Introduction
Cervical cancer is a frequent diagnosis in women
worldwide; however, its incidence is decreasing in developed
countries thanks to early diagnosis (1,2).
Although comprehensive cervical cancer genomic profiles have been
reported (3), the underlying genomic
mechanisms remain poorly understood. MicroRNAs (miRNAs) represent a
family of abundant endogenous, noncoding RNA molecules, ~22
nucleotides in length (4,5), which post-transcriptionally repress the
expression of target genes by typically binding the 3′untranslated
regions (3′UTRs) of messenger RNA (mRNA) (6,7). It is
becoming increasingly evident that now miRNAs play a role in
virtually every pathophysiological process (8,9). One
particular miRNA, namely miR-150, reportedly influences the
progression of many cancer types (10–17).
Notably, the expression of miR-150 in cervical carcinoma is
significantly higher than that in normal cervical tissue from
healthy donors, and its expression in cervical tissue correlates
with cancer stage progression (18).
Moreover, the overexpression of miR-150 promotes proliferation in
cervical cancer by targeting FOXO4, P2RX7, and SRCIN1
(18–20). FOXO4 regulates many cellular
pathways, including oxidative stress signaling, longevity, insulin
signaling, cell cycle progression, and apoptosis (21). In addition, FOXO4 regulates the
expression of cyclin D1, p27, BIM, and FASL (21). The activation of the P2X7 receptor
mediates apoptosis via the caspase-9 mitochondrial pathway
(22). SRCIN1 is another tumor
suppressor that plays a vital role in tumor cells (23). Furthermore, microRNA-150 promotes
cell proliferation, migration, and invasion of cervical cancer by
targeting PDCD4 (24).
However, the exact identities of the molecular targets of miR-150
in cervical cancer remain uncertain.
We predicted that miR-150 binds the 3′UTR of
CDKN1B using the TargetScan database (http://www.targetscan.org/). The CDKN1B gene
encodes the cyclin-dependent kinase inhibitor 1B
(p27Kip1), which integrates extracellular signals that
negatively regulate the cell cycle progression at the G1 stage
(25,26). In this study, we aimed to investigate
the interaction between miR-150 and CDKN1B, via dual-luciferase
reporter assay in the HeLa human cervical cancer cell line. We also
carried out oligonucleotide transfection and western blot analysis
to examine the effect of miR-150 on HeLa cell proliferation.
Materials and methods
Cell culture
The HeLa human cervical cancer cell line was
obtained from the RIKEN Cell Bank (Tsukuba, Japan) and cultured in
complete Dulbecco's Modified Eagle's medium (Sigma-Aldrich; Merck
KGaA) supplemented with 10% fetal bovine serum (Thermo Fisher
Scientific, Inc.) and 1% antibiotic/antimycotic solution (Thermo
Fisher Scientific, Inc.) at 37°C in a humidified 5% CO2
incubator.
Dual-luciferase reporter assay
To explore the mechanism through which miR-150
promotes tumor progression, we used TargetScan as a miRNA target
prediction algorithm. Based on the frequencies of miR-150 sites in
the 3′UTRs of mRNAs, more than 100 mRNAs were predicted to be
regulated by miR-150. In this study, we focused on CDKN1,
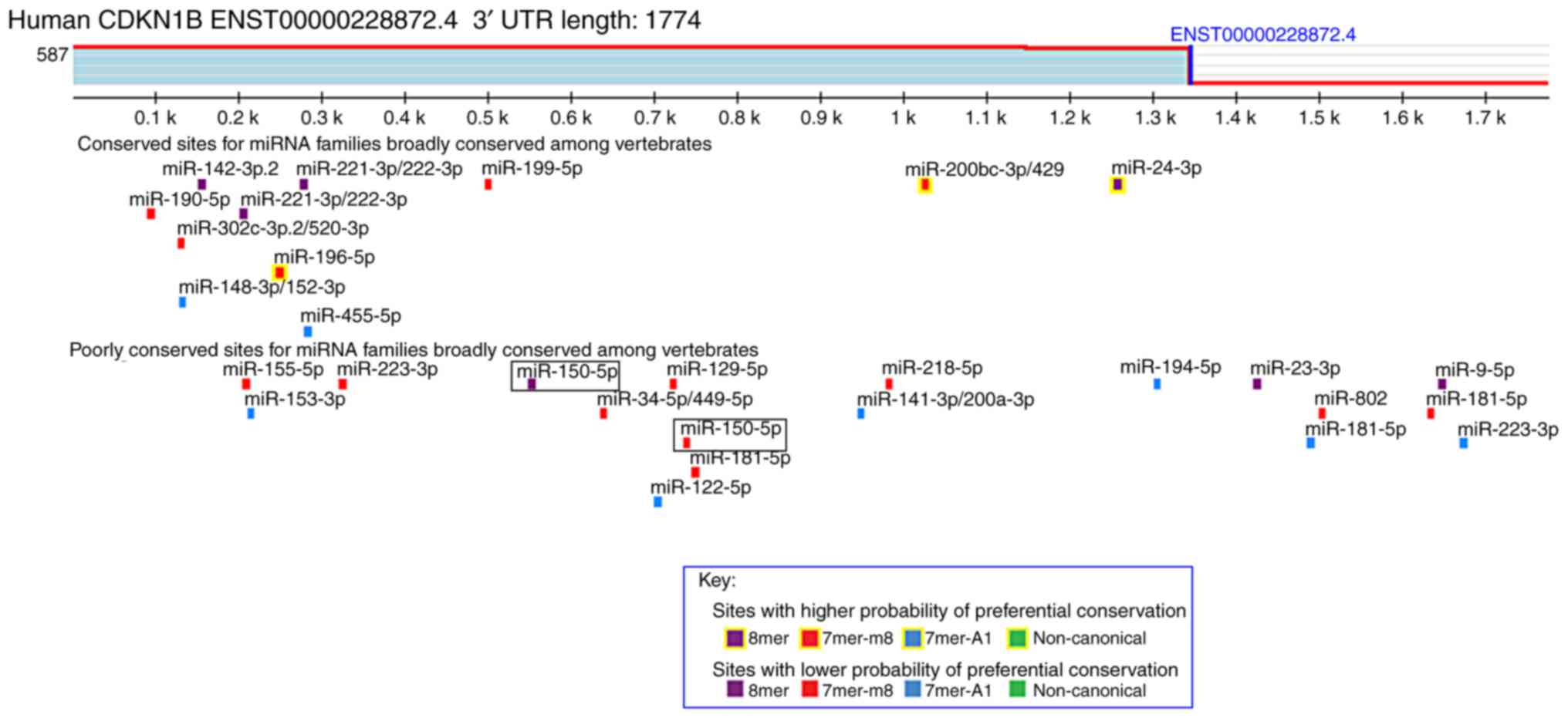
which encodes a cell cycle regulator. As shown in Fig. 1, this analysis revealed putative 8-
and 7-mer binding sites for miR-150 in the 3′UTR of the
CDKN1B transcript. The restriction sites in the psiCHECK-2
vector multiple cloning region are SgfI, XhoI,
PmeI, and NotI. Therefore, two pairs of wild-type and
mutant 3′UTR sequences of CDKN1B with XhoI and
NotI restriction enzyme digestion sites were chemically
synthesized (Table I). The oligo
DNAs of the paired sequences were annealed and digested by
XhoI and NotI restriction enzymes. These synthesized
sequences were integrated into the psiCHECK-2 vector (Promega
Corporation), and the recombinant plasmids were transformed into
One Shot chemically competent E. coli (Thermo Fisher
Scientific, Inc.) and purified according to the manufacturer's
instructions. The recombinant plasmids were named WT1, MUT1, WT2,
and MUT2 (Table I). HeLa cells were
co-transfected with the recombinant plasmids and 100 nM miR-150
mimics or miRNA control for 48 h using Lipofectamine 3000
Transfection reagent (Thermo Fisher Scientific, Inc.). After
transfection, both firefly and Renilla luciferase activities
were measured using the Dual-Luciferase Reporter Assay System
(Promega Corporation). Luciferase activities were normalized to
Renilla luciferase. This analysis was performed in
triplicate.
 | Table I.Chemically synthesized sequences for
dual-luciferase reporter assay |
Table I.
Chemically synthesized sequences for
dual-luciferase reporter assay
| Sequence |
| XhoI
site | 3′UTR of
CDKN1B | NotI
site |
|
|---|
| WT-1 (5′-3′) | GG | CTCGAG |
AAGUUUAUUCUCAUUUGGGAGA | GCGGCCGC | GG |
| WT-1 (3′-5′) | CC | GAGCTC |
UUCAAAUAAGAGUAAACCCUCU | CGCCGGCG | CC |
| MUT-1 (5′-3′) | GG | CTCGAG |
AAGUUUAUUCUCAUAACCCUCU | GCGGCCGC | GG |
| MUT-1 (3′-5′) | CC | GAGCTC |
UUCAAAUAAGAGUAUUGGGAGA | CGCCGGCG | CC |
| WT-2 (5′-3′) | GG | CTCGAG |
AAAAUCCGAGGUGCUUGGGAGU | GCGGCCGC | GG |
| WT-2 (3′-5′) | CC | GAGCTC |
UUUUAGGCUCCACGAACCCUCA | CGCCGGCG | CC |
| MUT-2 (5′-3′) | GG | CTCGAG |
AAAAUCCGAGGUGCAACCCUCU | GCGGCCGC | GG |
| MUT-2 (3′-5′) | CC | GAGCTC |
UUUUAGGCUCCACGUUGGGAGA | CGCCGGCG | CC |
Oligonucleotide transfection
HeLa cells were transfected with 100 nM
hsa-miR-150-5p mimics (miR-150 mimics), 100 nM hsa-miR-150-5p
inhibitors (miR-150 inhibitors), or 100 nM miRNA control (Bioneer)
for 72 h using Lipofectamine RNAiMAX Transfection reagent (Thermo
Fisher Scientific, Inc.) according to the manufacturer's
instructions.
Reverse-transcription quantitative
(RT-q) PCR
The expression level of miR-150 and CDKN1B
mRNA was quantified by real-time PCR. Total RNA from transfected
HeLa cells was extracted using the miRNeasy Mini Kit (Qiagen)
according to the manufacturer's instructions. The quantity and
quality of total genomic RNA were evaluated using the NanoDrop Lite
spectrophotometer (Thermo Fisher Scientific, Inc.). For miRNA
analysis, TaqMan MicroRNA Reverse Transcription kit (Thermo Fisher
Scientific, Inc.) was used according to the manufacturer's
protocol. The program was the following: 16°C for 30 min, 42°C for
30 min, and 85°C for 5 min. Quantitative PCR (qPCR) on miRNA was
performed using TaqMan Fast Advanced Master Mix (Thermo Fisher
Scientific, Inc.) and TaqMan miRNA assays (Thermo Fisher
Scientific, Inc.) for hsa-miR-150-5p (Assay ID, 000473) and RNU6B
(Assay ID, 001093) in accordance with the manufacturer's
instructions. RNU6B was used as a reference gene. The cycling
conditions were as follows; 95°C for 20 sec, followed by 40 cycles
at 95°C for 1 sec and 60°C for 20 sec. For mRNA analysis, the
extracted total RNA was reverse transcribed into cDNA using the
High-Capacity cDNA Reverse Transcription Kit (Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol.
Real-time PCR was performed using the PowerUp SYBR Green Master Mix
(Thermo Fisher Scientific, Inc.) according to the manufacturer's
instructions, starting with 100 ng of total RNA. The cycling
conditions were as follows; 95°C for 20 sec, followed by 40 cycles
at 95°C for 3 sec and 60°C for 30 sec. Primer sequences are shown
in Table II. GAPDH was
quantified as a reference gene. All real-time PCR experiments were
conducted with a StepOnePlus instrument (Thermo Fisher Scientific,
Inc.). The average cycle threshold value for the housekeeping gene
was used to normalize the raw cycle threshold data and calculate
ΔCq. The 2−ΔΔCq method was used to determine the
relative expression levels of miR-150 and CDKN1B. All
experiment was performed in triplicate.
| Table II.Primer sequences for
reverse-transcription quantitative PCR. |
Table II.
Primer sequences for
reverse-transcription quantitative PCR.
| Gene name | Orientation | Primer
sequences |
|---|
| CDKN1B | Forward |
5′-GGCCTCAGAAGACGTCAAAC-3′ |
|
| Reverse |
5′-CAGGATGTCCATTCCATGAAG-3′ |
| GAPDH | Forward |
5′-GCACCGTCAAGGCTGAGAAC-3′ |
|
| Reverse |
5′-TGGTGAAGACGCCAGTGGA-3′ |
Western blotting
Transfected HeLa cells were homogenized in cold
EzRIPA Lysis buffer (ATTO). A 100-µg protein extract was separated
by SDS-PAGE using a 10% polyacrylamide gel, and the proteins were
transferred onto polyvinylidene difluoride membranes (Merck
Millipore). After blocking with 5% non-fat dry milk at room
temperature for 1 h, the membranes were incubated with primary
antibodies against p27Kip1 (1:1,000; cat. no. 3686; Cell
Signaling Technology, Inc.) and β-actin (1:1,000; cat. no. 8457;
Cell Signaling Technology, Inc.) overnight at 4°C. Subsequently,
the membranes were washed and exposed to goat anti-rabbit-IgG HRP
(1:1,000; cat. no. 7074, Cell Signaling Technology, Inc.) for 1 h
at room temperature, which was followed by imaging with a
chemiluminescent substrate. This assay was performed in
triplicate.
Cell cycle analysis
To analyze the cell cycle, transfected cells were
collected, washed with phosphate-buffered saline (PBS), and fixed
in cold 70% ethanol at 4°C overnight. Fixed cells were washed with
PBS and incubated with 250 µg/ml RNase A and 50 µg/ml propidium
iodide for 30 min. The DNA content was determined using a Cell Lab
Quanta SC flow cytometer (Beckman Coulter) and the cell cycle
profile was determined with ModFit LT, which is the most
comprehensive flow cytometric DNA cell cycle analysis software
available (Verity Software House). This assay was performed in
triplicate.
Cell proliferation assay
Cell proliferation of transfected HeLa cells was
determined using the
2-(2-methoxy-4-nitrophenyl)-3-(4-nitrophenyl)-5-(2,4-disulfophenyl)-2H-tetrazolium
(WST-8) assay with the Cell Counting Kit-8 (CCK-8; Dojindo
Laboratories) every 24 h according to the manufacturer's
instructions. CCK-8 solutions were added to the cell culture, and
cells were incubated at 37°C for 2 h. Later, the absorbance was
measured at 450 nm. This assay was performed in triplicate.
Statistical analysis
Results are expressed as the mean ± standard
deviation. Statistical significance for the experiments was
determined using Dunn's test, a Mann-Whitney's U test, or a
one-way factorial ANOVA test. P<0.05 was considered to indicate
a statistically significant difference. StatMate V software was
used to perform statistical analyses.
Results
miR-150 directly targets the 3′UTR of
CDKN1B in transfected HeLa cells
miRNAs repress the expression of target genes by
binding the 3′UTR of target genes. The target sites of miRNA-150
were investigated using the TargetScan miRNA target prediction
database. CDKN1B is a predicted target gene of miR-150. Two
predicted miR-150-binding sites in the 3′UTR of CDKN1B were
identified. To verify these two binding sites, we designed
recombinant plasmids, including wild-type or mutant 3′UTR sequences
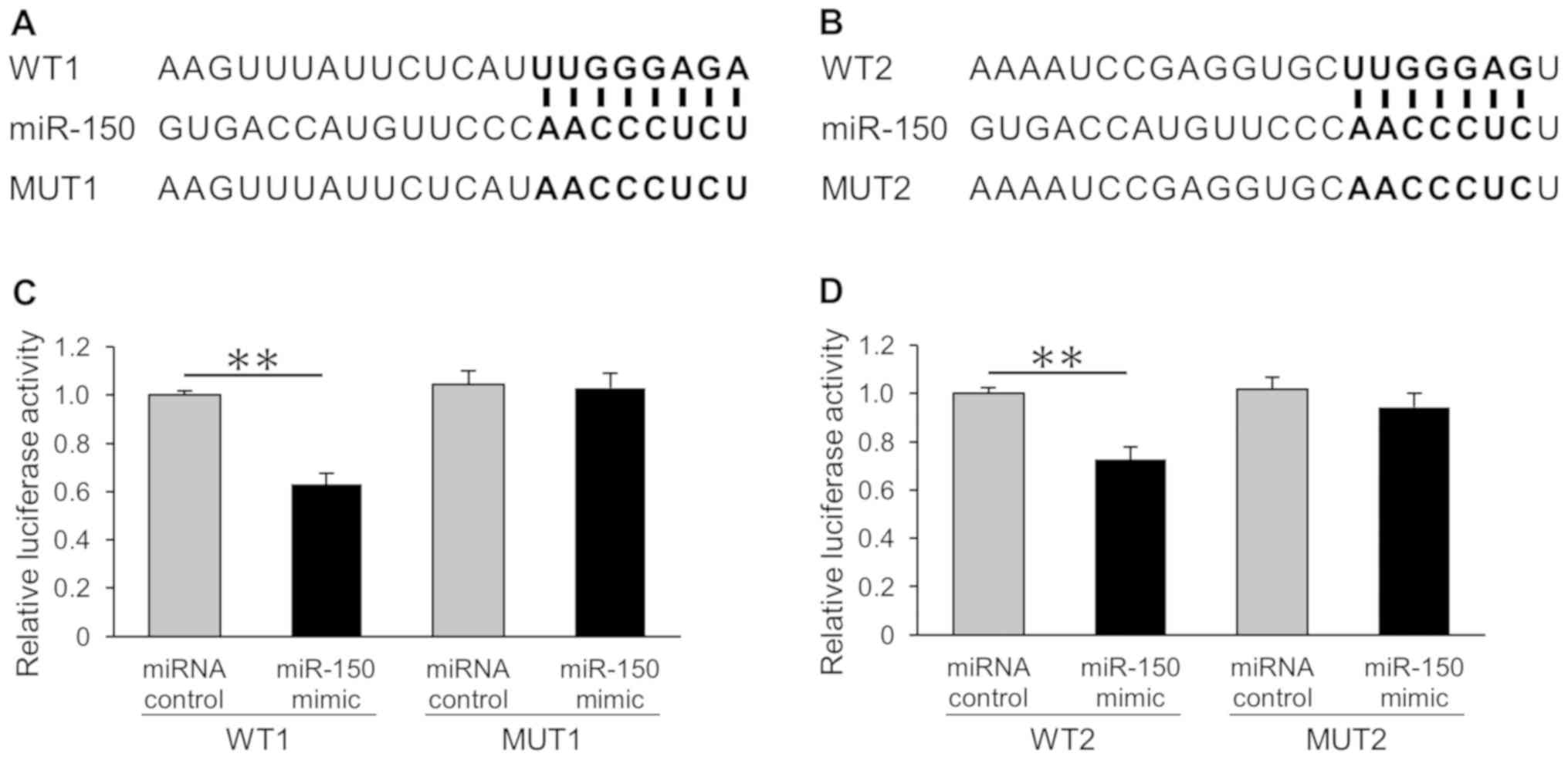
of CDKN1B (Fig. 2A and B).
Then, dual-luciferase reporter assays were performed to investigate
whether miR-150 could influence the expression of luciferase via
binding to the predicted sequences. Our results showed that miR-150
directly downregulated CDKN1B in the HeLa cells (Fig. 2C and D). Conversely, there was no
effect on the reporters with mutant sequences.
miR-150 reduces CDKN1B mRNA expression
and p27Kip1 protein expression in transfected HeLa
cells
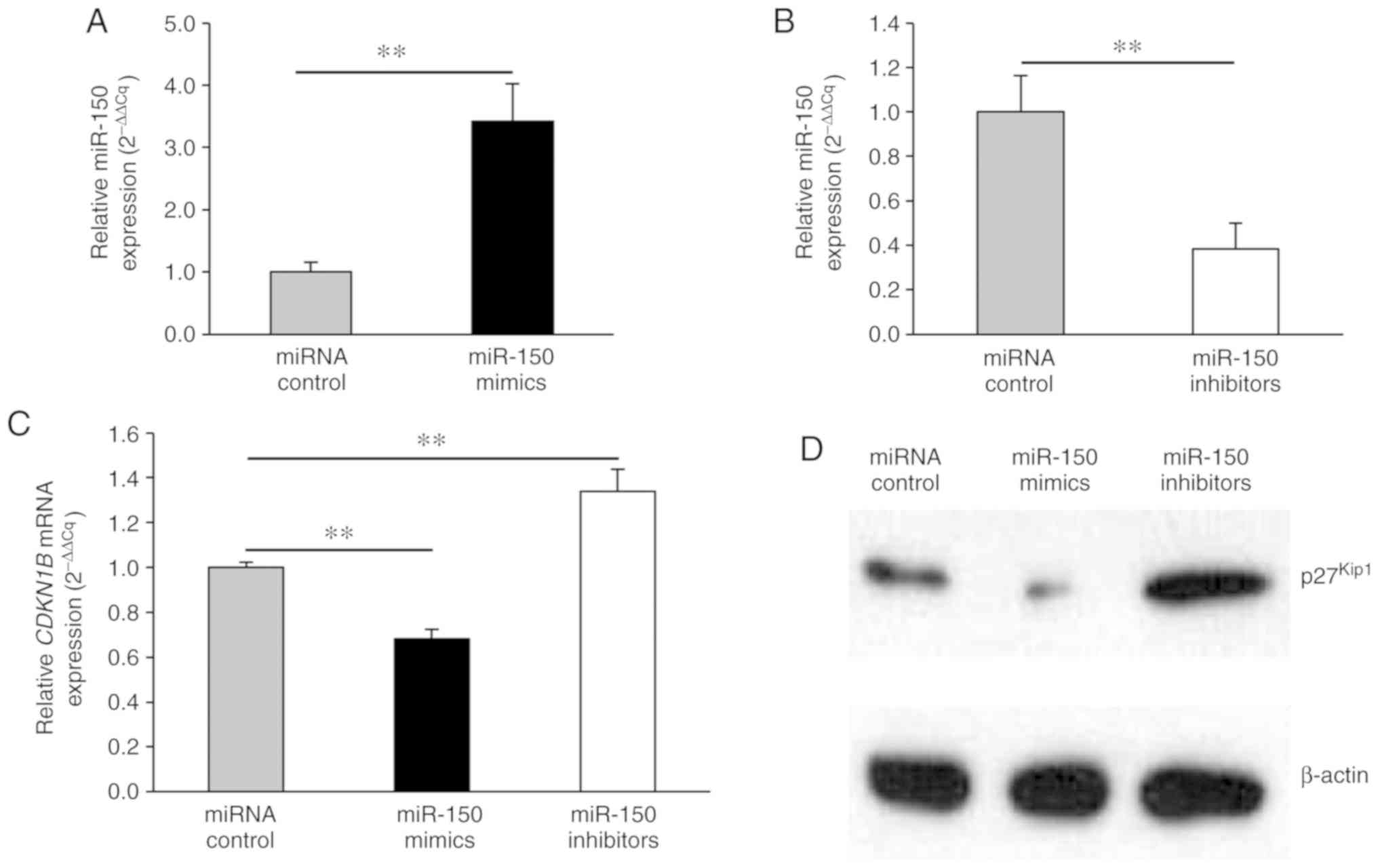
To investigate the molecular details of miR-150
effects, HeLa cells were transfected with miR-150 mimics, miR-150
inhibitors, or miRNA control. Firstly, qPCR for miRNA showed that
miR-150 expression levels were significantly higher and lower in
the miR-150 mimic-transfected and miR-150 inhibitor-transfected
cells, respectively, relative to the controls (Fig. 3A, B). Since miR-150 directly targeted
the 3′UTR of the CDKN1B gene, the expression levels of
CDKN1B mRNA and p27Kip1 protein were evaluated
after the transfection of miR-150 mimics or inhibitors using qPCR
and western blot analysis, respectively. miR-150 mimics
significantly decreased CDKN1B mRNA expression levels
(Fig. 3C). In contrast, miR-150
inhibitors significantly upregulated CDKN1B mRNA expression
levels (Fig. 3C). Consistent with
the RT-PCR results, the protein levels of p27Kip1 were
reduced by miR-150 mimics and increased by miR-150 inhibitors
(Fig. 3D).
miR-150 facilitates cell cycle
progression in transfected HeLa cells
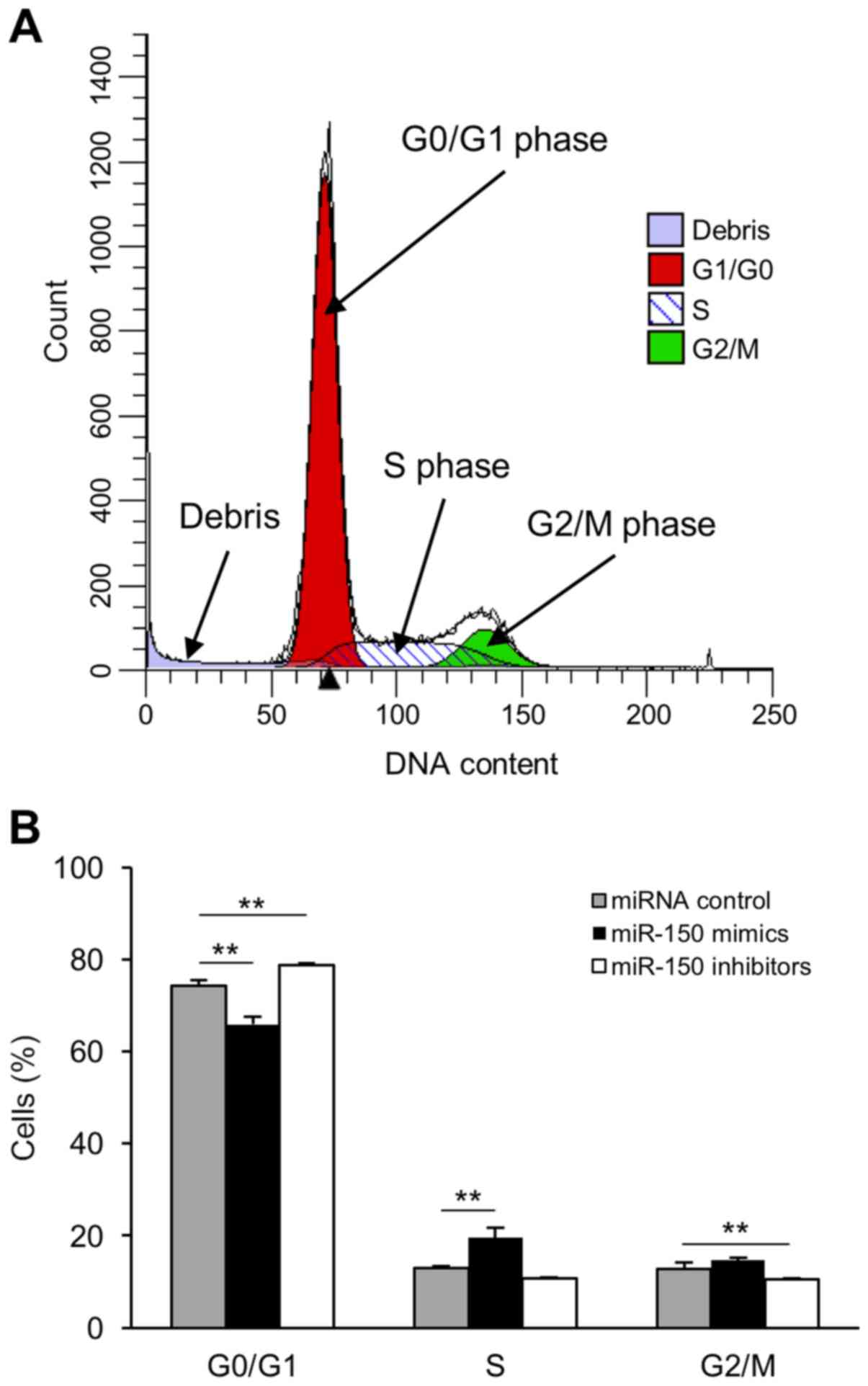
To investigate the role of miR-150 in cervical
cancer cells, we analyzed cell cycle progression in transfected
HeLa cells using a flow cytometer (Fig.
4A). There was an increased number of cells in the S phase in
miR-150 mimic-transfected cells compared to that in miRNA
control-transfected cells (Fig. 4B).
In contrast, transfection of miR-150 inhibitors induced G0/G1 phase
arrest (Fig. 4B).
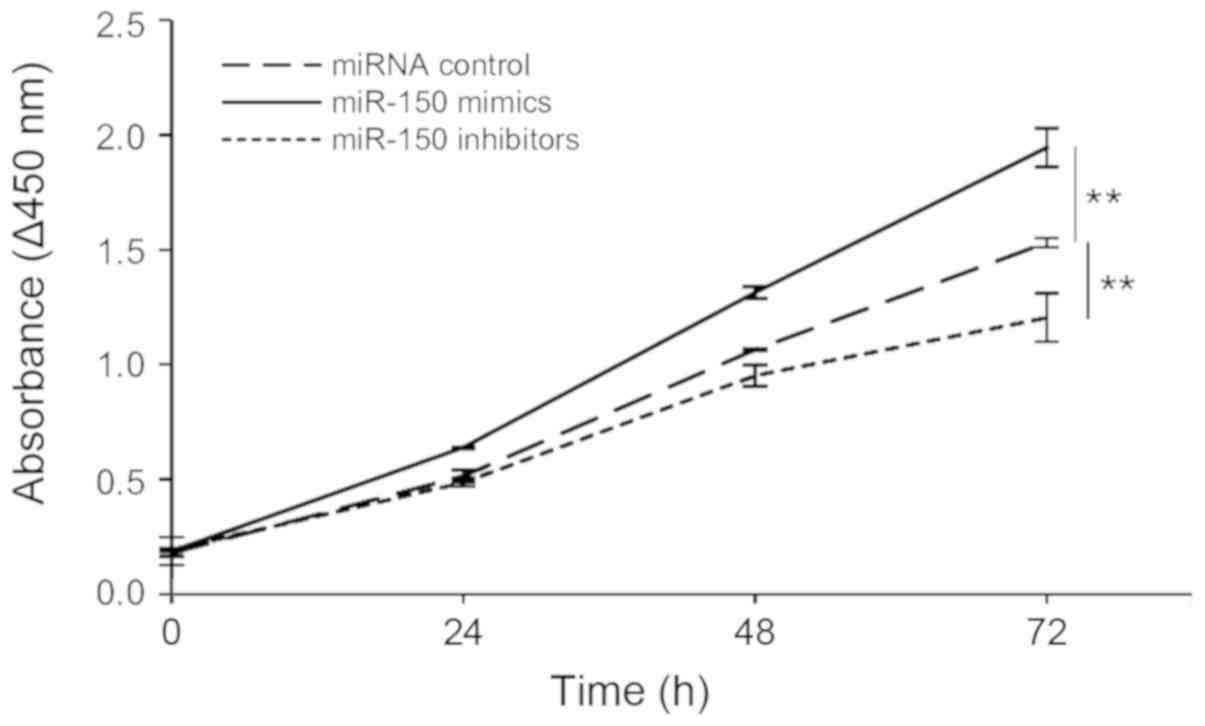
miR-150 promotes cell proliferation in
transfected HeLa cells
We hypothesized that miR-150 levels are involved in
cell proliferation in cervical cancer. The proliferation of
transfected HeLa cells was determined by measuring the absorbance
at 450 nm based on a colorimetric assay. Our results showed that
the overexpression of miR-150 led to a significant increase in cell
proliferation, whereas expressing miR-150 inhibitors led to a
significant decrease in HeLa cell proliferation (Fig. 5).
Discussion
miRNAs play critical roles in many diverse
biological and pathological processes, such as cell proliferation,
migration, apoptosis, and the pathogenesis of cancer (8,9). Among
them, recent studies have reported the influence of miR-150 on many
types of cancers (10–17). Previous studies have reported
decreased levels of miR-150 in colorectal cancer (27,28),
liver cancer (29), pancreatic
cancer (12), esophageal squamous
cell carcinoma (30), and ovarian
cancer (14). In contrast, the
upregulated expression of miR-150 has been found in breast cancer
(13), non-small cell lung cancer
(31–33), gastric cancer (34), and prostate cancer (15). A previous report indicated that
miR-150 expression levels are significantly higher in cervical
carcinoma cells than in para-carcinoma tissues (18). Moreover, the expression of miR-150 in
samples from patients with cervical carcinoma significantly
exceeded that of normal cervical tissue from healthy donors.
Moreover, miR-50 expression correlated with stage progression
(18). However, results depict a
controversial picture regarding the targets of miR-150 in cervical
cancer. Moreover, the consensus regarding a hypothetical influence
of miR-150 on cervical cancer is lacking.
In the present study, we analyzed the effect of
miR-150 on cervical cancer cells in vitro. We first
investigated the target sites of miRNA-150 using the TargetScan
miRNA target prediction database and predicted two binding sites on
the 3′UTR of CDKN1B. Further, the results of dual-luciferase
reporter assays showed that miR-150 directly bound and
downregulated CDKN1B in HeLa cells. Notably, miR-150
significantly decreased CDKN1B mRNA expression levels, and
western blot analysis showed that the transfection of miR-150
mimics reduced p27Kip1 protein levels. The
p27Kip1 protein transduces extracellular signals of cell
cycle regulators that negatively regulate G1 cell cycle progression
(25,26).
To investigate the role of miR-150 in HeLa cells, we
analyzed cell cycle progression by flow cytometry. In miR-150
mimic-transfected cells, a higher proportion of cells was in the S
phase. Moreover, the transfection of miR-150 inhibitors induced
cell cycle arrest at the G1/G0 phase. These results suggested that
miR-150 promotes cell cycle progression from the G0/G1 to S phase
in cervical cancer cells by suppressing the p27Kip1
function.
We then measured transfected HeLa cell numbers using
CCK-8, via a sensitive colorimetric assay for the determination of
cell proliferation. The cells transfected with miR-150 mimics
proliferated faster than control cells, whereas the cells
transfected with miR-150 inhibitors grew slower than control cells.
These findings suggest that miR-150 promotes cell cycle progression
and cell proliferation by suppressing p27Kip1
expression. Our results validated previously reported direct
targets of miR-150 in cervical cancer, namely FOXO4, P2RX7,
and SRCIN1 (18–20). However, our results also suggest that
miR-150 promotes proliferation by directly targeting CDKN1B.
Considering the well-known pleiotropic effects of miRNA, a single
miRNA could bind several hundred target mRNAs (35). Although the present study provides a
clearer understanding of the function and molecular mechanism of
miR-150 in HeLa cells, more comprehensive in vitro and in
vivo studies are needed to decipher the role of miR-150 in
cervical cancer.
In conclusion, our results indicate that miR-150
promotes cell cycle progression and cell proliferation of HeLa
human cervical cancer cells by directly suppressing
p27Kip1 expression. We identified CDKN1B as a
novel target of miR-150 in cervical cancer.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
WO conceived and designed all the experiments. WO
and KH performed the experiments. HT, KI, YY, AK, TN and NY
analyzed the experimental data. TN and NY contributed to writing of
the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of Kagawa Prefectural University of Health Sciences,
Japan.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2016. CA Cancer J Clin. 66:7–30. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hagemann T, Bozanovic T, Hooper S, Ljubic
A, Slettenaar VI, Wilson JL, Singh N, Gayther SA, Shepherd JH and
Van Trappen PO: Molecular profiling of cervical cancer progression.
Br J Cancer. 96:321–328. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lai EC: miRNAs: Whys and wherefores of
miRNA-mediated regulation. Curr Biol. 15:R458–R460. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Friedman RC, Farh KK, Burge CB and Bartel
DP: Most mammalian mRNAs are conserved targets of microRNAs. Genome
Res. 19:92–105. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ebert MS and Sharp PA: Roles for microRNAs
in conferring robustness to biological processes. Cell.
149:515–524. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Calin GA and Croce CM: MicroRNA signatures
in human cancers. Nat Rev Cancer. 6:857–866. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Farazi TA, Spitzer JI, Morozov P and
Tuschl T: miRNAs in human cancer. J Pathol. 223:102–115. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang F, Ren X and Zhang X: Role of
microRNA-150 in solid tumors. Oncol Lett. 10:11–16. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Feng J, Yang Y, Zhang P, Wang F, Ma Y, Qin
H and Wang Y: miR-150 functions as a tumour suppressor in human
colorectal cancer by targeting c-Myb. J Cell Mol Med. 18:2125–2134.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Srivastava SK, Bhardwaj A, Singh S, Arora
S, Wang B, Grizzle WE and Singh AP: MicroRNA-150 directly targets
MUC4 and suppresses growth and malignant behavior of pancreatic
cancer cells. Carcinogenesis. 32:1832–1839. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Huang S, Chen Y, Wu W, Ouyang N, Chen J,
Li H, Liu X, Su F, Lin L and Yao Y: miR-150 promotes human breast
cancer growth and malignant behavior by targeting the pro-apoptotic
purinergic P2X7 receptor. PLoS One. 8:e807072013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Jin M, Yang Z, Ye W, Xu H and Hua X:
MicroRNA-150 predicts a favorable prognosis in patients with
epithelial ovarian cancer, and inhibits cell invasion and
metastasis by suppressing transcriptional repressor ZEB1. PLoS One.
9:e1039652014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Dezhong L, Xiaoyi Z, Xianlian L, Hongyan
Z, Guohua Z, Bo S, Shenglei Z and Lian Z: miR-150 is a factor of
survival in prostate cancer patients. J BUON. 20:173–179.
2015.PubMed/NCBI
|
|
16
|
Zhang J, Luo N, Luo Y, Peng Z, Zhang T and
Li S: MicroRNA-150 inhibits human CD133-positive liver cancer stem
cells through negative regulation of the transcription factor
c-Myb. Int J Oncol. 40:747–756. 2012.PubMed/NCBI
|
|
17
|
Katada T, Ishiguro H, Kuwabara Y, Kimura
M, Mitui A, Mori Y, Ogawa R, Harata K and Fujii Y: MicroRNA
expression profile in undifferentiated gastric cancer. Int J Oncol.
34:537–542. 2009.PubMed/NCBI
|
|
18
|
Li J, Hu L, Tian C, Lu F, Wu J and Liu L:
MicroRNA-150 promotes cervical cancer cell growth and survival by
targeting FOXO4. BMC Mol Biol. 16:242015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhou L, Qi X, Potashkin JA, Abdul-Karim FW
and Gorodeski GI: MicroRNAs miR-186 and miR-150 down-regulate
expression of the pro-apoptotic purinergic P2X7 receptor by
activation of instability sites at the 3′-untranslated region of
the gene that decrease steady-state levels of the transcript. J
Biol Chem. 283:28274–2886. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhu J and Han S: miR-150-5p promotes the
proliferation and epithelial-mesenchymal transition of cervical
carcinoma cells via targeting SRCIN1. Pathol Res Pract.
215:738–747. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Boura E, Silhan J, Herman P, Vecer J, Sulc
M, Teisinger J, Obsilova V and Obsil T: Both the N-terminal loop
and wing W2 of the forkhead domain of transcription factor Foxo4
are important for DNA binding. J Biol Chem. 282:8265–8275. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang Q, Li X, Wang L, Feng YH, Zeng R and
Gorodeski G: Antiapoptotic effects of estrogen in normal and cancer
human cervical epithelial cells. Endocrinology. 145:5568–5579.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Cabodi S, del Pilar Camacho-Leal M, Di
Stefano P and Defilippi P: Integrin signalling adaptors: Not only
figurants in the cancer story. Nat Rev Cancer. 10:858–870. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang Z, Wang J, Li J, Wang X and Song W:
MicroRNA-150 promotes cell proliferation, migration, and invasion
of cervical cancer through targeting PDCD4. Biomed Pharmacother.
97:511–517. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Polyak K, Lee MH, Erdjument-Bromage H,
Koff A, Roberts JM, Tempst P and Massagué J: Cloning of p27Kip1, a
cyclin-dependent kinase inhibitor and a potential mediator of
extracellular antimitogenic signals. Cell. 78:59–66. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Toyoshima H and Hunter T: p27, a novel
inhibitor of G1 cyclin-Cdk protein kinase activity, is related to
p21. Cell. 78:67–74. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Pizzini S, Bisognin A, Mandruzzato S,
Biasiolo M, Facciolli A, Perilli L, Rossi E, Esposito G, Rugge M,
Pilati P, et al: Impact of microRNAs on regulatory networks and
pathways in human colorectal carcinogenesis and development of
metastasis. BMC Genomics. 14:5892013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ma Y, Zhang P, Wang F, Zhang H, Yang J,
Peng J, Liu W and Qin H: miR-150 as a potential biomarker
associated with prognosis and therapeutic outcome in colorectal
cancer. Gut. 61:1447–1453. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Sun W, Zhang Z, Wang J, Shang R, Zhou L,
Wang X, Duan J, Ruan B, Gao Y, Dai B, et al: MicroRNA-150
suppresses cell proliferation and metastasis in hepatocellular
carcinoma by inhibiting the GAB1-ERK axis. Oncotarget.
7:11595–11608. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yokobori T, Suzuki S, Tanaka N, Inose T,
Sohda M, Sano A, Sakai M, Nakajima M, Miyazaki T, Kato H and Kuwano
H: miR-150 is associated with poor prognosis in esophageal squamous
cell carcinoma via targeting the EMT inducer ZEB1. Cancer Sci.
104:48–54. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Li H, Ouyang R, Wang Z, Zhou W, Chen H,
Jiang Y, Zhang Y, Li H, Liao M, Wang W, et al: miR-150 promotes
cellular metastasis in non-small cell lung cancer by targeting
FOXO4. Sci Rep. 6:390012016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yin QW, Sun XF, Yang GT, Li XB, Wu MS and
Zhao J: Increased expression of microRNA-150 is associated with
poor prognosis in non-small cell lung cancer. Int J Clin Exp
Pathol. 8:842–846. 2015.PubMed/NCBI
|
|
33
|
Zhang N, Wei X and Xu L: miR-150 promotes
the proliferation of lung cancer cells by targeting P53. FEBS Lett.
587:2346–2351. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wu Q, Jin H, Yang Z, Luo G, Lu Y, Li K,
Ren G, Su T, Pan Y, Feng B, et al: miR-150 promotes gastric cancer
proliferation by negatively regulating the pro-apoptotic gene EGR2.
Biochem Biophys Res Commun. 392:340–345. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Filipowicz W, Bhattacharyya SN and
Sonenberg N: Mechanisms of post-transcriptional regulation by
microRNAs: Are the answers in sight? Nat Rev Genet. 9:102–114.
2008. View Article : Google Scholar : PubMed/NCBI
|