Introduction
Matriptase-2 is a member of the type II
transmembrane serine protease (TTSP) family, a family of proteases,
which has attracted significant scientific interest due to
differential expression patterns in normal and cancerous tissue and
cells (1–5). The matriptase-2 gene is located on
chromosome 22 encoding an 88,901-kDa molecular weight protein
(6). The protein structure of
matriptase-2 is characteristic of that of the TTSP group,
consisting of a mixture of domains common to this family of
proteases and has been described in a number of studies (7,8).
Matriptase-2 shares homology with matriptase-1, another TTSP whose
expression is altered in various human cancers (9). However, whereas the overexpression of
matriptase-1 leads to cancer progression (10,11),
matriptase-2 overexpression significantly reduces breast and
prostate cancer growth and the reduced expression of matriptase-2
is associated with poor patient outcome (12,13).
This difference in apparent functions of both proteases has
prompted more in depth investigation into the potential
anti-metastatic properties of matriptase-2. Currently, the
molecular mechanisms involved in the activation and regulation of
matriptase-2 remain largely unknown due to the small number of
studies currently investigating matriptase-2.
Cancer cells arise from normal cells via mutation.
These mutations allow the previously non-cancerous cells to gain
characteristics facilitating the progression to an aggressive
phenotype. A cancer cell relies on a number of vital biological
processes with which to establish itself in its environment and to
subsequently metastasise. The most significant of these include
changes in cell growth and invasive and migratory capacities.
Cancer cells can influence these processes in several ways, most
significantly by altering the expression of molecules that play key
roles in controlling these cellular traits. Matriptase-2 has
previously shown the ability to reduce the migration of PC3 and
DU145 cells and the adhesion of PC3 cells (13).
β-catenin was first described as a structural
component in adherens junction formation, a mechanism of cell-cell
adhesion and is normally maintained at low levels in the absence of
Wnt stimulation, due to its constitutive proteasomal degradation
(14). The reduction of β-catenin
degradation upon Wnt stimulation leads to the increased
participation of β-catenin in two major cellular processes: nuclear
signalling and cell-cell adhesion. β-catenin accumulation in the
cytoplasm results in its translocation into the nucleus and
interaction with DNA-binding transcription factors, the most
prominent of which are those of the T cell-specific transcription
factor (TCF)/lymphoid enhancer factor (LEF) family. Once localized
to the nucleus, the complex formed between β-catenin and its
DNA-binding transcription factors initiates the transcription of
its target genes. These target genes are highly involved in the
regulation of many proteins that are involved in maintaining the
proper functions of the cell. These target genes include c-MYC
(15), MMP-7 (16,17),
VEGF (18) and MET (19). Due to the importance of these target
genes in maintaining the correct function of the cell, the balance
between the degradation and activation of β-catenin must remain
tightly controlled. The deregulation of this pathway plays a
significant role in the progression and possibly the development of
cancer.
In the present study, the expression of matriptase-2
was knocked down in the normal prostate cell lines, PZHPV7 and
PNT2C2, using a ribozyme transgene targeting matriptase-2. The
biological functions of these cells were subsequently examined. In
order to investigate any link between matriptase-2 and β-catenin,
matriptase-2 expression was also forced in the aggressive prostate
cancer cell lines, PC3 and DU145. A mammalian expression plasmid
containing the matriptase-2 gene was used to produce the
overexpressed cell lines.
Materials and methods
Cell lines and culture conditions
The prostate epithelial cell line, PZHPV7, was
purchased from the American Type Culture Collection (ATCC;
Rockville, MD). PNT2C2 prostate epithelial cells were a kind gift
from Professor Norman Maitland (University of York, York, UK).
These cells were maintained in Dulbecco’s modified Eagle medium
(DMEM) (PAA Laboratories Ltd., Somerset, UK), supplemented with
penicillin, streptomycin and 10% foetal calf serum (PAA
Laboratories Ltd.) and incubated at 37°C, 5% CO2 and 95%
humidity.
Generation of cell lines displaying
altered expression of matriptase-2
The ribozyme transgene was generated using touchdown
PCR before being inserted into a pEF6/V5-His-TOPO plasmid and
cloned into E. coli. The plasmids were then checked to
ensure that the ribozyme was in the correct orientation. PZHPV7 and
PNT2C2 cells were subsequently transfected with this plasmid. The
matriptase-2 coding sequence had previously been cloned into a
pEF6/V5-His-TOPO plasmid vector in our laboratory (12). PC3 cells had also been previously
transfected with the matriptase-2 expression plasmid (13). These cells were utilised in this
study. To ensure that an efficient/stable transfection was
achieved, the cells were placed into a selection medium (5 μg/ml
blasticidin), utilising the blasticidin resistance gene coded on
the pEF6 plasmid to select cells containing the plasmid. Cells were
subsequently cultured in maintenance medium (0.5 μg/ml blasticidin)
and routinely tested to confirm matriptase-2 overexpression or
knockdown. Cells containing the ribozyme transgene were termed
PZHPV7mat2 rib1 and PNT2C2mat2 rib1, whereas
PC3 cells containing the expression plasmid were designated as
PC3mat2 exp. Similarly, cells containing a closed pEF6
plasmid (containing no expression sequence/ribozyme transgene, to
demonstrate the lack of effects exerted by the plasmid alone) were
termed PZHPV7pEF6, PNT2C2pEF6 and
PC3pEF6.
RNA extraction, quantification and
RT-PCR
Cells were grown to confluence in a 25
cm2 tissue culture flask before extracting RNA from the
cells using TRI reagent (Sigma, Dorset, UK) in accordance with the
provided protocol. Subsequently, RNA concentration was calculated
using a spectrophotometer (WPA UV 1101, Biotech Photometer,
Cambridge, UK) and samples were standardised to 250 ng prior to
undertaking reverse transcription (enhanced avian reverse
transcriptase-PCR-100; Sigma). The quality of the cDNA generated
was tested using GAPDH primers (full primer sequences are outlined
in Table I).
 | Table IPrimer sequences. |
Table I
Primer sequences.
| Molecule | Primer name | Primer sequence |
|---|
| Matriptase-2
ribozyme | Matrip2 RZ1F |
CTGCAGCACTAGAGATTCCCGGCGGGTAACTGATGAGTCCGTCAGGA |
| Matrip2 RZ1R |
ACTAGTTGTACTCAATCGGCACTTCTCCCAG |
| Matriptase-2 | Matrip2 F2 |
GAAAGACATAGCTGCATTG |
| Matrip2 R2 |
GTAGTAGCTGGGGAAGTACG |
| GAPDH | GAPDH F10 |
AGCTTGTCATCAATGGAAAT |
| GAPDH R10 |
CTTCACCACCTTCTTGATGT |
| Matriptase-2
(Q-PCR) | Matrip2 ZF |
ACTGAACCTGACCGTACAGCCGAGTACGAAGTGGAC |
| Matrip2 R2 |
GTAGTAGCTGGGGAAGTACG |
| GAPDH (Q-PCR) | GAPDH ZF |
ATGATATCGCCGCGCTCA |
| GAPDH R10 |
CTTCACCACCTTCTTGATGT |
| β-catenin | β-catenin F22 |
AAAGGCTACTGTTGGATTGA |
| β-catenin R22 |
TCCACCAGAGTGAAAAGAAC |
Quantitative-polymerase chain reaction
(Q-PCR)
Matriptase-2 transcript levels in each of the
samples were detected using the iCycler IQ system. The results are
presented as the number of transcripts/μg RNA, based on an internal
standard. Matriptase-2 transcript levels were normalised within
each sample in relation to the expression of GAPDH. For the Q-PCR
analysis, the Amplifluor system (Intergen Inc., Purchase, NY),
Q-PCR Master Mix (ABgene, Surrey, UK) and a universal probe
(Uniprimer) were used, recording emitted fluorescence. Conditions
for Q-PCR were: an initial 15 min 95°C period followed by 80 cycles
of 95°C for 15 sec, 55°C for 60 sec and 72°C for 20 sec. Full
details of primers used are presented in Table I.
SDS-PAGE and western blot analysis
Cells were grown to confluence, detached and lysed
in a lysis buffer containing 0.5% SDS, 1% Triton X-100, 2 mM
CaCl2, 100 mg/ml phenylmethylsulfonyl fluoride, 1 mg/ml
leupeptin, 1 mg/ml aprotinin and 10 mM sodium orthovanadate on a
rotor wheel for 1 h before the removal of insolubles by
centrifugation at 13,000 rpm. The Bio-Rad DC Protein Assay kit
(Bio-Rad Laboratories, Hemel Hempstead, Hertfordshire, UK) was used
to quantify protein levels in each sample and samples were
subsequently standardised to 2 mg/ml, diluted with Laemmli 2X
concentrate sample buffer (Sigma) and boiled for 5 min. Samples
were loaded, in equal quantities, onto a 10% acrylamide gel and
subjected to electrophoretic separation. The proteins were blotted
onto a Hybond-C Extra nitrocellulose membrane (Amersham Biosciences
UK Ltd., Bucks, UK), blocked in 10% milk and subjected to specific
antibody probing. Matriptase-2 expression was detected using
matriptase-2 stem region-specific antibodies (Abcam, Cambridge,
UK). The detection of the GAPDH housekeeping protein, to indicate
uniform protein expression levels in the samples, was undertaken
using an anti GAPDH antibody (Santa Cruz Biotechnology, Inc., Santa
Cruz, CA).
Immunocytochemistry (ICC)
A total of 20,000 cells in 200 μl DMEM medium were
seeded into chamber slides (Nalge Nunc International,
Lab-Tek®, Rochester, NY, USA) and left to incubate
overnight at 37°C, with 5% CO2. After incubation the
cells were fixed with 200 μl formalin for 20 min at 4°C. Subsequent
to fixation, the cells were rehydrated with 200 μl BSS for a
minimum of 30 min at room temperature. Cells were permeabilised
with 0.1% Triton X-100, then incubated with β-catenin primary
antibody for 1 h. After 4 washes with wash buffer, the cells were
incubated with 200 μl of working Vectastain® Universal
secondary antibody for 30 min, followed by extensive washing and
further incubation with ABC complex (Vector Laboratories Inc.,
Burlingame, USA). Following the removal of the ABC solution, a few
drops of DAB chromogen (Vector Laboratories Inc.) were added to the
cells and left to incubate for 5 min in the dark. The cells were
then counterstained with Mayer’s haematoxylin for approximately 1
min.
Immunofluorescent (IFC) staining
A total of 20,000 cells in 200 μl DMEM medium were
seeded into chamber slides (Nalge Nunc International,
Lab-Tek®) and left to incubate overnight at 37°C, with
5% CO2. After incubation, the medium was aspirated and
the cells fixed with 200 μl ice-cold pure ethanol for 20 min at
4°C. Subsequent to fixation, the cells were rehydrated with 200 μl
BSS for a minimum of 30 min at room temperature. Cells were
permeabilised with 0.1% Triton X-100, and incubated with β-catenin
primary antibody for 1 h. The primary antibody was then washed off
and replaced with the corresponding FITC-labelled secondary
antibody (Sigma). FluorSave (Calbiochem-Novabiochem Ltd.,
Nottingham, UK) was added to the cells before placing a cover-slip
over the slide. Following this, cells were viewed using a
fluorescent microscope.
In vitro growth assay
Cells were seeded into triplicate 96-well plates at
a density of 3,000 cells per well. Plates were then incubated for
overnight, 3- or 5-day periods before being fixed in 4%
formaldehyde (v/v) and stained with 0.5% (w/v) crystal violet. The
crystal violet stain absorbed by the cells was subsequently
extracted using 10% acetic acid (v/v) and the cell density was
determined by measuring the absorbance of this solution using a
Bio-Tek ELx800 multi-plate reader (Bio-Tek Instruments Inc,
Winooski, VT).
In vitro cell motility assay
A total of 500,000 cells were incubated in 10 ml of
growth medium containing 100 μl of cytodex-2 beads (GE Healthcare,
Cardiff, UK) for 3.5 h to allow cell adherence to the cytodex
beads. The beads were then carefully washed twice in 5 ml of normal
medium to remove non-adherent or dead cells. Following the second
wash, the beads were re-suspended in 5 ml of growth medium and 300
μl of this solution was then added to a 96-well plate and incubated
overnight at 37°C, 5% CO2 and 95% humidity. Following
incubation, the cells that had migrated from the cytodex-2 beads
and adhered to the base of the well were fixed in 4% formaldehyde
and stained with 0.5% crystal violet and counted.
In vitro Matrigel adhesion assay
A total of 45,000 cells were seeded into the wells
of a 96-well plate that had been previously coated with 5 μg of
Matrigel artificial basement membrane. Cells were incubated for 45
min before being subjected to washing in BSS to remove non-adherent
cells, after which adherent cells were fixed in 4% formaldehyde
(v/v), stained with 0.5% (w/v) crystal violet and counted under a
microscope.
In vitro invasion assay
A total of 40,000 cells were seeded into transwell
inserts containing 8-μm pores, that had previously been coated with
50 μg Matrigel, in a 24-well plate and were incubated for a period
of 3 days. Following incubation, cells which had degraded and
migrated through the Matrigel to the other side of the insert were
fixed in formaldehyde, stained with crystal violet and counted
under a microscope.
Statistical analysis
Statistical comparisons relative to the control cell
lines containing an empty pEF6 vector were made. Data were analysed
using the Minitab 14 software and statistical comparisons were
drawn using the two samples: two-tailed t-test or the Mann-Whitney
non-parametric test. All experimental procedures were repeated for
minimum of three times. A p-value <0.05 was considered to
indicate a statistically significant difference.
Results
Confirmation of knockdown in PZHPV7 and
PNT2C2 cells
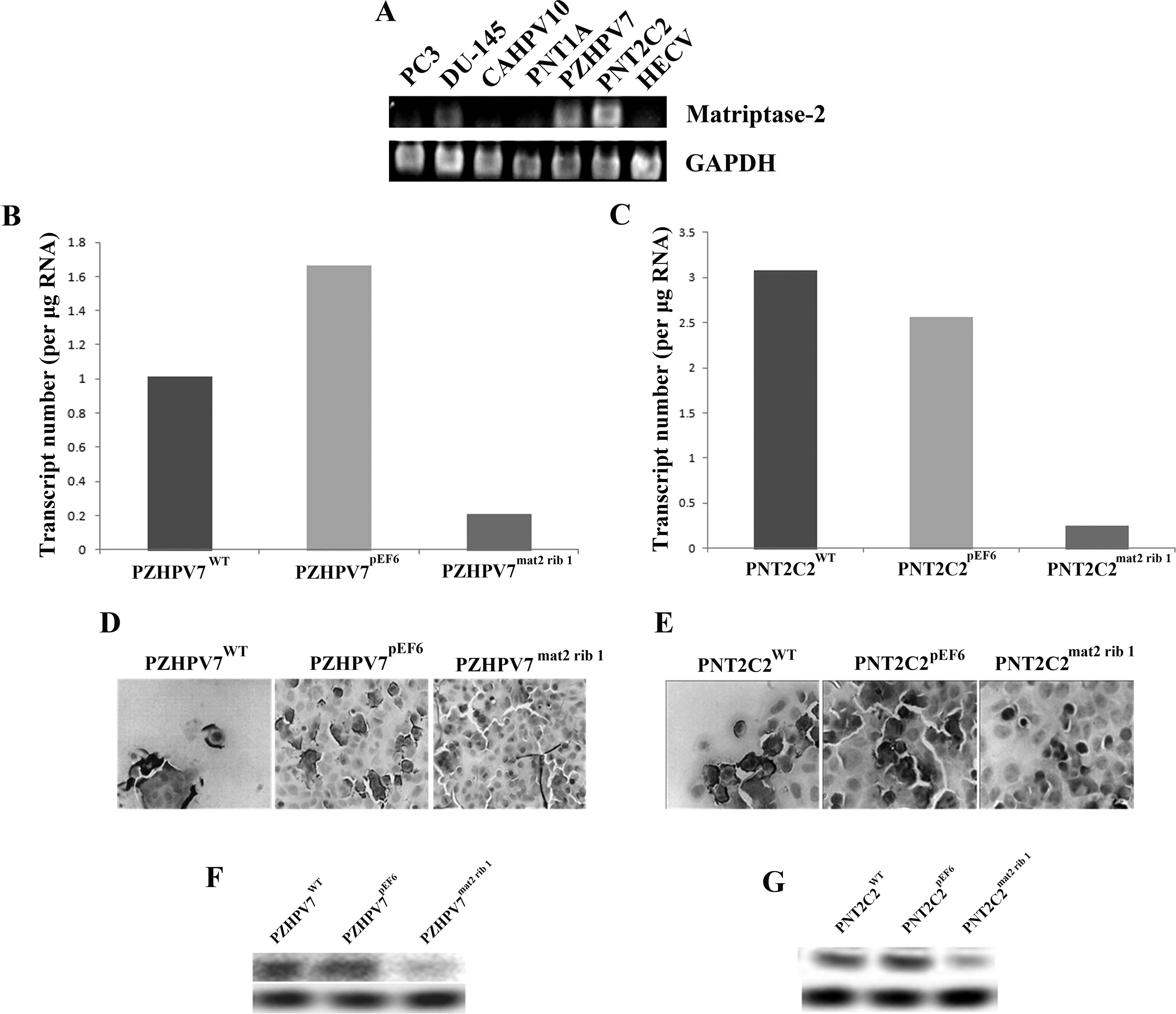
PZHPV7 and PNT2C2 cells were chosen as they
expressed matriptase-2 at a high level (Fig. 1A). To confirm matriptase-2 knockdown
at the mRNA levels, Q-PCR was performed and showed that there was a
visible reduction in matriptase-2 mRNA levels in both cells lines
compared to the respective wild-type and plasmid control cell lines
(Fig. 1B and C). To confirm
matriptase-2 knockdown at the protein level, two methods were used.
ICC staining revealed that there was indeed matriptase-2 knockdown
at the protein level. This also showed the position of matriptase-2
at the cell membrane (Fig. 1D and
E). Western blot analysis also confirmed matriptase-2 protein
knockdown in both the PZHPV7mat2 rib1 and
PNT2C2mat2 rib1 cells compared to the respective
wild-type and plasmid control cell lines (Fig. 1F and G).
Matriptase-2 has no effect on PZHPV7 or
PNT2C2 cell growth or adhesion
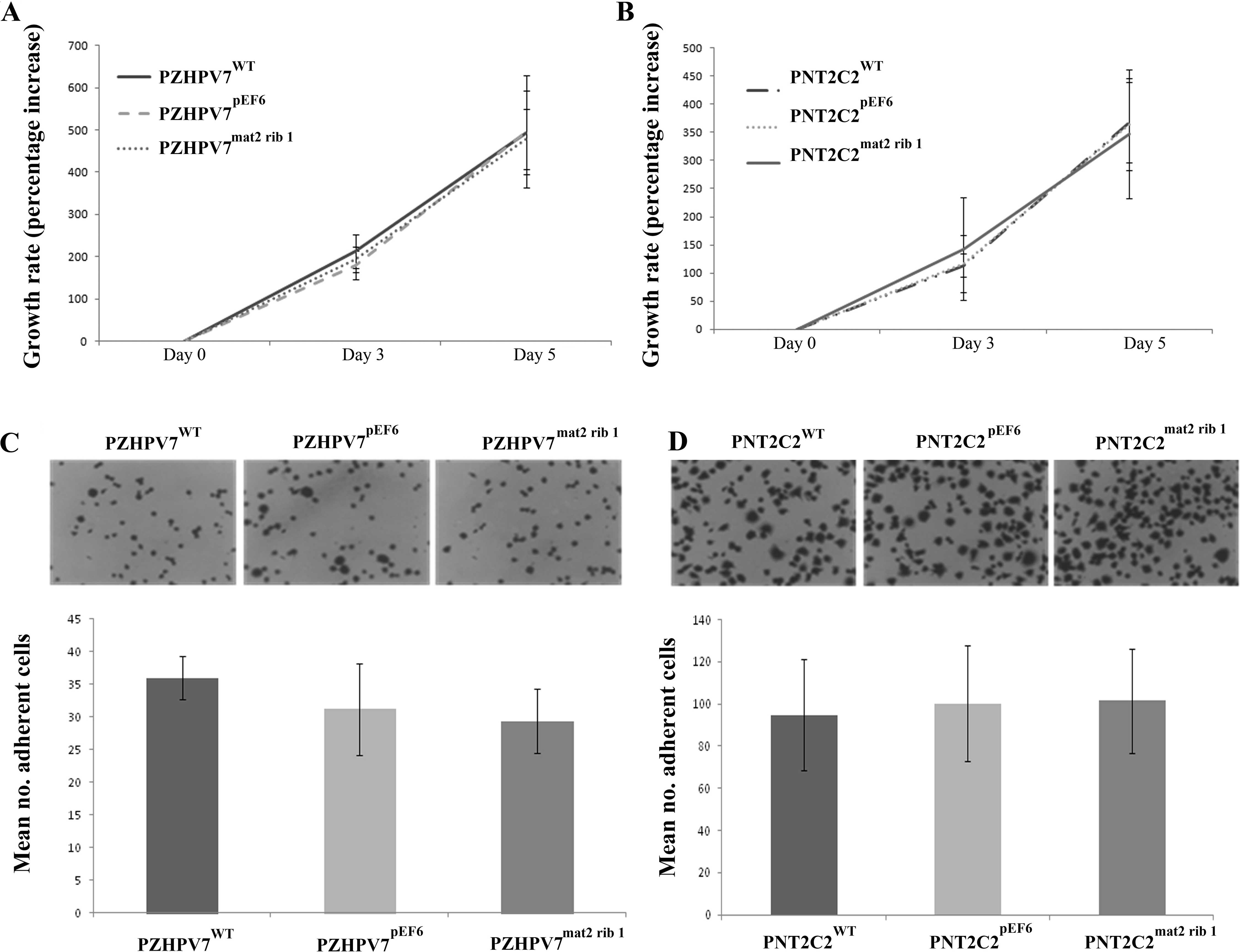
The impact of matriptase-2 knockdown appeared to
have little effect on the in vitro growth rates of PZHPV7
and PNT2C2 cells. There were no obvious differences in the growth
rates of PZHPV7mat2 rib1 cells compared to the control
cells (Fig. 2A). Cell growth at 5
days was not significantly different in the PZHPV7mat2
rib1 cells (478.20±72.27) compared to the control
PZHPV7pEF6 cells (495.88±132.99; p=0.716). Similarly,
there was also no significant difference in cell growth observed in
the PNT2C2 cell line. Cell growth at 5 days was not significantly
different in the PNT2C2mat2 rib1 cells (367.65±71.54)
compared to the PNT2C2pEF6 cells (347.14±115.04;
p=0.711) (Fig. 2B). The cells were
further analysed for their adhesive capacity in an in vitro
Matrigel adhesion assay. Cells displaying a suppressed level of
matriptase-2 displayed little difference in adhesive capacity
compared to the respective control lines. There was no significant
difference in cell adhesion in the PZHPV7mat2 rib1 cells
(29.36±4.90), compared to the control PZHPV7pEF6 cells
(31.20±7.03; p=0.465) (Fig. 2C).
Similarly, no significant difference in cell adhesion was observed
in the PNT2C2mat2 rib1 cells (101.53±24.64) compared to
the control PNT2C2pEF6 cells (100.33±27.25; p=0.900)
(Fig. 2D).
Knockdown of matriptase-2 significantly
increases the motility and invasive capabilities of PZHPV7 and
PNT2C2 cells
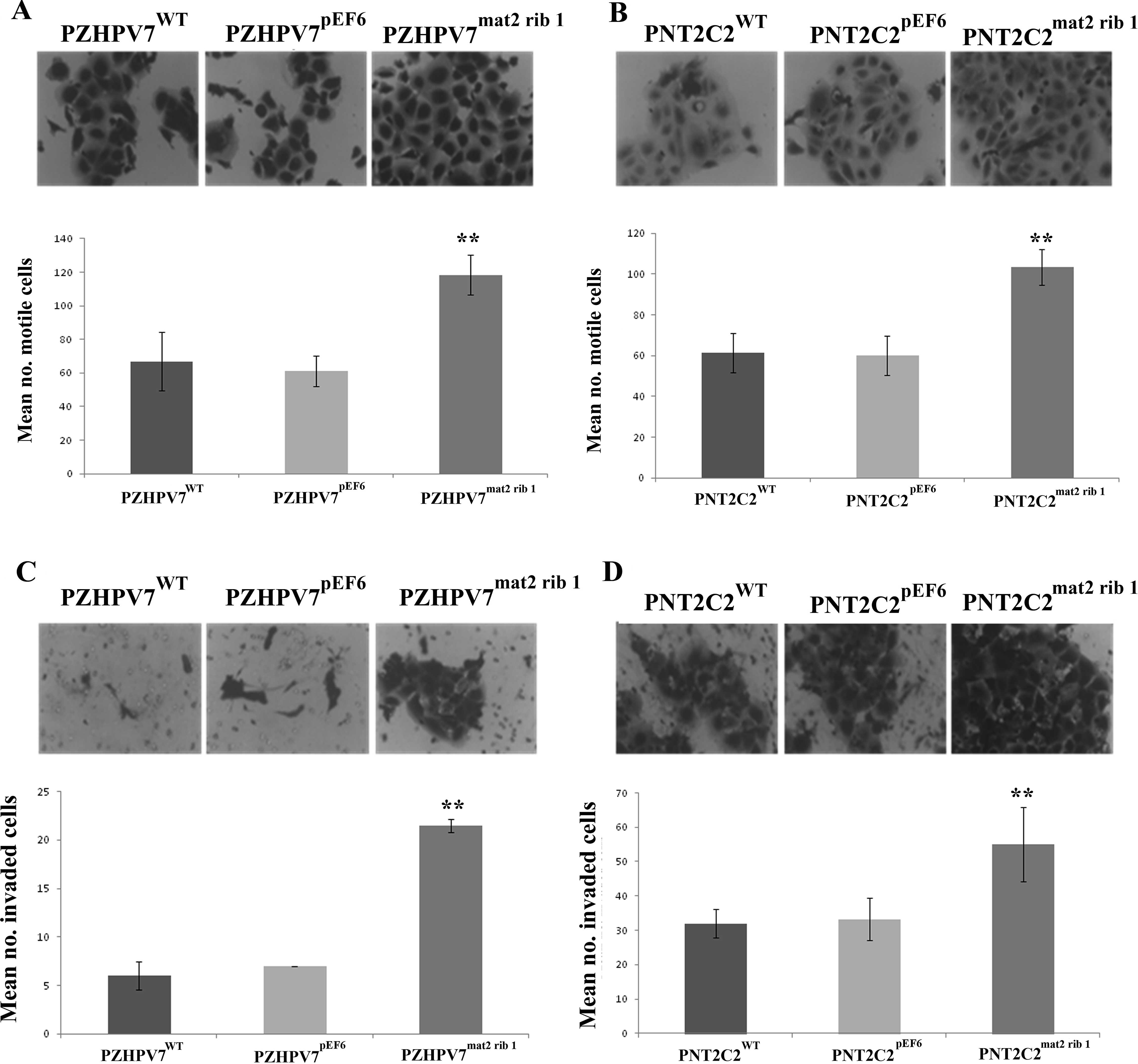
The motility of both cell lines was examined using a
cytodex-2 bead assay. The matriptase-2 knockdown cells displayed a
significant difference in their motility rates compared to control
cells. A significant increase in cell motility was observed in the
PZHPV7mat2 rib1 cells (118.33±12.02) compared to the
control PZHPV7pEF6 cells (61.20±9.05; p<0.001)
(Fig. 3A). There was also a
significant increase in cell motility in the PNT2C2mat2
rib1 cells (103.40±8.83), compared to the control
PNT2C2pEF6 cells (59.93±9.67; p<0.001) (Fig. 3B). The cells were further analysed
for their invasive capability. The knockdown of matriptase-2 in
these prostate epithelial cells significantly altered their
invasive capacities. A significant increase in invasion was
observed in the PZHPV7mat2 rib1 cells (21.50±0.70)
compared to the control PZHPV7pEF6 cells (7.00±0.00;
p=0.001) (Fig. 3C). In keeping with
this trend, a significant increase in invasiveness was also
observed in the PNT2C2mat2 rib1 cells (55.00±10.71),
compared to the control PNT2C2pEF6 cells (33.25±6.18;
p=0.007) (Fig. 3D).
Knockdown of matriptase-2 increases
expression of β-catenin protein
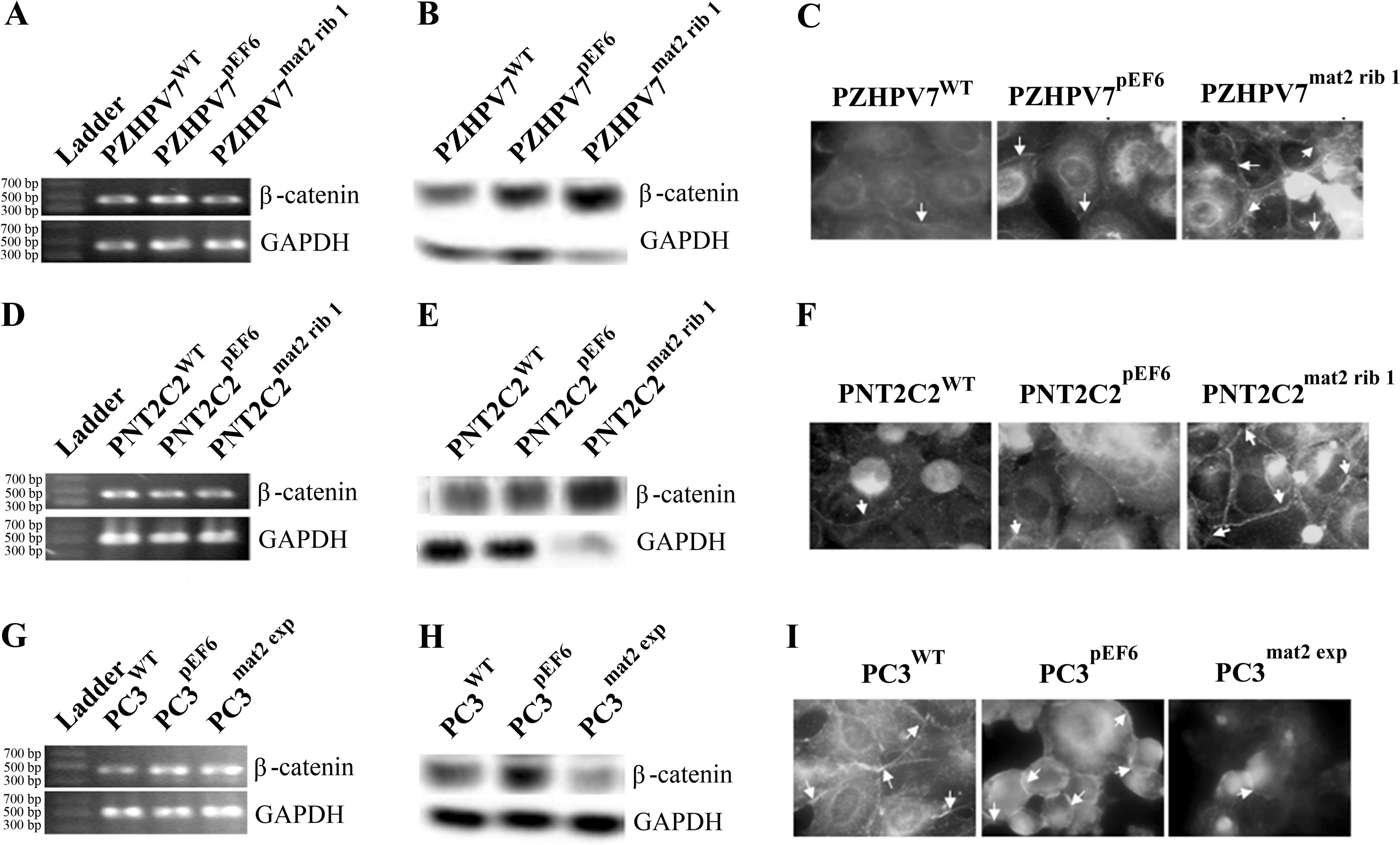
β-catenin expression was examined in PZHPV7mat2
rib1, PNT2C2mat2 rib1 and PC3mat2 exp
cells. This revealed no change in β-catenin mRNA levels in the
PZHPV7mat2 rib1 cells (Fig.
4A) but showed increased levels of β-catenin at the protein
level following western blot analysis (Fig. 4B). IFC revealed that the increased
levels of β-catenin protein were localised in the cell membrane
(Fig. 4C). PNT2C2 cells followed a
similar trend, with no obvious change in β-catenin mRNA expression
(Fig. 4D), but an increased level
of β-catenin protein expression (Fig.
4E), which was subsequently found to be localised in the cell
membrane (Fig. 4F). To further
investigate the effect of matriptase-2 on β-catenin, the PC3 cell
line overexpressing matriptase-2 was used in similar experiments.
There was no change in β-catenin mRNA levels in this cell line
(Fig. 4G). However, a decrease in
the β-catenin protein level was observed in the PC3mat2
exp cells compared to the control cells (Fig. 4H) and this reduction was apparent at
the cell membrane following IFC staining (Fig. 4I).
Discussion
An increase in the cellular levels of β-catenin
resulting from mutations in β-catenin itself, or in components of
the destruction complex, is frequently observed in many cancer
cells, including prostate cancer. The detection of β-catenin
mutations in prostate cancer samples has been reported previously,
with approximately 5% of samples revealing mutations at the serine
or threonine residues in the NH2-terminal of the β-catenin protein
(20–22). The examination of the β-catenin
protein by ICC has revealed the aberrant localization of the
protein in prostate cancer specimens (23,24).
Alterations of APC and b-TrCP1, which directly affect the
degradation of β-catenin, have also been observed in prostate
cancer samples (21). The direct
role of β-catenin in promoting prostate cancer cell growth has been
previously examined. The biological role of β-catenin in the
tumorigenesis of prostate cancer was further characterized using a
transgenic mouse model (25). The
specific expression of a mutant β-catenin, lacking exon 3, in
prostate tissues has been shown to result in the development of
prostate intraepithelial neoplasia (PIN), a precursor to prostate
cancer, in mice ranging between 10–21 weeks of age (25). However, using similar approaches,
another study observed hyperplasia of the prostate and squamous
metaplasia but no PIN lesion in a similar animal model (26). Although the precise mechanism of
β-catenin in the development of prostate cancer remains unclear,
the above data have provided a solid link between β-catenin and the
pathogenesis of prostate cancer.
To fully investigate the effect of matriptase-2 on
β-catenin, the aggressive prostate cancer cell line, PC3, was
utilised in the experiments, along with the prostate epithelial
cell lines, PZHPV7 and PNT2C2. The PC3mat2 exp cell line
was previously established within our laboratory, as this cell line
displays minimal expression levels of matriptase-2 (13). By contrast, the PZHPV7 and PNT2C2
cell lines expressed a high level of the matriptase-2 transcript
and were thus chosen as a model to target matriptase-2 expression.
Once matriptase-2 knockdown was confirmed, these model systems were
used to explore the potential correlation between matriptase-2 and
β-catenin.
Examination of the mRNA levels of β-catenin in the
matriptase-2-manipulated and respective control cell lines revealed
that matriptase-2 expression status had no effect on the transcript
levels of β-catenin. In light of this, the protein levels of
β-catenin were then examined using western blot analysis. The
western blots revealed that an overexpression of matriptase-2
greatly decreased the protein levels of β-catenin in the
PC3mat2 exp cell line. Conversely, in the matriptase-2
knockdown cells, PZHPV7mat2 rib1 and PNT2C2mat2
rib1, there was a large increase in β-catenin protein. These
results suggest a potential role for matriptase-2 in regulating
β-catenin protein expression.
To further examine the protein expression of
β-catenin, IFC was conducted on all cell lines. This also revealed
a decrease in β-catenin expression in the PC3mat2 exp
cells, where the lack of fluorescence was clearly visible. As with
the western blot analysis, the PZHPV7mat2 rib1 and
PNT2C2mat2 rib1 cells also showed an increase in
β-catenin protein staining. The regulation of β-catenin protein by
matriptase-2 was apparent at the cell membrane location. As
matriptase-2 is also found at the cell membrane, it is in the ideal
place to affect β-catenin. This change in β-catenin levels in the
cells may account for the changes in motility and invasion observed
in the cells, as β-catenin target genes are often involved in
regulating such cellular processes. The exact manner in which
matriptase-2 performs this regulatory action currently remains
unknown. One potential theory is that matriptase-2 may affect
extracellular signals that would have a knock-on effect on
β-catenin.
Matriptase-2 plays a complex role in prostate cancer
cells. This study has provided additional evidence that
matriptase-2 may have a protective function in preventing the
development and progression of prostate cancer. It may utilise its
proteolytic action toward the regulation of β-catenin to bring
about this protective function. Further investigation is required
in order to fully elucidate the correlation between matriptase-2
and β-catenin and to identify whether this may be a key mechanism
in the protective function of matriptase-2 in prostate cancer
progression.
Acknowledgements
The authors wish to thank Cancer Research Wales for
supporting this study.
References
|
1
|
Lang JC and Schuller DE: Differential
expression of a novel serine protease homologue in squamous cell
carcinoma of the head and neck. Br J Cancer. 84:237–243. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lee JW, Yong Song S, Choi JJ, et al:
Increased expression of matriptase is associated with
histopathologic grades of cervical neoplasia. Hum Pathol.
36:626–633. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Magee JA, Araki T, Patil S, et al:
Expression profiling reveals hepsin overexpression in prostate
cancer. Cancer Res. 61:5692–5696. 2001.PubMed/NCBI
|
|
4
|
Netzel-Arnett S, Hooper JD, Szabo R, et
al: Membrane anchored serine proteases: a rapidly expanding group
of cell surface proteolytic enzymes with potential roles in cancer.
Cancer Metastasis Rev. 22:237–258. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wallrapp C, Hahnel S, Muller-Pillasch F,
et al: A novel transmembrane serine protease (TMPRSS3)
overexpressed in pancreatic cancer. Cancer Res. 60:2602–2606.
2000.PubMed/NCBI
|
|
6
|
Velasco G, Cal S, Quesada V, Sanchez LM
and Lopez-Otin C: Matriptase-2, a membrane-bound mosaic serine
proteinase predominantly expressed in human liver and showing
degrading activity against extracellular matrix proteins. J Biol
Chem. 277:37637–37646. 2002. View Article : Google Scholar
|
|
7
|
Hooper JD, Campagnolo L, Goodarzi G,
Truong TN, Stuhlmann H and Quigley JP: Mouse matriptase-2:
identification, characterization and comparative mRNA expression
analysis with mouse hepsin in adult and embryonic tissues. Biochem
J. 373:689–702. 2003. View Article : Google Scholar
|
|
8
|
Szabo R and Bugge TH: Type II
transmembrane serine proteases in development and disease. Int J
Biochem Cell Biol. 40:1297–1316. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Shi YE, Torri J, Yieh L, Wellstein A,
Lippman ME and Dickson RB: Identification and characterization of a
novel matrix-degrading protease from hormone-dependent human breast
cancer cells. Cancer Res. 53:1409–1415. 1993.PubMed/NCBI
|
|
10
|
Tsai WC, Chu CH, Yu CP, et al: Matriptase
and survivin expression associated with tumor progression and
malignant potential in breast cancer of Chinese women: tissue
microarray analysis of immunostaining scores with
clinicopathological parameters. Dis Markers. 24:89–99. 2008.
View Article : Google Scholar
|
|
11
|
Uhland K: Matriptase and its putative role
in cancer. Cell Mol Life Sci. 63:2968–2978. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Parr C, Sanders AJ, Davies G, et al:
Matriptase-2 inhibits breast tumor growth and invasion and
correlates with favorable prognosis for breast cancer patients.
Clin Cancer Res. 13:3568–3576. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sanders AJ, Parr C, Martin TA, Lane J,
Mason MD and Jiang WG: Genetic upregulation of matriptase-2 reduces
the aggressiveness of prostate cancer cells in vitro and in vivo
and affects FAK and paxillin localisation. J Cell Physiol.
216:780–789. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hart M, Concordet JP, Lassot I, et al: The
F-box protein beta-TrCP associates with phosphorylated beta-catenin
and regulates its activity in the cell. Curr Biol. 9:207–210. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
He TC, Sparks AB, Rago C, et al:
Identification of c-MYC as a target of the APC pathway. Science.
281:1509–1512. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Brabletz T, Jung A, Dag S, Hlubek F and
Kirchner T: beta-catenin regulates the expression of the matrix
metalloproteinase-7 in human colorectal cancer. Am J Pathol.
155:1033–1038. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Crawford HC, Fingleton BM, Rudolph-Owen
LA, et al: The metalloproteinase matrilysin is a target of
beta-catenin transactivation in intestinal tumors. Oncogene.
18:2883–2891. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhang X, Gaspard JP and Chung DC:
Regulation of vascular endothelial growth factor by the Wnt and
K-ras pathways in colonic neoplasia. Cancer Res. 61:6050–6054.
2001.PubMed/NCBI
|
|
19
|
Boon EM, van der Neut R, van de Wetering
M, Clevers H and Pals ST: Wnt signaling regulates expression of the
receptor tyrosine kinase met in colorectal cancer. Cancer Res.
62:5126–5128. 2002.PubMed/NCBI
|
|
20
|
Chesire DR, Ewing CM, Sauvageot J, Bova GS
and Isaacs WB: Detection and analysis of beta-catenin mutations in
prostate cancer. Prostate. 45:323–334. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Gerstein AV, Almeida TA, Zhao G, et al:
APC/CTNNB1 (beta-catenin) pathway alterations in human prostate
cancers. Genes Chromosomes Cancer. 34:9–16. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Voeller HJ, Truica CI and Gelmann EP:
Beta-catenin mutations in human prostate cancer. Cancer Res.
58:2520–2523. 1998.PubMed/NCBI
|
|
23
|
Chesire DR, Ewing CM, Gage WR and Isaacs
WB: In vitro evidence for complex modes of nuclear
beta-catenin signaling during prostate growth and tumorigenesis.
Oncogene. 21:2679–2694. 2002. View Article : Google Scholar
|
|
24
|
de la Taille A, Rubin MA, Chen MW, et al:
Beta-catenin-related anomalies in apoptosis-resistant and
hormone-refractory prostate cancer cells. Clin Cancer Res.
9:1801–1807. 2003.PubMed/NCBI
|
|
25
|
Gounari F, Signoretti S, Bronson R, et al:
Stabilization of beta-catenin induces lesions reminiscent of
prostatic intraepithelial neoplasia, but terminal squamous
transdifferentiation of other secretory epithelia. Oncogene.
21:4099–4107. 2002. View Article : Google Scholar
|
|
26
|
Bierie B, Nozawa M, Renou JP, et al:
Activation of beta-catenin in prostate epithelium induces
hyperplasias and squamous transdifferentiation. Oncogene.
22:3875–3887. 2003. View Article : Google Scholar : PubMed/NCBI
|