Introduction
Hepatocellular carcinoma (HCC) is the main type of
primary liver cancer and is one of the most common malignancies
with poor survival (1). Hepatectomy
is a potentially curative treatment, but the recurrence rate of HCC
after surgery is remarkably high at approximately 70% (2). Therefore, further understanding of the
molecular mechanisms of HCC development and recurrence is
required.
More than 100 types of chemical modifications have
been identified in RNA (3).
Recently, internal modifications of mRNA have received attention
for their roles in mRNA metabolism. N6-methyladenosine
(m6A) is the most abundant modification of mRNA in
eukaryotes and was first reported in the 1970s (4). Recent evidence suggests that
m6A has various functions in RNA metabolism, such as
pre-mRNA splicing, 3′-end processing, nuclear export, translation
regulation, regulation of mRNA decay and noncoding RNA processing
(5–7). Furthermore, m6A methylation
has been revealed to have crucial roles in the initiation and
progression of cancer (8).
m6A readers are the proteins that
recognize and bind to m6A sites and thereby elicit
multiple effects (9). YT521-B
homology domain family 2 (YTHDF2) was the first identified
m6A binding protein (10).
YTHDF2 weakens mRNA stability by recognizing m6A,
while YT521-B homology domain family 1 (YTHDF1) promotes
mRNA translation efficiency (11).
In solid cancer, YTHDF2 and YTHDF1 have been reported
to have roles as both tumour promoters and suppressors (12–14).
However, the significance of YTHDF1 and YTHDF2 in
oncogenesis remains unclear.
In the current study, we assessed the expression of
YTHDF1 and YTHDF2 in both resected HCC tissues and
paired normal liver tissues collected from patients who underwent
surgery with curative intent. We also sought to discover novel
prognostic implications of m6A readers that could be
used to predict prognosis in patients with resected HCC.
Patients and methods
Patients and samples
A total of 177 frozen tumour specimens and paired
paratumor noncancerous tissues were collected from patients with
HCC who underwent surgery at Nagoya University Hospital (Nagoya
City, Japan) between January 1998 and April 2014. All fresh tissues
were immediately frozen in liquid nitrogen and stored at −80°C
until use. Patient characteristics are summarized in Table I. After surgery, all patients were
monitored via blood examinations, ultrasonography, and computed
tomography once every six months. Angiography was performed for
further information whenever recurrence was suspected. The median
follow-up duration of all patients was 48.8 months (range, 0.3 to
191 months). This study and all procedures were approved by the
Institutional Review Board at Nagoya University (Nagoya City,
Japan), and all patients provided written informed consent. All
clinical investigations were conducted in accordance with the
principles of the Declaration of Helsinki.
 | Table I.Clinicopathological characteristics
of patients with hepatocellular carcinoma (n=177). |
Table I.
Clinicopathological characteristics
of patients with hepatocellular carcinoma (n=177).
|
Characteristics | Value |
|---|
| Median age (range),
years | 65 (37–84) |
| Sex, male:female, n
(%) | 148 (84) : 29
(16) |
| Viral infection,
HBV:HCV:non-HBV/HCV, n (%) | 41 (23) : 106 (60)
: 30 (17) |
| Median albumin
(range), g/dl | 3.9 (2.3–4.9) |
| Median total
bilirubin (range), mg/dl | 0.7 (0.2–7.3) |
| Median PT (range),
% | 88.7
(46.9–138) |
| Median ICG-R15
(range), % | 11.4
(1.6–70.5) |
| Child-Pugh
classification, A:B, n (%) | 166 (94):10
(6) |
| Liver damage
classification, A:B:C, n (%) | 142 (83):28 (16):1
(1) |
| Tumour
multiplicity, solitary:multiple, n (%) | 138 (78):39
(22) |
| Median tumour size
(range), cm | 3.5 (0.15–15) |
| Median AFP (range),
ng/ml | 17
(0.8–119923) |
| Stage, I:II:III:IV,
n (%) | 19 (11):91 (52):44
(25):21 (12) |
RNA isolation and RT-qPCR
Total RNA was extracted from tissue samples using a
Qiagen miRNeasy mini-kit (Qiagen, Hilden, Germany). We used DNAse
to avoid contamination, and RNA quality was analysed by a NanoDrop
(Thermo Scientific Fisher, Waltham, MA, USA). Total RNA was
converted to complementary DNA by reverse transcription with M-MLV
Reverse Transcriptase (Invitrogen, Carlsbad, CA, USA). This total
cDNA was used as a template for the next step of quantitative PCR
(qPCR). qPCR was performed using SYBR Premix Ex Taq II (Takara
Clontech, Kyoto, Japan) under the following conditions:
Denaturation at 95°C for 10 sec and 40 cycles of denaturation at
95°C for 5 sec and annealing/extension at 60°C for 30 sec. The SYBR
Green signal was detected in real-time using a StepOne Plus
Real-Time PCR System (Life Technologies, Carlsbad, CA, USA). The
relative quantification method was used, and the expression level
of each gene was normalized to the expression level of the control
gene glyceraldehyde-3-phosphate dehydrogenase (GAPDH) for
each sample. The relative gene expression levels were determined
using the comparative threshold cycle (2−ΔCT)
method.
The PCR primers used in the current study were
specific for the 78-base-pair fragment of YTHDF1 (sense,
5′-TCCATCTTCGACGACTTTGCT-3′; antisense, 5′-TCGACTCTGCCGTTCCTTG-3′)
and for the 50-base-pair fragment of YTHDF2 (sense,
5′-GAGGATCTGAGAGCCATGTCG-3′; antisense,
5′-ATTTTGTACTGCTCCAAGAGGC-3′). GAPDH primers (sense,
5′-GAGTCCACTGGCGTCTTCAC-3′; antisense, 5′-GTTCACACCCATGACGAACA-3′)
were used to quantify the expression in each sample as an internal
control. The primers were designed as intron spanning. All qPCR
experiments were performed in duplicate, including the
template-omitted negative controls.
Acquisition of publicly available
data
Normalized TCGA RNA-sequencing data of HCC were
downloaded from the Broad GDAC Firehose (http://gdac.broadinstitute.org/, accessed on January
1st, 2020). This dataset consists of 50 noncancerous cases and 360
HCC cases, including seven HCC cases mixed with
hepatocholangiocarcinoma and two cases with fibrolamellar
carcinoma. Of the 360 cases, there were 266 cases with
recurrence-free survival (RFS) information and 336 cases with
overall survival (OS) information.
Statistical analysis
Continuous variables are expressed as the median
(range), and the expression of each target gene was compared by a
Wilcoxon signed-rank test and paired t-test. Categorical variables
were compared using χ2 or Fisher's exact tests, as
appropriate. The OS and RFS rates at each point of the follow-up
time were estimated using the Kaplan-Meier method and compared
using a log-rank test. A Cox proportional hazard regression model
was used to perform univariate analysis and multivariate analysis
for OS and RFS. In the multivariate analysis, variables that showed
statistical significance in the univariate Cox proportional hazard
regression were included. All statistical analyses were performed
using R version 3.5.3 (http//www.r-project.org/), and P<0.05 obtained
using two-tailed tests was considered to indicate statistical
significance.
Results
YTHDF1 and YTHDF2 in resected
specimens from HCC patients
First, expression analyses of m6A readers
were conducted with our surgically resected specimens. The
expression levels of YTHDF1 and YTHDF2 were measured
by qPCR. The expression of YTHDF1 and YTHDF2 was not
significantly different between tumour tissues and noncancerous
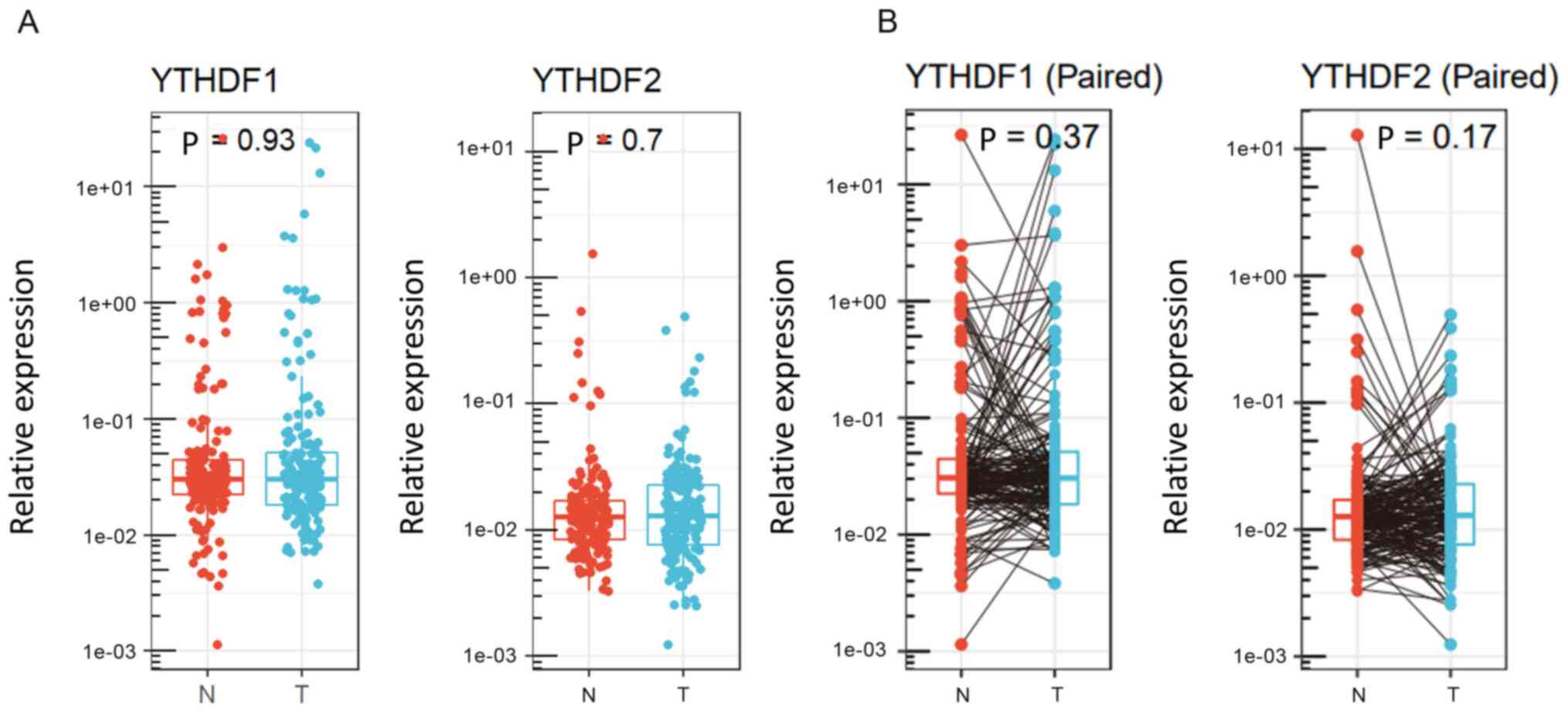
tissues (P=0.93 and P=0.7, respectively, Fig. 1A). Fig.
1B shows individual changes in YTHDF1 and YTHDF2
expression in paired analysis. Based on the results obtained by
qPCR, 177 HCC cases were divided into two groups according to
YTHDF1 and YTHDF2 expression in tumour tissues. We
selected the cut-off values that showed the best statistical
difference. Clinical features of the groups stratified by
YTHDF1 and YTHDF2 expression are shown in Table II. In HCC tissues, low YTHDF1
expression was associated with a lack of capsule (65% vs. 81%,
P=0.02), whereas low YTHDF2 expression was associated with
septal formation (73% vs. 53%, P=0.02).
| Table II.Clinical features of 177 patients
with hepatocellular carcinoma according to YTHDF1 and YTHDF2
expression. |
Table II.
Clinical features of 177 patients
with hepatocellular carcinoma according to YTHDF1 and YTHDF2
expression.
|
| YTHDF1 expression
(n=172)a |
| YTHDF2 expression
(n=174)b |
|
|---|
|
|
|
|
|
|
|---|
| Variables | Low (n=108) | High (n=64) | P-value | Low (n=137) | High (n=37) | P-value |
|---|
| Age, years |
|
|
|
|
|
|
|
<65 | 50 | 34 | 0.43 | 67 | 18 | 0.98 |
|
≥65 | 58 | 30 |
| 70 | 19 |
|
| Sex |
|
|
|
|
|
|
|
Female | 14 | 13 | 0.28 | 22 | 6 | 0.98 |
|
Male | 94 | 51 |
| 115 | 31 |
|
| Virus
infection |
|
|
|
|
|
|
|
Others | 44 | 25 | 0.87 | 58 | 13 | 0.46 |
|
HCV | 64 | 39 |
| 79 | 24 |
|
| Albumin, g/dl |
|
|
|
|
|
|
|
≥3.5 | 85 | 54 | 0.54 | 110 | 30 | 0.98 |
|
<3.5 | 22 | 10 |
| 26 | 7 |
|
| NA | 1 | 0 |
| 1 | 0 |
|
| PT, % |
|
|
|
|
|
|
|
≥70 | 91 | 57 | 0.50 | 114 | 34 | 0.29 |
|
<70 | 16 | 7 |
| 22 | 3 |
|
| NA | 1 | 0 |
| 1 | 0 |
|
| ICG-R15, % |
|
|
|
|
|
|
|
<15 | 58 | 35 | 0.91 | 75 | 19 | 0.15 |
|
≥15 | 19 | 12 |
| 21 | 11 |
|
| NA | 31 | 17 |
| 41 | 7 |
|
| Liver
cirrhosis |
|
|
|
|
|
|
|
Negative | 72 | 39 | 0.51 | 90 | 20 | 0.25 |
|
Positive | 36 | 25 |
| 47 | 17 |
|
| Child-Pugh
classification |
|
|
|
|
|
|
| A | 100 | 61 | 0.74 | 128 | 35 | 0.91 |
| B | 7 | 3 |
| 8 | 2 |
|
| NA | 1 | 0 |
| 1 | 0 |
|
| Liver damage |
|
|
|
|
|
|
| A | 84 | 54 | 0.53 | 110 | 29 | 0.80 |
| B or
C | 19 | 9 |
| 22 | 7 |
|
| NA | 5 | 1 |
| 5 | 1 |
|
| Tumour number |
|
|
|
|
|
|
|
Solitary | 86 | 48 | 0.57 | 109 | 27 | 0.38 |
|
Multiple | 22 | 16 |
| 28 | 10 |
|
| Tumour size,
cm |
|
|
|
|
|
|
|
<2 | 14 | 10 | 0.82 | 17 | 6 | 0.79 |
| ≥2 | 78 | 50 |
| 100 | 29 |
|
| NA | 16 | 4 |
| 20 | 2 |
|
| AFP, ng/ml |
|
|
|
|
|
|
|
<20 | 58 | 35 | 0.91 | 68 | 25 | 0.06 |
|
≥20 | 48 | 28 |
| 67 | 11 |
|
| NA | 2 | 1 |
| 2 | 1 |
|
|
Differentiation |
|
|
|
|
|
|
| Good or
moderate | 98 | 56 | 0.77 | 121 | 35 | 0.08 |
|
Poor | 8 | 6 |
| 14 | 0 |
|
| NA | 2 | 2 |
| 2 | 2 |
|
| Growth form |
|
|
|
|
|
|
|
Expansive | 88 | 54 | 0.52 | 114 | 31 | 0.42 |
|
Infiltrative | 18 | 8 |
| 22 | 3 |
|
| NA | 2 | 2 |
| 1 | 3 |
|
| Formation of
capsule |
|
|
|
|
|
|
|
Positive | 70 | 52 | 0.02 | 97 | 27 | 0.84 |
|
Negative | 38 | 12 |
| 40 | 10 |
|
| Infiltration to
capsule |
|
|
|
|
|
|
|
Negative | 52 | 22 | 0.08 | 57 | 17 | 0.71 |
|
Positive | 55 | 42 |
| 79 | 20 |
|
| NA | 1 | 0 |
| 1 | 0 |
|
| Septal
formation |
|
|
|
|
|
|
|
Positive | 74 | 42 | 0.61 | 98 | 19 | 0.02 |
|
Negative | 31 | 21 |
| 36 | 17 |
|
| NA | 3 | 1 |
| 3 | 1 |
|
| Serosal
invasion |
|
|
|
|
|
|
|
Negative | 85 | 50 | 0.69 | 107 | 31 | 0.81 |
|
Positive | 19 | 14 |
| 26 | 6 |
|
| NA | 4 | 0 |
| 4 | 0 |
|
| Portal vein or
hepatic vein invasion |
|
|
|
|
|
|
|
Negative | 80 | 45 | 0.60 | 101 | 26 | 0.68 |
|
Positive | 28 | 19 |
| 36 | 11 |
|
| Surgical
margin |
|
|
|
|
|
|
|
Negative | 91 | 53 | 0.66 | 114 | 31 | 0.92 |
|
Positive | 15 | 11 |
| 21 | 6 |
|
| NA | 2 | 0 |
| 2 | 0 |
|
| Stage |
|
|
|
|
|
|
|
<III | 65 | 42 | 0.62 | 85 | 24 | 0.83 |
|
≥III | 41 | 22 |
| 50 | 13 |
|
| NA | 2 | 0 |
| 2 | 0 |
|
Prognostic significance of YTHDF1 and
YTHDF2 in resected HCC cases
Next, the effects of the expression levels on RFS
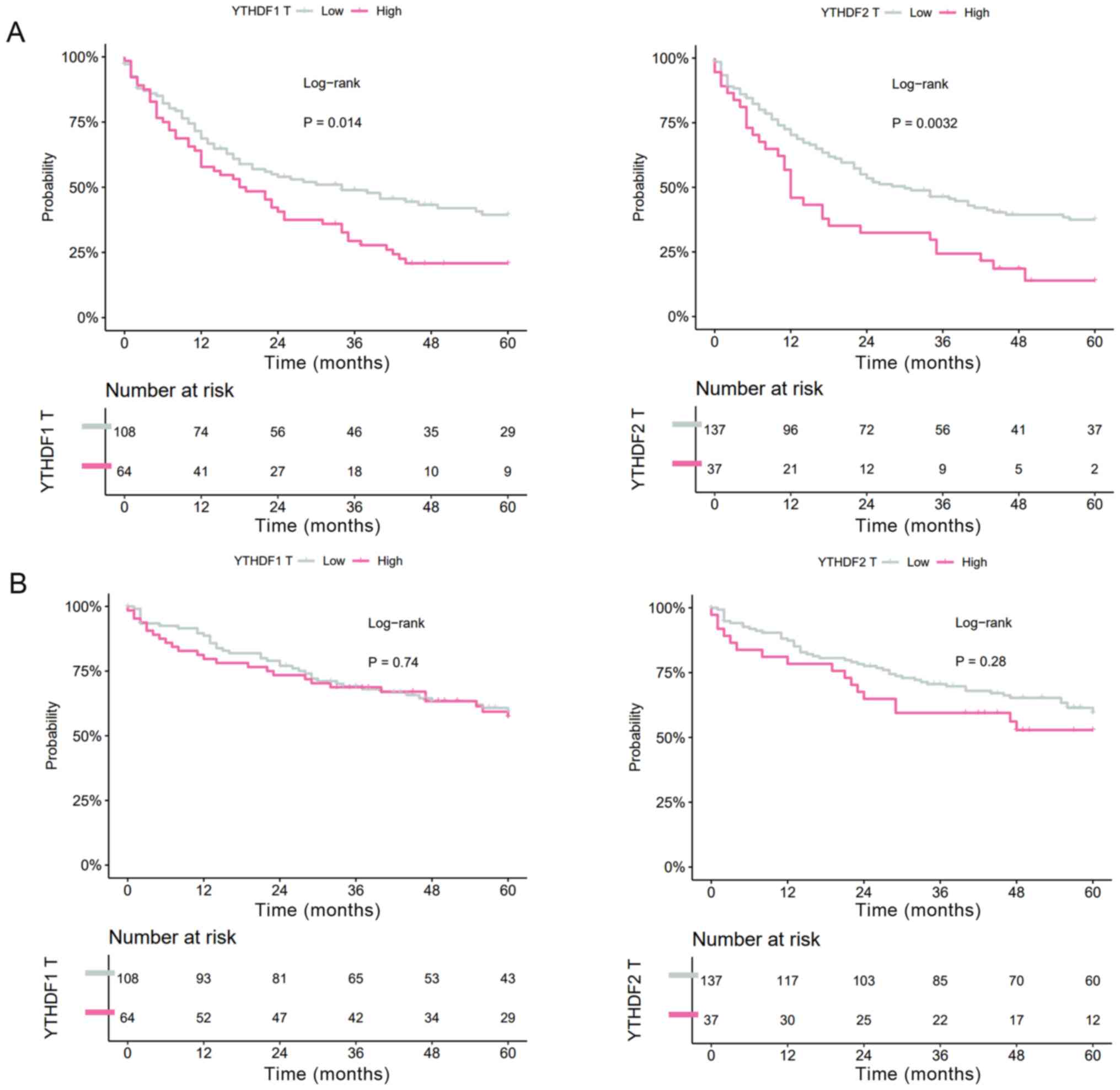
and OS were evaluated. In HCC tissues, both high YTHDF1
expression and high YTHDF2 expression were significantly
correlated with shorter RFS (YTHDF1: MST=34.0 vs. 19.0
months, P=0.014; YTHDF2: MST=30.1 vs. 12.9 months, P=0.0032,
Fig. 2A), whereas YTHDF1
expression and YTHDF2 expression were not correlated with OS
(YTHDF1: MST=99.4 vs. 70.2 months, P=0.74; YTHDF2:
MST=98.4 vs. 64.1 months, P=0.28, Fig.
2B).
YTHDF1 and YTHDF2 expression levels
and their correlation with HCC prognosis in a publicly available
dataset
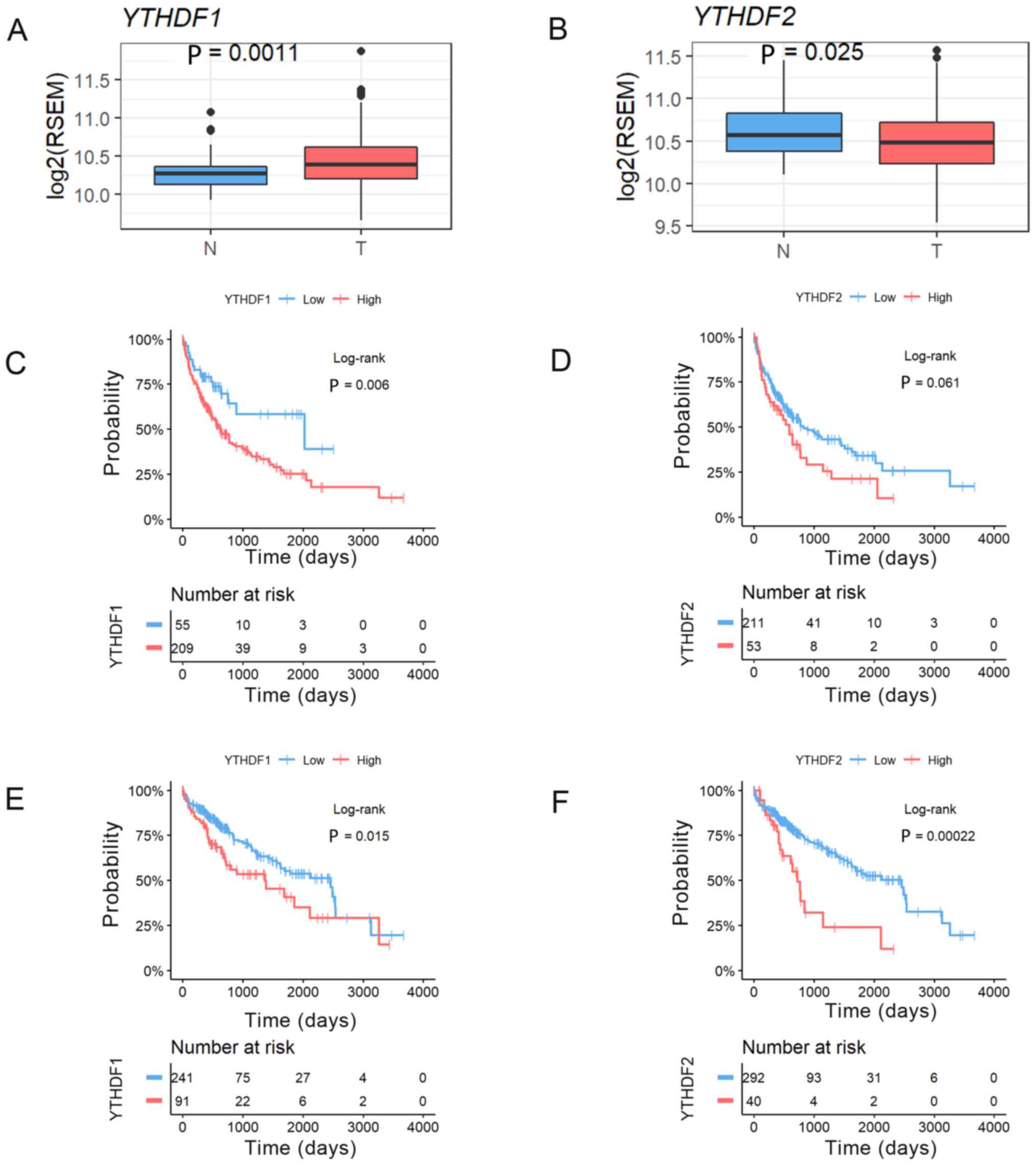
We analysed the expression levels of YTHDF1
and YTHDF2 in HCC and noncancerous tissues using a TCGA
RNA-sequence dataset. This analysis revealed that the expression of
YTHDF1 was significantly higher in HCC tumour tissues and
that YTHDF2 expression was significantly lower in HCC tumour
tissues than in noncancerous tissues (YTHDF1, P=0.0011;
YTHDF2, P=0.025, Fig. 3A and
B). We then confirmed the prognostic impact of YTHDF1
and YTHDF2 expression in resected HCC patients using the
same TCGA dataset. HCC cases were divided into two groups according
to the YTHDF1 and YTHDF2 expression in HCC tissues in
the normalized RNA-sequencing data. We also selected the cut-off
values that showed the best statistical difference. This analysis
revealed that the patients with high YTHDF1 expression had
significantly worse RFS (MST=754 vs. 489 days, P=0.006, Fig. 3C), and the patients with high
YTHDF2 expression tended to have worse RFS (MST=636 vs. 315
days, P=0.06, Fig. 3D). In addition,
the patients with high YTHDF1 expression had significantly
worse OS (MST=2456 vs. 1,372 days, P=0.015, Fig. 3E), and the patients with high
YTHDF2 expression had significantly worse OS (MST=2,456 vs.
724 days, P=0.0002, Fig. 3F).
Cox regression analysis of HCC
survival
Since the survival curves showed that YTHDF1
and YTHDF2 expression levels in HCC tissues were correlated
with RFS, we performed Cox proportional hazards analyses to further
investigate the prognostic value of YTHDF1 and YTHDF2
expression. The multivariate analysis identified serosal invasion
(hazard ratio (HR): 2.39, 95% confidence interval (95% CI):
1.30–4.42, P=0.005), portal vein or hepatic vein invasion (HR,
2.82, 95% CI: 1.26–6.28, P=0.01) and YTHDF2 expression in
HCC tissues (HR, 1.85, 95% CI: 1.09–3.15, P=0.02) as significant
independent factors for RFS (Table
III) and AFP (HR, 1.79, 95% CI: 1.10–2.92, P=0.02), serosal
invasion (HR, 1.99, 95% CI: 1.17–3.34, P=0.01), and portal vein or
hepatic vein invasion (HR, 3.02, 95% CI: 1.38–6.61, P=0.006) as
significant independent factors for OS (Table IV). Consequently, high expression of
YTHDF2 in HCC tissues was significantly associated with
recurrence after HCC surgery.
| Table III.Univariate and multivariate Cox
proportional-hazard regression analysis of recurrence free survival
in patients with hepatocellular carcinoma. |
Table III.
Univariate and multivariate Cox
proportional-hazard regression analysis of recurrence free survival
in patients with hepatocellular carcinoma.
|
| Univariate
analysis | Multivariate
analysis |
|---|
|
|
|
|
|---|
| Characteristic | HR | 95% CI | P-value | HR | 95% CI | P-value |
|---|
| Age, ≥65 vs. <65
years | 1.01 | 0.71–1.43 | 0.98 |
|
|
|
| Sex, male vs.
female | 1.27 | 0.77–2.10 | 0.35 |
|
|
|
| Virus infection,
HCV vs. others | 1.28 | 0.88–1.85 | 0.19 |
|
|
|
| Albumin, <3.5
vs. ≥3.5 g/dl | 1.74 | 1.13–2.68 | 0.01 | 1.17 | 0.59–2.35 | 0.65 |
| PT, <70 vs.
≥70% | 1.12 | 0.67–1.84 | 0.67 |
|
|
|
| ICG-R15, ≥15 vs.
<15% | 2.06 | 1.31–3.26 | 0.002 | 1.17 | 0.60–2.29 | 0.64 |
| Liver cirrhosis,
(+) vs. (−) | 1.31 | 0.91–1.88 | 0.14 |
|
|
|
| Child-Pugh
classification, B vs. A | 1.24 | 0.60–2.53 | 0.56 |
|
|
|
| Liver damage, B or
C vs. A | 1.97 | 1.26–3.08 | 0.003 | 1.91 | 0.88–4.14 | 0.10 |
| Tumour number,
multiple vs. solitary | 1.61 | 1.07–2.42 | 0.02 | 1.44 | 0.66–3.17 | 0.35 |
| Tumour size, ≥2 vs.
<2 cm | 1.73 | 0.97–3.09 | 0.06 |
|
|
|
| AFP, ≥20 vs. <20
ng/ml | 1.46 | 1.02–2.08 | 0.04 | 1.43 | 0.88–2.34 | 0.15 |
| Differentiation,
poor vs. good/moderate | 1.58 | 0.85–2.94 | 0.15 |
|
|
|
| Growth form,
infiltrative vs. expansive | 1.49 | 0.92–2.42 | 0.10 |
|
|
|
| Formation of
capsule, (−) vs. (+) | 1.27 | 0.84–1.91 | 0.25 |
|
|
|
| Infiltration to
capsule, (+) vs. (−) | 1.06 | 0.74–1.51 | 0.77 |
|
|
|
| Septal formation,
(−) vs. (+) | 1.00 | 0.68–1.47 | 0.99 |
|
|
|
| Serosal invasion,
(+) vs. (−) | 2.00 | 1.33–3.02 | 0.0009 | 2.39 | 1.30–4.42 | 0.005 |
| Portal vein or
hepatic vein invasion, (+) vs. (−) | 2.36 | 1.57–3.54 | <.0001 | 2.82 | 1.26–6.28 | 0.01 |
| Surgical margin,
(+) vs. (−) | 1.32 | 0.81–2.13 | 0.26 |
|
|
|
| Stage, III/IV vs.
I/II | 1.46 | 1.01–2.10 | 0.04 | 0.65 | 0.28–1.51 | 0.32 |
| YTHDF1 expression,
high vs. low | 1.60 | 1.11–2.31 | 0.01 | 1.37 | 0.83–2.27 | 0.21 |
| YTHDF2 expression,
high vs. low | 1.82 | 1.20–2.76 | 0.004 | 1.85 | 1.09–3.15 | 0.02 |
| Table IV.Univariate and multivariate cox
proportional-hazard regression analysis of overall survival in
patients with hepatocellular carcinoma. |
Table IV.
Univariate and multivariate cox
proportional-hazard regression analysis of overall survival in
patients with hepatocellular carcinoma.
|
| Univariate
analysis | Multivariate
analysis |
|---|
|
|
|
|
|---|
| Characteristic | HR | 95% CI | P-value | HR | 95% CI | P-value |
|---|
| Age, ≥65 vs. <65
years | 1.34 | 0.87–2.06 | 0.18 |
|
|
|
| Sex, female vs.
male | 1.01 | 0.57–1.80 | 0.96 |
|
|
|
| Virus infection,
HCV vs. others | 1.32 | 0.84–2.07 | 0.23 |
|
|
|
| Albumin, <3.5
vs. ≥3.5 g/dl | 1.75 | 1.05–2.92 | 0.03 | 1.34 | 0.63–2.85 | 0.45 |
| PT, <70 vs.
≥70% | 1.51 | 0.87–2.61 | 0.14 |
|
|
|
| ICG-R15, ≥15 vs.
<15% | 1.69 | 0.95–2.98 | 0.07 |
|
|
|
| Liver cirrhosis,
(+) vs. (−) | 1.38 | 0.90–2.13 | 0.14 |
|
|
|
| Child-Pugh
classification, B vs. A | 1.33 | 0.58–3.06 | 0.50 |
|
|
|
| Liver damage, B or
C vs. A | 2.08 | 1.24–3.49 | 0.005 | 1.73 | 0.80–3.75 | 0.16 |
| Tumour number,
multiple vs. solitary | 1.86 | 1.16–2.97 | 0.009 | 1.63 | 0.81–3.29 | 0.17 |
| Tumour size, ≥2 vs.
<2 cm | 1.78 | 0.82–3.89 | 0.14 |
|
|
|
| AFP, ≥20 vs. <20
ng/ml | 2.30 | 1.48–3,58 | 0.0002 | 1.79 | 1.10–2.92 | 0.02 |
| Differentiation,
poor vs. good/moderate | 2.02 | 1.04–3.93 | 0.04 | 1.18 | 0.50–2.76 | 0.71 |
| Growth form,
infiltrative vs. expansive | 1.69 | 0.99–2.90 | 0.05 |
|
|
|
| Formation of
capsule, (−) vs. (+) | 1.03 | 0.64–1.66 | 0.89 |
|
|
|
| Infiltration to
capsule, (−) vs. (+) | 1.10 | 0.71–1.70 | 0.66 |
|
|
|
| Septal formation,
(−) vs. (+) | 1.04 | 0.65–1.65 | 0.87 |
|
|
|
| Serosal invasion,
(+) vs. (−) | 1.90 | 1.17–3.09 | 0.008 | 1.99 | 1.17–3.34 | 0.01 |
| Portal vein or
hepatic vein invasion, (+) vs. (−) | 2.55 | 1.61–4.05 | <.0001 | 3.02 | 1.38–6.61 | 0.006 |
| Surgical margin,
(+) vs. (−) | 1.77 | 1.04–3.02 | 0.04 | 1.68 | 0.90–3.13 | 0.10 |
| Stage, III/IV vs.
I/II | 1.68 | 1.09–2.59 | 0.02 | 2.22 | 0.91–5.42 | 0.08 |
| YTHDF1 expression,
high vs. low | 1.22 | 0.78–1.90 | 0.38 |
|
|
|
| YTHDF2 expression,
high vs. low | 1.48 | 0.90–2.43 | 0.12 |
|
|
|
Discussion
In this study, we primarily evaluated the clinical
effects of m6A readers in resected HCC patients. The
expression of YTHDF1 and YTHDF2 in HCC tissues was
correlated with tumour recurrence. Furthermore, YTHDF2 was
an independent prognostic factor in resected HCC patients. Members
of the YT521-B homology (YTH) domain family, including YTHDF1,
YTHDF2, YTHDF3, YTHDC1 and YTHDC2, all have a conserved
m6A-binding domain and preferentially bind to
m6A-modified RNA at the RRm6ACH consensus sequence
(15). YTHDF2, the first
characterized m6A reader, accelerates the decay of
m6A-modified transcripts by facilitating the recruitment
of the CCR4-NOT complex directly (10). In contrast, YTHDF1 was
initially demonstrated to bind to m6A sites around the
stop codon and then cooperate with the translation initiation
machinery to improve the translation efficacy of target RNAs in
mammals (11).
We first compared the expression levels of
YTHDF1 and YTHDF2 in HCC and noncancerous liver
tissues from patients who underwent hepatectomy with curative
intent at our institution. Neither YTHDF1 expression nor
YTHDF2 expression was significantly different between HCC
tissues and noncancerous tissues. In the paired analysis, patients
with high expression in tumour tissues tended to have low
expression in noncancerous tissues. In another cohort in the TCGA
RNA-sequence dataset, however, the expression of YTHDF1 was
found to be significantly higher in HCC tissues, and YTHDF2
was significantly lower in HCC tissues. Li et al reported
that upregulation of YTHDF2 was observed in TCGA prostate cancer
tissues compared with normal controls (16). Bai et al also reported that
YTHDF1 is significantly upregulated in tumour compared with
adjacent normal tissues in colorectal cancer (17). In the TCGA dataset, there were
relatively small numbers of noncancerous tissues available, but we
studied the expression of YTHDF1 and YTHDF2 in both
tumour and noncancerous tissues from 177 HCC patients. This might
have caused the discrepancy in results between the TCGA cases and
our cases. In addition, our data showed that the reader expression
in tumour tissues was different from that in noncancerous tissues.
This change could be more important than the absolute value. In
addition, the prognostic analysis stratified by YTHDF1 and
YTHDF2 expression in our study revealed that the expression
of these two m6A readers in HCC tissues is not
associated with OS. On the other hand, high expression of both
YTHDF1 and YTHDF2 in HCC tissues was associated with
significantly worse RFS. In the public dataset, high expression of
YTHDF1 or YTHDF2 was associated with worse prognosis
than low expression. In particular, our Cox regression analysis
showed that YTHDF2 was an independent risk factor for
recurrence in resected HCC. Thus, YTHDF1 and YTHDF2
might have inherent effects in HCC carcinogenesis and influence the
long-term outcome after HCC resection, for example, by causing
sporadic recurrence.
Evidence of RNA modifications in cancer development
and progression has been increasing. The RNA methyltransferase
METTL3 is the first characterized component of the
m6A methyltransferase complex. METTL3 promotes
tumour proliferation and invasion in several cancers (18–22). The
m6A demethylases FTO and ALKBH5 were
identified in the 2010s. FTO and ALKBH5 also play an
important role in human cancer (23–25).
However, the functions of YTHDF1 and YTHDF2 in HCC
have not been uncovered. Zhao et al reported that
YTHDF1 played a vital role in the regulation of HCC
metabolism (26). Qu et al
reported that m6A RNA methylation modulators, including
YTHDF1, affected OS in HCC patients (27). YTHDF2 was able to degrade both
tumour promoter and suppressor gene mRNAs. Zhang et al
reported that YTHDF2 promotes the cancer stem cell liver
phenotype and cancer metastasis by modulating the m6A
methylation of OCT4 mRNA (28). In
contrast, YTHDF2 may act as a tumour suppressor to repress
cell proliferation and growth by destabilizing EGFR mRNA in HCC
(29). Further investigations are
required to reveal the role of YTHDF1 and YTHDF2 in
HCC.
Although we showed important aspects of
YTHDF1 and YTHDF2, there are some inherent
limitations to the present study. First, more data are necessary
because we used specimens from a single institute in this study.
Second, more detailed molecular mechanisms through which specific
m6A methylation enhances HCC development need to be
discovered. Further investigation is necessary before the clinical
utility of our findings can be determined.
In conclusion, our study revealed that high
YTHDF2 expression in HCC tissues is related to cancer
recurrence. Our results may pave the way for discovering the
clinical utility of m6A methylation and associated genes
in HCC therapy in the future.
Acknowledgements
The authors would like to thank Dr Raju Kandimalla
(Bristol Myers Squibb, San Diego, CA, USA) for useful discussions
and Ms. Yoko Nishikawa (Nagoya University Graduate School of
Medicine, Nagoya, Japan) for technical assistance with the
experiments.
Funding
The present study was partly supported by the Japan
Society for the Promotion of Science KAKENHI Grant-in-Aid for
Scientific Research (grant nos. 18H06176 and 19K18110).
Availability of data and materials
The datasets used and/or analysed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
NN, FS and YK conceived and designed the study. FS
and YK provided financial support. SY and YK provided
administrative support. SY, FS, YS, YI, HT, MH, MKa, CT, GN and MKo
provided study materials and patients. SY, FS, YS, YI, HT, MH, MKa,
CT, GN and MKo assisted with analyses and manuscript preparation.
NN, FS, KT and YS collected and assembled the data. NN and KT
performed quantitative PCR data analysis and interpretation. NN, FS
and YK wrote the manuscript and confirmed the authenticity of all
the raw data. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The study and all procedures were approved by the
Institutional Review Board at Nagoya University (approval no.
2013-0295; Nagoya, Japan), and all patients provided written
informed consent. All clinical investigations were conducted in
accordance with the principles of the Declaration of Helsinki.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2020. CA Cancer J Clin. 70:7–30. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Tabrizian P, Jibara G, Shrager B, Schwartz
M and Roayaie S: Recurrence of hepatocellular cancer after
resection: Patterns, treatments, and prognosis. Ann Surg.
261:947–955. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Roundtree IA, Evans ME, Pan T and He C:
Dynamic RNA modifications in gene expression regulation. Cell.
169:1187–1200. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Adams JM and Cory S: Modified nucleosides
and bizarre 5′-termini in mouse myeloma mRNA. Nature. 255:28–33.
1975. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Camper SA, Albers RJ, Coward JK and
Rottman FM: Effect of undermethylation on mRNA cytoplasmic
appearance and half-life. Mol Cell Biol. 4:538–543. 1984.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Finkel D and Groner Y: Methylations of
adenosine residues (m6A) in pre-mRNA are important for formation of
late simian virus 40 mRNAs. Virology. 131:409–425. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gilbert WV, Bell TA and Schaening C:
Messenger RNA modifications: Form, distribution, and function.
Science. 352:1408–1412. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ma S, Chen C, Ji X, Liu J, Zhou Q, Wang G,
Yuan W, Kan Q and Sun Z: The interplay between m6A RNA methylation
and noncoding RNA in cancer. J Hematol Oncol. 12:1212019.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Allis CD and Jenuwein T: The molecular
hallmarks of epigenetic control. Nat Rev Genet. 17:487–500. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Du H, Zhao Y, He J, Zhang Y, Xi H, Liu M,
Ma J and Wu L: YTHDF2 destabilizes m(6)A-containing RNA through
direct recruitment of the CCR4-NOT deadenylase complex. Nat Commun.
7:126262016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang X, Zhao BS, Roundtree IA, Lu Z, Han
D, Ma H, Weng X, Chen K, Shi H and He C: N(6)-methyladenosine
modulates messenger RNA translation efficiency. Cell.
161:1388–1399. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chen J, Sun Y, Xu X, Wang D, He J, Zhou H,
Lu Y, Zeng J, Du F, Gong A and Xu M: YTH domain family 2
orchestrates epithelial-mesenchymal transition/proliferation
dichotomy in pancreatic cancer cells. Cell Cycle. 16:2259–2271.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Nishizawa Y, Konno M, Asai A, Koseki J,
Kawamoto K, Miyoshi N, Takahashi H, Nishida N, Haraguchi N, Sakai
D, et al: Oncogene c-Myc promotes epitranscriptome m6A
reader YTHDF1 expression in colorectal cancer. Oncotarget.
9:7476–7486. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Shi Y, Fan S, Wu M, Zuo Z, Li X, Jiang L,
Shen Q, Xu P, Zeng L, Zhou Y, et al: YTHDF1 links hypoxia
adaptation and non-small cell lung cancer progression. Nat Commun.
10:48922019. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang X, Lu Z, Gomez A, Hon GC, Yue Y, Han
D, Fu Y, Parisien M, Dai Q, Jia G, et al:
N6-methyladenosine-dependent regulation of messenger RNA stability.
Nature. 505:117–120. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li J, Xie H, Ying Y, Chen H, Yan H, He L,
Xu M, Xu X, Liang Z, Liu B, et al: YTHDF2 mediates the mRNA
degradation of the tumor suppressors to induce AKT phosphorylation
in N6-methyladenosine-dependent way in prostate cancer. Mol Cancer.
19:1522020. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bai Y, Yang C, Wu R, Huang L, Song S, Li
W, Yan P, Lin C, Li D and Zhang Y: YTHDF1 regulates tumorigenicity
and cancer stem cell-like activity in human colorectal carcinoma.
Front Oncol. 9:3322019. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Han J, Wang JZ, Yang X, Yu H, Zhou R, Lu
HC, Yuan WB, Lu JC, Zhou ZJ, Lu Q, et al: METTL3 promote tumor
proliferation of bladder cancer by accelerating pri-miR221/222
maturation in m6A-dependent manner. Mol Cancer. 18:1102019.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lin S, Choe J, Du P, Triboulet R and
Gregory RI: The m(6)A Methyltransferase METTL3 promotes translation
in human cancer cells. Mol Cell. 62:335–345. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Miao W, Chen J, Jia L, Ma J and Song D:
The m6A methyltransferase METTL3 promotes osteosarcoma progression
by regulating the m6A level of LEF1. Biochem Biophys Res Commun.
516:719–725. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang Q, Chen C, Ding Q, Zhao Y, Wang Z,
Chen J, Jiang Z, Zhang Y, Xu G, Zhang J, et al: METTL3-mediated
m6A modification of HDGF mRNA promotes gastric cancer
progression and has prognostic significance. Gut. 69:1193–1205.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Xia T, Wu X, Cao M, Zhang P, Shi G, Zhang
J, Lu Z, Wu P, Cai B, Miao Y and Jiang K: The RNA m6A
methyltransferase METTL3 promotes pancreatic cancer cell
proliferation and invasion. Pathol Res Pract. 215:1526662019.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Li J, Han Y, Zhang H, Qian Z, Jia W, Gao
Y, Zheng H and Li B: The m6A demethylase FTO promotes the growth of
lung cancer cells by regulating the m6A level of USP7 mRNA. Biochem
Biophys Res Commun. 512:479–485. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Niu Y, Lin Z, Wan A, Chen H, Liang H, Sun
L, Wang Y, Li X, Xiong XF, Wei B, et al: RNA N6-methyladenosine
demethylase FTO promotes breast tumor progression through
inhibiting BNIP3. Mol Cancer. 18:462019. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhang S, Zhao BS, Zhou A, Lin K, Zheng S,
Lu Z, Chen Y, Sulman EP, Xie K, Bögler O, et al: m6A
Demethylase ALKBH5 maintains tumorigenicity of glioblastoma
stem-like cells by sustaining FOXM1 expression and cell
proliferation program. Cancer Cell. 31:591–606.e6. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhao X, Chen Y, Mao Q, Jiang X, Jiang W,
Chen J, Xu W, Zhong L and Sun X: Overexpression of YTHDF1 is
associated with poor prognosis in patients with hepatocellular
carcinoma. Cancer Biomark. 21:859–868. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Qu N, Qin S, Zhang X, Bo X, Liu Z, Tan C,
Wen G and Jiang H: Multiple m6A RNA methylation
modulators promote the malignant progression of hepatocellular
carcinoma and affect its clinical prognosis. BMC Cancer.
20:1652020. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhang C, Huang S, Zhuang H, Ruan S, Zhou
Z, Huang K, Ji F, Ma Z, Hou B and He X: YTHDF2 promotes the liver
cancer stem cell phenotype and cancer metastasis by regulating OCT4
expression via m6A RNA methylation. Oncogene. 39:4507–4518. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhong L, Liao D, Zhang M, Zeng C, Li X,
Zhang R, Ma H and Kang T: YTHDF2 suppresses cell proliferation and
growth via destabilizing the EGFR mRNA in hepatocellular carcinoma.
Cancer Lett. 442:252–261. 2019. View Article : Google Scholar : PubMed/NCBI
|