Introduction
For advanced gastric cancers, fluoropyrimidine- and
platinum-based combination chemotherapy is currently the most
widely accepted treatment. However, its benefit has not been
translated into higher overall survival rates. Therefore, there is
a need for more effective therapies for gastric cancers.
Overexpression of HER family proteins is often
associated with tumor malignancy. Also in gastric cancers, EGFR,
HER2, and HER3 overexpression has been identified and was shown to
be associated with prognosis (1–5).
Hence, inhibiting the HER family signal transduction is likely to
contribute to improved gastric cancer therapies. Indeed, the ToGA
trial (6) demonstrated that
chemotherapeutic regimens including trastuzumab
(Herceptin®), a humanized anti-HER2 antibody,
significantly prolonged overall survival of HER2-overexpressing
metastatic gastric cancer patients compared to comparison regimens
not including trastuzumab.
Trastuzumab emtansine (T-DM1) is a HER2-targeted
antibody-drug conjugate (ADC), composed of microtubule
polymerization inhibitor DM1 (a derivative of maytansine) linked to
trastuzumab. T-DM1 is designed to deliver DM1 into
HER2-overexpressing tumor cells. Namely, T-DM1 binds to HER2 on
tumor cells, followed by internalization and degradation in
lysosomes. As a consequence, active DM1 is liberated
intracellularly from T-DM1 (7–9) and
inhibits microtubule assembly causing cell apoptosis/death
(10,11). In addition, T-DM1 has been shown to
retain the mechanism of action of trastuzumab as an antibody,
including antibody-dependent cellular cytotoxicity (ADCC),
inhibition of cell signaling through the phosphatidylinositol
3-kinase (PI3K)/AKT pathway, and inhibition of HER2 shedding
(12,13). To investigate the clinical
effectiveness of T-DM1, two phase III trials, EMILIA (evaluating
T-DM1 compared with lapatinib plus capecitabine for patients with
HER2-positive metastatic breast cancer previously treated with
trastuzumab and a taxane) and MARIANNE (evaluating T-DM1 plus
placebo vs. T-DM1 plus pertuzumab vs. trastuzumab plus a taxane for
patients with previously untreated HER2-positive metastatic breast
cancer) (14) are under way.
Pertuzumab is a humanized anti-HER2 antibody that
exerts antitumor activity in a different manner from trastuzumab.
Whereas trastuzumab binds to domain IV of the HER2 extracellular
domain (ECD), pertuzumab binds to domain II of the HER2 ECD, the
region responsible for dimer formation. Thus, pertuzumab inhibits
the dimerization of HER2 with other HER family proteins such as
EGFR or HER3, which results in prevention of ligand-dependent HER2
signaling (15). Recently, the
efficacy of pertuzumab in HER2-positive metastatic breast cancer
was demonstrated in a phase III trial (CLEOPATRA, evaluating
trastuzumab plus docetaxel vs. trastuzumab plus docetaxel plus
pertuzumab), which showed significantly prolonged progression-free
survival (PFS) (16).
Thus, therapies containing anti-HER2 antibodies are
expected to be of great clinical significance. In a previous study,
we showed an enhanced antitumor activity of trastuzumab in
combination with pertuzumab and described its mechanism of action
in a HER2-positive gastric cancer model (17). In this context, we believe that the
combination of T-DM1 with pertuzumab could be a new potential
therapy for advanced gastric cancer. Therefore, in the present
study, we investigated the combination effects of T-DM1 and
pertuzumab on antitumor activities in vitro and in
vivo using HER2-positive human gastric cancers.
Materials and methods
Test agents
T-DM1 and pertuzumab were provided by F. Hoffmann-La
Roche (Basel, Switzerland). Both antibodies were diluted with
saline or culture medium in in vivo or in vitro
experiments, respectively. Human immunoglobulin G (IgG) was
purchased from MP Biomedicals, Inc. (Aurora, OH, USA) and
reconstituted with water and diluted with saline.
Animals
Five-week-old male BALB-nu/nu mice
(CAnN.Cg-Foxn1<nu>/CrlCrlj nu/nu) were obtained from Charles
River Laboratories Japan (Yokohama, Japan). All animals were
acclimatized for 1 week prior to the study. The health of the mice
was monitored daily. Chlorinated water and irradiated food were
provided ad libitum, and the animals were maintained under a
controlled light-dark cycle (12 h-12 h). All animal experiments
were conducted in accordance with the Institutional Animal Care and
Use Committee.
Cell lines and culture
Five human gastric cancer cell lines were used in
the present study: NCI-N87, SNU-16, SCH, MKN-28 and 4-1ST. NCI-N87
and SNU-16 cells were purchased from ATCC. SCH cells were purchased
from Japan Health Science Foundation (Osaka, Japan). MKN-28 cells
were purchased from Immuno-Biological Laboratories Co., Ltd.
(Fujioka, Japan). NCI-N87, SNU-16, SCH and MKN-28 were maintained
in RPMI-1640 (Sigma-Aldrich, St. Louis, MO, USA), supplemented with
10% FBS at 37°C under 5% CO2. 4-1ST cells were purchased
from the Central Institute for Experimental Animals (Yokohama,
Japan) and maintained in BALB-nu/nu mice by subcutaneous (s.c.)
inoculation of pieces of tumor. CD16(158V)/NK-92 cells were
constructed as previously described (17) and were maintained in MEMα (Wako Pure
Chemical Industries, Osaka, Japan) supplemented with 12.5% FBS,
12.5% horse serum, 0.02 mM folic acid, 0.1 mM 2-mercaptoethanol,
0.2 mM inositol, 0.5 mg/ml G418, and 20 ng/ml recombinant human
IL-2 at 37°C under 5% CO2.
Immunohistochemistry and fluorescent in
situ hybridization
As previously described, HER2 protein expression and
HER2 gene amplification in tumors were examined by
immunohistochemistry (IHC) using HercepTest™ (Dako Japan, Tokyo,
Japan) and fluorescent in situ hybridization (FISH) using
Path Vysion® (Abbott Japan, Tokyo, Japan), respectively
(18).
In vivo tumor growth inhibition
studies
Each mouse was inoculated subcutaneously in the
right flank with either 5×106 cells of human gastric
cancer cell lines NCI-N87, SCH, SNU-16 or MKN-28, or with an
~8-mm3 piece of 4-1ST tumor tissue. Several weeks after
tumor inoculation, mice were randomly allocated to control and
treatment groups.
The administration of anticancer agents was started
when the tumor volumes reached ~150–350 mm3. T-DM1 was
administered intravenously once every 3 weeks and pertuzumab was
administered intraperitoneally once a week for 6 weeks. To evaluate
the antitumor activity of the test agents, tumor volume was
measured twice a week and the percentage of tumor growth inhibition
(TGI%) was calculated as previously described (19).
In vitro anti-proliferation assays
NCI-N87 cells were seeded on 96-well plates at
1×104 cells/well and pre-cultured for 24 h. Then the
cells were starved in serum-free RPMI-1640 for 24 h and treated
with pertuzumab, T-DM1, or both. Following 30 min of incubation,
cells were exposed to 100 ng/ml of EGF or heregulin (HRG)-α and
incubated for 2 days. Cells were fixed with 10% formalin neutral
buffer solution. Crystal violet staining and extraction were
performed as previously described (18) and absorbance was measured at 595 nm.
Cells pre-cultured for 24 h were also detected by crystal violet
staining as the absorbance of the pre-cultured well. The percentage
of cell proliferation (% proliferation) was calculated as follows:
% proliferation = (absorbance of treatment well - absorbance of
pre-cultured well)/(absorbance of unstimulated well - absorbance of
pre-cultured well) ×100.
Apoptosis assay
NCI-N87 cells were seeded on 96-well plates at
1×104 cells/well and cultured in 0.1% FBS medium with
100 ng/ml of EGF or HRG-α added for 24 h. After that, cells were
treated with pertuzumab, T-DM1, or both. After 24 h of treatment,
Caspase-Glo® 3/7 Assay (Promega, Madison, WI, USA) was
used to measure caspase 3/7 activity, which was calculated as
follows: caspase 3/7 activity = (luminescent unit of treatment well
- luminescent unit of blank well)/(mean luminescent unit of control
well).
Western blotting
NCI-N87 cells were seeded in 0.1% FBS RPMI-1640 for
24 h and then treated with IgG as the control or with 40 μg/ml
pertuzumab, 5 μg/ml T-DM1, or both. Following incubation for 1 h,
cells were exposed to EGF or HRG-α for 1 or 5 min, respectively.
Cells were washed with ice-cold PBS and then lysed with Cell Lysis
Buffer (Cell Signaling Technology, Beverly, MA, USA) with 10 mM
NaF, 1 μg/ml aprotinin, and 1 mM PMSF. After centrifugation (4°C,
14,000 × g, 5 min), the resultant supernatants were used for the
western blot assays.
To prepare in vivo samples, tumor samples
were taken on day 5 (4 days after treatment) and immediately frozen
in liquid nitrogen and stored at −80°C. Tumor samples were
homogenized in Cell Lysis Buffer, and centrifuged (4°C, 10,000 × g,
20 min). The resultant supernatants were used for the western blot
assays.
The protein concentration of the supernatant was
quantified using the DC protein assay kit (Bio-Rad Laboratories,
Hercules, CA, USA). The supernatants were denatured for 5 min at
95°C, electrophoresed on SDS-PAGE and transferred to a
polyvinylidene difluoride membrane (Millipore, Bedford, MA, USA).
The membrane was blocked in SuperBlock® T20 (TBS)
Blocking Buffer (Thermo Scientific, Waltham, MA, USA) and probed
with each antibody against HER3 (sc-285), (Santa Cruz
Biotechnology, Santa Cruz, CA, USA), HER2 (#2248), EGFR (#2963),
pEGFR (Tyr1068) (#3777), pHER2 (Tyr1248) (#2247), pHER3 (Tyr1289)
(#4791), AKT (#9272), pAkt (Ser473) (#9271), ERK1/2 (#9102),
pERK1/2 (Thr202/Tyr204) (#9106) (Cell Signaling Technology), and
actin (A2228) (Sigma-Aldrich) as primary antibodies. These proteins
were detected by horseradish peroxidase-conjugated secondary
antibodies. The bands were visualized using Chemi-Lumi One Super
(Nacalai Tesque, Kyoto, Japan).
ADCC assays
We used Real-time Cell Analyzer (xCELLigence, Roche
Diagnostics, Tokyo, Japan) to monitor ADCC activity in real-time.
The system detects electrical impedance on the bottom of the tissue
culture E-Plates, which contain interdigitated electrodes as the
Cell Index. NCI-N87 cells were seeded on E-plates at
5×103 cells/well and pre-cultured for 24 h.
CD16(158V)/NK-92 cells were added as effector cells at a target
ratio of 1:1. Cells were treated with pertuzumab, T-DM1, or both
concurrently. The Cell Index was measured every 5 min for the first
4 h and every 10 min thereafter. The Normalized Cell Index was
calculated as follows: (Cell Index at each point)/(Cell Index at
the point when pertuzumab and T-DM1 treatment started).
Statistical analysis
The Wilcoxon test was used to detect differences in
tumor volume for in vivo experiments and Student's t-test
was used for in vitro experiments, with P<0.05 considered
statistically significant. Statistical analyses were carried out
using the SAS preclinical package (SAS Institute, Tokyo,
Japan).
Results
Antitumor activity of T-DM1 as a single
agent in human gastric cancer mouse xenograft models
Antitumor activity of T-DM1 was evaluated in 5
gastric tumor models with various HER2 levels. NCI-N87, 4-1ST and
SCH xenografted tumors exhibited high HER2 expression with IHC
scores of 3+ or 2+ and a FISH HER2/CEP17 ratio >2.0. SNU-16 and
MKN-28 showed low HER2 expression with IHC scores of 1+ or 0 and a
<2.0 FISH ratio. On day 22 (21 days after treatment), T-DM1 (20
mg/kg) showed significant antitumor activity in the xenograft
models with high HER2 expression and decreasing tumor volumes.
However, T-DM1 did not show significant efficacy for tumors with
low HER2 expression (Table I).
 | Table IAntitumor activity of T-DM1 as a
single agent in human gastric cancer mouse xenograft models. |
Table I
Antitumor activity of T-DM1 as a
single agent in human gastric cancer mouse xenograft models.
| Tumor name | HER2 IHC score | HER2/CEP17 ratio
(FISH) | Tumor volume
(mm3) | TGI% |
|---|
|
|---|
| Treatment | Day 1 | Day 22 |
|---|
| NCI-N87 | 3+ | 8.4b | Control | 174±25 | 667±152 | 133 |
| | | T-DM1 | 174±24 | 11±9a | |
| 4-1ST | 3+ | 16.5 | Control | 283±75 | 1320±396 | 126 |
| | | T-DM1 | 283±72 | 12±18a | |
| SCH | 2+ | 2.1 | Control | 240±51 | 1565±223 | 109 |
| | | T-DM1 | 241±63 | 121±25a | |
| SNU-16 | 1+ | 1.4b | Control | 185±23 | 485±116 | 39 |
| | | T-DM1 | 184±16 | 369±47 | |
| MKN-28 | 0 | 1.0b | Control | 271±50 | 1662±286 | 27 |
| | | T-DM1 | 269±52 | 1290±507 | |
Antitumor activity of T-DM1 in
combination with pertuzumab in a HER2-positive gastric cancer mouse
xenograft model
We assessed the combined efficacy of T-DM1 with
pertuzumab using an NCI-N87 xenograft model which expressed high
levels of HER2 and was sensitive to T-DM1 treatment. Compared to
each antibody alone, T-DM1 in combination with pertuzumab showed
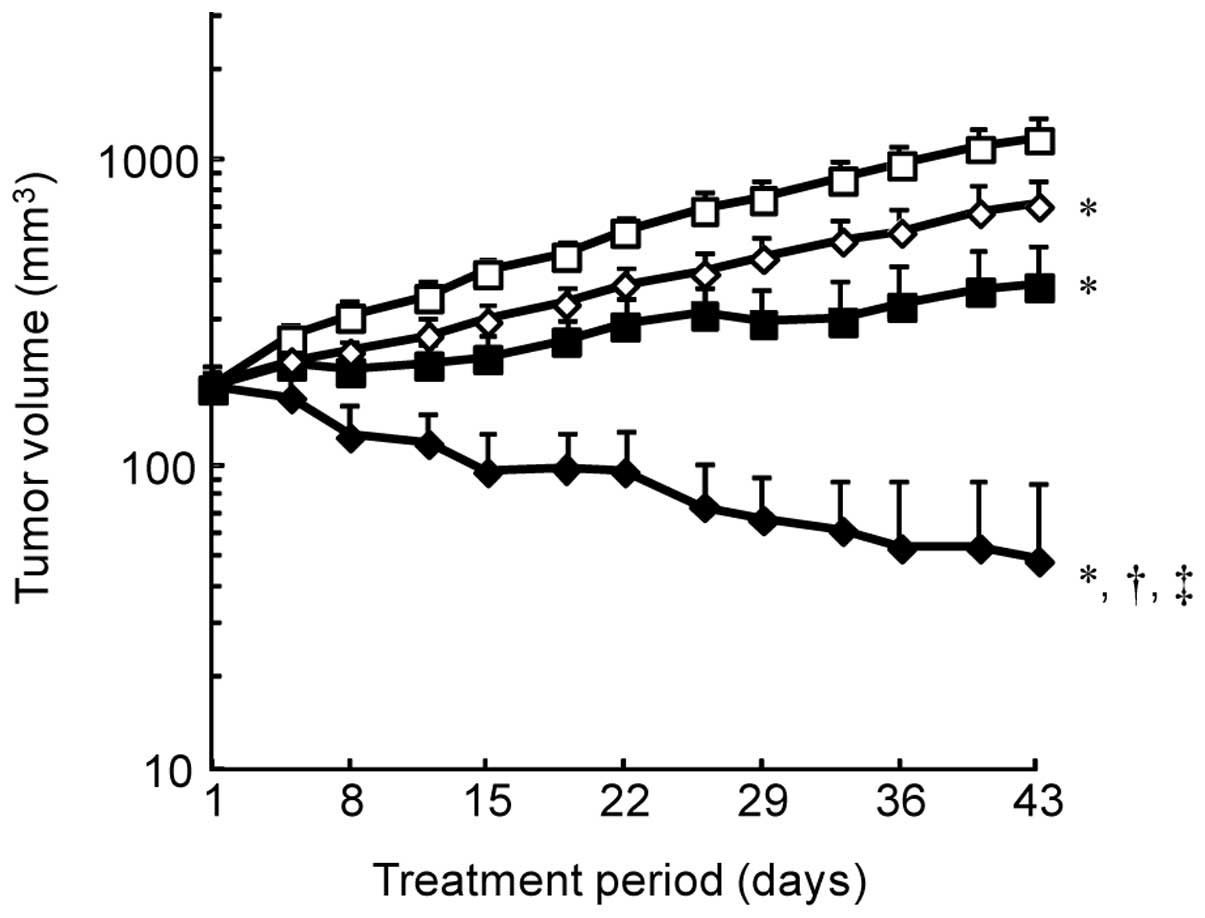
significantly enhanced antitumor activity (Fig. 1).
Combination effect of T-DM1 and
pertuzumab on proliferation and apoptosis in a HER2-positive human
gastric cancer cell line
To investigate the mechanism of action of T-DM1 in
combination with pertuzumab, we examined the growth inhibitory
effect of T-DM1 and pertuzumab in NCI-N87 cells. These cells have
been previously shown to be positive for both HER2 and EGFR and
also express low levels of HER3 (17). Pertuzumab completely blocked
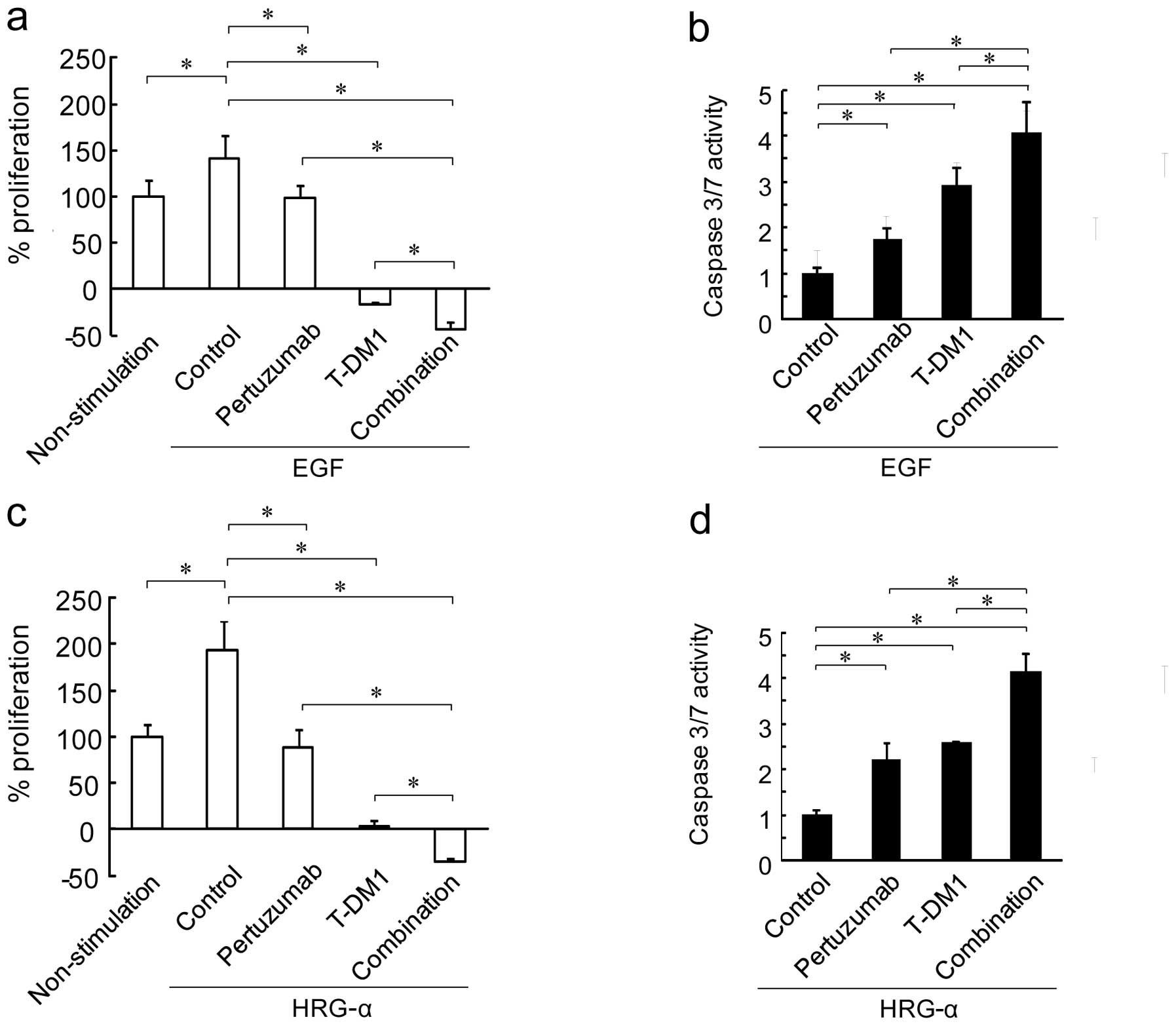
EGF-stimulated cell proliferation, and T-DM1 stopped cell growth at
the initial cell number. In the presence of EGF, T-DM1 combined
with pertuzumab significantly enhanced the inhibition of cell
proliferation compared to T-DM1 alone (Fig. 2a). We also examined the apoptotic
activity of these agents by measuring caspase 3/7 activity.
Treatment with both pertuzumab and T-DM1 significantly induced
caspase 3/7 activity compared to treatment with either single
agent, indicating that pertuzumab in combination with T-DM1
significantly enhanced the apoptosis-inducing activity of T-DM1
(Fig. 2b).
Then, we evaluated the growth inhibition and
apoptotic activity induced by T-DM1 in combination with pertuzumab
in NCI-N87 cells when stimulated with HRG-α. Similar to the
stimulation with EGF, combining pertuzumab and T-DM1 significantly
enhanced the anti-proliferative activity in HRG-α-stimulated cells.
Likewise, the increased caspase 3/7 activity seen with pertuzumab
and T-DM1 as single agents was further enhanced by their
combination under HRG-α (Fig. 2c and
d). These results suggest that T-DM1 in combination with
pertuzumab has the potential to suppress ligand-dependent cell
growth by inducing apoptosis.
Effect of T-DM1 in combination with
pertuzumab on HER2 signal transduction
To examine whether pertuzumab and T-DM1 could
inhibit HER2 signal transduction, we examined the phosphorylation
status of the molecules in HER2-EGFR or HER2-HER3 signaling
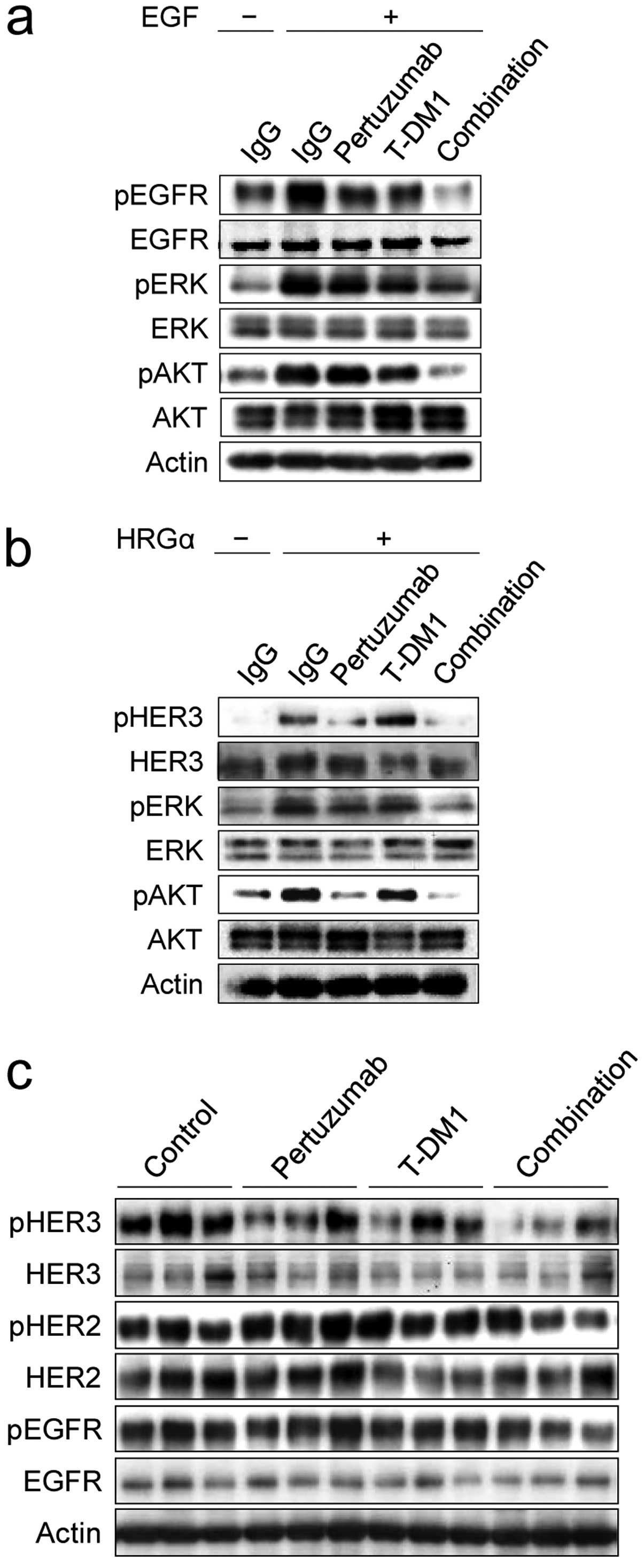
pathways. Under EGF stimulation, only T-DM1 combined with
pertuzumab strongly suppressed phosphor(p)-EGFR, pERK and pAKT
(Fig. 3a). Under HRG-α stimulation,
pertuzumab alone strongly inhibited pHER3 and, in combination with
T-DM1, almost completely suppressed pHER3 and downstream pAKT.
T-DM1 in combination with pertuzumab further reduced pERK in
comparison with pertuzumab or T-DM1 alone (Fig. 3b).
To confirm whether the HER family signaling pathway
was also inhibited by this therapy in xenografted tumors, tumor
specimens collected 4 days after treatment were tested for the
phosphorylation level of HER family receptors: pHER2, pEGFR and
pHER3. pHER2 and pEGFR were slightly reduced whereas pHER3 was
clearly reduced by the combination of T-DM1 with pertuzumab
compared to either T-DM1 or pertuzumab alone (Fig. 3c).
ADCC activity of T-DM1 in combination
with pertuzumab
One way to interpret the different antitumor potency
observed in vitro and in vivo is to consider an
immune effector-mediated activity, such as ADCC. Hence, we examined
whether T-DM1-induced ADCC activity in NCI-N87 cells was further
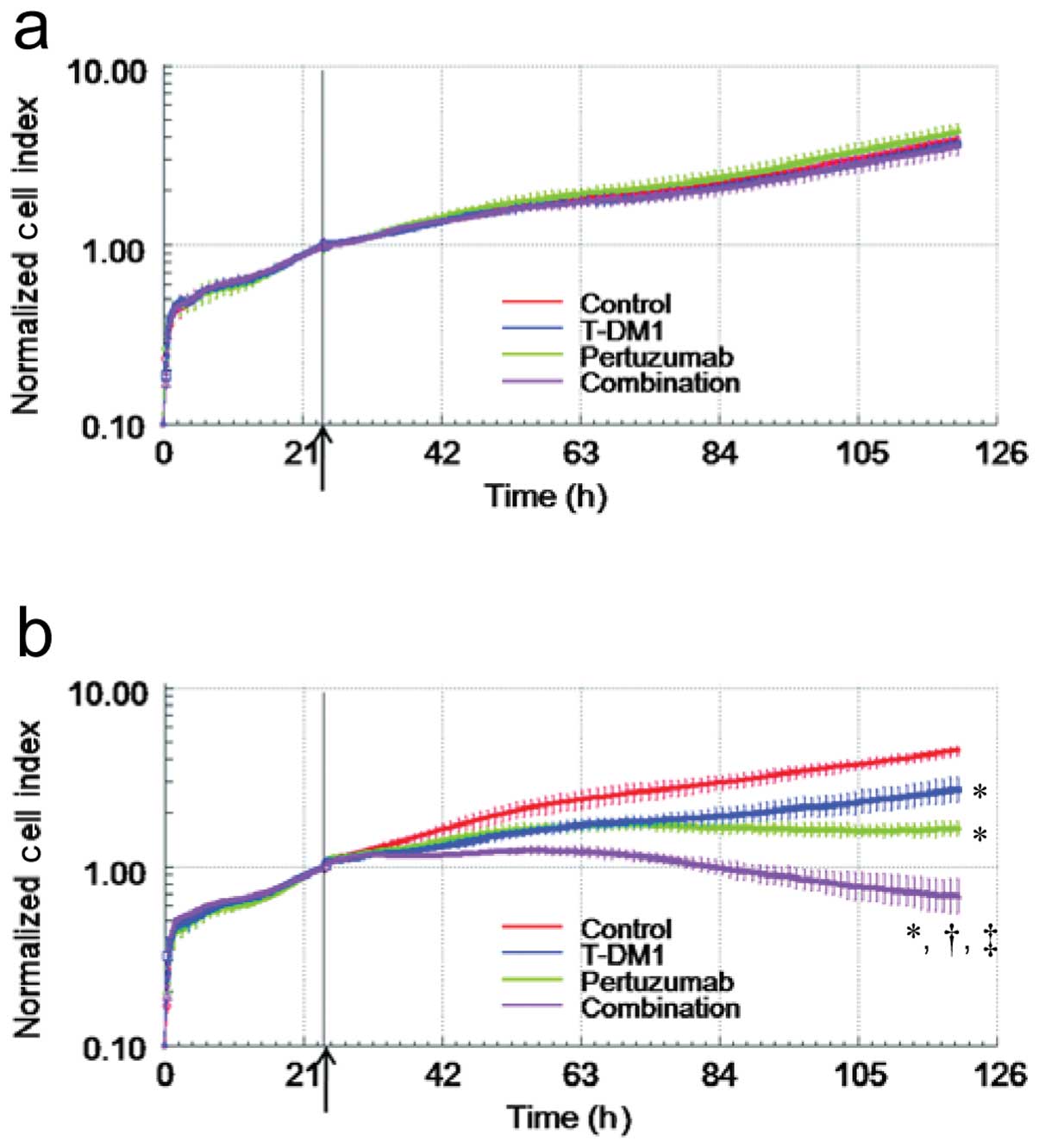
enhanced by pertuzumab, using a real-time cell analyzer. In the
wells without effector cells, the addition of pertuzumab and T-DM1
did not reduce the Normalized Cell Index (Fig. 4a). With effector cells, however, the
Normalized Cell Index was significantly reduced by the addition of
each single agent. Furthermore, T-DM1 in combination with
pertuzumab significantly enhanced the reduction of the Normalized
Cell Index induced by either T-DM1 or pertuzumab (Fig. 4b), indicating that the combination
treatment of T-DM1 and pertuzumab enhanced ADCC activity in
vitro.
Discussion
First, we clarified whether the relationship between
HER2 expression level and T-DM1 efficacy in a gastric cancer model
was comparable to that of trastuzumab. In breast cancer cells, it
is known that T-DM1 shows great antitumor potency to cancer cells
with high HER2 expression but less efficacy to those with low HER2
level (8). Our study using gastric
cancer xenograft models similarly showed that T-DM1 demonstrated
significant antitumor activity only in cancers in which HER2 status
was positive (the IHC score was 3+ or 2+, and the FISH ratio was
>2.0), suggesting that, as with trastuzumab, efficacy of T-DM1
is associated with HER2 status. On the other hand, antitumor
potency of T-DM1 for NCI-N87 (TGI% was 133%) was much higher than
that of trastuzumab administered at the maximum effective dose in a
previous study (TGI% was 89%) (18). Even in SCH, in which trastuzumab did
not show significant antitumor activity, T-DM1 showed clear
antitumor efficacy (TGI was 109%). Also T-DM1 was efficacious to
trastuzumab-insensitive xenograft tumors which expressed HER2
(8). These results confirmed that
T-DM1 has more potent antitumor activity than trastuzumab. Further
research both in clinical and preclinical studies, however, is
required to examine the efficacy of T-DM1 for tumors expressing low
levels of HER2, or that are trastuzumab-refractory.
There are some clinical studies indicating that the
combination of pertuzumab and trastuzumab was active and well
tolerated in patients with metastatic HER2-positive breast cancer
(20). In addition, the CLEOPATRA
trial showed that pertuzumab significantly improved PFS of breast
cancer patients in combination with trastuzumab plus docetaxel
(16) and the combination was thus
approved by the FDA. T-DM1 was reported to have significantly
improved the PFS of the combination of trastuzumab and docetaxel
(21). In addition, the potency of
T-DM1 and pertuzumab in combination as a first line treatment in
HER2-positive metastatic breast cancers is currently being
investigated in the MARIANNE trial. Also, in the ToGA trial
investigating gastric cancer, patients with HER2-positive gastric
cancer who received trastuzumab in addition to chemotherapy
achieved a significant improvement in overall survival compared to
treatment with chemotherapy alone (6). From these lines of clinical evidence,
next, we studied the combined efficacy of T-DM1 and pertuzumab in
HER2-positive gastric cancers. In our in vivo study, the
combination treatment of T-DM1 with pertuzumab showed significantly
stronger antitumor activity compared to either single agent in the
HER2-positive NCI-N87 model, similarly to trastuzumab in
combination with pertuzumab (17).
T-DM1 in combination with pertuzumab may show strong efficacy in
HER2-positive gastric cancer.
Our study demonstrated that the combination of two
different anti-HER2 antibodies with different HER2-binding sites
enhanced the inhibition of HER2 signal transduction in cultured
NCI-N87 cells. Under EGF stimulation, the combination strongly
reduced the phosphorylation of EGFR and its downstream factors, ERK
and AKT compared to either single agent. Similar results were shown
in our previous study in which the combination of trastuzumab and
pertuzumab suppressed pEGFR and pHER2 and their downstream pERK and
pAKT by inhibiting the EGFR-HER2 heterodimerization more strongly
than either single agent (17).
These results suggest that T-DM1 in combination with pertuzumab
also suppressed heterodimer formation of EGFR-HER2.
In the NCI-N87 cells stimulated with HRG-α,
concomitant treatment of T-DM1 and pertuzumab almost completely
reduced the phosphorylation of HER3 and AKT, and more potently
inhibited cell growth and increased apoptotic cells. Furthermore,
in the xenografted NCI-N87 tumors, western blot analysis revealed
that the combination suppressed the phosphorylation of HER3 not but
EGFR. Considering the studies that HER2-HER3 heterodimer is the
most active signal transducer among HER family complexes (22,23),
it is possible that disruption of the signal of HER3 rather than
EGFR may play an important role in the antitumor efficacy of the
combination treatment, even though HER3 expression levels were low
in NCI-N87 tumors. Although further study is required to elucidate
the mechanism of action, suppression of AKT signal followed by
apoptosis induction in tumor may contribute to the enhanced
antitumor efficacy by pertuzumab combined with T-DM1.
T-DM1 retains the ADCC that is reported as a key
mechanism of trastuzumab (24), and
pertuzumab also had ADCC (25,26).
The results of the present study, which are consistent with the
enhanced ADCC activity of pertuzumab combined with trastuzumab in
our previous study (17),
demonstrate that in the combination treatment using T-DM1 with
pertuzumab the ADCC activity was significantly enhanced compared to
either single agent alone. On the other hand, a study has shown
that the simultaneous binding of trastuzumab and pertuzumab onto
target cells did not mediate a synergistic effect on ADCC when they
were used at maximum effective doses (25). The authors showed that the level of
ADCC of the combination of trastuzumab and pertuzumab was the same
as those of the single antibodies, when the final total antibody
concentration was identical. The discrepancy may arise from the
difference in Materials and methods; that is, we used a real-time
cell analyzer to measure ADCC and the sum of the concentration of
each antibody was used as the combinational concentration. The
results of the present study confirm that, in addition to enhanced
inhibition of HER2 signaling, ADCC may be another mechanism of
action that explains the increased efficacy of the combination.
In conclusion, T-DM1 in combination with pertuzumab
achieved significant antitumor activity by enhancing HER family
signal inhibition and ADCC activity in a HER2-positive human
gastric cancer model. These findings suggest that T-DM1 in
combination with pertuzumab is a rational and promising treatment
option for HER2-positive gastric cancer.
Acknowledgements
We would like to thank Kumiko Kondo, Hiromi Sawamura
and Masako Takahashi (Product Research department, Chugai) for
their excellent technical assistance; Editing Services, Chugai for
helpful advice about drafting the manuscript; and Dr Kazushige Mori
for helpful discussion and comments regarding the study.
References
|
1
|
Garcia I, Vizoso F, Martin A, et al:
Clinical significance of the epidermal growth factor receptor and
HER2 receptor in resectable gastric cancer. Ann Surg Oncol.
10:234–241. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Yonemura Y, Ninomiya I, Yamaguchi A, et
al: Evaluation of immunoreactivity for erbB-2 protein as a marker
of poor short term prognosis in gastric cancer. Cancer Res.
51:1034–1038. 1991.PubMed/NCBI
|
|
3
|
Tanner M, Hollmen M, Junttila TT, et al:
Amplification of HER-2 in gastric carcinoma: association with
Topoisomerase IIalpha gene amplification, intestinal type, poor
prognosis and sensitivity to trastuzumab. Ann Oncol. 16:273–278.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Nicholson RI, Gee JM and Harper ME: EGFR
and cancer prognosis. Eur J Cancer. 37(Suppl 4): S9–S15. 2001.
View Article : Google Scholar
|
|
5
|
Hayashi M, Inokuchi M, Takagi Y, et al:
High expression of HER3 is associated with a decreased survival in
gastric cancer. Clin Cancer Res. 14:7843–7849. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bang YJ, Van Cutsem E, Feyereislova A, et
al: Trastuzumab in combination with chemotherapy versus
chemotherapy alone for treatment of HER2-positive advanced gastric
or gastro-oesophageal junction cancer (ToGA): a phase 3,
open-label, randomised controlled trial. Lancet. 376:687–697. 2010.
View Article : Google Scholar
|
|
7
|
Kovtun YV and Goldmacher VS: Cell killing
by antibody-drug conjugates. Cancer Lett. 255:232–240. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lewis Phillips GD, Li G, Dugger DL, et al:
Targeting HER2-positive breast cancer with trastuzumab-DM1, an
antibody-cytotoxic drug conjugate. Cancer Res. 68:9280–9290.
2008.PubMed/NCBI
|
|
9
|
Erickson HK, Lewis Phillips GD, Leipold
DD, et al: The effect of different linkers on target cell
catabolism and pharmacokinetics/pharmacodynamics of trastuzumab
maytansinoid conjugates. Mol Cancer Ther. 11:1133–1142. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Oroudjev E, Lopus M, Wilson L, et al:
Maytansinoid-antibody conjugates induce mitotic arrest by
suppressing microtubule dynamic instability. Mol Cancer Ther.
9:2700–2713. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lopus M, Oroudjev E, Wilson L, et al:
Maytansine and cellular metabolites of antibody-maytansinoid
conjugates strongly suppress microtubule dynamics by binding to
microtubules. Mol Cancer Ther. 9:2689–2699. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Junttila TT, Li G, Parsons K, Phillips GL
and Sliwkowski MX: Trastuzumab-DM1 (T-DM1) retains all the
mechanisms of action of trastuzumab and efficiently inhibits growth
of lapatinib insensitive breast cancer. Breast Cancer Res Treat.
128:347–356. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Barok M, Tanner M, Koninki K and Isola J:
Trastuzumab-DM1 is highly effective in preclinical models of
HER2-positive gastric cancer. Cancer Lett. 306:171–179. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
LoRusso PM, Weiss D, Guardino E, Girish S
and Sliwkowski MX: Trastuzumab emtansine: a unique antibody-drug
conjugate in development for human epidermal growth factor receptor
2-positive cancer. Clin Cancer Res. 17:6437–6447. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Franklin MC, Carey KD, Vajdos FF, Leahy
DJ, de Vos AM and Sliwkowski MX: Insights into ErbB signaling from
the structure of the ErbB2-pertuzumab complex. Cancer Cell.
5:317–328. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Baselga J, Cortes J, Kim SB, et al:
Pertuzumab plus trastuzumab plus docetaxel for metastatic breast
cancer. N Engl J Med. 366:109–119. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yamashita-Kashima Y, Iijima S, Yorozu K,
et al: Pertuzumab in combination with trastuzumab shows
significantly enhanced antitumor activity in HER2-positive human
gastric cancer xenograft models. Clin Cancer Res. 17:5060–5070.
2011. View Article : Google Scholar
|
|
18
|
Fujimoto-Ouchi K, Sekiguchi F, Yasuno H,
Moriya Y, Mori K and Tanaka Y: Antitumor activity of trastuzumab in
combination with chemotherapy in human gastric cancer xenograft
models. Cancer Chemother Pharmacol. 59:795–805. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Fujimoto-Ouchi K, Sekiguchi F and Tanaka
Y: Antitumor activity of combinations of anti-HER-2 antibody
trastuzumab and oral fluoropyrimidines capecitabine/5′-dFUrd in
human breast cancer models. Cancer Chemother Pharmacol. 49:211–216.
2002.PubMed/NCBI
|
|
20
|
Baselga J, Gelmon KA, Verma S, et al:
Phase II trial of pertuzumab and trastuzumab in patients with human
epidermal growth factor receptor 2-positive metastatic breast
cancer that progressed during prior trastuzumab therapy. J Clin
Oncol. 28:1138–1144. 2010. View Article : Google Scholar
|
|
21
|
Hurvitz SA, Dirix L, Kocsis J, Gianni L,
Lu MJ, Vinholes J, Song C, Tong B, Chu Y-W and Perez EA:
Trastuzumab emtansine (T-DM1) vs trastuzumab plus docetaxel (H+T)
in previously untreated HER2-positive metastatic breast cancer
(MBC): primary results of a randomized, multicenter, open-label
phase II study (TDM4450g/BO21976). Eur J Cancer. 47:S3302011.
View Article : Google Scholar
|
|
22
|
Baselga J and Swain SM: Novel anticancer
targets: revisiting ERBB2 and discovering ERBB3. Nat Rev Cancer.
9:463–475. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
23
|
Olayioye MA, Neve RM, Lane HA and Hynes
NE: The ErbB signaling network: receptor heterodimerization in
development and cancer. EMBO J. 19:3159–3167. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Spector NL and Blackwell KL: Understanding
the mechanisms behind trastuzumab therapy for human epidermal
growth factor receptor 2-positive breast cancer. J Clin Oncol.
27:5838–5847. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Scheuer W, Friess T, Burtscher H,
Bossenmaier B, Endl J and Hasmann M: Strongly enhanced antitumor
activity of trastuzumab and pertuzumab combination treatment on
HER2-positive human xenograft tumor models. Cancer Res.
69:9330–9336. 2009. View Article : Google Scholar
|
|
26
|
El-Sahwi K, Bellone S, Cocco E, et al: In
vitro activity of pertuzumab in combination with trastuzumab in
uterine serous papillary adenocarcinoma. Br J Cancer. 102:134–143.
2010. View Article : Google Scholar : PubMed/NCBI
|