 overexpression
plasmids were synthesized by Shanghai GenePharma Co., Ltd. Specific
small interfering (si) RNAs targeting SNHG1 and negative control
(NC) siRNA were chemically synthesized by Guangzhou RiboBio Co.,
Ltd. To obtain cells with stable knockdown or overexpression of
METTL3, 3×105 HT29 and LOVO cells were transfected with
the aforementioned constructed plasmids (140 µg/ml; 2 µl; siSHNG1)
and selected with Geneticin (G418; 800 µg/ml; Procell Life Science
& Technology Co., Ltd.) for 4 weeks. METTL3 expression levels
in selected cell clones were confirmed using RT-qPCR (as per the
aforementioned method) and the constructed cell clones with stable
overexpressed or knocked down METTL3 were used for further
functional assays. To determine the effect of SNHG1 on METTL3, HT29
and LOVO cells with stable overexpression of METTL3 were
transfected with SNHG1 siRNAs at 37°C for 24 husing the RiboFECT™
Transfection Kit (166T; Guangzhou RiboBio Co., Ltd.), performed
according to the manufacturer's protocols. RT-qRCR assay was
performed 24 h after transfection to determine knockdown
efficiency, and then follow-up experiments were performed. The
sequences of siRNAs are listed in Table IB.
Methylated RNA immunoprecipitation
(MeRIP) qPCR
MeRIP-qPCR assays were performed to determine the
m6A modification level of SNHG1 as previously reported (13). All procedures were carried out
using the Magna MeRIP™ m6A Kit (cat. no. 1710499; Merck KGaA),
according to the manufacturer's instructions. In brief, total RNA
was isolated as described above and fragmented using 2 µl RNA
fragmentation buffer. The input control was one-tenth of the
isolated RNA saved. The magnetic bead A/G blend was washed three
times, resuspended and incubated with 500 µl MeRIP reaction fluid
(containing fragmental RNAs, RNase inhibitor and IP buffer) at 4°C
for 2 h. The beads were then harvested by The beads were then
harvested by Magnetic rack adsorption to remove superessence and
eluted with elution buffer (containing IP buffer, 20 mM m6A and
RNase inhibitor; from the MeRIP kit) in a vertical mixer (4°C, 5
min, 10 rpm/min). Eluted RNA fragments were harvested and purified
using the A&D Pure TRIzol Total RNA Purification Kit (A&D
Co, Ltd). Enrichment levels were determined using qPCR (as per the
aforementioned method) and the corresponding m6A enrichment level
of each sample was calculated by normalizing the input data (RNA
from target sample without m6A antibodies).
Statistical analysis
Data were analysed and evaluated using GraphPad
Prism V7.0 (GraphPad Software; Dotmatics) software and SPSS 19.0
statistical software (IBM Corp.). Differences between groups were
analysed using unpaired Student's t test or one-way ANOVA followed
by a post hoc LSD or Tukey's test. Kaplan-Meier analysis was used
to estimate overall survival using the log-rank test. P<0.05 was
considered to indicate a statistically significant difference. All
data were from three independent repeated experiments and are
expressed as the mean ± standard deviation.
Results
METTL3 is upregulated in patients with
CRC
METTL3 mRNA expression was initially analysed using
the GEO datasets GSE41258 and GSE41328. CRC tissues demonstrated
significantly higher METTL3 protein expression compared with that
in normal colonic tissues (Fig. 1A and
B). Additionally, METTL3 expression was significantly increased
in CRC tissue samples compared with that in paired adjacent
non-tumour tissue samples (Fig. 1C and
D). METTL3 protein expression was also demonstrated to be
upregulated in CRC cells (Fig.
1H). High METTL3 expression was significantly associated with
lymph node and metastasis stages and a significantly shorter
overall survival in patients with CRC, compared with those with low
METTL3 expression (Fig. 1E-G).
However, other factors were not considered. These findings
indicated that METTL3 may be a promising biomarker of CRC.
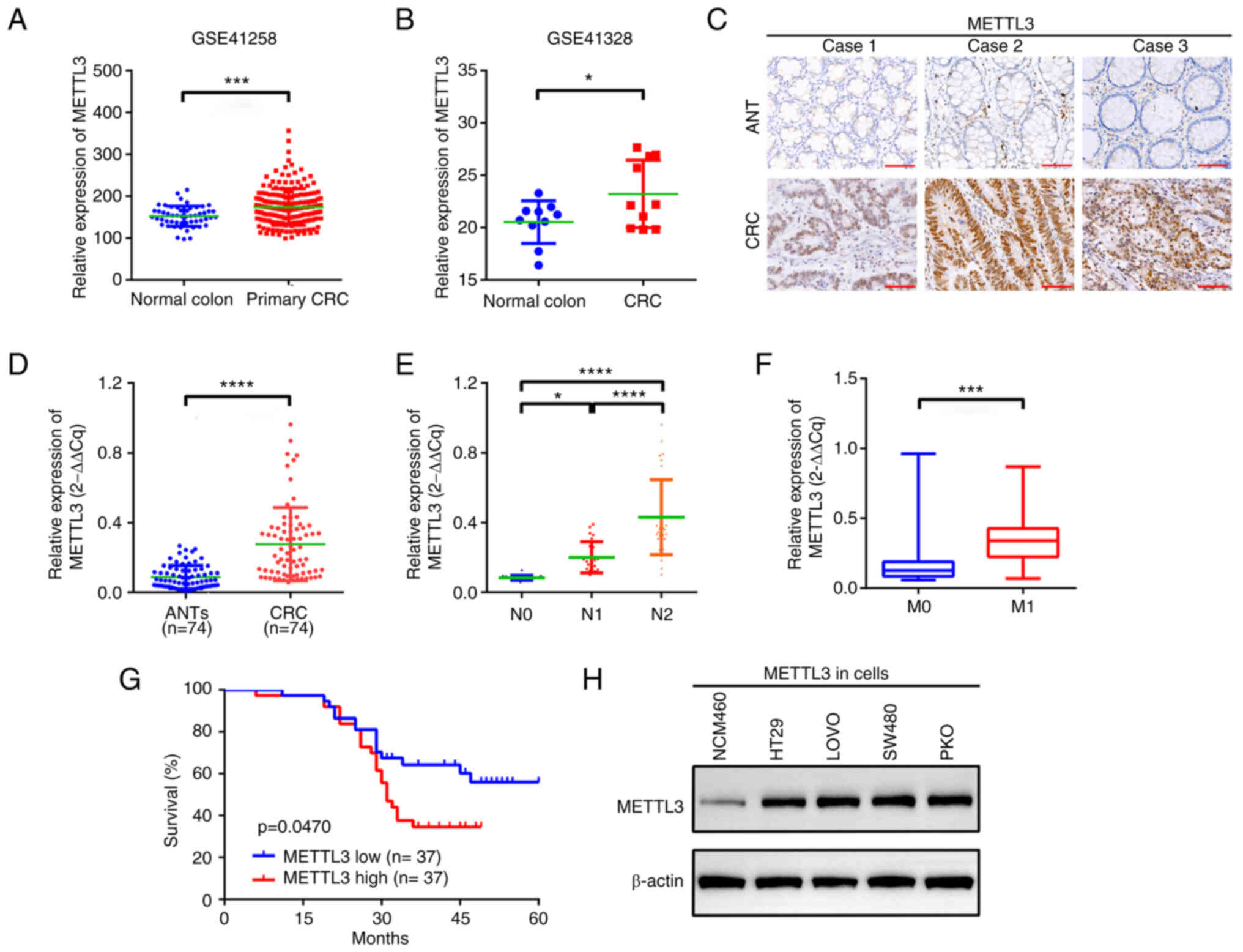 | Figure 1.METTL3 is upregulated in patients
with CRC. METTL3 expression in CRC, demonstrated by analysis of
datasets from two genome-wide studies, (A) GSE41258 and (B)
GSE41328. (C) METTL3 expression in CRC and in paired ANTs, assessed
using immunohistochemistry. Scale bar, 20 µm. (D) Differential
expression of METTL3 in CRC tissues and ANTs, evaluated using
reverse transcription-quantitative PCR. (E) The expression of
METTL3 in patients with advanced N stage compared with those with
low N stage. (F) METTL3 expression in patients with M1
stage compared with those with M0 stage. (G)
Kaplan-Meier analysis indicating overall survival of patients with
high METTL3 expression compared with that of patients with low
METTL3 expression. n=37 for each group. (H) Expression of METTL3 in
NCM460, LOVO, RKO, SW480 and HT29 cells, measured using western
blotting. *P<0.05; ***P<0.001; ****P<0.0001. METTL3,
methyltransferase-like 3; CRC, colorectal cancer; ANT, adjacent
nontumor tissue; N stage, lymph node stage; M stage; metastasis
stage. |
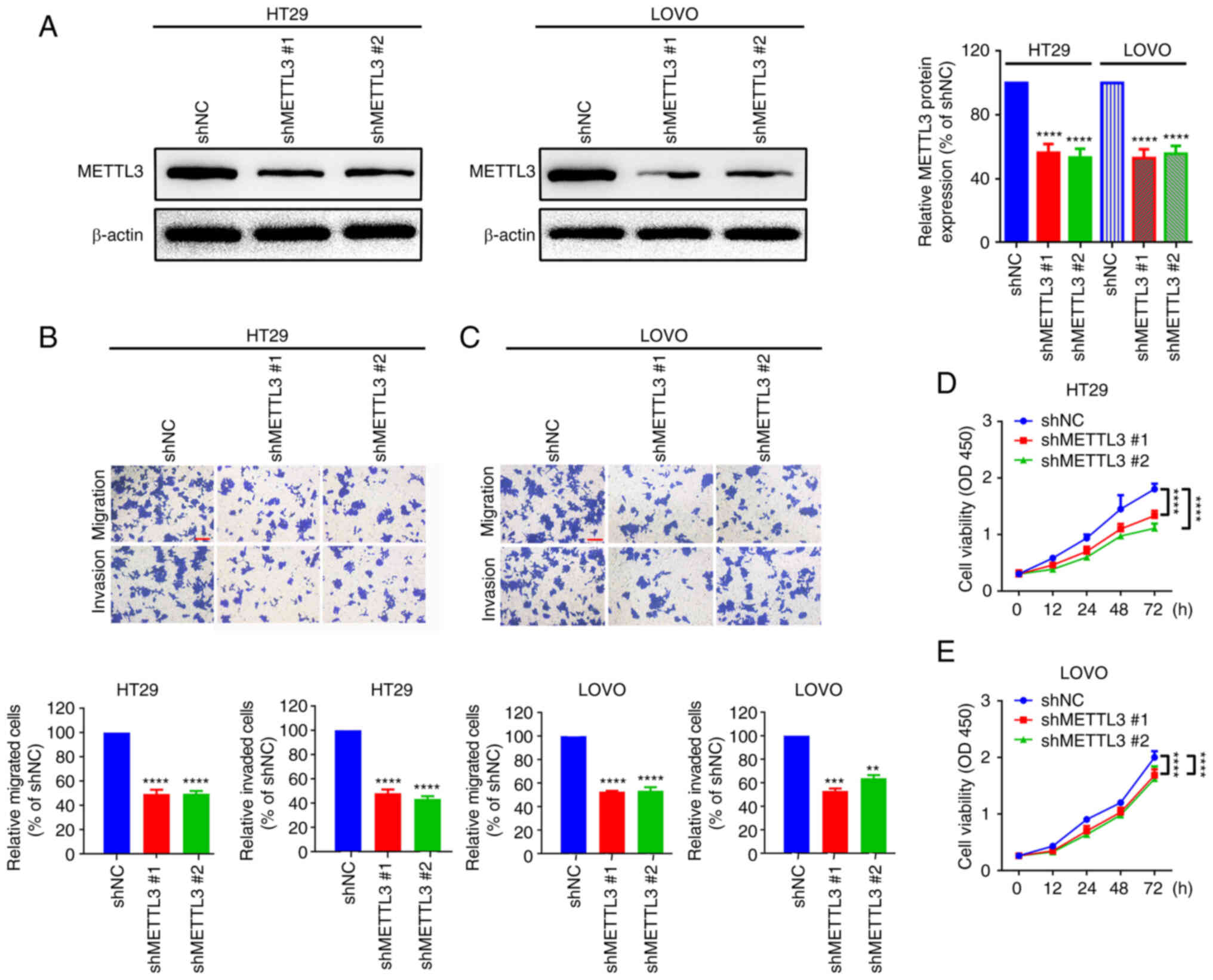
The data above showed that METTL3
knockdown inhibits CRC migration and proliferation
METTL3 shRNAs were transfected into two CRC cell
lines, HT29 and LOVO, to determine the functional role of METTL3 in
CRC cells. Western blotting demonstrated that METTL3 was
successfully knocked down in HT29 and LOVO cells (Fig. 2A). The migration of HT29 and LOVO
cells was then assessed and was demonstrated to have significantly
decreased after METTL3 knockdown, compared with that in the shNC
groups (Fig. 2B and C). Finally, a
CCK-8 assay was performed, and compared with that of the shNC
group, the proliferation of HT29 and LOVO cells was significantly
decreased when METTL3 was knocked down (Fig. 2D and E). The data showed that
METTL3 knockdown inhibits CRC cell migration and proliferation.
METTL3-mediated m6A modification is
associated with SNHG1 stability in HT29 and LOVO cells
Previous studies reported that METTL3 is closely
implicated in certain lncRNA m6A modifications (28–30).
SNHG1 is an oncogenic lncRNA in certain cancers, including CRC
(16,31,32).
Analysis of METTL3 and SNHG1 expression data from TCGA using the
online software UALCAN (22), both
METTL3 and SNHG1 were demonstrated to be notably upregulated in CRC
primary tumour samples, compared with that in normal samples
(Fig. 3A-D). Additionally, a
significant positive correlation was demonstrated between METTL3
and SNHG1 expression in CRC (Fig. 3E
and F). Moreover, SNHG1 contained 18 m6A modification sites,
demonstrated using RMBase 2.0 (23) (Table
SI) and METTL3 provided six m6A binding sites for SNHG1
(Table SII; Fig. 3G and H). The m6A level of SNHG1 in
cells with different METTL3 expression levels was then assessed and
compared with that in the shNC groups, the m6A level of SNHG1 was
significantly decreased in the METTL3 knockdown groups in both HT29
and LOVO cells (Fig. 3I). SNHG1
expression was also significantly reduced in METTL3 knockdown
groups in both HT29 and LOVO cells, compared with that in shNC
groups (Fig. 3J and K).
Furthermore, to evaluate the relationship between METTL3-mediated
m6A modification and SNHG1 upregulation, 2 µmol/l actinomycin D was
added to HT29 and LOVO cells with knocked down METTL3 and RT-qPCR
was used to assess the half-life of SNHG1. It was demonstrated that
the half-life of SNHG1 was significantly reduced in groups with
downregulated METTL3 in HT29 and LOVO cells, compared with that in
the shNC groups (Fig. 3L and M).
These results indicated that METTL3 affected the stability of SNHG1
in an m6A-dependent manner.
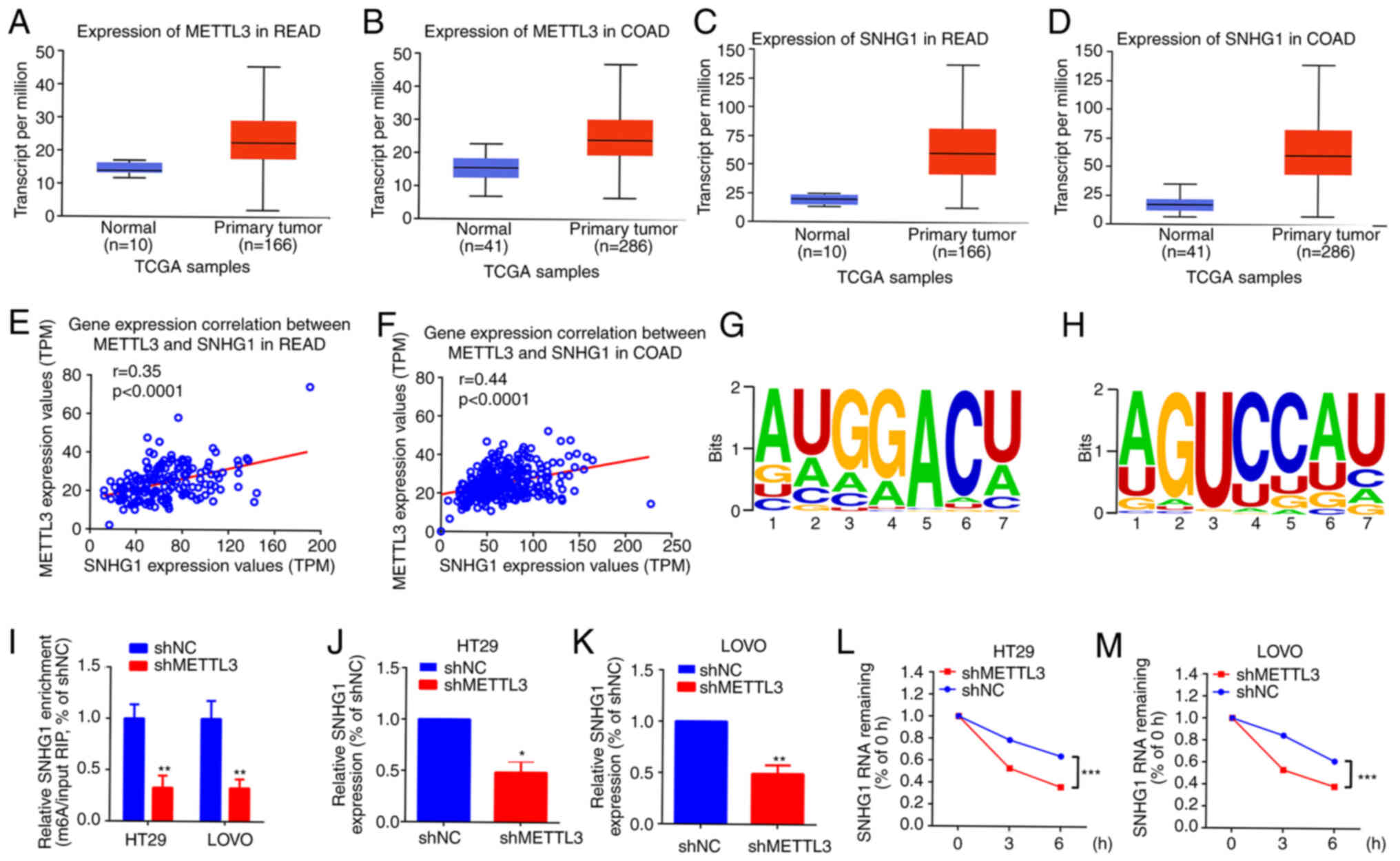 | Figure 3.METTL3-mediated m6A modification is
associated with SNHG1 upregulation in HT29 and LOVO cells. The
expression of METTL3 in (A) READ and (B) COAD and the expression of
SNHG1 in (C) READ and (D) COAD, using data from TCGA, analysed
using the online software UALCAN (http://ualcan.path.uab.edu/index.html). The
correlation between the expression of METTL3 and SNHG1 in (E) READ
r=0.35 and (F) COAD r=0.44, assessed using Spearman correlation
analysis. (G) and (H) METTL3 binding motifs in the exon region of
SNHG1, predicted using RMBase (version 2.0; http://rna.sysu.edu.cn/rmbase/). (I) m6A level of
SNHG1 in different METTL3-expressing cells, assessed using
methylated RNA immunoprecipitation quantitative PCR. The expression
of SNHG1 after METTL3 knockdown in (J) HT29 and (K) LOVO cells,
measured using RT-qPCR. The level of SNHG1 in METTL3-expressing
cells (L) HT29 and (M) LOVO after actinomycin D intervention,
quantified using RT-qPCR. *P<0.05; **P<0.01; ***P<0.001.
METTL3, methyltransferase-like 3; m6A,
N6-methyladenosine; SNHG1, small nucleolar RNA host gene
1; READ, rectal adenocarcinoma; COAD, colon adenocarcinoma; TCGA,
The Cancer Genome Atlas; UALCAN, University of ALabama at
Birmingham CANcer data analysis portal; RT-qPCR, reverse
transcription-quantitative PCR; TPM, transcripts per million; RIP,
RNA immunoprecipitation; NC, negative control; sh, short hairpin
RNA. |
SNHG1 knockdown partially attenuates
the facilitative effect of METTL3 on migration and proliferation in
HT29 and LOVO cells
SNHG1-related loss-of-function assays were performed
to further assess the relationship between SNHG1- and
METTL3-mediated promotion of cell migration and proliferation.
Initially, HT29 and LOVO cell models with stable METTL3
overexpression were constructed (Fig.
4A). Specific SNHG1 siRNAs were transfected into
METTL3-overexpressing HT29 and LOVO cells and SNHG1 expression was
quantified by RT-qPCR (Fig. S1A and
B). Overexpression of METTL3 significantly increased the level
of SNHG1 expression, compared with vector group, whilst specific
SNHG1 siRNAs significantly reduced SNHG1 expression, compared with
METTL3-siNC group (Fig. 4B and C).
Functional CCK-8 and Transwell assays were then performed to
evaluate the role of SNHG1 in METTL3-mediated cell proliferation
and cell motility. Overexpression of METTL3 significantly promoted
proliferation and migration compared with the empty vector control
group; however, this facilitative effect was significantly reversed
by knockdown of SNHG1 in HT29 and LOVO cells when compared with the
METTL3-siNC group (Fig. 4D and E).
These findings indicated that knockdown of SNHG1 partially
attenuated the facilitative effect of METTL3 on HT29 and LOVO cell
migration and proliferation.
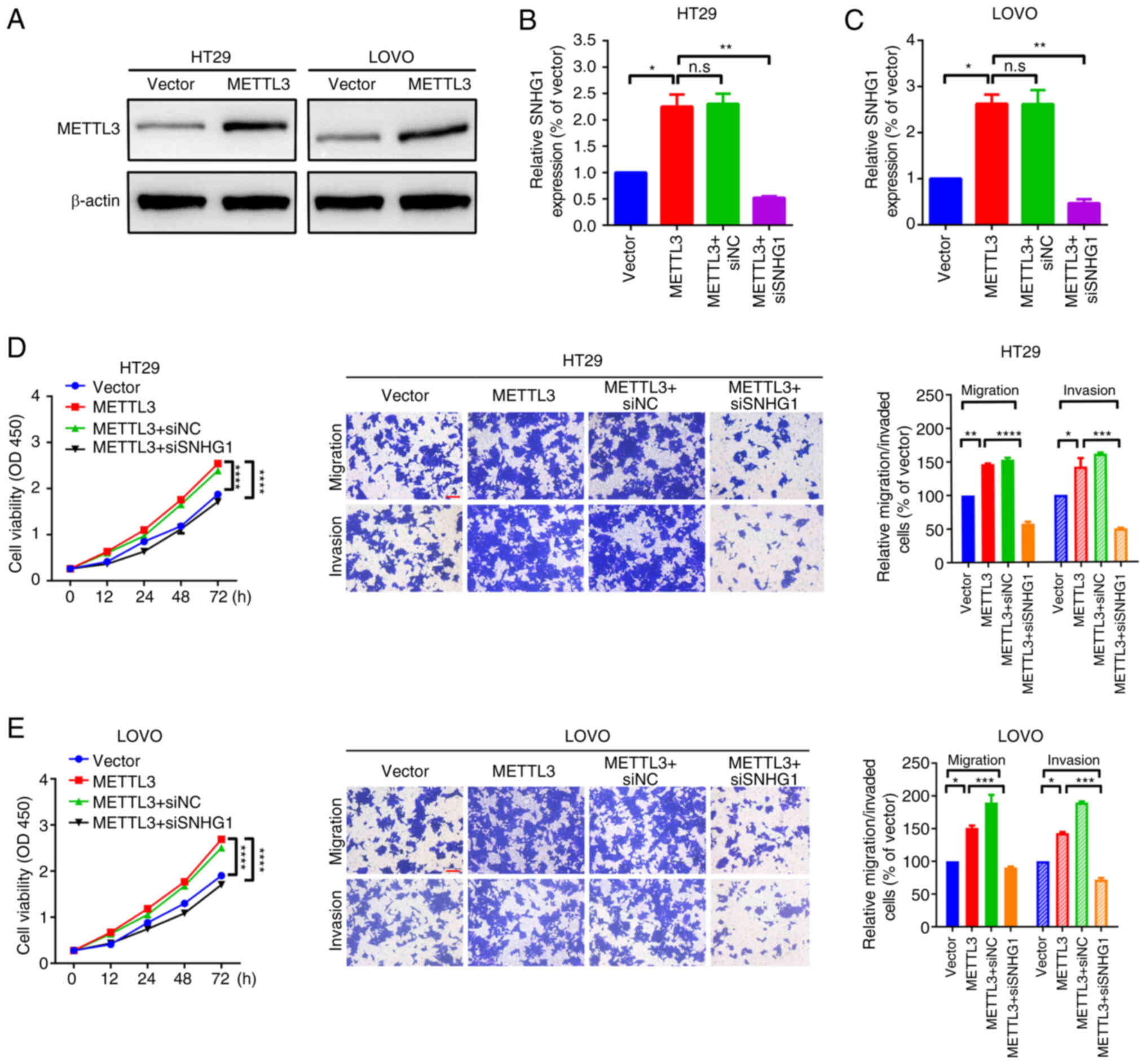 | Figure 4.Knockdown of SNHG1 partially
attenuated the facilitative effect of METTL3 on migration and
proliferation in HT29 and LOVO cells. (A) The expression of METTL3
protein in HT29 and LOVO cells, measured using western blotting.
The expression level of SNHG1 in (B) HT29 and (C) LOVO cells,
assessed using RT-qPCR. Cell proliferation, invasion and migration
of (D) HT29 and (E) LOVO cells, determined using CCK-8 assays (left
panel) and Transwell assays (right panel). Scale bar, 200 µm.
*P<0.05; **P<0.01; ***P<0.001; ****P<0.0001. SNHG1,
small nucleolar RNA host gene 1; METTL3, methyltransferase-like 3;
CCK-8; Cell Counting Kit-8; NC, negative control; si, small
interfering RNA; n.s, non-significance; OD, optical density. |
Discussion
RNA modifications are common features in epigenetic
regulation in numerous human diseases, including cancer (33). One of the most common RNA
modifications is m6A, accounting for ~50% of total methylated
ribonucleotides and 0.1-0.4% of all adenosine in total cellular RNA
(34). m6A regulates cellular
processes, including cell self-renewal, differentiation, invasion
and apoptosis, and it is also extensively implicated in neoplastic
diseases, including CRC, osteosarcoma, lung cancer and ovarian
cancer (29,35–37).
m6A methyltransferases, demethylases and reader proteins write,
remove and recognise m6A, respectively (38).
METTL3, first purified in 1994 from HeLa cell nuclei
(39), is the core component of
the m6A methyltransferase complex (MTC) and contains 580 amino
acids; it functions as the catalytic subunit in the MTC, using
adenosylmethionine as the methyl donor (9,40).
METTL3 acts as an m6A methyltransferase in numerous cancers,
including liver, gastric, lung, pancreatic, bladder, prostate and
breast cancers, CRC and acute myeloid leukaemia (10). In line with previously reported
findings, analysis of GEO datasets GSE41258 and GSE41328 and
experimental research in the present study demonstrated that METTL3
was upregulated and correlated with poor features such as lymph
node and metastasis stages, and a significantly shorter overall
survival time in patients with CRC. Functional CCK-8 and Transwell
assays demonstrated that METTL3 knockdown suppressed the migration
and proliferation of HT29 and LOVO cells, suggesting that METTL3
serves as an oncogene in CRC. The downstream targets of METTL3 in
CRC include SOX2, chromobox 8, hexokinase 2 and solute carrier
family 2 member 1 (13,41,42).
Additionally, METTL3 participates in the m6A modification of
several noncoding RNAs, including lncRNA RP11-138 J23.1,
primary-microRNA-1246 and circular RNA NOP2/Sun RNA
methyltransferase 2 (43–45). In the present study, only the
relationship between METTL3 and SNHG1 was evaluated.
SNHG1, located in the 11q12.3 region of the
chromosome, contains 11 exons and is a host to eight small
nucleolar RNAs from its spliced intron (46). As an oncogenic lncRNA, SNHG1 is
aberrantly expressed in numerous malignancies, including
colorectal, liver, lung and prostate cancer, and promotes
tumourigenesis via diverse signalling pathways (16,31).
SNHG1 upregulation promotes CRC progression through multiple
mechanisms, such as microRNA sponging, Wnt/β-catenin signal
activation and p53 pathway modulation (14,18,32,47).
Previous studies have reported that m6A modification affects the
stability of several lncRNAs, such as X inactive-specific
transcript, growth arrest-specific 5 and metastasis associated lung
adenocarcinoma transcript 1 (48–50).
In the present study, it was demonstrated that SNHG1 was positively
correlated with METTL3. Additionally, it was demonstrated that
METTL3 affected the m6A level of SNHG1 and the half-life of SNHG1.
Furthermore, it was demonstrated via online bioinformatics analysis
that SNHG1 supplied 18 m6A modification sites. Furthermore,
knockdown of SNHG1 was demonstrated to have partially attenuated
the facilitative effect of METTL3 on migration and proliferation in
CRC cells. The findings from the present study together suggest
that SNHG1 is a downstream target of METTL3 in CRC.
CRC carcinogenesis is an intricate biological
process involving numerous factors and molecules. In the present
study, it was demonstrated that METTL3 promotes the proliferation
and migration of CRC cells via SNHG1 m6A modification. The findings
of the present study therefore suggest that the inhibition of
METTL3 may provide a new therapeutic target in the molecular
treatment of CRC.
Supplementary Material
Supporting Data
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was supported by the Science and Technology
Innovation Fund for Master's Students of Shenyang Medical College
(grant no. Y20210515).
Availability of data and materials
The datasets used and/or analysed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XW conceived and designed the study, and revised the
manuscript. YX, YB and GQ performed the experiments, prepared the
figures, and designed Table I,
SI and SII. YB and GQ confirm the authenticity
of all the raw data. YX, HY and MH developed the methods and
performed data analysis. YX and MH acquired and interpreted the
data, and wrote and revised the paper. All authors have read and
approved the final version of the manuscript.
Ethics approval and consent to
participate
All experiments were approved by the Animal Ethics
and Laboratory Committee of Affiliated Central Hospital of Shenyang
Medical Science (approval no. 20220528). All individuals provided
written informed consent for the use of their tissues in the
present study
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Morgan E, Arnold M, Gini A, Lorenzoni V,
Cabasag CJ, Laversanne M, Vignat J, Ferlay J, Murphy N and Bray F:
Global burden of colorectal cancer in 2020 and 2040: Incidence and
mortality estimates from GLOBOCAN. Gut. 72:338–344. 2023.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2018. CA Cancer J Clin. 68:7–30. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
De Rosa M, Pace U, Rega D, Costabile V,
Duraturo F, Izzo P and Delrio P: Genetics, diagnosis and management
of colorectal cancer (Review). Oncol Rep. 34:1087–1096. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Manfredi S, Lepage C, Hatem C, Coatmeur O,
Faivre J and Bouvier AM: Epidemiology and management of liver
metastases from colorectal cancer. Ann Surg. 244:254–259. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Liu N, Dai Q, Zheng G, He C, Parisien M
and Pan T: N(6)-methyladenosine-dependent RNA structural switches
regulate RNA-protein interactions. Nature. 518:560–564. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wang X, Zhao BS, Roundtree IA, Lu Z, Han
D, Ma H, Weng X, Chen K, Shi H and He C: N(6)-methyladenosine
modulates messenger RNA translation efficiency. Cell.
161:1388–1399. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wang X, Lu Z, Gomez A, Hon GC, Yue Y, Han
D, Fu Y, Parisien M, Dai Q, Jia G, et al:
N6-methyladenosine-dependent regulation of messenger RNA stability.
Nature. 505:117–120. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Alarcón CR, Lee H, Goodarzi H, Halberg N
and Tavazoie SF: N6-methyladenosine marks primary microRNAs for
processing. Nature. 519:482–485. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang X, Feng J, Xue Y, Guan Z, Zhang D,
Liu Z, Gong Z, Wang Q, Huang J, Tang C, et al: Structural basis of
N(6)-adenosine methylation by the METTL3-METTL14 complex. Nature.
534:575–578. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zeng C and Huang W, Li Y and Huang W:
Roles of METTL3 in cancer: Mechanisms and therapeutic targeting. J
Hematol Oncol. 13:1172020. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yue B, Song C, Yang L, Cui R, Cheng X,
Zhang Z and Zhao G: METTL3-mediated N6-methyladenosine modification
is critical for epithelial-mesenchymal transition and metastasis of
gastric cancer. Mol Cancer. 18:1422019. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Han J, Wang JZ, Yang X, Yu H, Zhou R, Lu
HC, Yuan WB, Lu JC, Zhou ZJ, Lu Q, et al: METTL3 promote tumor
proliferation of bladder cancer by accelerating pri-miR221/222
maturation in m6A-dependent manner. Mol Cancer. 18:1102019.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Li T, Hu PS, Zuo Z, Lin JF, Li X, Wu QN,
Chen ZH, Zeng ZL, Wang F, Zheng J, et al: METTL3 facilitates tumor
progression via an m6A-IGF2BP2-dependent mechanism in
colorectal carcinoma. Mol Cancer. 18:1122019. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhu Y, Li B, Liu Z, Jiang L, Wang G, Lv M
and Li D: Up-regulation of lncRNA SNHG1 indicates poor prognosis
and promotes cell proliferation and metastasis of colorectal cancer
by activation of the Wnt/β-catenin signaling pathway. Oncotarget.
8:111715–111727. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Avazpour N, Hajjari M, Kazemi Nezhad SR
and Tahmasebi Birgani M: SNHG1 long noncoding RNA is potentially
up-regulated in colorectal adenocarcinoma. Asian Pac J Cancer Prev.
21:897–901. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Thin KZ, Tu JC and Raveendran S: Long
non-coding SNHG1 in cancer. Clin Chim Acta. 494:38–47. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Sun X, Wang Z and Yuan W: Down-regulated
long non-coding RNA SNHG1 inhibits tumor genesis of colorectal
carcinoma. Cancer Biomark. 20:67–73. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Bai J, Xu J, Zhao J and Zhang R: lncRNA
SNHG1 cooperated with miR-497/miR-195-5p to modify
epithelial-mesenchymal transition underlying colorectal cancer
exacerbation. J Cell Physiol. 235:1453–1468. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sheffer M, Bacolod MD, Zuk O, Giardina SF,
Pincas H, Barany F, Paty PB, Gerald WL, Notterman DA and Domany E:
Association of survival and disease progression with chromosomal
instability: A genomic exploration of colorectal cancer. Proc Natl
Acad Sci USA. 106:7131–7136. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lin G, He X, Ji H, Shi L, Davis RW and
Zhong S: Reproducibility Probability Score-incorporating
measurement variability across laboratories for gene selection. Nat
Biotechnol. 24:1476–1477. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Barrett T, Wilhite SE, Ledoux P,
Evangelista C, Kim IF, Tomashevsky M, Marshall KA, Phillippy KH,
Sherman PM, Holko M, et al: NCBI GEO: Archive for functional
genomics data sets-update. Nucleic Acids Res 41(Database issue).
D991–D995. 2013.PubMed/NCBI
|
|
22
|
Chandrashekar DS, Bashel B, Balasubramanya
SAH, Creighton CJ, Ponce-Rodriguez I, Chakravarthi BVSK and
Varambally S: UALCAN: A portal for facilitating tumor subgroup gene
expression and survival analyses. Neoplasia. 19:649–658. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Xuan JJ, Sun WJ, Lin PH, Zhou KR, Liu S,
Zheng LL, Qu LH and Yang JH: RMBase v2.0: Deciphering the map of
RNA modifications from epitranscriptome sequencing data. Nucleic
Acids Res 46(D1). D327–D334. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Haderk F, Olivas V and Bivona TG:
Immunohistochemistry to study YAP in human tissue samples. Methods
Mol Biol. 1893:89–95. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wang Y, Zeng X, Wang N, Zhao W, Zhang X,
Teng S, Zhang Y and Lu Z: Long noncoding RNA DANCR, working as a
competitive endogenous RNA, promotes ROCK1-mediated proliferation
and metastasis via decoying of miR-335-5p and miR-1972 in
osteosarcoma. Mol Cancer. 17:892018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Shang A, Gu C, Wang W, Wang X, Sun J, Zeng
B, Chen C, Chang W, Ping Y, Ji P, et al: Exosomal circPACRGL
promotes progression of colorectal cancer via the
miR-142-3p/miR-506-3p-TGF-β1 axis. Mol Cancer. 19:1172020.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zuo X, Chen Z, Gao W, Zhang Y, Wang J,
Wang J, Wang J, Cao M, Cai J, Wu J and Wang X: M6A-mediated
upregulation of LINC00958 increases lipogenesis and acts as a
nanotherapeutic target in hepatocellular carcinoma. J Hematol
Oncol. 13:52020. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Xue L, Li J, Lin Y, Liu D, Yang Q, Jian J
and Peng J: m6 A transferase METTL3-induced lncRNA
ABHD11-AS1 promotes the Warburg effect of non-small-cell lung
cancer. J Cell Physiol. 236:2649–2658. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Jin D, Guo J, Wu Y, Du J, Yang L, Wang X,
Di W, Hu B, An J, Kong L, et al: m6A mRNA methylation
initiated by METTL3 directly promotes YAP translation and increases
YAP activity by regulating the MALAT1-miR-1914-3p-YAP axis to
induce NSCLC drug resistance and metastasis. J Hematol Oncol.
12:1352019. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Huang L, Jiang X, Wang Z, Zhong X, Tai S
and Cui Y: Small nucleolar RNA host gene 1: A new biomarker and
therapeutic target for cancers. Pathol Res Pract. 214:1247–1252.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Fu Y, Yin Y, Peng S, Yang G, Yu Y, Guo C
and Qin Y, Zhang X, Xu W and Qin Y: Small nucleolar RNA host gene 1
promotes development and progression of colorectal cancer through
negative regulation of miR-137. Mol Carcinog. 58:2104–2117. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Barbieri I and Kouzarides T: Role of RNA
modifications in cancer. Nat Rev Cancer. 20:303–322. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Desrosiers R, Friderici K and Rottman F:
Identification of methylated nucleosides in messenger RNA from
Novikoff hepatoma cells. Proc Natl Acad Sci USA. 71:3971–3975.
1974. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Fang Z, Hu Y, Hu J, Huang Y, Zheng S and
Guo C: The crucial roles of N6-methyladenosine
(m6A) modification in the carcinogenesis and progression
of colorectal cancer. Cell Biosci. 11:722021. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wang C, Meng Y, Zhao J, Ma J, Zhao Y, Gao
R, Liu W and Zhou X: Deubiquitinase USP13 regulates glycolytic
reprogramming and progression in osteosarcoma by stabilizing
METTL3/m6A/ATG5 axis. Int J Biol Sci. 19:2289–2303.
2023. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Bi X, Lv X, Liu D, Guo H, Yao G, Wang L,
Liang X and Yang Y: METTL3-mediated maturation of miR-126-5p
promotes ovarian cancer progression via PTEN-mediated PI3K/Akt/mTOR
pathway. Cancer Gene Ther. 28:335–349. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Chen XY, Zhang J and Zhu JS: The role of
m6A RNA methylation in human cancer. Mol Cancer.
18:1032019. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Bokar JA, Rath-Shambaugh ME, Ludwiczak R,
Narayan P and Rottman F: Characterization and partial purification
of mRNA N6-adenosine methyltransferase from HeLa cell nuclei.
Internal mRNA methylation requires a multisubunit complex. J Biol
Chem. 269:17697–17704. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wang P, Doxtader KA and Nam Y: Structural
basis for cooperative function of Mettl3 and Mettl14
methyltransferases. Mol Cell. 63:306–317. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Shen C, Xuan B, Yan T, Ma Y, Xu P, Tian X,
Zhang X, Cao Y, Ma D, Zhu X, et al: m6A-dependent
glycolysis enhances colorectal cancer progression. Mol Cancer.
19:722020. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Zhang Y, Kang M, Zhang B, Meng F, Song J,
Kaneko H, Shimamoto F and Tang B: m6A
modification-mediated CBX8 induction regulates stemness and
chemosensitivity of colon cancer via upregulation of LGR5. Mol
Cancer. 18:1852019. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Peng W, Li J, Chen R, Gu Q, Yang P, Qian
W, Ji D, Wang Q, Zhang Z, Tang J and Sun Y: Upregulated METTL3
promotes metastasis of colorectal Cancer via miR-1246/SPRED2/MAPK
signaling pathway. J Exp Clin Cancer Res. 38:3932019. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Chen RX, Chen X, Xia LP, Zhang JX, Pan ZZ,
Ma XD, Han K, Chen JW, Judde JG, Deas O, et al:
N6-methyladenosine modification of circNSUN2 facilitates
cytoplasmic export and stabilizes HMGA2 to promote colorectal liver
metastasis. Nat Commun. 10:46952019. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Wu Y, Yang X, Chen Z, Tian L, Jiang G,
Chen F, Li J, An P, Lu L, Luo N, et al: m6A-induced
lncRNA RP11 triggers the dissemination of colorectal cancer cells
via upregulation of Zeb1. Mol Cancer. 18:872019. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Tycowski KT, Shu MD and Steitz JA:
Requirement for intron-encoded U22 small nucleolar RNA in 18S
ribosomal RNA maturation. Science. 266:1558–1561. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Zhao Y, Qin ZS, Feng Y, Tang XJ, Zhang T
and Yang L: Long non-coding RNA (lncRNA) small nucleolar RNA host
gene 1 (SNHG1) promote cell proliferation in colorectal cancer by
affecting P53. Eur Rev Med Pharmacol Sci. 22:976–984.
2018.PubMed/NCBI
|
|
48
|
Patil DP, Chen CK, Pickering BF, Chow A,
Jackson C, Guttman M and Jaffrey SR: m(6)A RNA methylation promotes
XIST-mediated transcriptional repression. Nature. 537:369–373.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Ni W, Yao S, Zhou Y, Liu Y, Huang P, Zhou
A, Liu J, Che L and Li J: Long noncoding RNA GAS5 inhibits
progression of colorectal cancer by interacting with and triggering
YAP phosphorylation and degradation and is negatively regulated by
the m6A reader YTHDF3. Mol Cancer. 18:1432019.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Warda AS, Kretschmer J, Hackert P, Lenz C,
Urlaub H, Höbartner C, Sloan KE and Bohnsack MT: Human METTL16 is a
N6-methyladenosine (m6A) methyltransferase
that targets pre-mRNAs and various non-coding RNAs. EMBO Rep.
18:2004–2014. 2017. View Article : Google Scholar : PubMed/NCBI
|