Introduction
The incidence of cervical cancer has decreased in
advanced countries due to more frequent health checkups and the
development of vaccines. In the United States in 2010, there were
12,200 cases of cervical cancer and 4,210 deaths related to this
disease, reflecting a decreasing tendency (1). However, cases of cervical cancer
worldwide have increased from 378,000 in 1980 to 454,000 in 2010
(2). Death due to cervical cancer
has decreased, but there were still about 200,000 deaths worldwide
in 2010, including 46,000 females aged 15–49 years in developing
countries (2).
Cervical cancer occurs due to infection with the
human papillomavirus (HPV) (3).
More than 100 genotypes of HPV have been detected and about 40 have
been found to infect the genital tract. HPV is classified into
high-risk types causing cervical cancer, including HPV16 and HPV18,
and low-risk types causing conditions other than cancer, such as a
polyp. HPV16, the most frequent genotype, is detected in
approximately half of patients with cervical cancer (4). After infecting cells, HPV produces
oncoproteins E6 and E7, which inhibit controlling the cell cycle
and apoptosis, and therefore have important roles in oncogenesis.
E6 is associated with p53 and induces p53 degradation by E3
ubiquitin ligase via the function of E6-associated protein (E6AP).
E7 inactivates the retinoblastoma tumor suppressor gene product,
pRb and its family members (5).
Epigenetic DNA methylation of tumor suppressor genes
in the promoter region is generally important in carcinogenesis
(6–8). In cervical cancer, carcinogenesis is
related to aberrant methylation of CpG islands of p16, FHIT,
retinoic acid receptor β, E-cadherin, death-associated protein
kinase, HIC-1, APC, and Ras association domain family 1A genes
(9–12). Methylation of CpG islands in the
WRN promoter region is related to carcinogenesis in various
cancers (13). The chromosomal
WRN locus on the short arm of chromosome 8, is composed of
35 exons, and has a length >250 kb (14). WRN encodes the WRN
protein, which is a member of the RecQ helicase family and is also
an exonuclease. Loss of WRN causes abnormalities in DNA
repair, replication, and telomere maintenance.
Werner syndrome (WS) is an autosomal recessive
genetic disease that is caused by mutation of the WRN gene.
WS symptoms include aging at an early stage and various secondary
symptoms associated with aging, including bilateral cataract, skin
change, short stature and graying hair; in addition, diabetes
mellitus, osteoporosis, atherosclerosis and cancer also develop
frequently (15). Malignant
complications include sarcomas of mesenchymal origins, including
soft-tissue sarcoma and osteosarcoma, suggesting that the mechanism
of carcinogenesis in WS differs from that in carcinogenesis in
other cancers (16). The mean age
at death of WS patients is 46–54 years and one of major causes is
the high prevalence of malignancy (17,18).
Several studies have shown a relationship between
WRN expression and malignancy and have indicated that
epigenetic inactivation of WRN is of importance in
carcinogenesis. In many tumors, loss of heterozygosity is detected
in chromosome 8p, in which WRN is located, but WRN
somatic mutation has not been found, suggesting that epigenetic
control has a significant effect (19,20).
Epigenetic DNA methylation in the promoter region of WRN and
a methylation-induced decrease in WRN expression have been
found in colorectal, lung, gastric, prostate and breast cancer. The
methylation-induced decrease in WRN expression increases
chromosomal instability (13).
Reduced WRN expression is also related to
sensitivity to camptothecin (CPT-11), a topoisomerase I (Top-I)
inhibitor (13,21). CPT-11 is an alkaloid found in plants
such as Camptotheca acuminata. A single-strand break (SSB)
occurs after Top I binds to DNA and generates a Top I-DNA cleavable
complex. CPT-11 stabilizes this complex and inhibits reconnection
of the SSB, resulting in inhibition of DNA synthesis (22). CPT-11 also inhibits replication fork
progression, resulting in DNA double-strand breaks (DSBs) and
apoptosis (23). Inactivation of
WRN in cancer cells increases the effect of CPT-11 (13,21),
and overall survival of patients with colorectal cancer treated
with irinotecan, a camptothecin analogue, is dependent on the
methylation status of CpG islands in the WRN promoter
(13).
Cisplatin is a key drug in chemotherapy for cervical
cancer (24). CPT-11 is a similarly
important drug and has a high response rate of 24% (25). Evaluation of WRN expression
as a marker of sensitivity to CPT-11 may be clinically useful in
treatment of cervical cancer. Thus, in this study, the associations
among cervical cancer, WRN expression, and cancer cell
sensitivity to CPT-11 were investigated.
Materials and methods
Subjects and cytologic specimens
Samples were obtained from 21 cervical cancer smears
collected using a ThinPrep system (Cytyc, Boxborough, MA, USA) and
kept in preservation fluid (PreservCyt Solution, Cytyc) (26). Informed consent was obtained before
collection. Pathological diagnosis was performed by cervical
histology, and the cytological and histological results were
consistent for all smears. Of the 21 cervical cancer smears, 10
were squamous carcinoma and 11 were adenocarcinoma. The
histological type and stage were determined according to the
General Rules for Clinical Cervical Cancer in Japan published by
the Japan Society of Obstetrics and Gynecology.
Cultured cell lines
The human cervical squamous cell carcinoma-derived
cell lines, SKG-I, SKG-II, SKG-IIIa and SKG-IIIb, and the human
cervical adenocarcinoma-derived cell lines, HeLa and TCO-I, were
used in the study. HeLa cells were incubated in DMEM (Sigma, St.
Louis, MO, USA) with 10% fetal bovine serum (FBS) (Sanko Junyaku
Co., Tokyo, Japan) and TCO-I cells were incubated in MEM medium
(Sigma) with 10% FBS. All other cell lines were incubated in F12
medium (Sigma) with 10% FBS. Cells were incubated in 10-cm dishes
at 37°C in a 5% CO2 atmosphere.
DNA extraction and methylation-specific
PCR (MSP) assay of the WRN gene
DNA was extracted from 21 cervical smears and 6
cervical carcinoma-derived cell lines using a Get Pure DNA kit
(Dojin Glocal, Kumamoto, Japan). DNA (1 μg) extracted from cervical
smears was diluted with 50 μl of distilled water and incubated in
5.5 μl of 3 N NaOH at 37°C for 15 min. To this solution, 30 μl of
10 mM hydroquinone (Sigma) and 520 μl of 3 M sodium bisulfite
(prepared at pH 5.5 with 10 N NaOH, Sigma) were added with mixing.
Mineral oil was laid over the solution to prevent evaporation, and
the solution was incubated overnight at 50°C. The lower layer of
the reaction solution was mixed with 1 ml of Clean-up Resin
(Promega, Madison, WI, USA) and then injected into a column. After
rinsing with 2 ml of 80% isopropanol, the mixture was centrifuged
at 15,000 rpm for 3 min to remove isopropanol. Hot (70°C) distilled
water (50 μl) was added and the mixture was centrifuged at 15,000
rpm for 2 min to elute DNA. The DNA was then incubated with 5.5 μl
of 2 N NaOH at 37°C for 20 min. Next, 66 μl of 5 N ammonium acetate
and 243 μl of 95% ethanol were added and the mixture was incubated
at −80°C for one hour and centrifuged at 15,000 rpm for 30 min to
precipitate DNA. Supernatant exceeding 50 μl was removed, 1 ml of
60% ethanol was added, and the mixture was centrifuged at 15,000
rpm for 30 min and rinsed. The precipitated DNA was dried in air
and dissolved in 20 μl of distilled water. DNA solution (2 μl) was
used as the MSP template. In the PCR assay, AmpliTaq Gold and 10X
PCR buffer/MgCl2 with dNTP (Applied Biosystems, Foster
City, CA, USA) were used and the results were analyzed with a
GeneAmp PCR System 9700 (Applied Biosystems).
The primer sequences were 5′-CGGGTAGGGGTATCG
TTCGC-3′ (sense) and 5′-AACGAAATCCACCGCCCGCC-3′ (antisense), 159
bp. Primer sequences for the unmethylated reaction were
5′-GTAGTTGGGTAGGGGTATTGTTTGT-3′ (sense) and
5′-AAACAACCTCCACCACCCACCCC-3′ (antisense), 165 bp. PCR was
performed for 35 cycles (95, 65 and 72°C for 30 sec, respectively).
DNA extracted from the cultured cell lines was prepared similarly
for use in MSP analysis of the WRN gene.
RNA extraction and RT-PCR assay of WRN
expression
Total RNA from 6 cervical cancer-derived cell lines
was extracted using an RNeasy mini-kit (Qiagen, Valencia, CA, USA).
cDNA was synthesized from 1 μg of total RNA using SuperScriptII
Reverse Transcriptase (Invitrogen, Carlsbad, CA, USA). WRN
expression was analyzed in an RT-PCR assay using 1 μl of
first-strand cDNA as template. AmpliTaq Gold and 10X PCR
buffer/MgCl2 with dNTP were used in the PCR assay, with
analysis using a GeneAmp PCR System 9700 (Applied Biosystems). The
primer sequences were 5′-GCATGTGTTCGGAAGAGTGTTT-3′ (sense) and
5′-TGACATGGAAGAAACGTGGAA-3′ (antisense), 258 bp. PCR was performed
for 30 cycles (94, 57 and 72°C for 30 sec, respectively).
Demethylation treatment
Cervical carcinoma-derived SKGII and TCO-I cells
with aberrant methylation of WRN were plated on 10-cm dishes
at 106 cell/dish and incubated for 72 h.
5-Aza-2-deoxycytidine (5-aza) (Sigma), a demethylating agent, was
then added at a final concentration of 1 μM in culture medium.
After 48 h of incubation, 5-aza was added again and DNA and RNA
were extracted 48 h after the second addition of 5-aza.
In vitro test of sensitivity to
anticancer agents
The sensitivity of cervical carcinoma-derived cell
lines to anti-cancer agents was determined using the collagen gel
droplet-embedded culture drug sensitivity test (CD-DST) (27). Cells were pretreated with cell
dispersion enzyme EZ (Nitta Gelatin Inc., Tokyo, Japan) for 2 h,
followed by centrifugation to collect the cells. In a flask
containing collagen gel, the cells were pre-incubated for 24 h and
surviving cells that adhered to collagen gel were collected.
Cellmatrix Type CD solution was added to the collected cells, and
the suspension of cells and collagen gel was dropped onto a 6-well
plate to prepare 3 drops of 30 μl each. The suspension was left to
stand in an incubator at 37°C in a 5% CO2 atmosphere for
1 h for gelling and then overlaid with 4 ml/well of medium.
Cisplatin (CDDP), doxorubicin (ADM), or CPT-11 was then added to
the suspension at final concentrations of 0.2, 0.02 and 0.03 μg/ml,
respectively. After 24 h, the anticancer agents were removed by
rinsing and the cells were incubated without serum at 37°C in 5%
CO2 for 7 days. The cells were dyed with neutral red,
fixed with formalin, and dried. Images were collected by scanning
using an image analyzer and the ratio of the volume of living
cancer cells in the treated group (T) to that in the control group
(C) (T/C ratio) was determined. In general, cells are considered to
be highly sensitive to the agent when the T/C ratio is ≤50%
(28).
Transfection of small interfering RNA
(siRNA)
SKG-IIIb cells were plated on 60-mm dishes at
4×105 cell/dish and transfected 48 h later with siRNA
using siFector (B-Bridge International Inc., Cupertino, CA, USA).
In this procedure, 4.5 μl of siRNA stock solution (100 μM) and
295.5 μl of serum-free MEM were mixed in a test tube. In another
tube, 13.5 μl of siFector and 286.5 μl of serum-free MEM were
mixed. The solutions from the two tubes were mixed and incubated at
room temperature for 30 min. Each dish containing SKG-IIIb cells
was rinsed twice with 2 ml of serum-free MEM and 2.4 ml of
serum-free MEM was then added. The incubated siRNA mixture was
added to the dish at 0.6 ml/dish and incubated at 37°C in 5%
CO2 for 6 h. After incubation, 3 ml of MEM containing
20% serum was added to the dish. A negative control siRNA was used
as designed by B-Bridge International Inc. The siRNA sequence
corresponding to the WRN gene was
5′-GUUCUUGUCACGUCCUCUGdTdT-3′. The expression levels of mRNA and
protein were determined 48 h after siRNA addition. Anticancer
agents were added 48 h after siRNA addition and the sensitivity of
the cells to each agent was analyzed using the CD-DST.
Immunoblotting
siRNA-transfected SKG-IIIb cells were rinsed with
PBS, trypsinized, and centrifuged at 15,000 rpm for 5 min at 4°C.
Protein was extracted using a Mammalian Cell Extraction kit
(BioVision Research Products, Mountain View, CA, USA). The sample
(200 μg of protein) was mixed with sample buffer (Bio-Rad
Laboratories, Hercules, CA, USA) containing the equivalent volume
of 5% β-mercaptoethanol (Bio-Rad Laboratories) and the mixture was
boiled for 5 min. After boiling, the mixture was electrophoresed on
a 10% polyacrylamide gel and the proteins were transferred to
nitrocellulose membranes (Bio-Rad Laboratories). The membranes were
soaked in PBS containing 1% BSA and 0.1% Tween-20 and incubated at
room temperature for 1 h for blocking. They were then reacted with
anti-β-actin antibody (AC-74, Sigma-Aldrich, St. Louis, MO, USA;
5000-fold diluted) and anti-WRN antibody (4H12, Abcam,
Cambridge, UK; 500-fold diluted) at 4°C overnight, followed by
rinsing three times with PBS containing 0.1% Tween (PBS-T) for 10
min each. The anti-β-actin and anti-WRN antibodies were reacted
with anti-mouse IgG antibody (PK-6102, Vector Laboratories,
Burlingame, CA, USA) and anti-goat IgG antibody (BA-5000, Vector
Laboratories; 250-fold diluted), respectively, at room temperature
for 1 h. The membranes were rinsed with PBS-T three times and
reacted with ABC complex (PK-6102, Vector Laboratories; pre-reacted
at 4°C for 30 min) at room temperature for 1 h, then rinsed with
PBS-T twice and PBS once, and visualized with DAB (Sigma).
Cell cycle analysis using flow
cytometry
The cell cycle was evaluated 96 h after siRNA
addition. Cells were trypsinized and rinsed twice with PBS.
Supernatant was separated from the cell pellets by centrifugation
at 1,000 rpm for 5 min, and 450 μl of PBS was added to the pellets
and the mixture was pipetted well. As the mixture was vortexed, 1
ml of cool 100% ethanol was added. The mixture was then incubated
at room temperature for 30 min for cell fixation. The cells were
rinsed twice with PBS and 500 μl of RNase was added to the pellets
after supernatant removal. The cells were then incubated at room
temperature for 20 min. Subsequently, 500 μl of propidium iodide
solution was added, the mixture was poured into a cell strainer,
and the cell cycle was determined by flow cytometry using an Epics
XL MCL (Beckman Coulter, Inc., Fullerton, CA, USA).
Results
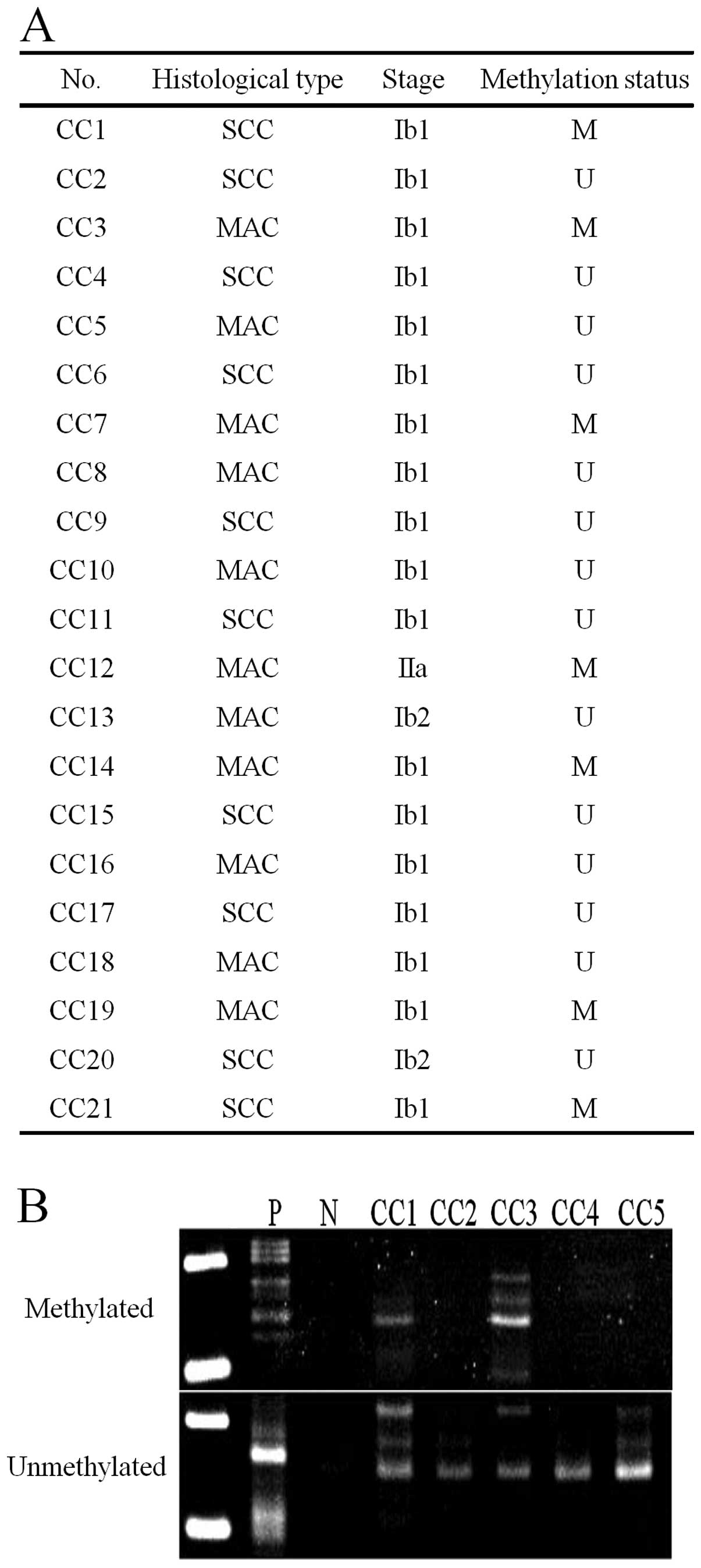
Aberrant methylation of the WRN gene in
cervical cancer was examined using specimens collected for cytology
(Fig. 1). Aberrant methylation was
detected in 7 (33.3%) of 21 patients, including in 2 (20%) of 10
cases of squamous cell carcinoma and 5 (45.5%) of 11 cases of
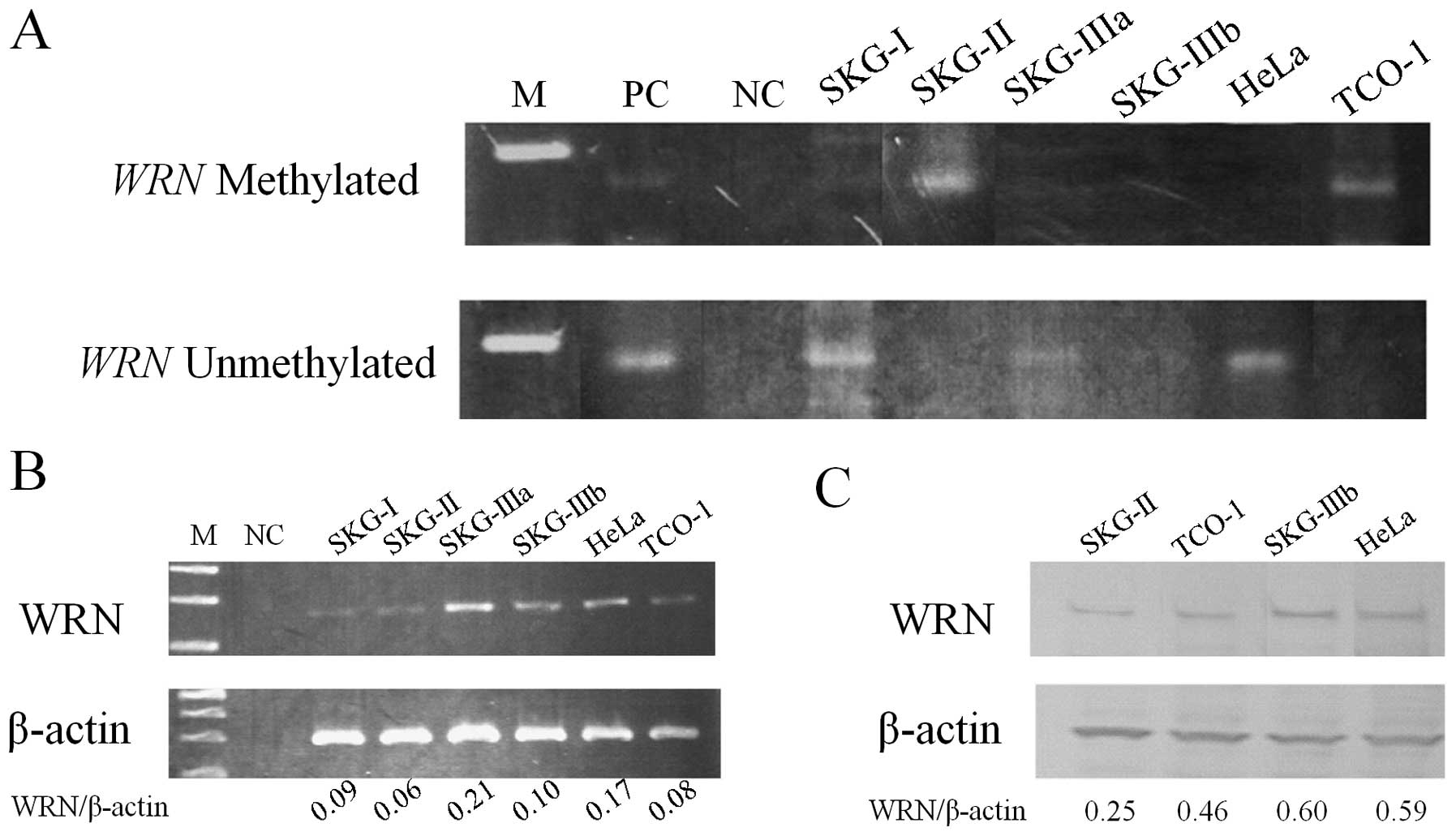
adenocarcinoma. Among 6 cervical cancer-derived cell lines,
aberrant methylation was detected in 2 cell lines, SKG-II and TCO-I
(Fig. 2A), and mRNA and protein
levels for WRN were lower in these cells (Fig. 2B and C).
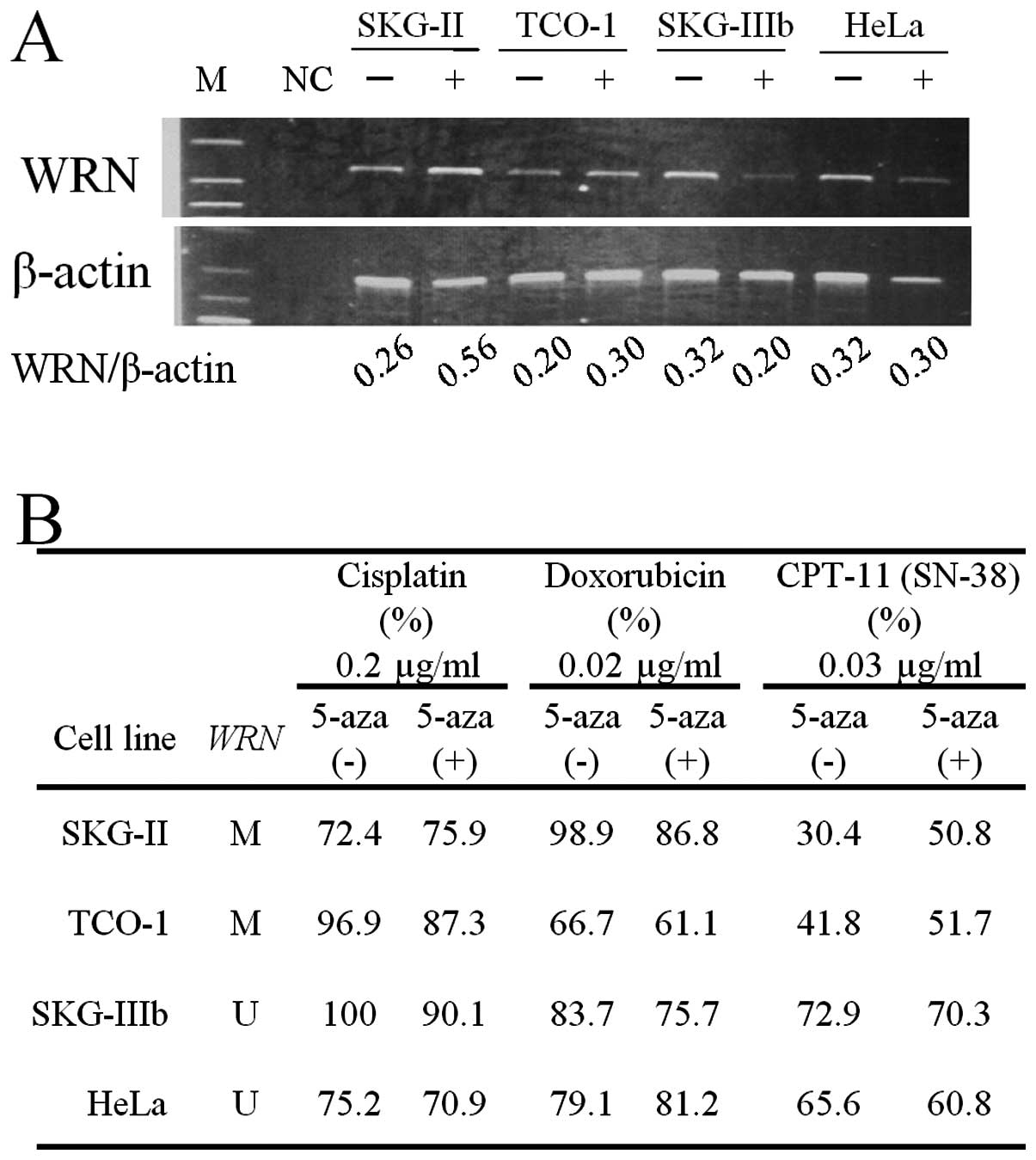
Changes in WRN mRNA levels in cervical
cancer-derived cell lines were analyzed before and after treatment
with 5-aza, a demethylating agent (Fig.
3A). After administration of 5-aza, WRN mRNA increased
in SKG-II and TCO-I cells, in which aberrant methylation of
WRN was found. Sensitivity to anticancer drugs before and
after treatment with 5-aza was analyzed by CD-DST, based on the T/C
ratio (Fig. 3B). Sensitivity to
CDDP and ADM did not change in 4 cell lines after administration of
5-aza. For CPT-11, the T/C ratio increased to >50% in SKG-II and
TCO-I cells after administration of 5-aza, showing decreased
sensitivity to CPT-11.
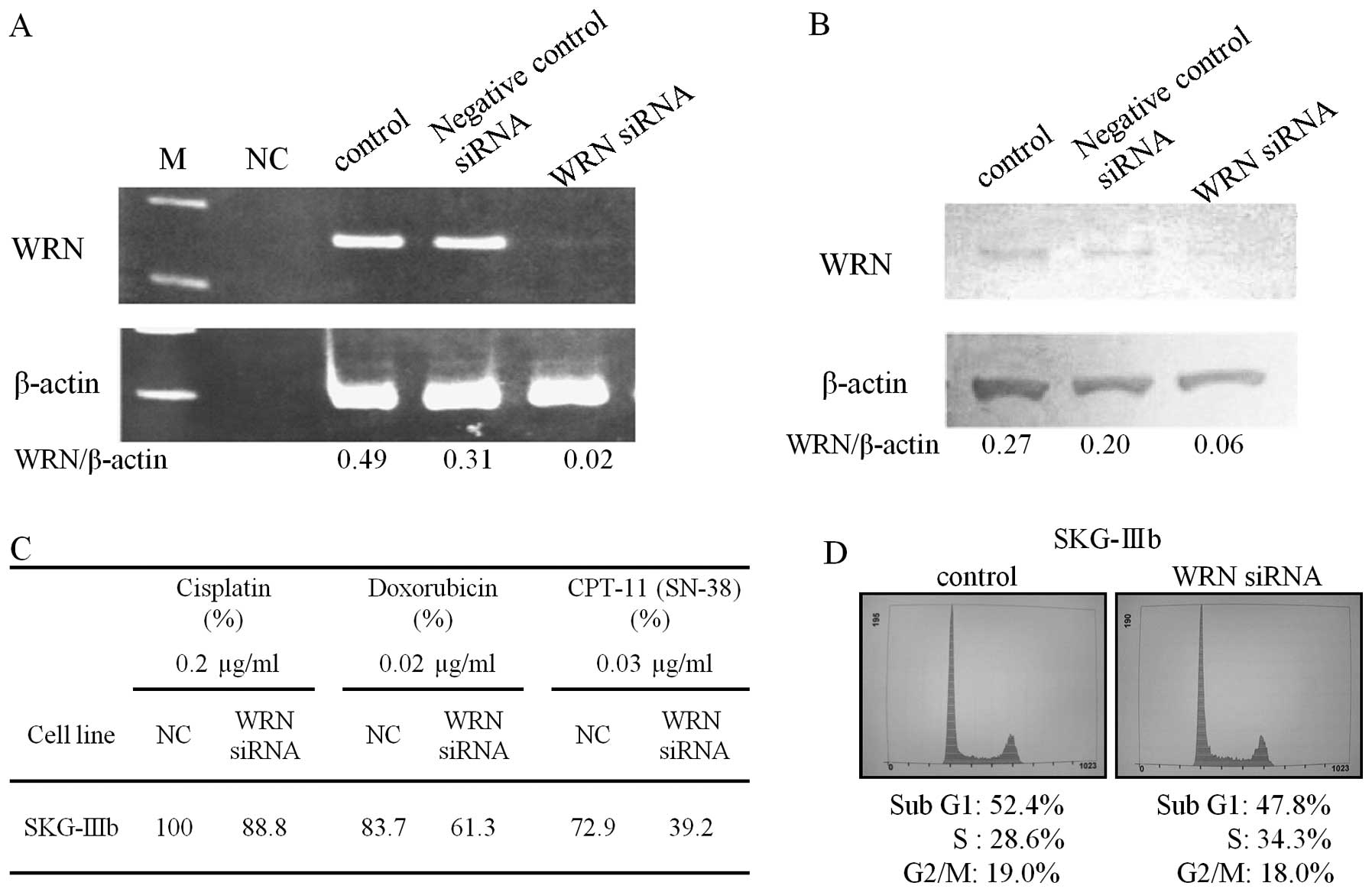
Introduction of siRNA for WRN in SKG-IIIb
cells decreased the levels of WRN mRNA and protein (Fig. 4A and B). The sensitivity of the
cells to CPT-11 was increased by siRNA treatment based on the
marked decrease in the T/C ratio in the CD-DST (Fig. 4C). Flow cytometry indicated that the
percentage of S-phase cells increased from 28.6 to 34.3% after
siRNA for WRN was introduced into SKG-IIIb cells (Fig. 4D).
Discussion
This study provided the first evidence of a
relationship of WRN promoter methylation with cervical
cancer, with aberrant methylation of WRN detected in 33.3%
of specimens of cervical cancer and in two cervical cancer-derived
cell lines. Decreased WRN mRNA and protein levels were also
found in both cell lines. These results suggested that aberrant
methylation of WRN plays an important role in cervical
cancer. The sensitivity to CPT-11 of cervical cancer cells with
aberrant methylation of WRN was decreased by treatment with
a demethylating agent. This treatment also increased the level of
WRN mRNA, consistent with the general effect of
demethylating agents on expression of many genes.
Selective downregulation of WRN expression
using siRNA increased the sensitivity of cervical cancer cells to
CPT-11, but not to other anticancer agents. Several previous
studies have shown that WRN inactivation increases the
anticancer effect of CPT-11 (13,21).
CPT-11 acts on the covalent complex of topoisomerase I (Top-I) and
DNA, and inhibits DNA replication and causes strand breaks
(29–31). CPT-11 acts in S-phase and causes
activation of S-phase checkpoint function via the ATR (ataxia and
Rad-related protein)-CHK1 (checkpoint kinase-1) pathway (32–34).
WRN is involved in the ATR-CHK1 pathway by recognizing the
Top-I-DNA complex and detecting replication-derived DNA structures
or unresolved positive supercoils (35,36).
Thus, if ATR, CHK1 and WRN have reduced activity, cells are
hypersensitive to CPT-11, and administration of CPT-11 to
inactivated WS cells increases S-phase DSBs and unresolved
recombination structures. A similar effect does not occur with
etoposide, a topoisomerase II inhibitor (37). The S-phase checkpoint is activated
upon DNA damage and is regulated by ataxia telangiectasia mutated
(ATM) and ATM and Rad3-related protein (ATR) kinases. WRN is
related to both kinases (38,39).
Cells with inactivated WRN proceed to S-phase earlier than
wild-type cells; however, the slow progress in late S-phase for
cells with inactivated WRN causes the final S-phase lengths
to be equal (40). In this study,
siRNA for WRN produced a small, but insignificant, increase
in the number of S-phase cells.
In chemotherapy for cervical cancer, the response
rate to cisplatin is 20–30% (24)
and that to monotherapy with CPT-11 has been found to be 24%
(25). A combination of cisplatin
and CPT-11 has a good response rate of 59% and is effective therapy
(41). Taxanes also play an
important role in chemotherapy for cervical cancer, with response
rates of 17% for taxane monotherapy (42) and 46% for cisplatin and paclitaxel
(TP) combination therapy for recurrent or advanced cervical cancer,
making this regimen the current standard of care (43). We previously showed a relationship
between aberrant methylation of CHFR and sensitivity to
taxanes, and suggested that aberrant methylation of CHFR
could serve as a molecular marker for the sensitivity of cervical
cancer to anticancer drugs (44).
Thus, examination of aberrant methylation of CHFR and
WRN in specimens collected for cytology may be useful for
prediction of treatment outcome before administration of anticancer
drugs.
The standard treatment for cervical cancer is
concurrent chemoradiotherapy from stages IB bulky to IIB (45). Neoadjuvant chemotherapy (NAC) is a
promising approach for reduction of tumor size to increase the
number of patients indicated for surgery and reduce distant
metastasis through an effect on micrometastasis (46). It has been suggested that NAC could
be used in stages IB2 to IIIB (47), but the efficacy of NAC remains
inconclusive (48). A disadvantage
of NAC is that the tumor may grow prior to the main therapy in
cases that are non-responsive to NAC. However, this concern may be
avoided by choice of the most effective chemotherapy based on
evaluation of methylation of CHFR and WRN in
specimens collected before initiation of NAC. This approach may
represent a new therapeutic strategy for cervical cancer.
In conclusion, our data suggest that aberrant
methylation of WRN plays an important role in carcinogenesis
and sensitivity to CPT-11 of cervical cancer. This is the first
report to show a relationship between the methylation of the
WRN gene and sensitivity to CPT-11 in gynecologic
cancer.
Acknowledgements
The authors gratefully acknowledge grant support
from the Japan Society for the Promotion of Science (JSPS) through
a Grant-in-Aid for Scientific Research (KAKENHI), a Grant-in-Aid
for Scientific Research (B) (22390313), a Grant-in-Aid for
Scientific Research (C) (22591866), and a Grant-in-Aid for Young
Scientists (B) (21791573); the Ichiro Kanehara Foundation;
Kobayashi Foundation for Cancer Research; and the Keio University
Medical Science Fund through a Research Grant for Life Sciences and
Medicine.
References
|
1
|
Jemal A, Siegel R, Xu J and Ward E: Cancer
Statistics, 2010. CA Cancer J Clin. 60:277–300. 2010. View Article : Google Scholar
|
|
2
|
Forouzanfar MH, Foreman KJ, Delossantos
AM, et al: Breast and cervical cancer in 187 countries between 1980
and 2010: a systematic analysis. Lancet. 378:1461–1484. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
zur Hausen H: Papillomaviruses in
anogenital cancer as a model to understand the role of viruses in
human cancers. Cancer Res. 49:4677–4681. 1989.PubMed/NCBI
|
|
4
|
Munoz N, Bosch FX, de Sanjosé S, et al:
Epidemiologic classification of human papillomavirus types
associated with cervical cancer. N Engl J Med. 348:518–527. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
zur Hausen H: Papillomaviruses and cancer:
from basic studies to clinical application. Nat Rev Cancer.
2:342–350. 2002.PubMed/NCBI
|
|
6
|
Jones PA and Laird PW: Cancer epigenetics
comes of age. Nat Genet. 21:163–167. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Baylin SB and Herman JG: DNA
hypermethylation in tumorigenesis: epigenetics joins genetics.
Trends Genet. 16:168–174. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Esteller M: Epigenetic gene silencing in
cancer: the DNA hypermethylome. Hum Mol Genet. 16:50–59. 2007.
View Article : Google Scholar
|
|
9
|
Virmani AK, Muller C, Rathi A,
Zoechbauer-Mueller S, Mathis M and Gazdar AF: Aberrant methylation
during cervical carcinogenesis. Clin Cancer Res. 7:584–589.
2001.PubMed/NCBI
|
|
10
|
Wong Y, Chung TK, Cheung T, et al:
Methylation of p16INK4A in primary gynecologic
malignancy. Cancer Lett. 136:231–235. 1999.PubMed/NCBI
|
|
11
|
Dong SM, Kim H-S, Rha S-H and Sidransky D:
Promoter hypermethylation of multiple genes in carcinoma of the
uterine cervix. Clin Cancer Res. 7:1982–1986. 2001.PubMed/NCBI
|
|
12
|
Kuzmin I, Liu L, Dammann R, et al:
Inactivation of RAS association domain family 1A gene in cervical
carcinomas and the role of human papillomavirus infection. Cancer
Res. 63:1888–1893. 2003.PubMed/NCBI
|
|
13
|
Agrelo R, Cheng W-H, Setien F, et al:
Epigenetic inactivation of the premature aging Werner syndrome gene
in human cancer. Proc Natl Acad Sci USA. 103:8822–8827. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yu CE, Oshima J, Wijsman EM, et al:
Mutations in the consensus helicase domains of the Werner syndrome
gene. Werner’s Syndrome Collaborative Group. Am J Hum Genet.
60:330–341. 1997.PubMed/NCBI
|
|
15
|
Muftuoglu M, Oshima J, Kobbe C, Cheng W-H,
Leistritz DF and Bohr VA: The clinical characteristics of Werner
syndrome: molecular and biochemical diagnosis. Hum Genet.
124:369–377. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Goto M, Miller RW, Ishikawa Y and Sugano
H: Excess of rare cancers in Werner syndrome (adult progeria).
Cancer Epidemiol Biomarkers Prev. 5:239–246. 1996.PubMed/NCBI
|
|
17
|
Epstein CJ, Martin GM, Schultz AL and
Motulsky AG: Werner’s syndrome a review of its symptomatology,
natural history, pathologic features, genetics and relationship to
the natural aging process. Medicine. 45:177–221. 1966.
|
|
18
|
Huang S, Lee L, Hanson NB, et al: The
spectrum of WRN mutations in Werner syndrome patients. Hum Mutat.
27:558–567. 2006. View Article : Google Scholar
|
|
19
|
Shaheen AC, Malcolm CC, Neil RJC, et al:
Two novel regions of interstitial deletion on chromosome 8p in
colorectal cancer. Oncogene. 18:657–665. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Armes JE, Hammet F, de Silva M, et al:
Candidate tumor-suppressor genes on chromosome arm 8p in
early-onset and high-grade breast cancers. Oncogene. 23:5697–5702.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Futami K, Takagi M, Shimamoto A, Sugimoto
M and Furuichi Y: Increased chemotherapeutic activity of
camptothecin in cancer cells by siRNA-induced silencing of WRN
helicase. Biol Pharm Bull. 30:1958–1961. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hsiang YH, Hertzberg R, Hecht S and Liu
LF: Camptothecin induces protein-linked DNA breaks via mammalian
DNA topoisomerase I. J Biol Chem. 260:14873–14878. 1985.PubMed/NCBI
|
|
23
|
Hsiang YH, Lihou MG and Liu LF: Arrest of
replication forks by drug-stabilized topoisomerase I-DNA cleavable
complexes as a mechanism of cell killing by camptothecin. Cancer
Res. 49:5077–5082. 1989.
|
|
24
|
Thigpen T: The role of chemotherapy in the
management of carcinoma of the cervix. Cancer J. 9:425–432. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Takeuchi S, Dobashi K, Fujimoto S, et al:
A late phase II study of CPT-11 on uterine cervical cancer and
ovarian cancer. Research Groups of CPT-11 in Gynecologic Cancers.
Gan To Kagaku Ryoho. 18:1681–1689. 1991.(In Japanese).
|
|
26
|
Susumu N, Aoki D, Noda T, et al:
Diagnostic clinical application of two-color fluorescence in situ
hybridization that detects chromosome 1 and 17 alterations to
direct touch smear and liquid-based thin-layer cytologic
preparations of endometrial cancers. Int J Gynecol Cancer.
15:70–80. 2005. View Article : Google Scholar
|
|
27
|
Kawaguchi M, Banno K, Susumu N, et al:
Successful analysis of anticancer drug sensitivity by CD-DST using
pleural fluid and ascites from patients with advanced ovarian
cancer: case reports. Anticancer Res. 25:3547–3551. 2005.PubMed/NCBI
|
|
28
|
Kawamura M, Gika M, Abiko T, et al:
Clinical evaluation of chemosensitivity testing for patients with
unresectable non-small cell lung cancer (NSCLC) using collagen gel
droplet embedded culture drug sensitivity test (CD-DST). Cancer
Chemother Pharmacol. 59:507–513. 2007. View Article : Google Scholar
|
|
29
|
Leppard JB and Champoux JJ: Human DNA
topoisomerase I: relaxation, roles, and damage control. Chromosoma.
114:75–85. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Covey JM, Jaxel C, Kohn KW and Pommier Y:
Protein-linked DNA strand breaks induced in mammalian cells by
camptothecin, an inhibitor of topoisomerase I. Cancer Res.
49:5016–5022. 1989.PubMed/NCBI
|
|
31
|
Shao RG, Cao CX, Zhang H, Kohn KW, Wold MS
and Pommier Y: Replication-mediated DNA damage by camptothecin
induces phosphorylation of RPA by DNA-dependent protein kinase and
dissociates RPA:DNA-PK complexes. EMBO J. 18:1397–1406. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wan S, Capasso H and Walworth NC: The
topoisomerase I poison camptothecin generates a Chk1-dependent DNA
damage checkpoint signal in fission yeast. Yeast. 15:821–828. 1999.
View Article : Google Scholar
|
|
33
|
Cliby WA, Lewis KA, Lilly KK and Kaufmann
SH: S phase and G2 arrests induced by topoisomerase I poisons are
dependent on ATR kinase function. J Biol Chem. 277:1599–1606. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Seiler JA, Conti C, Syed A, Aladjem MI and
Pommier Y: The intra-S-phase checkpoint affects both DNA
replication initiation and elongation: single-cell and -DNA fiber
analyses. Mol Cell Biol. 27:5806–5818. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Koster DA, Palle K, Bot ESM, Bjornsti M-A
and Dekker NH: Antitumour drugs impede DNA uncoiling by
topoisomerase I. Nature. 448:213–217. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Patro BS, Frøhlich R, Bohr VA and
Stevnsner T: WRN helicase regulates the ATR-CHK1-induced S-phase
checkpoint pathway in response to topoisomerase-I-DNA covalent
complexes. J Cell Sci. 124:3967–3979. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Christmann M, Tomicic MT, Gestrich C, Roos
WP, Bohr VA and Kaina B: WRN protects against topo I but not topo
II inhibitors by preventing DNA break formation. DNA Repair.
7:1999–2009. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Abraham RT: Cell cycle checkpoint
signaling through the ATM and ATR kinases. Genes Dev. 15:2177–2196.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Shiloh Y: ATM and related protein kinases:
safeguarding genome integrity. Nat Rev Cancer. 3:155–168. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Versini G, Comet I, Wu M, Hoopes L, Schwob
E and Pasero P: The yeast Sgs1 helicase is differentially required
for genomic and ribosomal DNA replication. EMBO J. 22:1939–1949.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Sugiyama T, Yakushiji M, Noda K, et al:
Phase II study of irinotecan and cisplatin as first-line
chemotherapy in advanced or recurrent cervical cancer. Oncology.
58:31–37. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
McGuire WP, Blessing JA, Moore D, Lentz SS
and Photopulos G: Paclitaxel has moderate activity in squamous
cervix cancer. A Gynecologic Oncology Group study. J Clin Oncol.
14:792–795. 1996.PubMed/NCBI
|
|
43
|
Rose PG, Blessing JA, Gershenson DM and
McGehee R: Paclitaxel and cisplatin as first-line therapy in
recurrent or advanced squamous cell carcinoma of the cervix: a
gynecologic oncology group study. J Clin Oncol. 17:2676–2680.
1999.
|
|
44
|
Banno K, Yanokura M, Kawaguchi M, et al:
Epigenetic inactivation of the CHFR gene in cervical cancer
contributes to sensitivity to taxanes. Int J Oncol. 31:713–720.
2007.PubMed/NCBI
|
|
45
|
Cervical cancer guideline (Version 1.
2012). NCCN Clinical Practice Guidelines in Oncology.
|
|
46
|
Treatment Guidelines for cervical cancer.
Japan Society of Gynecologic Oncology. Kanehara & Co; 2011
|
|
47
|
Benedetti-Panici P, Greggi S, Colombo A,
et al: Neoadjuvant chemotherapy and radical surgery versus
exclusive radiotherapy in locally advanced squamous cell cervical
cancer: results from the Italian multicenter randomized study. J
Clin Oncol. 20:179–188. 2002. View Article : Google Scholar
|
|
48
|
Chang TC, Lai CH, Hong JH, et al:
Randomized trial of neoadjuvant cisplatin, vincristine, bleomycin,
and radical hysterectomy versus radiation therapy for bulky stage
IB and IIA cervical cancer. J Clin Oncol. 18:1740–1747. 2000.
|