Introduction
Proteins must be folded into proper conformations in
order to carry out their cellular functions (1). The endoplasmic reticulum (ER) is the
site of synthesis and folding of proteins destined for either
secretion or locations in the cell membrane, Golgi apparatus,
lysosomes and elsewhere (2).
However, when protein folding in the ER is impaired due to various
physiological and pathological conditions, unfolded or misfolded
proteins can threaten cell survival. The unfolded protein response
(UPR) of the cell stresses the ER and triggers an ER
quality-control system that recognizes and removes nascent proteins
that fail to fold or assemble properly (3,4).
The UPR system is composed of two ER-associated
degradation (ERAD) systems, the ubiquitin/proteasome and the
autophagy/lysosome (5). Fujita
et al (5) used
dystrophy-associated fer-1-like protein (dysferlin) degradation to
investigate two ERAD models and found that when misfolded dysferlin
aggregated on the ER membrane excessively, the cell chose the
autophagy/lysosome ERAD system rather than the ubiquitin/proteasome
(6,7). It is known that autophagy is a
self-digestion process that degrades intracellular structures in
response to stresses, whose purpose is cell survival. However, if
autophagy was prolonged, it leads to cell death (8). Therefore, the autophagy/lysosome ERAD
system may open a door for purposively causing cell death.
In recent years, the relationship between the
intracellular aggregation of unfolded or misfolded proteins and ER
stress has been intensively examined (9). However, there are few reports in the
cell biological literature regarding ER stress and autophagy
induced by a heterologous protein. In this study, we found that
recombinant Lz-8 (rLz-8), a protein from the fruiting body of the
bracket fungus Ganoderma lucidum, aggregated on the ER of
human gastric cancer SGC-7901 cells. Accumulation of rLz-8 induced
ER stress and autophagy, leading to massive death of cells,
independent of caspase activity. The aims of this study were to
determine how cells deal with a heterologous protein such as rLz-8
that accumulates on the ER, and the role of ER stress and autophagy
in triggering cell death.
Ganoderma lucidum, a popular medicinal
mushroom, has been widely used in traditional Chinese medicine in
many Asian countries during the past two millennia. It has been
reported to be effective in modulating immune functions, inhibiting
tumor cell growth and allergies (10) and in the treatment of chronic
hepatitis, hypertension, and hyperglycemia (11). G. lucidum polysaccharide and
triterperoid were the major bioactive substances until
immunomodulatory proteins, Lz-8, was isolated and purified from the
mycelia of G. lucidum in 1989 (12–14).
Previously, we demonstrated the crystal structure of Lz-8 which was
a noncovalently linked homodimer with an apparent molecular weight
of 24 kDa. Each monomer consists of 110 aa residues with an
acetylated N terminus and a molecular mass of 12 kDa (15). Liao et al also reported that
reFIP-gts, another immunomodulatory protein from the closely
related Ganoderma tsugae, inhibited the growth of A549 cancer cells
significantly and selectively (16). However, until now there have been no
studies demonstrating how Lz-8 induces cell death and the
mechanisms involved in this process.
Herein we report for the first time that an
excessive heterologous protein aggregation of rLz-8 from
Ganoderma lucidum on the ER of human cancer cells induces
autophagy-dependent cell death, not a caspase-dependent cell death
or apoptosis, and provides a novel strategy for cancer
treatment.
Materials and methods
Recombinant plasmid construction and
Pichia pastoris transformation
The total DNA of G. lucidum was extracted as
described by Al-Samarrai and Schmid (17). The rLz-8 gene was amplified from the
total DNA sample by PCR, and SnaBI and EcoRI
restriction sites were designed for flanking the PCR product at the
5′- and 3′-termini, respectively. To build the expression plasmid,
the PCR fragment was cloned into the SnaBI/EcoRI site
of pPIC9K (Invitrogen, CA, USA) and transformed into Pichia
pastoris Gs115 (Mut+; Invitrogen).
Media and culture conditions for rLz-8
expression
The P. pastoris transformants were cultured
in a 100 ml flask containing 1000 ml buffered minimal glycerol-rich
yeast (BMGY) medium, supplemented with 1% (v/v) glycerol as a
carbon source and 200 μg/ml G418 (Geneticin) as a selection
pressure. Cells were grown at 28˚C and shaken at 300 rpm until an
OD600 value reached 15 making a 10-fold dilution. The
cells were then harvested by centrifugation at 3000 × g and 4˚C for
5 min. BMGY was replaced by buffered minimal methanol-rich yeast
(BMMY) medium containing 0.5% (v/v) methanol. Methanol was used to
induce the AOXI promoter. BMGY and BMMY media were also prepared
according to the manufacturer's instructions (Invitrogen). After 48
h of induction, the supernatant was collected by centrifugation at
4˚C and 12,000 × g for 20 min. rLz-8 protein was purified by a
nickel affinity column Ni-sepharose (GE Lifescience, USA) and
eluted by a gradient of 30–100 mM imidazol.
Cell culture and materials
Cells of the human gastric cancer cell line SGC7901
were obtained from the Institute of Biochemistry and Cell Biology
(Shanghai, China). Cells were grown in Dulbecco's modified Eagle's
medium (DMEM; Invitrogen) supplemented with 2 mM L-glutamine, 100
U/ml penicillin, 100 μg/ml streptomycin, and 10% fetal bovine serum
(FBS) at 37˚C in a humidified atmosphere of 95% air and 5%
CO2. Anti-ATF4 antibody was purchased from Proteintech
Group (PTG, USA). Anti-CHOP, Anti-LC3, Anti-GAPDH antibodies and
secondary antibody were purchased from Santa Cruz Biotechnology
(Santa Cruz, Canada). Tm, TG and 3-MA were obtained from
Sigma-Aldrich (USA).
Electron microscopy
SGC7901 cells were double fixed in PBS-buffered
glutaraldehyde (2.5%) and osmium tetroxide (1%), dehydrated, and
embedded using the EMBed-812 kit (EMS, USA) according to the
manufacturer's instructions. Ultrathin sections (70 nm) were made
and double stained with uranyl acetate and lead citrate, and viewed
in a transmission electron microscope (Hitachi H-7500, Japan).
Western blotting
Cells were washed with PBS and lysed in Lysis-M
Reagent supplemented with complete mini-protease inhibitor cocktail
tablets (Roche, Indianapolis, IN, USA). The lysates were then
incubated for 5 min at room temperature with gentle shaking, and
centrifuged at 14,000 × g for 10 min. Equal amounts of protein were
subjected to 12% sodium dodecyl sulfate polyacrylamide gel
electrophoresis and then transferred to polyvinylidene difluoride
membranes (Millipore, USA). The membranes after blocking in 5%
skimmed milk powder in TBS were incubated with each primary
antibody followed by a peroxidase conjugated anti-rabbit secondary
antibody. The corresponding bands were detected using an ECL
Advance kit (GE Biosciences, USA).
Real-time quantitative PCR
The total RNA was isolated using Trizol (Invitrogen)
from cells after treatment with or without rLz-8 for the indicated
times. cDNA was synthesized using a reverse transcription system
(Takara, Japan). The real-time PCR was performed with the use of
LineGene 3320 Bioanalyzer (Bioer, China) following the
manufacturer's instructions. SYBR Green I kit (Takara) was used.
PCR conditions were 95˚C for 30 sec followed by 40 cycles of 95˚C
for 5 sec and 60˚C for 20 sec. Bioer 3320 software was used to
analyze the standards and to carry out the quantification. The
relative quantity of each mRNA was normalized to the relative
quantity of GAPDH mRNA. ATF4 sense primer: 5′-TCTCC AGCGACAAGGCT
AAG-3′; ATF4 anti-sense primer: 5′-TACCCAACAGGGCATC CAAG-3′; CHOP
sense primer: 5′-GAACCAGGAAACGGAA ACAG-3′; CHOP anti-sense primer:
5′-CATTCACCATTCGG TCAATCA-3′; LC3 sense primer: 5′-AGCAGCATCCAACC
AAAATC-3′; LC3 anti-sense primer: 5′-CTGTGTCCGTTCA CCAACAG-3′;
GAPDN sense primer: 5′-GCACCGTCAAGG CTGAGAAC-3′; GAPDH anti-sense
primer: 5′-ATGGTGGTG AAGACGCCAGT-3′.
Cell proliferation assays
The inhibition of cell proliferation and viability
in SGC7901 cells was determined using the WST-1 (Roche) assay.
Cells were placed at 8,000 per well in 96-well plates in their
respective growth medium with FBS reduced to 5%. The cells were
allowed to grow for 24 h and then treated with different drugs.
After 24 h, WST-1 reagent was added to the plates according to
manufacturer's protocol, and absorbance was read at 450 nm with an
ELISA reader (Tecan).
Confocal microscopy
ERtracker, mitotracker and lysotracker (Invitrogen)
was used for the assessment of cellular lysosomal content,
according to the manufacturer's instructions. Briefly, cells were
cultured on 24-well plates and treated with rLz-8- fluorescein
isothiocyanate (FITC) or rLz-8 (5 μg/ml). After several time
courses, cells were incubated with 1 μM ER tracker, 300 nM
mitotracker, or 75 nM Lysotracker for 45 min at 37˚C in 5%
CO2, and nuclear stained with 1 μg/ml Hoechst33342
(Sigma, USA) for 15 min. Cell were washed thrice with fresh medium
and were visualized using an Olympus laser scanning confocal
fluorescence microscope fitted with the appropriate filters. The
results of Lysotracker fluorescence were selected, five fields of
view (512×512 pixel) per section, and fluorescence images were
analyzed using ImageJ software (NIH).
Measurement of caspase activity
Caspase-3 activity in SGC7901 cells was measured
with a NucView 488 Caspase-3 Assay Kit for live cells according to
the manufacturer's instructions (Biotium, Hayward, CA, USA).
Briefly, the cells were grown in 24-well plates at a density of
1×105 cells per well. After rLz-8 treatment for 24 h,
cells were incubated with 3 μm NucView 488 Caspase-3 substrate in
medium for 30 min. Cells (200/group) were visualized using a
fluorescence microscope fitted with the appropriate filters. For
the inhibition of caspases, cells were grown in 96-well plates.
After 24 h, the cells were pre-incubated with 50 μM z-VAD-fmk (a
pan-caspase inhibitor) for 30 min and then treated with rLz-8 for
48 h. The results were measured with the WST-1 assay.
Cell cycle analysis
Cells were placed into 6-well plates at a density of
5×105 cells per well and incubated with media or rLz-8
at 5 μg/ml for the indicated times. Cells were harvested by
centrifugation at appropriate times and fixed with 70% ethanol at
4˚C overnight. After fixation, cells were washed by centrifugation
with PBS and resuspended in 1 ml PI staining buffer (0.1% Triton
X-100, 100 μg/ml RNase A, 50 μg/ml PI in PBS) at 37˚C for 30 min.
The cell cycle phase was analyzed using a flow cytometer (Becton
Dickinson, USA) with Cellquest software.
Assessment of apoptosis
The effects of rLz-8 on nuclear morphology in cells
were examined by staining with Hoechst 33342 (Sigma). In brief,
cells (1×105 cells/well; 24 wells) were incubated with
rLz-8 (5 μg/ml) in the growth medium for 24 h at 37˚C. The cells
were stained with Hoechst 33342 (1 μg/ml) for 15 min and washed
twice with PBS. The images were observed under a fluorescence
microscope (Olympus X71, Japan).
Evaluation of apoptotic cell death was performed
with the Annexin V-FITC Apoptosis Detection kit (Bipec Biopharma,
USA) according to the manufacturer's recommendations. In brief,
after the treatment, cells were washed twice with cold PBS and
collected by centrifugation. The pellet was resuspended in 1X
binding buffer and stained with FITC-labeled Annexin V (5 μl) for
15 min at 4–8˚C in the dark. PI (10 μl) was then added for 5 min at
4–8˚C in the dark. Cells were analyzed using a flow cytometer
(Becton Dickinson) with Cellquest software.
Mitochondrial membrane potential
Mitochondrial membrane potential was examined by
JC-1, according to the manufacturer's instructions. Briefly, cells
were placed on 6-well plates and treated with rLz-8 (5 μg/ml).
After 24 h, cells were incubated with 10 μg/ml JC-1 for 30 min at
37˚C in 5% CO2 and then analyzed using a flow
cytometer.
Small interfering RNA (siRNA)
The ATF4 siRNA or CHOP siRNA duplexes used in this
study were purchased from Santa Cruz Biotechnology. SGC7901 cells,
plated 1 day earlier on 6-well culture dishes to achieve 50–60%
confluence, were transfected with siRNAs (80 pM) by using 6 μl of
Lipofectmine 2000 (Invitrogen). After 24 h of transfection, cells
were treated with rLz-8 for up to 24 h.
Statistical analyses
Statistical analysis was conducted using Student's
t-test and one-way ANOVA. The data were presented as mean ±
standard error (SE). Experiments were performed at least three
times. Results were considered to be statistically significant at
P<0.05.
Results
rLz-8 inhibits cell growth and induces
cell death in human gastric cancer cell line SGC-7901
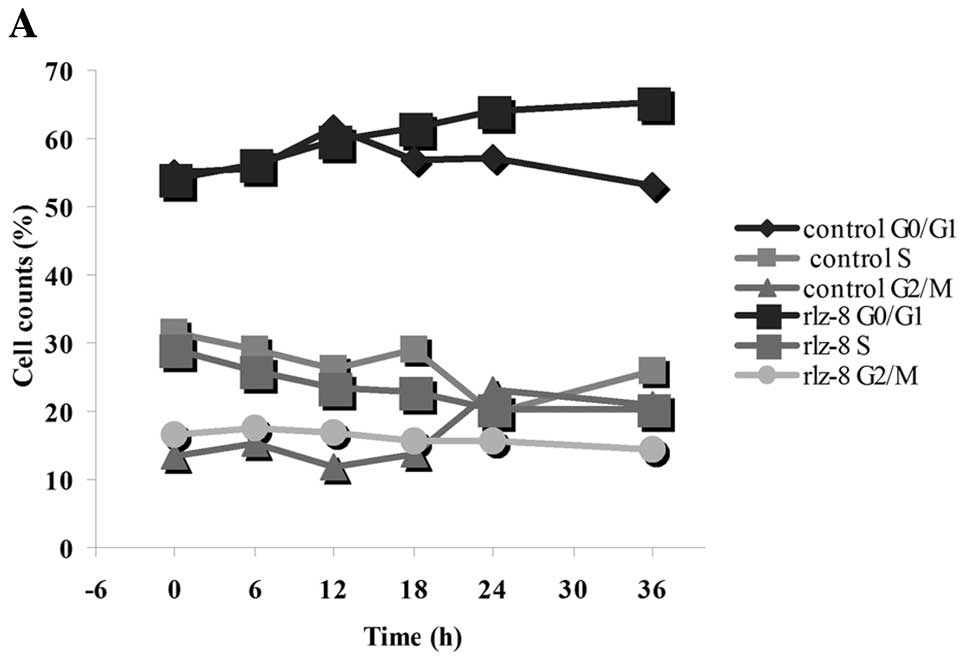
rLz-8 caused a detectable accumulation of nuclei in
G1 phase with a concomitant decrease of nuclei in S phase. This was
most evident after 12, 18, 24 and 36 h (Fig. 1A). After 12, 18, 24 and 36 h of
stimulation with 5 mg rLz-8, staining of cells of the gastric cell
line SGC-7901 with DNA-specific fluorochrome propidium iodide (PI)
showed 59.8, 61.7, 64 and 65.2% of SGC-7901 cells with G0/G1 DNA
content, while 23.3, 22.8, 20.3 and 20.4% of cells were in S-phase,
respectively.
Furthermore, treatment of SGC-7901 cells with rLz-8
for 24, 36, 48 and 72 h at different concentrations (0.5, 2.5, 5.0,
20, 50 μg/ml) also resulted in cell growth inhibition. Slight
inhibition of proliferation was detected in cells exposed to 0.5
μg/ml of rLz-8, whereas cell proliferation was markedly inhibited
in cells treated with 2.5 or 5 μg/ml. Treatment with 50 μg/ml rLz-8
for up to 72 h significantly reduced the number of viable human
gastric cancer SGC-7901 cells (Fig.
1B). All results of the cell viability and proliferation assay
WST-1 showed that rLz-8 inhibited SGC-7901 cell proliferation in a
dose- and time-dependent manner, with IC50 at 24 h of 5
μg/ml.
rLz-8 accumulating in nucleus triggers
endoplasmic reticulum stress (ERS)-mediated cell death
The underlying molecular mechanism governing the
effects of rLz-8 on the inhibition of SGC-7901 cell proliferation
is still unclear. The first step in examining the possible roles of
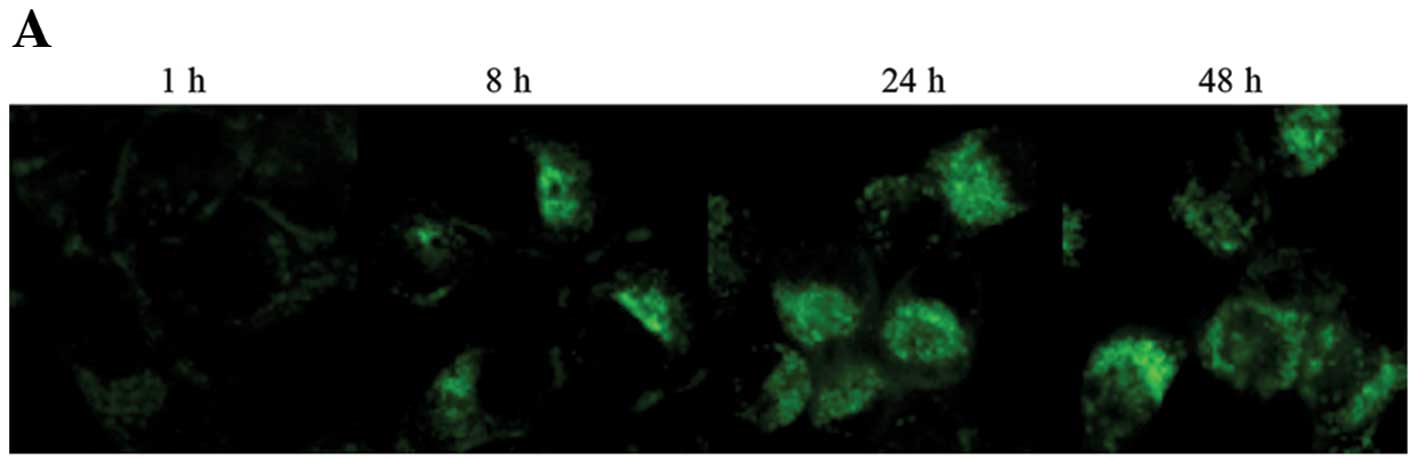
rLz-8 was to determine its cellular localization. Colocalization
studies were performed using specific markers for the cellular
organelles and rLz-8 in SGC-7901 cells. As shown in Fig. 2A and B, fluorescein isothiocyanate
(FITC)-labeled rLz-8 and the ER's specific marker overlapped,
suggesting that rLz-8 localized at the ER, into which it
diffuses.
In the following studies, we examined whether rLz-8
triggered ER stress as it accumulated in the ER by determining the
effects of rLz-8 on the expression of classical ER stress markers,
such as the transcription factors CHOP/GADD153 and ATF4. When ER
stress occurs, a large number of unfolded proteins accumulate in
the ER, GRP78 dissociates from protein kinase RNA-like ER kinase
(PERK) by binding these proteins. Unbound PERK oligomerizes and is
activated by trans-autophosphorylation. Activated PERK
phosphorylates eIF2α, then its phosphorylation promotes translation
of ATF4, escalating ER stress further. ATF4 can function as a
homodimer or heterodimer with members of the binding protein family
of transcription factors, such as the cell death protein
CHOP/GADD153, and also promotes translation of CHOP/GADD153
(18,19). Real-time quantitative RT-PCR
(qRT-PCR) analyses showed that rLz-8 induced CHOP (4- to 9-fold),
ATF4 (1- to 2-fold) and GRP78 (3- to 4-fold) mRNA 3- to 10-fold in
SGC-7901 cells at 5 and 20 μg/ml (Fig.
2C), and the expression of ATF4 was confirmed by Western
blotting (Fig. 2D). However, in the
same test for expression of CHOP, we did not observe positive
results. Leena et al (20)
reported that some commercially available CHOP antibodies were
non-specific for detecting CHOP in Western blotting and
hypothesized that antibody specificity may influence the
results.
To determine the role of ER stress in the
rLz-8-induced death of SGC-7901 cells, we transiently transfected
cells with siRNA duplexes specific for CHOP and ATF4. Results
showed that CHOP or ATF4 knockdown cells treated with rLz-8 had
higher survival rates than the control cells (SCR; Fig. 3E). Therefore, rLz-8 serves as an
inducer for death in SGC-7901 cells as it diffuses into the ER and
triggers ER stress.
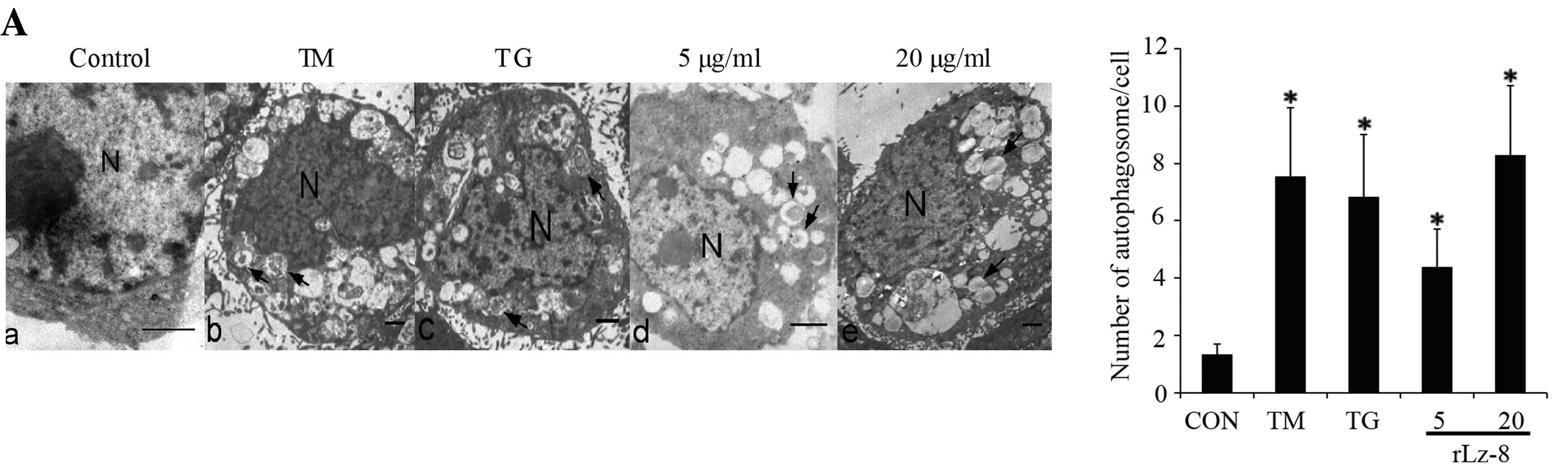 | Figure 3Induction of autophagy in SGC7901
cells treated with rLz-8. (A) Electron micrographs were taken of
SGC7901 cells untreated (a) or treated (b) with 2 μg/ml tunicamycin
(TM) for 24 h, (c) 300 nM thapsigargin (TG) for 24 h, (d) 5 μg/ml
and (e) 20 μg/ml rLz-8 for 48 h. Double membrane vacuoles are
denoted by an arrow, and the nucleus is denoted by N. Scale bar: 1
μm. The histogram is the numbers of autophagosomes in SGC7901 cells
after exposure to the indicated stimuli for 24 h. A total of 25
electron microscopic sections were prepared. Standard error
represents 3 independent experiments. P-value represents the
significant difference between conditions at *P<0.05.
(B) Protein levels of LC3B (15 kDa) were measured by immunoblot
analysis. SGC7901 cells were treated for 24 h with 300 nM
thapsigargin (TG), 0.5, 5, and 20 μg/ml rLz-8. (C) Quantitative
analysis of intensities of Western blotting. CON, control. Standard
error represents 3 independent experiments. P-value represents the
significant difference between conditions at *P<0.05.
(D) Gene expression of LC3B induced by rLZ-8. SGC7901 cells treated
with 0.5, 5, and 20 μg/ml rLz-8 for 24 h. Relative mRNA levels were
measured by real-time qPCR, n=3. CON, control. (E) Induction of
lysosomes in SGC7901 cells treated with rLz-8. SGC7901 cells were
treated with 5 μg/ml of rLz-8 for 24 h. Nuclear stained with
Hoechst 33342 (blue) and Lysotracker (red) staining are shown. The
histogram is quantitative analysis of fluorescence intensities.
Scale bar: 10 μm. (F) Protein levels of LC3B (15 kDa) were measured
by immunoblot analysis. Cells were RNAi against control (SCR), ATF4
and CHOP. After 24 h, cells were treated with 5 μg/ml rLz-8 for 24
h. (G) Quantitative analysis of intensities of Western blotting.
CON, control. Standard error represents three independent
experiments. P-value represents the significant difference between
conditions at *P<0.05. (H) Gene expression of LC3B
with siRNA. Cells were RNAi against control (SCR), ATF4 and CHOP.
Cells were treated 24 h later with 5 μg/ml or 20 μg/ml rLz-8 for 24
h. Relative mRNA levels were measured by real-time qPCR, n=3. |
The rLz-8-triggered ER stress induces an
autophagic response in SGC-7901 cells
Considering the role of ATF4 in ER stress and
autophagy, we next investigated whether rLz-8 induced an autophagic
response in SGC-7901 cells. Electron microscopy remains one of the
most accurate methods for the detection of autophagy and
quantification of autophagic accumulation. Firstly, autophagic
bodies and autophagosomes with double or single membrane structures
in the cytoplasm were examined by electron microscopy for
autophagic topical changes. In Fig.
3Aa, the electron micrograph depicts the intact cell and
nuclear membrane, rough endoplasmic reticulum, distinct nucleus,
and homogeneous cytoplasm of normal SGC7901 cells (control).
SGC7901 cells treated with different doses of rLz-8 are shown in
Fig. 3Ad and e. Membrane-bound
autophagosomes in the cytoplasm are observed. The nucleus is
condensed, but the nuclear membrane is intact and distinct.
Tunicamycin (TM) and thapsigargin (TG) are triggers of the
autophagic ER stress-induced response in cells. As shown in the
electron micrograph in Fig. 3Ab and
c, SGC7901 cells treated with TM and TG show an abnormally
large increase in the number of autophagosomes, condensed nuclei,
and damaged nuclear membranes. The number of autophagosomes was
significantly increased in cells treated with these stimuli,
compared to non-treated cells (Fig.
1B).
Microtubule-associated protein 1 light chain 3
(MAP1-LC3) is essential for autophagy and is associated with
autophagosome membranes. MAP1-LC3 has two subunits, specifically
MAP1-LC3A (MAP1-LC3α) and MAP1-LC3B (MAP1-LC3β) (21). There are two forms of LC3B, the
cytosolic LC3-I and the membrane-bound LC3-II. During autophagy,
LC3B is synthesized in large quantities. LC3-I is formed by the
removal of the C-terminal 22 amino acids from newly synthesized
LC3B, and a fraction of LC3-I is converted into LC3-II (22,23).
Therefore, LC3B is thought to be an important marker for autophagy.
During rLz-8 treatment, the protein and mRNA levels of LC3B
increased significantly in a dose- and time-dependent manner
(Fig. 3B and C), and there was an
increase in the processed form of LC3B, which was indicative of
increased autophagy. The result of real-time reverse-transcription
PCR analysis showed that for the negative control (CON), positive
control (TG) and rLz-8-treated groups (0.5, 5 and 20 μg/ml), mRNA
levels of LC3B were up-regulated 1.8-, 3.9-, and 4.2-fold,
respectively.
During rLz-8 treatment, the protein and mRNA levels
of LC3, a recognized marker, increased significantly in a dose- and
time-dependent manner (Fig. 3D),
and there was an increase in the processed form of LC3. This was
indicative of increased autophagy, particularly with 5 and 20
μg/ml, between the negative control (CON), positive control (TM,
TG) and rLz-8-treated groups.
When autophagy is induced in old or unwanted
organelles, proteins and microbes are delivered to the lysosome,
accompanied by a rapid increase in the number of lysosomes.
Lysotracker, a lysosome-specific dye, was used to examine the
number of lysosomes in rLz-8-treated SGC-7901 cells. Results
indicated that compared to the negative control group, cells
treated with rLz-8 at 5 μg/ml for 24 h demonstrated a significant
increase in lysosomal mass (Fig.
3E).
The relationship between autophagy and ER-stress was
investigated by using siRNA duplexes specific for the ER-stress
markers, CHOP and ATF. When CHOP and ATF4 were rendered
dysfunctional, the LC3B protein and mRNA levels were greatly
reduced compared to the control siRNA-transfected cells (Fig. 3F–H). Hence, we suggest that
ER-stress is the main factor triggering the autophagy response.
The role of autophagy in the death of
SGC7901 cells treated with rLz-8
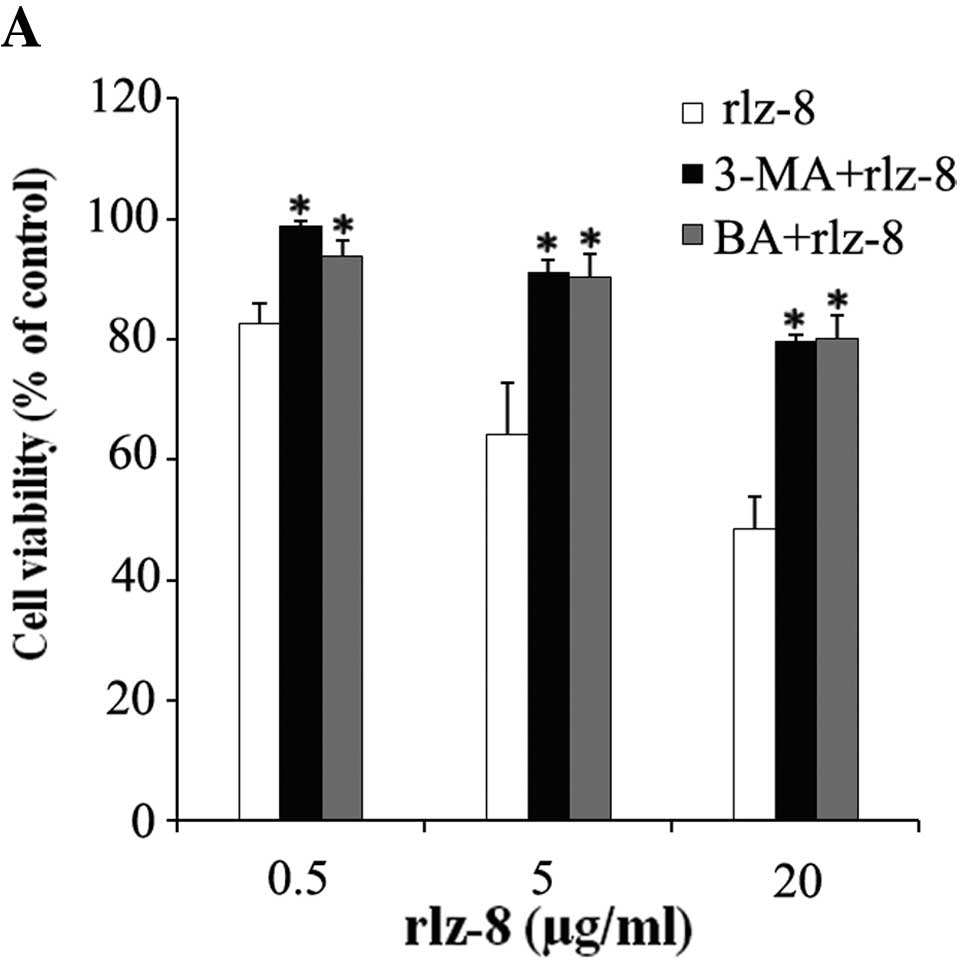
Over the years many studies have proven that
autophagy plays a dual role in the biological process of cell
death. To examine whether the autophagy induced by rLz-8 plays any
roles in cell survival or death, cells in which autophagy was
blocked by 3-methyladenine (3-MA; a specific inhibitor of
autophagic/lysosomal protein degradation) were treated with rLz-8.
A colorimetric WST-1 assay was used to determine the number of
viable cells. Results showed that cell death that would otherwise
be induced by rLz-8 was inhibited: the number of rLz-8-treated
living cells was more than the control that was not treated with
3-MA (Fig. 4A). We next transiently
transfected cells with siRNA duplexes specific for LC3B, then
detected the survival rate of LC3B knockdown cells treated with
rLz-8 for 24 or 48 h. Compared with the non-siRNA LC3B control in
which expression of LC3B was inhibited, the rLz-8 killing effect
declined rapidly (Fig. 4D). Hence,
this suggests that autophagy plays a crucial role in the process of
cell death induced by rLz-8.
Caspase inhibitors do not prevent
rLz-8-induced cell death
When apoptosis occurs, significant morphological
changes such as chromatin condensation, nuclear shrinkage, and the
disappearance of the nuclear envelope and nucleolus may be seen in
cells. To investigate the morphological changes of organelles in
rLz-8-treated SGC-7901 cells, laser scanning confocal microscopy
was used to analyze Hochest 33342-stained nuclei. As shown in
Fig. 2A, normal cells have a
beam-shaped nucleus, smooth nuclear envelope, and non-condensed
zones of chromatin. The cancer cell line SGC-7901 was treated with
rLz-8 for 24 and 48 h at concentrations ranging from 2.5 to 10
μg/ml. Compared with normal cells, no apoptotic morphological
changes were seen in any of the rLz-8-treated groups (Fig. 2A). Most significantly, in the
high-dose rLz-8 treatment group (10 μg/ml), nearly all cells were
dead, but we did not observe, by transmission electron microscopy
(TEM), any apoptotic morphological changes that had occurred during
the process of death (Fig. 3A). We
also analyzed the apoptosis rate of the treatment groups by using
flow cytometry (Fig. 5B), but
results indicated that rLz-8 could not induce apoptosis in SGC-7901
cancer cells in a time- and dose-dependent manner.
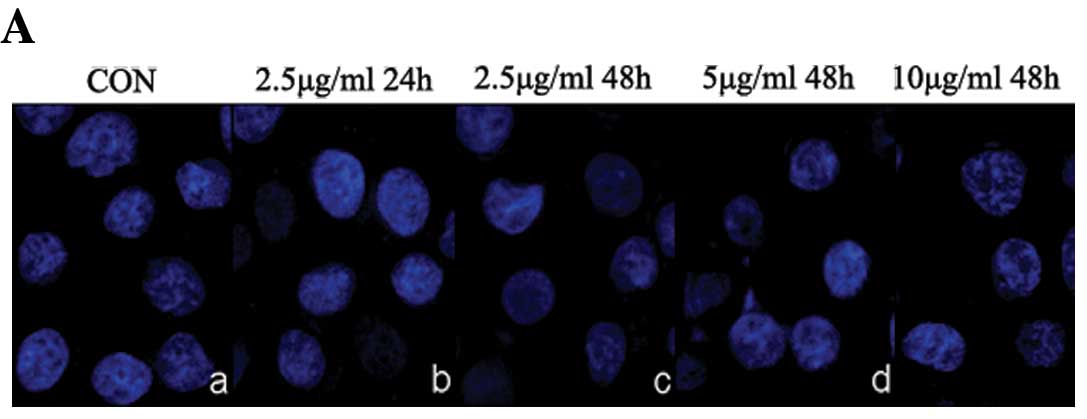 | Figure 5(A) Morphological changes of nuclear
in SGC7901 cells treated with rLz-8. SGC7901 cells untreated (a) or
treated (b–e) with 2.5, 5, or 10 μg/ml rLz-8 for several time
courses (24 h, 48 h), and then 1 μg/ml Hoechst 33342 stained
nuclear for 20 min. (B) Effect of rLz-8 on cell viability of
SGC7901 cells. AV/PI double staining was performed on SGC7901 cells
before and exposure to 2.5, 5, and 10 μg/ml rLz-8 for different
periods of time (24, 36 and 48 h).
AV-/PI+(Q1): percentage early necrotic cells;
AV+/PI+(Q2): percentage late apoptotic and
dead cells, AV−/PI−(Q3): percentage viable
cells, AV+/PI−(Q4): percentage early
apoptotic cells. (C) Effect of caspases in SGC7901 cells treated
with rLz-8. SGC7901 cells untreated or treated with 50 μm Z-VAD-FMK
for 30 min, and then treated with 0.5 to 20 μg/ml rLz-8 for 48 h.
The result measured with WST1. Standard error represents 4
independent experiments. (D) Effect of rLz-8 on mitochondrial
membrane potential of SGC7901 cells. SGC7901 cells untreated (a) or
treated (b–d) with 0.5, 5, and 20 μg/ml rLz-8 for 24 h, and then
cells were incubated with 10 μg/ml JC-1 for 30 min. |
Although it is accepted that apoptosis is mediated
by caspase activity, the specific role of caspases is debatable
(17,24–29). A
caspase inhibition assay was performed to examine the involvement
of caspase in rLz-8-induced cell death. The caspase inhibitor
z-VAD-fmk (50 mM) did not significantly affect the cell viability
of rLz-8-treated SGC-7901 cells (Fig.
5C). These results proved that rLz-8-induced cell death is
independent of caspase activation. It is well known that when
apoptosis occurs, a change in the electric potential of the
mitochondrial membrane is a typical event. Herein, flow cytometry
was used to investigate changes in mitochondrial membrane potential
in rLz-8-treated SGC7901 cells. These data together with the siRNA
assay results corroborate that rLz-8-induced cell death occurs by
autophagy.
Discussion
Disturbances in normal functions of the ER lead to
the UPR, an evolutionarily conserved cell stress response which is
aimed initially at compensating for damage but can eventually
trigger cell death if ER dysfunction is severe or prolonged. The
mechanisms by which ER stress leads to cell death remain enigmatic,
with multiple potential participants described with little clarity,
with specific death effectors dominating in particular cellular
contexts (30).
As we know, types of cell death induced by ER stress
include apoptotic cell death, necrosis, and autophagic cell death
(31). Among these, apoptotic cell
death depends greatly on caspase activity. Many important relevant
mechanisms involved in caspase-dependent cell death have already
been studied extensively (32).
In our study, rLz-8 exerted a promising killing
effect on SGC-7901 cells. We used classical methods to examine
whether caspase-independent cell death rather than
caspase-dependent apoptosis was triggered by rLz-8-induced ER
stress. The results of TEM indicated that no morphological feature
of caspase-dependent apoptosis was observed in rLz-8-treated cells,
i.e., chromatin condensation, loss of cytoplasmic processes, round
cell shape or fading of the nuclear membrane (33–36).
Moreover, when caspase families were inhibited with the pan-caspase
inhibitor z-VAD-fmk in SGC-7901 cells, we found that rLz-8 still
had the same killing effect, affirming that rLz-8-induced cell
death is not caspase-dependent, and may not even be apoptosis
(37–41).
Regarding rLz-8-induced cell death, our findings
suggest: 1) In SGC-7901 cells, rLz-8 diffuses and accumulates
gradually in the ER, reaching saturation after 24 h. rLz-8 was not
transported out of the ER or degraded until the cells died. 2)
After rLz-8 accumulated in the ER, a significant increase in the
number of lysosomes was observed, by fluorescence microscopy. The
results of TEM also confirmed dilated and vacuolized smooth ER and
an increased number of lysosomes. 3) rLz-8-treated cells showed
increased mRNA and protein levels of CHOP/GADD153 and ATF4,
respectively. 4) When rLz-8 gradually accumulated in the ER of
SGC-7901 cells, autophagy and cell death were observed. When
autophagy was inhibited by 3-MA treatment, the number of dead cells
decreased, suggesting that autophagy played a role in promoting
cell death in rLz-8 treated cells.
Therefore, how does the cell deal with the
aggregation of rLz-8 in the ER? Excess amounts of unfolded or
misfolded proteins in the ER are retrotranslocated to the cytoplasm
and degraded by the ERAD system, which includes a
ubiquitin/proteasome and an autophagy/lysosome system (15). A larger number of autophagosomes and
lysosomes appeared in rLz-8-treated cells. This suggests that the
autophagy/lysosome system was activated, and the cell may employ
this system to degrade rLz-8. In the study of Fujita et al
(5), native and mutant dysferlin
were degraded by the ubiquitin/proteasome ERAD system. However,
when mutant dysferlin was not sufficiently degraded by
ubiquitin/proteasome ERAD system, it aggregated in the ER and
stimulated autophagy formation and LC3 conversion via the ER
stress-related PERK-eIF2a-ATF4-CHOP/GADD153 pathway. This pathway
was also activated in rLz-8-treated cells. The mutant dysferlin
aggregates on the ER membrane could be recognized by the elongating
membrane that forms the autophagosome, and is then degraded by the
autolysosome upon fusion with a lysosome. Although there were no
experimental results in our study verifying that rLz-8 was
recognized and degraded by the ubiquitin/proteasome ERAD system, we
used a Bayesian discriminant method (42,43) to
predict the rLz-8 ubiquitination sites. However, specific
ubiquitination sites were not found.
In general, the ERAD system degrades aberrant
proteins to ensure cell survival, but in this study the SGC-7901
cells were killed by the accumulation of rLz-8 in the ER. This is
similar to the apoptotic death of C2C5 cells caused by dysferlin
aggregates, or protein overexpressing in yeast. It is known that
excessive ER stress, or ER-stress mediated cell death, can kill
cells but what are the roles of ERAD and autophagy in this process?
According to a recent study, autophagy formation was a cellular
defense mechanism against polyQ72-induced ER-stress-mediated cell
death, whose mechanism involved the degradation of polyQ72
aggregates (44). Similar studies
also indicated that autophagy protects cells from ER stress arising
from protein aggregations via the ERAD system (45,46).
However, we have determined that while aggregations of rLz-8
induced ER stress and triggered the ERAD system, the resulting
autophagy contributed to cell death rather than survival. Before
this, it was very rare that a heterologous protein aggregating in
the ER of a tumor cell could trigger autophagy-dependent cell
death.
Next, what role did ER stress play in the
rLz-8-induced cell death? As we know, when ER stress exceeds a
threshold it mediates many types of cell death, including classic
apoptosis, autophagic cell death and necrosis. The principal
challenge with any cellular strategy for preventing death caused by
ER stress lies with the multitude of parallel pathways that
potentially lead to downstream cell death mechanisms. Multiple
pathways linking ER stress to cell death have been reported. Of
these, ATF4-CHOP/GADD153 is the first identified protein that
mediates ER stress-induced apoptosis and much is known of the roles
of this molecule in cell death. Nevertheless, little is known of
just how ATF4-CHOP/GADD153 induces cell death, especially
considering that CHOP-deficient cells are resistant to ER
stress-mediated cell death. Our study showed that when expression
of ATF4 or CHOP was inhibited in rLz-8-treated SGC-7901 cells, dead
cells decreased significantly, and LC-3II conversion was
down-regulated. This proves that the ATF4-CHOP pathway and
ER-stress both are the key factors for inducing cell death, and
also that the ATF4-CHOP pathway acts as a bridge between ER stress
and autophagy.
In conclusion, rLz-8 aggregation induces autophagic
cell death and provides a novel mechanism for protein-based
anti-cancer medicine. This may lead to new strategies to develop
therapeutic drugs that will target cancer cells to undergo
autophagic cell death independent of apoptosis.
References
|
1
|
Parodi AJ: Protein glucosylation and its
role in protein folding. Annu Rev Biochem. 69:69–93. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lai E, Teodoro T and Volchuk A:
Endoplasmic reticulum stress: signaling the unfolded protein
response. Physiol (Bethesda). 22:193–201. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Boelens J, Lust S, Offner F, Bracke ME and
Vanhoecke BW: The endoplasmic reticulum: a target for new
anticancer drugs. In Vivo. 21:215–226. 2007.PubMed/NCBI
|
|
4
|
McGuckin MA, Eri RD, Das I, Lourie R and
Florin TH: ER stress and the unfolded protein response in
intestinal inflammation. Am J Physiol Gastrointest Liver Physiol.
298:G820–G832. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Fujita E, Kouroku Y, Isoai A, Kumagai H,
Misutani A, Matsuda C, Hayashi YK and Momoi T: Two endoplasmic
reticulum-associated degradation (ERAD) systems for the novel
variant of the mutant dysferlin: ubiquitin/proteasome ERAD(I) and
autophagy/lysosome ERAD(II). Hum Mol Genet. 16:618–629. 2007.
View Article : Google Scholar
|
|
6
|
Ohsumi Y: Molecular dissection of
autophagy: two ubiquitin-like systems. Nat Rev Mol Cell Biol.
2:211–216. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Komatsu M, Waguri S, Ueno T, Iwata J,
Murata S, Tanida I, Ezaki J, Mizushima N, Ohsumi Y and Uchiyama Y:
Impairment of starvation-induced and constitutive autophagy in
Atg7-deficient mice. J Cell Biol. 169:425–434. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chen Y, McMillan-Ward E, Kong J, Israels
SJ and Gibson SB: Oxidative stress induces autophagic cell death
independent of apoptosis in transformed and cancer cell. Cell Death
Differ. 15:171–182. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kopito RR and Ron D: Conformational
disease. Nat Cell Biol. 2:E207–E209. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Miyazaki T and Nishijima M: Studies on
fungal polysaccharides. XXVII. Structural examination of a
water-soluble, antitumor polysaccharide of Ganoderma
lucidum. Chem Pharm Bull (Tokyo). 29:3611–3616. 1981.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang SY, Hsu ML, Hsu HC, Tzeng CH, Lee SS,
Shiao MS and Ho CK: The anti-tumor effect of Ganoderma
lucidum is mediated by cytokines released from activated
macrophages and T lymphocytes. Int J Cancer. 70:699–705. 1997.
|
|
12
|
Nonaka Y, Shibata H, Nakai M, Kurihara H,
Ishibashi H, Kiso Y, Tanaka T, Yamaguchi H and Abe S: Anti-tumor
activities of the antlered form of Ganoderma lucidum in
allogeneic and syngeneic tumor-bearing mice. Biosci Biotechnol
Biochem. 70:2028–2034. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kino K, Yamashita A, Yamaoka K, Watanabe
J, Tanaka S, Ko K, Shimizu K and Tsunoo H: Isolation and
characterization of a new immunomodulatory protein, ling zhi-8
(LZ-8), from Ganoderma lucidum. J Biol Chem. 264:472–478.
1989.PubMed/NCBI
|
|
14
|
Tanaka S, Ko K, Kino K, Tsuchiya K,
Yamashita A, Murasugi A, Sakuma S and Tsunoo H: Complete amino acid
sequence of an immunomodulatory protein, Ling Zhi-8 (Lz-8). An
immunomodulator from a fungus, Ganoderma lucidum, having
similarity to immunoglobulin variable regions. J Biol Chem.
264:16372–16377. 1989.PubMed/NCBI
|
|
15
|
Huang L, Sun F, Liang C, He YX, Bao R, Liu
L and Zhou CZ: Crystal structure of LZ-8 from the medicinal fungus
Ganoderma lucidum. Proteins. 75:524–527. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liao CH, Hsiao YM, Sheu GT, Chang JT, Wang
PH, Wu MF, Wu MF, Shieh GJ, Hsu CP and Ko JL: Nuclear translocation
of telomerase reverse transcriptase and calcium signaling in
repression of telomerase activity in human lung cancer cells by
fungal immunomodulatory protein from Ganoderma tsugae.
Biochem Pharmacol. 74:1541–1554. 2007. View Article : Google Scholar
|
|
17
|
Momoi T: Caspases involved in ER
stress-mediated cell death. J Chem Neuroanat. 28:101–105. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Harding H, Zhang Y and Ron D: Translation
and protein folding are coupled by an endoplasmic reticulum
resident kinase. Nature. 397:271–274. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Rzymski T and Harris AL: The unfolded
protein response and integrated stress response to anoxia. Clin
Cancer Res. 13:2537–2540. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Leena H, Tatyana G, Huang CJ and Butler
PC: Many commercially available antibodies for detection of CHOP
expression as a marker of endoplasmic reticulum stress fail
specificity evaluation. Cell Biochem Biophys. 51:105–107. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Mann SS and Hammarback JA: Gene
localization and developmental expression of light chain 3: a
common subunit of microtubule-associated protein 1A (MAP1A) and
MAP1B. J Neurosci Res. 43:535–544. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kabeya Y, Mizushima N, Ueno T, Yamamoto A,
Kirisako T, Noda T, Kominami E, Ohsumi Y and Yoshimori T: LC3, a
mammalian homolog of yeast Apg8p, is localized in autophagosome
membrane after processing. EMBO J. 19:5720–5728. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Shintani T and Klionsky DJ: Autophagy in
health and disease: a double-edged sword. Science. 306:990–995.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yoneda T, Imaizumi K, Oono K, Yui D, Gomi
F, Katayama T and Tohyama M: Activation of caspase-12, an
endoplastic reticulum (ER) resident caspase, through tumor necrosis
factor receptor-associated factor 2-dependent mechanism in response
to the ER stress. J Biol Chem. 276:13935–13940. 2001.
|
|
25
|
Fischer H, Koenig U, Eckhart L and
Tschachler E: Human caspase 12 has acquired deleterious mutations.
Biochem Biophys Res Comm. 293:722–726. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Szegezdi E, Fitzgerald U and Samali A:
Caspase-12 and ER-stress-mediated apoptosis: the story so far. Ann
NY Acad Sci. 1010:186–194. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hitomi J, Katayama T, Eguchi Y, Kudo T,
Taniguchi M, Koyama Y, Manabe T, Yamagishi S, Bando Y, Imaizumi K,
Tsujimoto Y and Tohyama M: Involvement of caspase-4 in endoplasmic
reticulum stress-induced apoptosis and Aβ-induced. J Cell Biol.
165:347–356. 2004.
|
|
28
|
Martinez JA, Zhang Z, Svetlov SI, Hayes
RL, Wang KK and Larner SF: Calpain and caspase processing of
caspase-12 contribute to the ER stress-induced cell death pathway
in differentiated PC12 cells. Apoptosis. 15:1480–1493. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Salvesen GS: Caspases and apoptosis.
Essays Biochem. 38:9–19. 2002.
|
|
30
|
Kim I, Xu W and Reed JC: Cell death and
endoplasmic reticulum stress: disease relevance and therapeutic
opportunities. Nat Rev Drug Discov. 7:1013–1030. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Bursch W, Ellinger A, Gerner C, Fröhwein U
and Schulte-Hermann R: Programmed cell death (PCD). Apoptosis,
autophagic PCD, or others? Ann NY Acad Sci. 926:1–12. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kim R, Emi M and Tanabe K:
Caspase-dependent and -independent cell death pathways after DNA
damage. Oncol Rep. 14:595–599. 2005.PubMed/NCBI
|
|
33
|
Stuppia L, Gobbi P, Zamai L, Palka G,
Vitale M and Falcieri E: Morphometric and functional study of
apoptotic cell chromatin. Cell Death Differ. 3:397–405.
1996.PubMed/NCBI
|
|
34
|
Doonan F and Cotter TG: Morphological
assessment of apoptosis. Methods. 44:200–204. 2008. View Article : Google Scholar
|
|
35
|
Lin JC, Ho YS, Lee JJ, Liu CL, Yang TL and
Wu CH: Induction of apoptosis and cell-cycle arrest in human colon
cancer cells by meclizine. Food Chem Toxicol. 45:935–944. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Moodley YP, Caterina P, Scaffidi AK, Misso
NL, Papadimitriou JM, McAnulty RJ, Laurent GJ, Thompson PJ and
Knight DA: Comparison of the morphological and biochemical changes
in normal human lung fibroblasts and fibroblasts derived from lungs
of patients with idiopathic pulmonary fibrosis during FasL-induced
apoptosis. J Pathol. 202:486–495. 2004. View Article : Google Scholar
|
|
37
|
Nicotera P, Leist M, Single B and
Volbracht C: Execution of apoptosis: converging or diverging
pathways? Biol Chem. 380:1035–1040. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Nicotera P, Leist M, Fava E, Berliocchi L
and Volbracht C: Energy requirement for caspase activation and
neuronal cell death. Brain Pathol. 10:276–282. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Cahir-McFarland E and Kieff E: NF-kappaB
inhibition in EBV-transformed lymphoblastoid cell lines. Recent
Results Cancer Res. 159:44–48. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Graczyk PP: Caspase inhibitors as
anti-inflammatory and antiapoptotic agents. Prog Med Chem. 39:1–72.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Masuoka HC, Guicciardi ME and Gores GJ:
Caspase inhibitors for the treatment of hepatitis C. Clin Liver
Dis. 13:467–475. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Tung CW and Ho SY: Computational
identification of ubiquitylation sites from protein sequences. BMC
Bioinformatics. 9:3102008. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Xue Y, Chen H, Jin C, Sun Z and Yao X:
NBA-Palm: prediction of palmitoylation site implemented in Naive
Bayes algorithm. BMC Bioinformatics. 7:4582006. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Kouroku Y, Fujita E, Tanida I, Ueno T,
Isoai A, Kumagai H, Ogawa S, Kaufman RJ, Kominami E and Momoi T: ER
stress (PERK/eIF2a phosphorylation) mediates the
polyglutamine-induced LC3 conversion, an essential step for
autophagy formation. Cell Death Differ. 14:230–239. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Ravikumar B, Duden R and Rubinsztein DC:
Aggregate-prone proteins with polyglutamine and polyalanine
expansions are degraded by autophagy. Hum Mol Genet. 11:1107–1117.
2002. View Article : Google Scholar
|
|
46
|
Ravikumar B, Vacher C, Berger Z, Davies
JE, Luo S, Oroz LG, Scaravilli F, Easton DF, Duden R, O'Kane CJ and
Rubinsztein DC: Inhibition of mTOR induces autophagy and reduces
toxicity of polyglutamine expansions in fly and mouse models of
Huntington disease. Nat Genet. 36:585–595. 2004. View Article : Google Scholar
|