Introduction
Liver cancer is a common malignant tumor, which due
to its high metastatic potential, has become the primary cause of
cancer-related death (1,2). although great progress has been made in
the diagnosis and treatment of liver cancer, patient prognosis is
still unsatisfactory (3–5). Thus, exploring its potential
pathological mechanism is particularly important to find new
strategies that can improve treatment of patients with liver
cancer.
X-linked ubiquitin-specific peptidase 9 (USP9X) is a
member of the deubiquitinase family that has been shown to be
involved in a variety of biological pathways, such as regulating
cell transformation and survivalf6,7). Biochemical studies have
identified that USP9X is widely expressed in tissues, which has
been confirmed in TCGA database, and it can regulate the protein
function by the deubiquitination pathway (8–14). USP9X
plays important roles in the development and progression of several
human cancers (15). For example,
the USP9X protein is significantly increased in non-small cell lung
cancer, breast cancer, leukemia, cervical cancer, follicular
lymphoma, colon cancer and esophageal squamous cell carcinoma
(8–14). Overexpression of USP9X in cancer
activates multiple important pathways, such as the
Ras/mitogen-activated protein kinase, PI3K/AKT, Rho/ Rho-associated
protein kinase, Jagged/Notch, NK-κB and Wnt/β-catenin pathways
(16). In addition, USP9X plays an
important role in promoting liver cancer cell proliferation, but
low USP9X expression can inhibit the bioactivity of these cells
(17). This indicates that the
protein stability of USP9X plays an important role in regulating
the proliferation and cell activity of liver cancer cells. However,
the molecular mechanism by which USP9X regulates the proliferation,
invasiveness and migration of liver cancer cells is remains to be
clarified.
The Janus kinase 2 (JAK2)/STAT3 pathway plays an
important part in several cellular activities, including processes
involved with the cytoplasmic domains of several cytokine receptors
and mediating signals triggered by several hematopoietic growth
factors, including erythropoietin (Epo), thrombopoietin (Tpo), and
granulocyte colony-stimulating factor (G-CSF) (18,19).
This pathway can regulate cytokines, adhesion molecules and other
inflammatory mediators (18,20,21), and
transduce signals from activated receptors or intracellular kinases
to the nucleus to activate and regulate gene transcription
(22). JAK2/STAT3 signaling also
serves a critical role in tumor cell proliferation, invasiveness
and migration (23,24), where the oncogene STAT3 is a key
factor (25). Several studies have
shown that STAT3 activation can improve the expression of c-Myc,
which promotes cellular proliferation (26). On the other hand, the STAT3
activation is associated with the upregulation of matrix
metalloproteinase-2 (MMP2), and further promote tumor invasive
growth (27–31).
In the present study, the expression of USP9X in
liver cancer tissues, and its role in the proliferation,
invasiveness and migration of liver cancer cells were investigated
to provide novel insight into the role of USP9X in liver
cancer.
Materials and methods
Tissue samples
A total of 14 patient samples were collected from
seven liver cancer patients and seven patients without liver cancer
at the No. 1 People's Hospital of Xuzhou, (Xuzhou, China). Part of
the surgically removed tissues were used for histological
diagnosis, and remainder was immediately frozen in liquid nitrogen
and stored at −80°C. All specimens were pathologically diagnosed.
The present study was approved by the Research Ethics Committee of
the No. 1 People's Hospital of Xuzhou and written informed consent
was obtained from each patient.
Cell culture
The 293T cell line and the HepG2 liver cancer cell
line were purchased from The Cell Bank of Type Culture Collection
of the Chinese Academy of Sciences, Shanghai. All cells were
identified by STR profiling. Cells were cultured in DMEM (Hyclone;
Cytiva) supplemented with 10% fetal bovine serum (FBS; Invigentech)
in a 5% CO2 incubator at 37°C. The inhibitor of JAK2
(Fedratinib, http://www.medchemexpress.cn/tg-101348.html?src=bd-product)
was added 24 h post-transfection at a concentration of 10 µg/ml,
and the cells were collected 6 h later.
Lentivirus production
The short hairpin (sh) RNA USP9X plasmid and the
associated shControl (which contained a non-targeting oligomers and
doesn't affect the wild-type USP9X) were purchased from Shanghai
GenePharma Co., Ltd. shUSP9X sequence:
5′-CCGGCCACCTCAAACCAAGGATCAACTCGAGTTGATCCTTGGTTTGAGGTGGTTTTT-3′;
shControl sequence: 5′-GATCTTCTCCGAACGTGT-
CACGTTTCAAGAGAACGTGACACGTTCGGAGAAT TTTTTG-3′. Transfection was
performed with PolyJet transfection reagent (SignaGen Laboratories)
according to the manufacturer's instructions. The lentiviruses were
produced by co-transfecting 293T cells with both the core and the
packaging plasmids. The lentivirus was collected after incubation
at 37°C for 72 h.
Establishment of stable cell
lines
The plasmids of control and shUSP9X were GFP-tagged.
Moderate amounts (1 ml) of lentiviruses were used to infect HepG2
cells (5×105 cells/well in 6-well plate) and then
cultured at 37°C for 72 h. Cells were continuously cultured in the
DMEM medium containing 2.5 µg/ml puromycin (Sigma Aldrich; Merck
KGaA). The surviving cells were cultured into cell lines stably
expressing control and shUSP9X. Cell lines stably expressing the
control or shUSP9X constructs were confirmed by western blot
analysis.
Transient overexpression of USP9X
The Myc-USP9X was generated by cloning the human
USP9X cDNA into the expression vector pcDNA3.1-Myc at the
BamHI and XhoI restriction sites (Youbio, http://www.youbio.cn). An empty carrier containing the
Myc tag (Myc-Vector) was used as a negative control (Youbio). The
final concentration of all plasmids was ~1 mg/ml. Transfection of
USP9X was performed using PolyJet transfection reagent (SignaGen
Laboratories, http://signagen.com) according to the
manufacturer's instructions and then incubated at 37°C for 24 h.
Subsequent experiments were performed 24 h post-transfection. The
efficiency of overexpression was verified by western blot
analysis.
5-ethynyl-20-deoxyuridine (EdU)
incorporation assay
The effect of USP9X on the proliferation of HepG2
cells was measured by EdU incorporation using an EdU assay kit
(Guangzhou RiboBio Co., Ltd.) according to the manufacturer's
instructions. Briefly, a total of 8×104 cells/well were
cultured in 48-well plates for 24 h, followed by a 2-h incubation
with 50 µmol/l EdU at 37°C. Cells were then washed with PBS and
fixed with 4% paraformaldehyde for 30 min, followed by
permeabilization with 0.5% Triton X-100 for a further 30 min.
Afterwards, the cells were washed five times with PBS and incubated
with 100 µl 1X Apollo® reaction cocktail (apart of the
EdU kit) for 30 min at room temperature. Finally, the cell nuclei
were dyed with 100 μl of Hoechst solution (5 µg/ml) for 30 min and
visualized with a fluorescent microscope (IX71; Olympus,
Corporation). The results are analyzed through Image J v1.8.0.
Cell viability assay
Cell viability was assessed with the Cell Counting
Kit-8 (CCK-8) assay (Dojindo Molecular Technologies, Inc.). A total
of 1×104 cells/well were cultured in 96-well plates and
at the designated time point, 10 µl of CCK-8 reagent was added into
the medium. After 4 h at 37°C, the absorbances were measured at 450
nm using a Synergy Mx Multi-Mode Microplate Reader. The cell
viability was calculated according to the absorbances. The
absorbances were normalized against a DMSO control.
Colony formation assay
A total of 300 cells were seeded into 6-cm dishes
and cultured for 15 days at 37°C. Cells were fixed with methanol at
room temperature for 1 h and stained with 0.1% crystal violet at
room temperature for 30 min to assess the number of colonies. Cells
were washed twice with PBS and the plate images were captured using
a digital camera (Alpha 7 II, SONY, http://www.sonystyle.com.cn/products/ilc/ilce_7m2/ilce_7m2.html).
Positive colony formation, defined as colonies with >50 cells,
was confirmed by manual counting.
Transwell invasion assay
Transwell assays were performed with a polycarbonate
filter membrane with a diameter of 6.5 mm and pore size of 8 µm
(Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol. To assess invasion ability, the filters
were pre-coated with 10 µg Matrigel. A cell suspension of
3×104 cells (in each group) in serum-free culture media
was added into the inserts, and each insert was placed into the
lower chamber filled with culture media containing 10% FBS as a
chemoattractant. After 48 h in culture at 37°C, the non-invasive
cells were removed from the upper chamber with cotton-tipped swabs,
and the filters were fixed with methanol for 30 min and stained
with a 0.1% crystal violet solution for a further 30 min at room
temperature. Finally, five fields of adherent cells in each well
were randomly selected and counted with an inverted microscope and
counted.
Wound healing assay
Cells (5×106) cultured in DMEM
supplemented with 10% fetal bovine serum were seeded into 6-well
plates and incubated in a 5% CO2 incubator at 37°C for
12 h. Then, scratches were made with a pipette tip in the middle of
the wells, and the cells were washed with PBS to remove cell debris
before incubation in serum-free media for 50 h. At the designated
time, five randomly selected fields were imaged with an inverted
microscope. The number of cells across the wound was normalized to
that of the control group. The results were analyzed using Image J
v1.8.0 (National Institutes of Health).
Western blotting
At the designated times, the cells were lysed using
RIPA lysis buffer (Beyotime Institute of Biotechnology). The
protein concentration was determined using the BCA kit (cat. no.
P0010; Beyotime Institute of Biotechnology), according to the
manufacturer's protocol. Equal amounts of protein lysate (80 µg)
were resolved using 10% SDS-PAGE. The proteins were then
transferred onto PVDF membranes and blocked with 3% BSA at room
temperature for 2 h. Membrane were incubated with primary
antibodies at 4°C overnight, and subsequently incubated with HRP
goat anti mouse/rabbit IgG secondar antibody (1:4,000; cat. nos.
sa00001-1 and sa00001-2; ProteinTech Group, Inc.) at room
temperature for 2 h. 1X TBST was used for washing. The antibodies
used were against: USP9X [1:1,000; cat. no. 14898; Cell Signaling
Technology (CST), Inc.], phosphorylated (Tyr1007/1008)-JAK2
(1:1,000; cat. no. 3771, CST, Inc.), p-STAT3 (Tyr705; 1:1,000; cat.
no. 9145; CST, Inc.), JAK2 (1:1,000; cat. no. 17670-1-AP;
ProteinTech Group, Inc.), STAT3 (1:1,000, 10253-2-AP, ProteinTech
Group, Inc.), MMP2 (1:1,000; cat. no. 10373-2-AP; ProteinTech
Group, Inc.), c-Myc (1:1,000; cat. no. 10828-1-AP; ProteinTech
Group, Inc.), β-actin (1:5,000; cat. no. 60008-1-Ig; ProteinTech
Group, Inc.) and Myc-tag (1:1,000; cat. no. 16286-1-AP; ProteinTech
Group, Inc.). Bound antibodies were detected with ECL Plus Western
Blotting Substrate (Pierce; Thermo Fisher Scientific, Inc.) and
exposed to X-ray films. Band densities were quantified using ImageJ
software (version 1.8.0; National Institutes of Health). The
relative amount of protein was determined by normalizing the
densitometry value of interest to that of the loading control.
Relative protein expression was determined by normalizing the
densitometry value of target protein bands and corresponding
internal parameters.
Immunohistochemistry
Immunohistochemical staining was performed using the
procedures provided by the S-P immunohistochemical kit (OriGene
Technologies, Inc.). The sections were fixed with 4%
paraformaldehyde for 3 days and sealed with 10% goat serum (Vicmed,
http://www.vicmed.cn/a/dbyjxgcp/myzhxg/399.html) for
30 min at room temperature. The sections were then incubated with
USP9X antibody (1:100; cat. no. 14898; CST, Inc.) at 4°C overnight,
followed by incubation with biotin-conjugated goat anti-rabbit IgG
(original concentration; cat. no. PV-9001) and HRP-conjugated
streptavidin (1:20; cat. no. ZLI-9018; both from Beijing Zhongshan
Golden Bridge Biotechnology Co., Ltd.). The reaction was developed
using 3,3′diaminobenzidine reagent (OriGene Technologies, Inc.).
Sections were stained with hematoxylin to stain the nuclei for 2
min and dehydrated by incubation in increasing concentrations of
ethanol (50, 75, 80, 90 and 100%) followed by 100% xylene for 5
min. All steps were performed at room temperature. The slides were
observed under the Olympus BX-53 light microscope (magnification,
×40; Olympus, Corporation), and images processed using Image-Pro
Plus software (version 6.0; Media Cybernetics, Inc.).
Survival analysis
The association between USP9X and survival of
patients with liver cancer was assessed using the GEPIA database
(http://gepia.cancer-pku.cn/) in order to
understand the clinical significance of USP9X.
Statistical analysis
The results are representative of ≥3 experimental
repeats. All quantitative data are presented as the mean ± SEM.
Statistical analysis was performed using SPSS (version 16.0; SPSS,
Inc.). One-way analysis of variance followed by Tukey's post hoc
test were used to compare differences between multiple groups.
Paired Student's t-test was used to compare differences between two
groups. P<0.05 was considered to indicate a statistically
significant difference.
Results
USP9X is highly expressed in human
liver cancer tissues
To understand the clinical significance of USP9X,
the association between USP9X protein expression levels and the
survival of patients with liver cancer was analyzed using The
Cancer Genome Atlas database. The results show that high levels of
USP9X is associated with poor prognosis in patients with liver
cancer (Fig. 1A). The USP9X protein
expression level of seven non-tumorous liver tissues and seven
liver cancer tissues were also examined by western blot analysis.
The results indicate that USP9X is expressed to a greater degree in
liver cancer tissues than non-tumorous tissues (Fig. 1B). These results were also consistent
with immunohistochemical analysis (Fig.
1C and D). These results revealed that USP9X may play an
important role in the development of liver cancer.
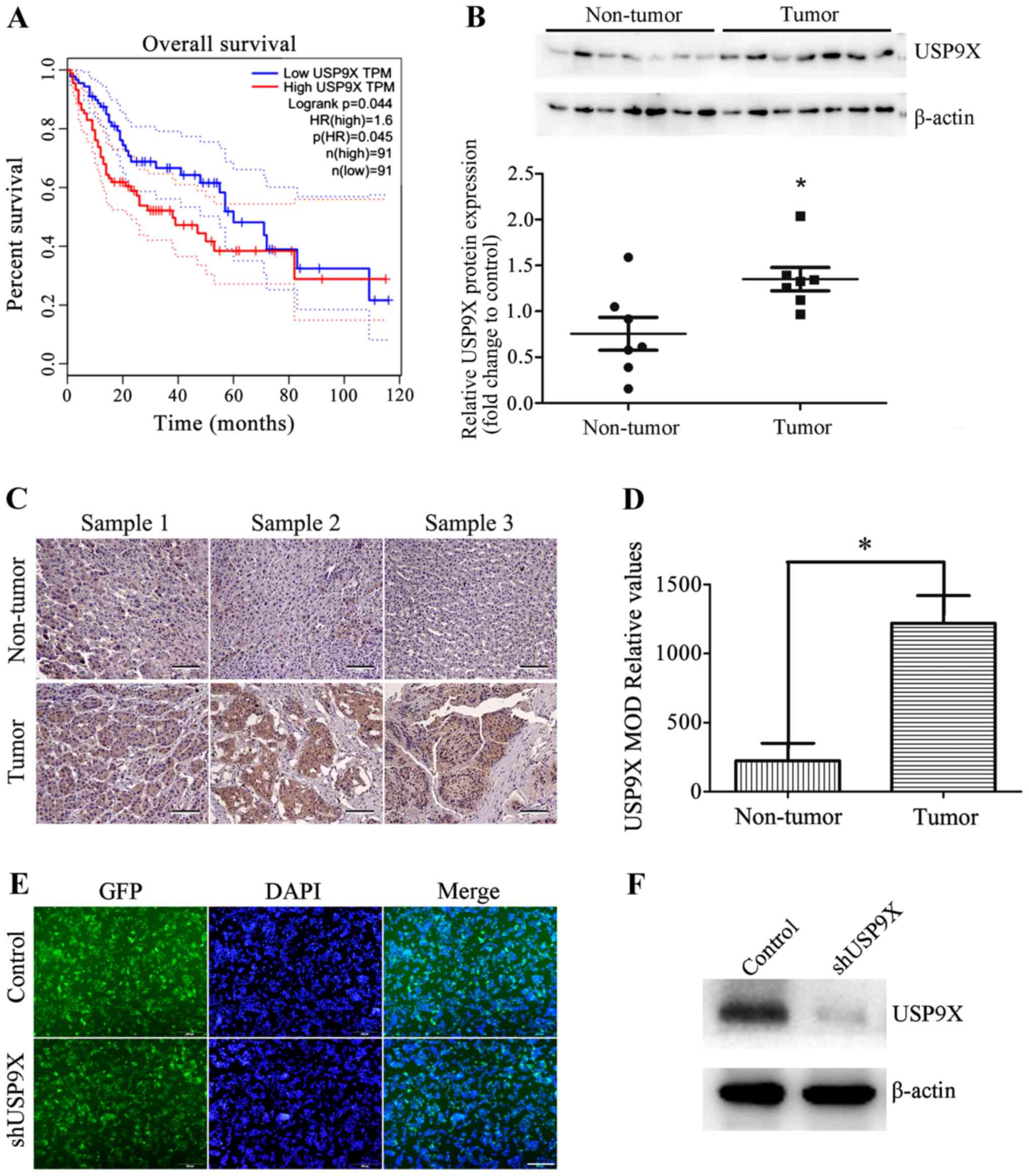 | Figure 1.Expression of USP9X in patients with
liver cancer. (A) The Cancer Genome Atlas database analysis showed
that high expression levels of USP9X were associated with poor
prognosis in patients with liver cancer. (B) Representative western
blots and quantification data show that the protein expression
levels of USP9X were higher in liver cancer tissues than in
non-tumor liver tissues. (C) Representative images of USP9X
expression innormal liver tissues and liver cancer tissues
determined by immunohistochemistry. Scale bar, 100 µm. (D)
Histogram showing the quantification of the relative levels of
USP9X from immunohistochemistry analysis. (E) GFP images show that
shUSP9X and the control were successfully transfected into HepG2
cells, generating stable USP9X-knockdown cell lines. Scale bar, 500
µm. (F) Silencing efficiency of shUSP9X in HepG2 cells was examined
by western blot. *P<0.05. GFP, green-fluorescent protein; MOD,
mean optical density; sh, short hairpin; USP9X, X-linked
ubiquitin-specific peptidase 9; TPM, total productive maintenance;
p(HR), proportion of high resistance; n, number. |
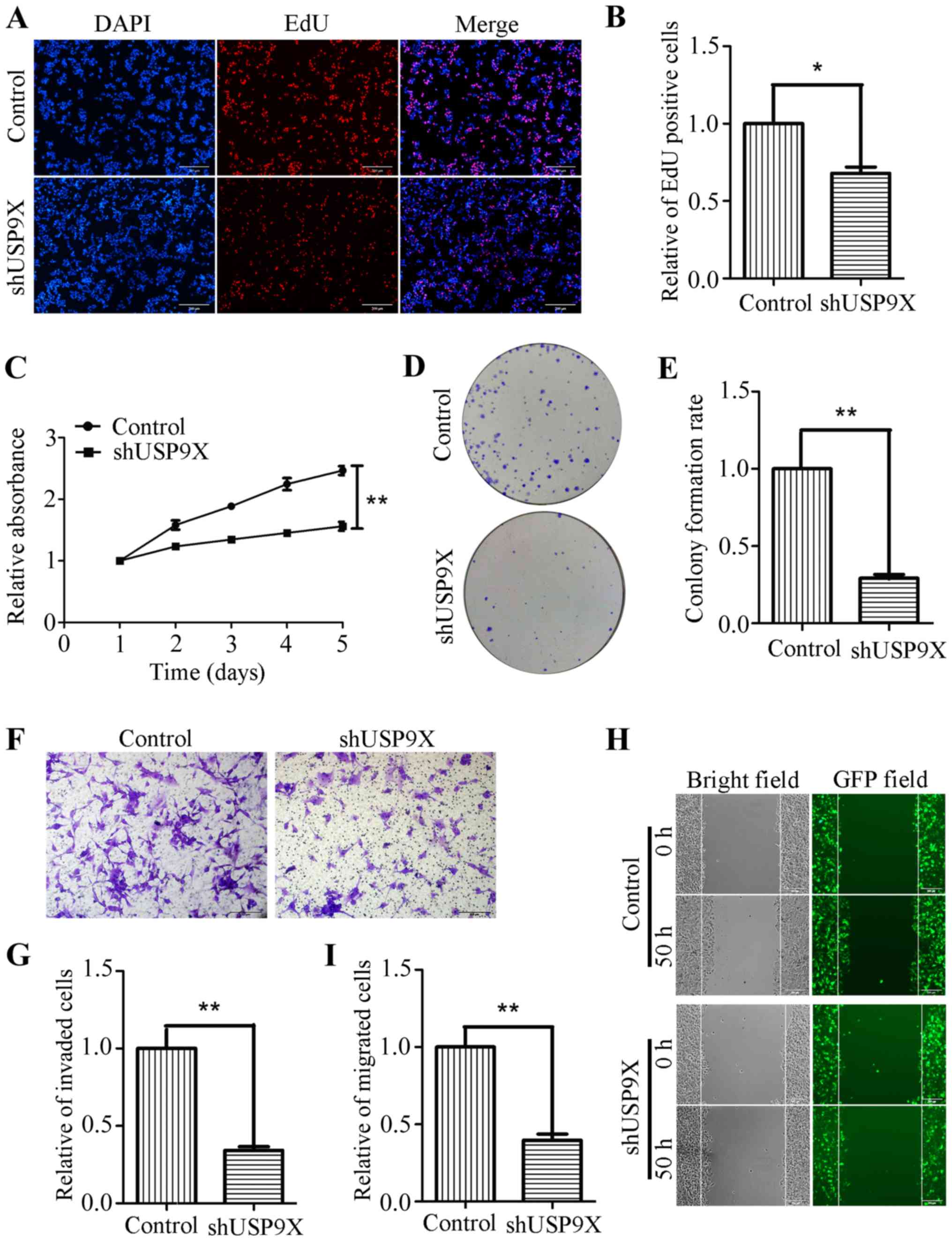
Silencing of USP9X inhibits the
proliferation, invasiveness and migration of human liver cancer
cells
Due to the increased expression of USP9X in liver
cancer tissues, it was hypothesized that USP9X might be associated
the proliferation, invasiveness and migration of liver cancer
cells. To address this problem, USP9X expression was silenced using
shRNA. shUSP9X and the respective control lentiviruses were
packaged into 293T cells before being transfection into the HepG2
cell line to successfully generate stable cell lines with loss of
USP9X (Fig. 1E and F). To determine
whether USP9X-knockdown could affect the proliferation, invasion
and migration of liver cancer cells, a number of functional
experiments were performed. The EdU incorporation assay, showed
that the number of EdU-positive cells was markedly reduced in HepG2
cells transfected with shUSP9X, compared with the control (Fig. 2A and B). In addition, the CCK-8 assay
revealed that the proliferation rate of the shUSP9X group was also
significantly decreased compared with the control (Fig. 2C). The ability of HepG2 cells to form
colonies was also significantly decreased upon silencing of USP9X
compared to the control group (Fig. 2D
and E). Furthermore, Transwell assays (with Matrigel coating)
showed that cell invasiveness was inhibited in HepG2 cells
transfected with shUSP9X (Fig. 2F and
G). The wound healing assay indicated that, compared with the
control group, the number of migrated cells at 50 h was
significantly decreased in shUSP9X HepG2 cells compared to the
control group (Fig. 2H and I). These
results show that silencing of USP9X significantly inhibits the
proliferation, invasiveness and migration of liver cancer
cells.
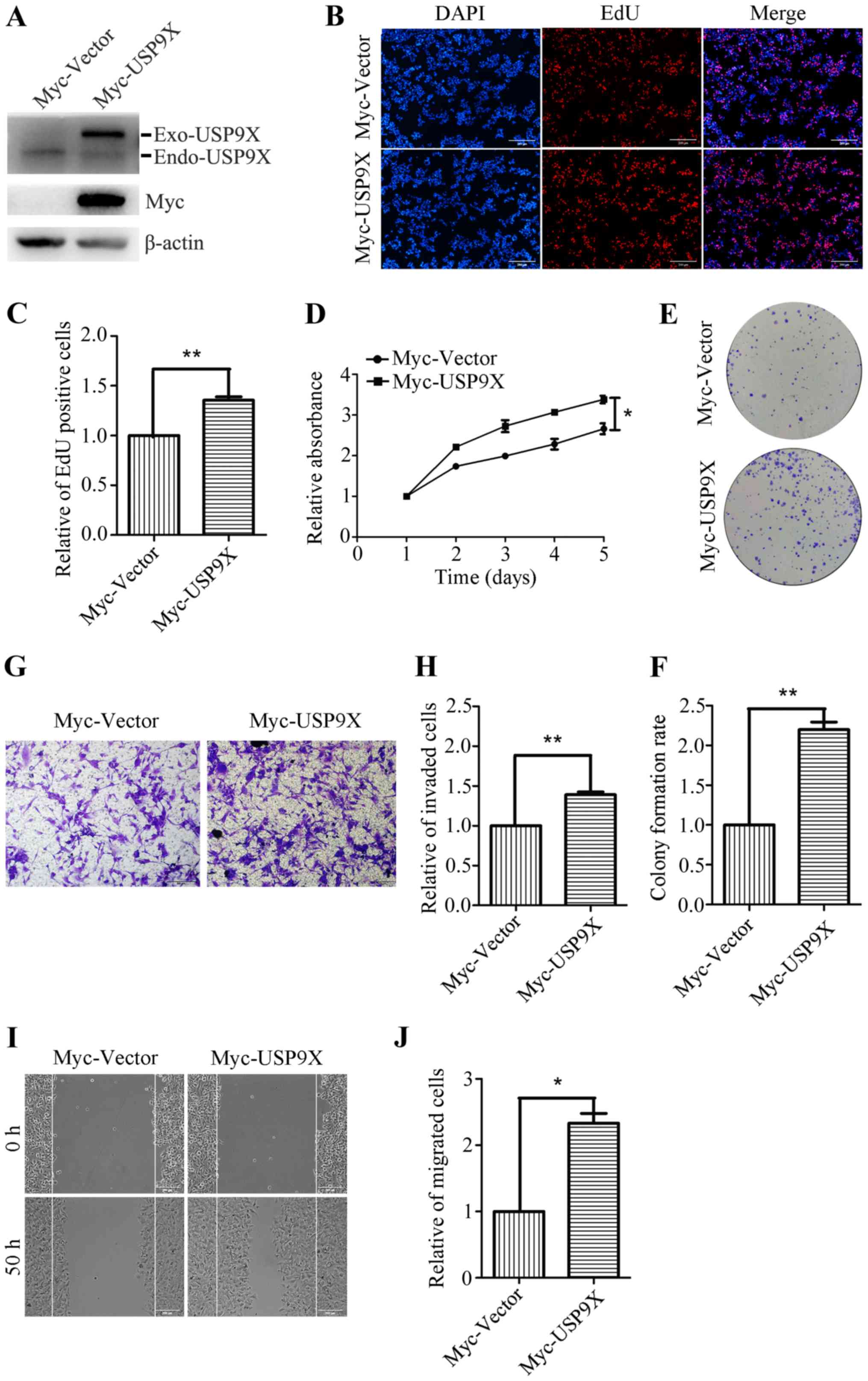
Overexpression of USP9X promotes the
proliferation, invasion and migration of human liver cancer
cells
To further investigate the role of USP9X in the
proliferation and metastatic potential of human liver cancer cells,
HepG2 cells were transiently transfected with Myc-Vector and
Myc-USP9X plasmids. Transfection efficiency was confirmed by
western blotting, and the Myc-tag antibody was used to detect the
exogenous USP9X. (Fig. 3A). After 24
h of transfection, the effects of overexpression of USP9X on cell
proliferation and metastasis were investigated. The EdU
incorporation assay showed that the number of EdU positive cells
was significantly increased in the USP9X overexpressing cells
(Fig. 3B and C). The CCK-8 assay
also revealed that the proliferation rate of the USP9X
overexpressing cells was increased compared with the control
(Fig. 3D). Moreover, the ability of
USP9X overexpressing cells to form colonies was also significantly
increased compared with the control cells (Fig. 3E and F). The Transwell assay (with
Matrigel coating) showed that the number of invasive cells was
increased by ~40% in USP9X overexpressing cells (Fig. 3G and H). Furthermore, the wound
healing assay indicated that the number of migratory cells was
notably increased upon Myc-USP9X transfection, compared with the
control (Fig. 3I and J). These
results reveal that USP9X overexpression significantly promotes the
proliferation, invasiveness and migration of liver cancer
cells.
USP9X positively regulates the
JAK2/STAT3 pathway to promote the proliferation, invasiveness and
migration of liver cancer cells
The aforementioned results have shown that USP9X is
important for the proliferation, invasivenessand migration of liver
cancer cells. However, the underlying mechanisms were yet to be
addressed. By western blot analysis, USP9X silencing was found to
block downstream effectors of the JAK2/STAT3 pathway, thereby
inhibiting the expression of c-Myc and MMP2 (Fig. 4A and B). By contrast, overexpression
of USP9X significantly upregulated the protein expression levels of
the JAK2/STAT3 signaling pathway thus increasing the expression of
c-Myc and MMP2. The relative ratios of the experimental group and
the control group to their respective internal parameters were used
for statistical analysis (Fig. 4A and
C). When evaluating the effect of the JAK2 inhibitor Fedratinib
on USP9X overexpressing cells, it was found that JAK2 inhibition
was able to effectively rescue the expression of JAK2/STAT3 pathway
components, c-Myc and MMP2 (Fig. 4D and
E), and also restore the cellular proliferation, invasiveness
and migration induced by USP9X overexpression (Fig. 4F-K). In conclusion, these results
suggest that USP9X positively regulates the JAK2/STAT3 pathway,
thereby promoting the proliferation, invasiveness and migration of
liver cancer cells.
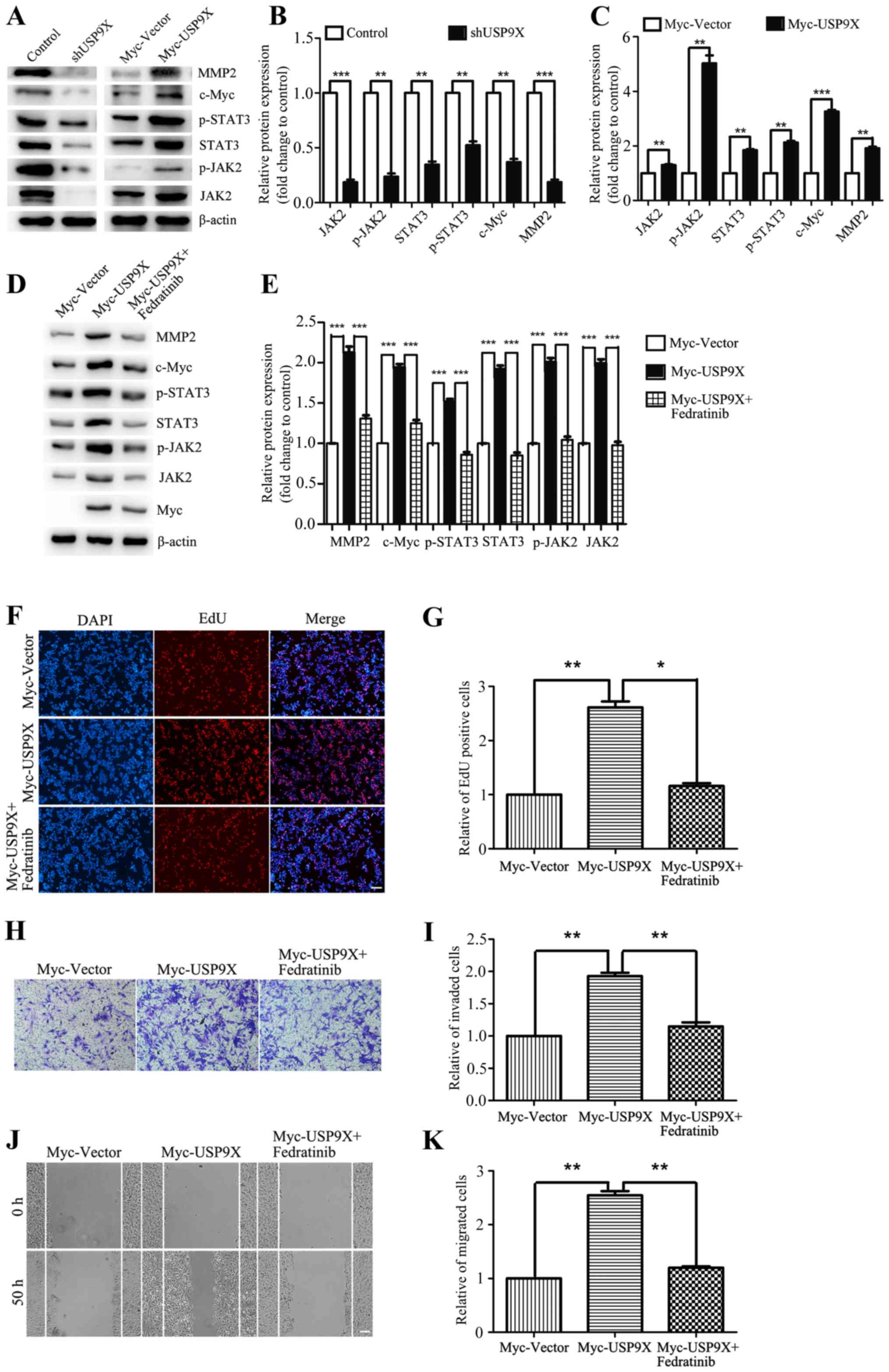 | Figure 4.USP9X positively regulates the
JAK2/STAT3 signaling pathway. (A-C) Western blot assay and
quantification showed that in HepG2 cells, the protein levels of
JAK2/STAT3 pathway components, c-Myc and MMP2 were reduced in the
context of USP9X loss by shRNA, while the results were contrary
when USP9X increased in expression. (D and E) Western blot assay
showed that the JAK2 inhibitor Fedratinib effectively rescued the
expression of JAK2/STAT3 pathway components, c-Myc and MMP2. The
inhibitor was added 24 h after transfection at a concentration of
10 µg/ml, and the cells were collected 6 h later. (F-K) EdU
incorporation assay, Transwell invasion assay and wound healing
assay showed that Fedratinib effectively restored pcell
roliferation, invasiveness and migration induced by overexpressing
USP9X. *P<0.05, **P<0.01 and ***P<0.001. EdU,
5-ethynyl-20-deoxyuridine; JAK2, Janus kinase 2; MMP2, matrix
metalloproteinase-2; p-, phosphorylated; USP9X,
X-linkedubiquitin-specific peptidase 9. |
Discussion
USP9X is involved in the regulation of numerous key
biological processes, such as proteasome activity, organ
production, tumor production and transcriptional regulation
(6,7). It can simultaneously affect the
ubiquitination process to regulate apoptosis, proliferation and
adhesion of cells (8). Previous
research has found that USP9X is significantly increased in human
non-small cell lung cancer, breast cancer, leukemia, cervical
cancer, follicular lymphoma, colon cancer and esophageal squamous
cell carcinoma, which indicates that USP9X could be a potential
biomarker in human cancers (8–14).
However, the functions of USP9X in liver cancer have not yet been
clarified. In the present study, the protein expression of USP9X in
normal liver tissues and liver cancer tissues was detected. The
results show that USP9X was unregulated in patients with liver
cancer, compared with those in normal liver tissues. When analyzing
the effects of USP9X on the proliferation, invasiveness and
migration of liver cancer cells, the results indicated that
silencing USP9X inhibited cell proliferation, invasiveness and
migration, whereas USP9X overexpression had the opposite effect.
These findings suggest that USP9X is a oncogene in liver cancer, as
well as being a potential biomarker.
It is well known that inhibition of tumor-suppressor
genes or activation of oncogenes is a key event for the development
and malignant progression of liver cancer (32–35).
USP9X plays an important role in the proliferation of liver cancer
cells (17). In the present study,
USP9X was found to be involved in the regulation of the JAK2/STAT3
pathway. Western blot analyses confirmed that USP9X overexpression
could increase the levels of JAK2 and STAT3, along with c-Myc and
MMP2. A number of studies have previously indicated that STAT3
activation improves the expression of c-Myc, which subsequently
promotes cellular proliferation (26). Conversely, STAT3 activation has been
associated with the upregulation of MMP2, and further promotion of
tumor invasive (27–31). At the same time, USP9X-knockdown
inhibits the expression of c-Myc and MMP2, thereby inhibiting the
proliferation, invasiveness and migration of liver cancer cells.
Overexpressing USP9X exerted the opposite effect, which was rescued
by the JAK2 inhibitor Fedratinib. Therefore, upregulation of USP9X
may be an important event in the development and malignant
progression of liver cancer.
To the best of our knowledge, the present study
shows for the first time evidence linking USP9X to the
proliferation and malignant progression of human liver cancer by
regulating the JAK2/STAT3 pathway. However, the specific mechanism
by which USP9X regulates the JAK2/STAT3 pathway requires further
investigation. These results lay the foundation for further study
into the role of USP9X in liver cancer, and exploring the diagnosis
and treatment of liver cancer.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
All authors participated in the design,
interpretation and analysis of the data and reviewed the
manuscript. XS and WY performed the experiments, WY and YL designed
the experiments, CW drafted the initial manuscript, and YH and YL
critically revised the manuscript for important intellectual
content.
Ethics approval and consent to
participate
The present study was approved by the Research
Ethics Committee of the No. 1 People's Hospital of Xuzhou (Xuzhou,
China; approval no. xyy11[2020]-XJSXZ-010). Written informed
consent was provided by all patients prior to the study start.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Allemani C, Matsuda T, Di Carlo V,
Harewood R, Matz M, Nikšić M, Bonaventure A, Valkov M, Johnson CJ,
Estève J, et al CONCORD Working Group, : Global surveillance of
trends in cancer survival 2000-14 (CONCORD-3): Analysis of
individual records for 37 513 025 patients diagnosed with one of 18
cancers from 322 population-based registries in 71 countries.
Lancet. 391:1023–1075. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2016. CA Cancer J Clin. 66:7–30. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yang L, Parkin DM, Li LD, Chen YD and Bray
F: Estimation and projection of the national profile of cancer
mortality in China: 1991-2005. Br J Cancer. 90:2157–2166. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kulik L, Heimbach JK, Zaiem F, Almasri J,
Prokop LJ, Wang Z, Murad MH and Mohammed K: Therapies for patients
with hepatocellular carcinoma awaiting liver transplantation: A
systematic review and meta-analysis. Hepatology. 67:381–400. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Khut PY, Tucker B, Lardelli M and Wood SA:
Evolutionary and expression analysis of the zebrafish
deubiquitylating enzyme, usp9. Zebrafish. 4:95–101. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Pérez-Mancera PA, Rust AG, van der Weyden
L, Kristiansen G, Li A, Sarver AL, Silverstein KA, Grützmann R,
Aust D, Rümmele P, et al Australian Pancreatic Cancer Genome
Initiative, : The deubiquitinase USP9X suppresses pancreatic ductal
adenocarcinoma. Nature. 486:266–270. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kapuria V, Peterson LF, Fang D, Bornmann
WG, Talpaz M and Donato NJ: Deubiquitinase inhibition by
small-molecule WP1130 triggers aggresome formation and tumor cell
apoptosis. Cancer Res. 70:9265–9276. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kerscher O, Felberbaum R and Hochstrasser
M: Modification of proteins by ubiquitin and ubiquitin-like
proteins. Annu Rev Cell Dev Biol. 22:159–180. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang Y, Liu Y, Yang B, Cao H, Yang CX,
Ouyang W, Zhang SM, Yang GF, Zhou FX, Zhou YF, et al: Elevated
expression of USP9X correlates with poor prognosis in human
non-small cell lung cancer. J Thorac Dis. 7:672–679.
2015.PubMed/NCBI
|
|
11
|
Peng J, Hu Q, Liu W, He X, Cui L, Chen X,
Yang M, Liu H, Wei W, Liu S, et al: USP9X expression correlates
with tumor progression and poor prognosis in esophageal squamous
cell carcinoma. Diagn Pathol. 8:1772013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bernassola F, Karin M, Ciechanover A and
Melino G: The HECT family of E3 ubiquitin ligases: Multiple players
in cancer development. Cancer Cell. 14:10–21. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Deng S, Zhou H, Xiong R, Lu Y, Yan D, Xing
T, Dong L, Tang E and Yang H: Over-expression of genes and proteins
of ubiquitin specific peptidases (USPs) and proteasome subunits
(PSs) in breast cancer tissue observed by the methods of RFDD-PCR
and proteomics. Breast Cancer Res Treat. 104:21–30. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Rolén U, Kobzeva V, Gasparjan N, Ovaa H,
Winberg G, Kisseljov F and Masucci MG: Activity profiling of
deubiquitinating enzymes in cervical carcinoma biopsies and cell
lines. Mol Carcinog. 45:260–269. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
15
|
Burger AM and Seth AK: The
ubiquitin-mediated protein degradation pathway in cancer:
Therapeutic implications. Eur J Cancer. 40:2217–2229. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Pardali K and Moustakas A: Actions of
TGF-β as tumor suppressor and pro-metastatic factor in human
cancer. Biochim Biophys Acta. 1775:21–62. 2007.PubMed/NCBI
|
|
17
|
Hu H, Tang C, Jiang Q, Luo W, Liu J, Wei
X, Liu R and Wu Z: Reduced ubiquitin-specific protease 9X
expression induced by RNA interference inhibits the bioactivity of
hepatocellular carcinoma cells. Oncol Lett. 10:268–272. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Liu F, Zhao X, Perna F, Wang L, Koppikar
P, Abdel-Wahab O, Harr MW, Levine RL, Xu H, Tefferi A, et al:
JAK2V617F-mediated phosphorylation of PRMT5 downregulates its
methyltransferase activity and promotes myeloproliferation. Cancer
Cell. 19:283–294. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yu JH, Kim KH and Kim H: SOCS 3 and PPAR-γ
ligands inhibit the expression of IL-6 and TGF-β1 by regulating
JAK2/STAT3 signaling in pancreas. Int J Biochem Cell Biol.
40:677–688. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Griffiths DS, Li J, Dawson MA, Trotter MW,
Cheng YH, Smith AM, Mansfield W, Liu P, Kouzarides T, Nichols J, et
al: LIF-independent JAK signalling to chromatin in embryonic stem
cells uncovered from an adult stem cell disease. Nat Cell Biol.
13:13–21. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
21
|
Berishaj M, Gao SP, Ahmed S, Leslie K,
Al-Ahmadie H, Gerald WL, Bornmann W and Bromberg JF: Stat3 is
tyrosine-phosphorylated through the interleukin-6/glycoprotein
130/Janus kinase pathway in breast cancer. Breast Cancer Res.
9:R322007. View
Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wu R, Liu Y, Zhao Y, Bi Z, Yao Y, Liu Q,
Wang F, Wang Y and Wang X: m6A methylation controls pluripotency of
porcine induced pluripotent stem cells by targeting
SOCS3/JAK2/STAT3 pathway in a YTHDF1/YTHDF2-orchestrated manner.
Cell Death Dis. 10:1712019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kong E, Sucic S, Monje FJ, Savalli G, Diao
W, Khan D, Ronovsky M, Cabatic M, Koban F, Freissmuth M, et al:
STAT3 controls IL6-dependent regulation of serotonin transporter
function and depression-like behavior. Sci Rep. 5:90092015.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chua CY, Liu Y, Granberg KJ, Hu L,
Haapasalo H, Annala MJ, Cogdell DE, Verploegen M, Moore LM, Fuller
GN, et al: IGFBP2 potentiates nuclear EGFR-STAT3 signaling.
Oncogene. 35:738–747. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hansen MF, Greibe E, Skovbjerg S, Rohde S,
Kristensen AC, Jensen TR, Stentoft C, Kjær KH, Kronborg CS and
Martensen PM: Folic acid mediates activation of the pro-oncogene
STAT3 via the Folate Receptor alpha. Cell Signal. 27:1356–1368.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Shirogane T, Fukada T, Muller JMM, Shima
DT, Hibi M and Hirano T: Synergistic roles for Pim-1 and c-Myc in
STAT3-mediated cell cycle progression and antiapoptosis. Immunity.
11:709–719. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kasza A: Signal-dependent Elk-1 target
genes involved in transcript processing and cell migration. Biochim
Biophys Acta. 1829:1026–1033. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ou Y, Liu L, Xue L, Zhou W, Zhao Z, Xu B,
Song Y and Zhan Q: TRAP1 shows clinical significance and promotes
cellular migration and invasion through STAT3/MMP2 pathway in human
esophageal squamous cell cancer. J Genet Genomics. 41:529–537.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Feng J, Yu SY, Li CZ, Li ZY and Zhang YZ:
Integrative proteomics and transcriptomics revealed that activation
of the IL-6R/JAK2/STAT3/MMP9 signaling pathway is correlated with
invasion of pituitary null cell adenomas. Mol Cell Endocrinol.
436:195–203. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wang H, Huo X, Yang XR, He J, Cheng L,
Wang N, Deng X, Jin H, Wang N, Wang C, et al: STAT3-mediated
upregulation of lncRNA HOXD-AS1 as a ceRNA facilitates liver cancer
metastasis by regulating SOX4. Mol Cancer. 16:1362017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hu B, Zhang K, Li S, Li H, Yan Z, Huang L,
Wu J, Han X, Jiang W, Mulatibieke T, et al: HIC1 attenuates
invasion and metastasis by inhibiting the IL-6/STAT3 signalling
pathway in human pancreatic cancer. Cancer Lett. 376:387–398. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Cao K, Gong H, Qiu Z, Wen Q, Zhang B, Tang
T, Zhou X, Cao T, Wang B, Shi H, et al: Hepatitis B virus X protein
reduces the stability of Nrdp1 to up-regulate ErbB3 in
hepatocellular carcinoma cells. Tumour Biol. 37:10375–10382. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Huang J, Zheng C and Shao J, Chen L, Liu X
and Shao J: Overexpression of eEF1A1 regulates G1-phase progression
to promote HCC proliferation through the STAT1-cyclin D1 pathway.
Biochem Biophys Res Commun. 494:542–549. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wang C, Yao B, Xu M and Zheng X: RIP1
upregulation promoted tumor progression by activating AKT/Bcl-2/BAX
signaling and predicted poor postsurgical prognosis in HCC. Tumour
Biol. 37:15305–15313. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Xia W, Zhuang J, Wang G, Ni J, Wang J and
Ye Y: P4HB promotes HCC tumorigenesis through downregulation of
GRP78 and subsequent upregulation of epithelial-to-mesenchymal
transition. Oncotarget. 8:8512–8521. 2017. View Article : Google Scholar : PubMed/NCBI
|