Introduction
Atherosclerotic lesions are characterized by the
formation and accumulation of foam cells in the arterial intimal
layer (1). Generally, foam cell
formation is primarily a result of uncontrolled uptake of modified
low-density lipoprotein (LDL) into the subendothelial space of
macrophages (2,3). In macrophages, scavenger receptors
such as class A scavenger receptor (SR-A) and cluster of
differentiation 36 (CD36) are reported to mediate the
internalization of oxidized LDL (oxLDL) that promotes the cellular
accumulation of cholesterol (4).
Conversely, reverse cholesterol transporters (RCTs) including
SR-BI, ATP-binding cassette transporter A1 and G1 (ABCA1 and ABCG1)
are responsible for cholesterol efflux (5). Although it is known that the
expression of SRs and RCTs can be affected by various atherogenic
factors, such as insulin and endothelin-1 (4,6), their
dysregulation in the setting of atherosclerosis has not yet been
fully investigated.
Porphyromonas gingivalis lipopolysaccharide
(P. gingivalis LPS), a major etiological agent in localized
chronic periodontitis, was shown to attack the tissue enclosing the
teeth, ultimately leading to tooth loss (7). Beside its role in periodontitis,
increasing evidence suggests that P. gingivalis LPS exerts
several atherogenic effects for cardiovascular diseases. For
instance, P. gingivalis LPS triggers the secretion of
inflammatory cytokines and facilitates mononuclear cell adhesion to
vascular endothelium (8,9). Elevated levels of P. gingivalis
LPS appear to present a risk factor for the development of
atherosclerosis (10). Furthermore,
chronic infusion of P. gingivalis LPS increased
atherosclerosis formation in ApoE−/− mice
(11). To this end, the interaction
between P. gingivalis LPS and macrophages regarding their
atherogenic role appears to be reciprocal. On the one hand, many
studies have suggested that LPS isolated from P. gingivalis
activates macrophages to convert into foam cells (12–16).
On the other hand, only two studies have reported the mechanisms of
macrophage-derived foam cell formation stimulated by P.
gingivalis LPS. Lei et al(12) reported that P. gingivalis
LPS-induced foam cell formation may be mediated through the
enhanced activation of the nuclear factor-κB pathway. Moreover,
Kuramitsu et al(17)
suggested that the modification of LDL induced by P.
gingivalis LPS is likely to promote foam cell formation. In
view of the complex nature of cellular lipid level regulation, the
detail mechanisms underlying the effect of P. gingivalis LPS
on foam cell formation remain to be further investigated.
It is well-established that SRs and RCTs dynamically
mediate intracellular cholesterol levels (5). Thus, their expression levels are
important for foam cell formation. The expression of these
mediators may be altered by transcriptional regulation including
factors such as activator protein-1 (AP-1) for CD36 (18,19)
and HO-1 for ABCA1 (20).
Meanwhile, the expression of RCTs may be altered through
post-translational regulation which is related to protein stability
(21,22). To date, the molecular mechanisms by
which P. gingivalis LPS influences the expression of RCTs
and SRs resulting in increased lipid accumulation in foam cells are
still unclear.
The purpose of this study was firstly to examine the
effect of P. gingivalis LPS on cellular lipid levels in
THP-1-derived macrophages; secondly, to investigate the effect of
P. gingivalis LPS on the expression of SRs and RCTs; and
thirdly, to delineate the possible molecular mechanisms involved in
P. gingivalis LPS-induced changes in the expression of RCTs
and SRs as well as foam cell formation.
Materials and methods
Cells and reagents
The THP-1 human monocyte-derived cell line was
obtained from the American Type Culture Collection (ATCC, Manassas,
VA, USA), and maintained in RPMI-1640 medium supplemented with 10%
fetal bovine serum (FBS), 20 IU/ml penicillin and 20 g/ml
streptomycin. Cells were maintained at 37°C in an atmosphere of 5%
CO2-95% room air and differentiated into macrophages by
the addition of phorbol 12-myristate 13-acetate (PMA; 200 ng/ml)
for 48 h. Rabbit anti-c-Fos, rabbit anti-HO-1, rabbit anti-c-Jun
antibodies were from Cell Signaling Technology, Inc. (Beverly, MA,
USA). Mouse anti-ABCA1, rabbit anti-CD36, rabbit anti-SR-BI, rabbit
anti-calpain and rabbit anti-ABCG1 antibodies were obtained from
Abcam (Cambridge, MA, USA). Rabbit anti-calpastatin and goat
anti-SR-A antibody were purchased from Santa Cruz Biotechnology,
Inc. (Santa Cruz, CA, USA). Scrambled and HO-1 small hairpin (sh)
RNA were from Shanghai GenePharma Co., Ltd. (Shanghai, China).
3-Dodecanoyl-NBD-cholesterol was obtained from Cayman Chemical Co.
(Ann Arbor, MI, USA). PMA was from Sigma (St. Louis, MO, USA).
Cholesterol assay kit was obtained from Nanjing Jiancheng
Bioengineering Institute (Nanjing, China). Assay kit for calpain
activity was supplied by Biovision (Lyon, France).
shRNA transfection
shRNA transfection experiments were conducted as
previously described (19). In
brief, cell transfections were performed with the SuperFect
fragment (Qiagen, Valencia, CA, USA) according to the
manufacturer's instructions using scrambled or HO-1 shRNA in a
6-well plate or 50 ml flask. Cells were incubated for 24 h after
transfection and used for the indicated experiments.
Oil Red O staining
Oil Red O staining was used to observe lipid
accumulation in foam cells derived from macrophages. After the
culture of THP-1-derived macrophages with 50 μg/ml oxLDL in the
presence or absence of P. gingivalis LPS for 24 h, cells
were fixed in 4% paraformaldehyde and stained with Oil Red O and
hematoxylin. After Oil Red O staining, the density of the lipid
content was examined by alcohol extraction. The absorbance at 540
nm was evaluated by a microplate reader (BioTek Instruments, Inc.,
Winooski, VT, USA).
Cholesterol efflux assay
Cholesterol efflux experiments were performed as
previously described (19). After a
24-h treatment with various concentrations of P. gingivalis
LPS, the THP-1-derived macrophages were incubated with the
equilibration of NBD-cholesterol (1 μg/ml) for an additional 6 h in
the presence of P. gingivalis LPS. The
NBD-cholesterol-labeled cells were incubated in RPMI-1640 medium
for 6 h after washing with PBS. A multilabel counter (Perkin-Elmer
Life Sciences, Waltham, MA, USA) was used to detect the
fluorescence-labeled cholesterol released from the cells into the
medium. Cholesterol efflux was calculated as a percentage of
fluorescence in the medium relative to the total amount of
fluorescence.
Quantitative real-time polymerase chain
reaction (qRT-PCR)
Total RNA was isolated using RNAiso Plus and was
converted into complementary DNA by the PrimeScript RT reagent kit
(Perfect Real Time; Takara Bio, Inc., Shiga, Japan). The reaction
of qRT-PCR was performed using the iQ™ SYBR-Green Supermix
(Bio-Rad, Hercules, CA, USA) under the following conditions: 3 min
at 95°C for 1 cycle, 10 sec at 95°C, 30 sec at 60°C for 39 cycles,
and 95°C for 5 sec. The following primers were used: GAPDH forward,
5′-GGTGAAGGTCGGTGTGAACG-3′ and reverse, 5′-CTCGCTCCTGGAAGATGGTG-3′;
SR-BI forward, 5′-ACCCTAACCCAAAGGAGCAT-3′ and reverse,
5′-CACAGCAACGGCAGAACTAC-3′; ABCG1 forward,
5′-GCCTACTACCTGGCAAAGACC-3′ and reverse,
5′-GAACAGCACAAAACGCACAG-3′; ABCA1 forward,
5′-CAATGTCAAGGTGTGGTTCAAT-3′ and reverse, 5′-GC
TGCTGTTTAGTGAGGTTCAA-3′; SR-A forward, 5′-TCCTTGATTTCGTCAGTCCAG-3′
and reverse, 5′-CCTCCTGTTGCTTTGCTGTAG-3′; CD36 forward,
5′-TCGCTTCCACATTTCCTACAT-3′ and reverse,
5′-CCCAGTCTCATTTAGCCACAG-3′.
Western blot analysis
The methods for nuclear extracts and western blot
analysis were described in our previous study (19). THP-1-derived macrophages were
harvested and lysed with 180 μl RIPA lysis buffer (Beyotime,
Jiangsu, China). Proteins from total cell lysates, measured using
the bicinchoninic acid protein assay kit (Biomed Biotech Co., Ltd.,
Beijing, China), were boiled in 5X SDS-sample buffer for 10 min,
separated on 8–12% SDS-polyacrylamide minigels and then transferred
onto nitrocellulose membranes (Amersham, Buckinghamshire, UK).
After blocking with 5% non-fat milk in PBST, the membrane was
incubated with primary antibodies overnight at 4°C. The protein
expression was detected by an enhanced chemiluminescence kit (ECL;
Perkin-Elmer Life Sciences) and densitometric analysis was
performed using the 720 BK/01837 System (Bio-Rad).
Immunoprecipitation
Cell lysates containing equal amounts of protein
(1,000 mg) from THP-1-derived macrophages treated with or without
P. gingivalis LPS for 24 h were incubated with the specific
primary antibody overnight at 4°C, and then with protein
A/G-Sepharose for 2 h. Immune complexes were collected and eluted
in lysis buffer. Eluted protein samples were then boiled in
SDS-PAGE loading buffer for subsequent western blot analysis.
Measurement of calpain activity
Calpain activity was measured as previously
described (20). Briefly, cellular
lysates (100 mg) were mixed with reaction buffer and fluorogenic
substrate Ac-LLY-AFC. The level of released AFC was measured over 1
h at 37°C by fluorometry using 400-nm excitation and 505-nm
emission filters.
Statistical analysis
Data are presented as the means ± SEM and were
analyzed using one-way analysis of variance (ANOVA), and the
Newman-Keuls test was used to determine any significant differences
identified by ANOVA. Differences were considered statistically
significant at P<0.05. All experiments were performed at least 3
times.
Results
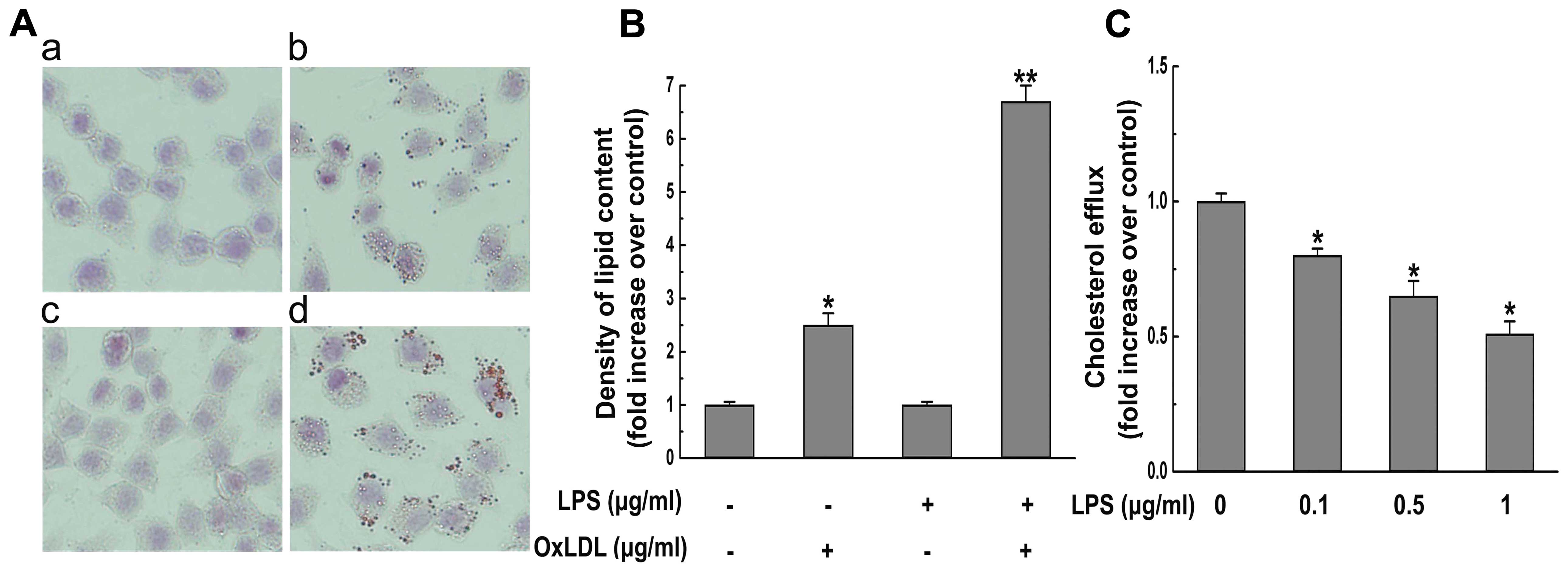
P. gingivalis LPS enhances intracellular
lipid accumulation and decreases cholesterol efflux in
oxLDL-induced macrophages
We investigated the effect of P. gingivalis
LPS on oxLDL-induced foam cell formation. Treatment with oxLDL
caused an increase in lipid accumulation and a decrease in cellular
cholesterol efflux. Notably, the effects of oxLDL on lipid
accumulation and cholesterol efflux were significantly exacerbated
by additional treatment with P. gingivalis LPS (Fig. 1). These results indicate that P.
gingivalis LPS possesses pro-atherogenic activities during the
formation of foam cells.
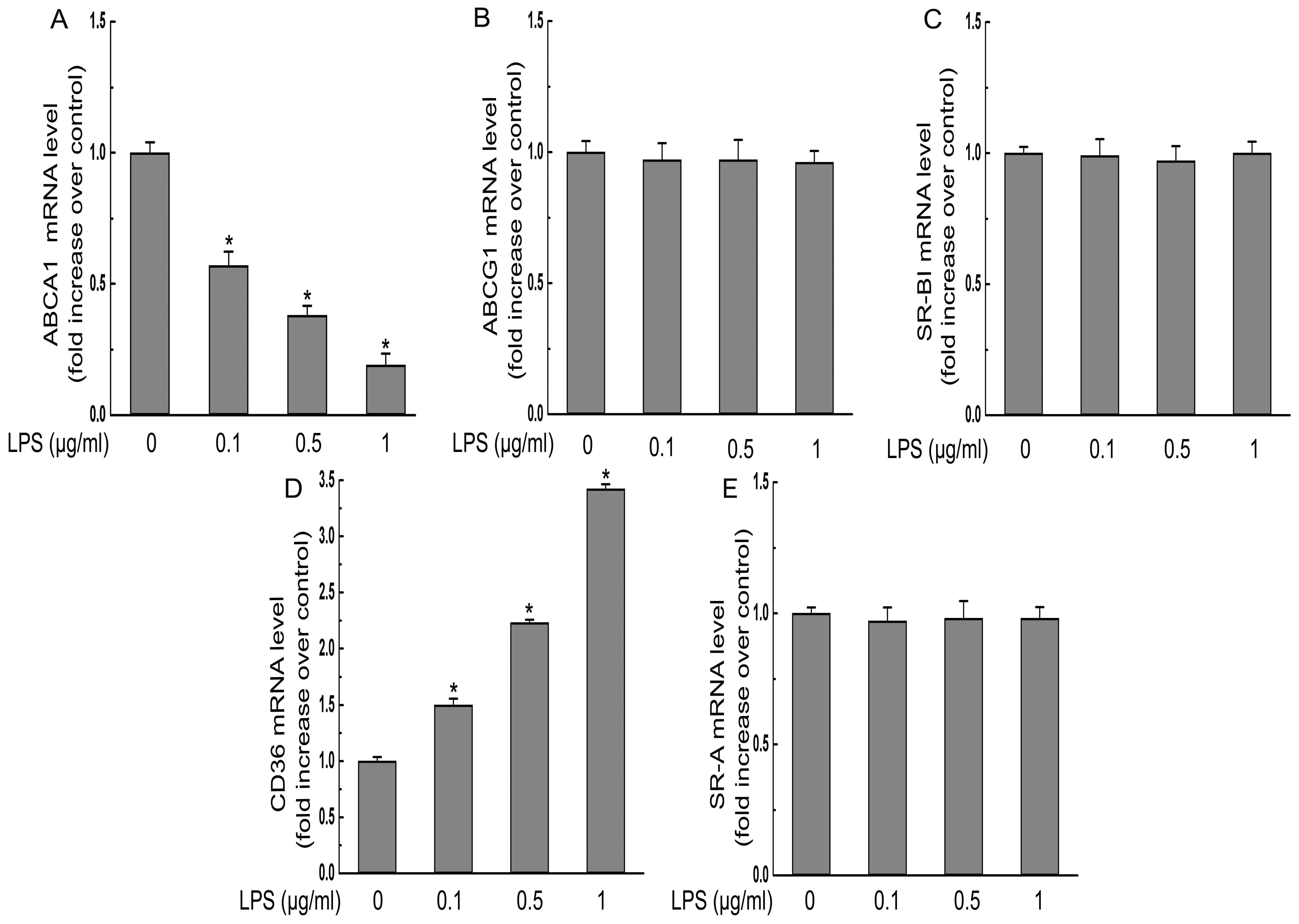
P. gingivalis LPS decreases the
expression of ABCA1, but increases the expression of CD36 in
macrophages
To investigate the mechanisms underlying the
exacerbation of foam cell formation by P. gingivalis LPS,
the effects of P. gingivalis LPS on RCTs and SRs were
examined. Treatment with P. gingivalis LPS at various
concentrations (0.1, 0.5 and 1 μg/ml) for 24 h dose-dependently
decreased the mRNA and protein expression of ABCA1 without
affecting the expression of ABCG1 and SR-BI (Figs. 2 and 3). Moreover, the expression of CD36 was
significantly increased at the protein and mRNA levels in response
to P. gingivalis LPS treatment, while the protein and mRNA
levels of SR-A were not altered by P. gingivalis LPS
incubation (Figs. 2 and 3).
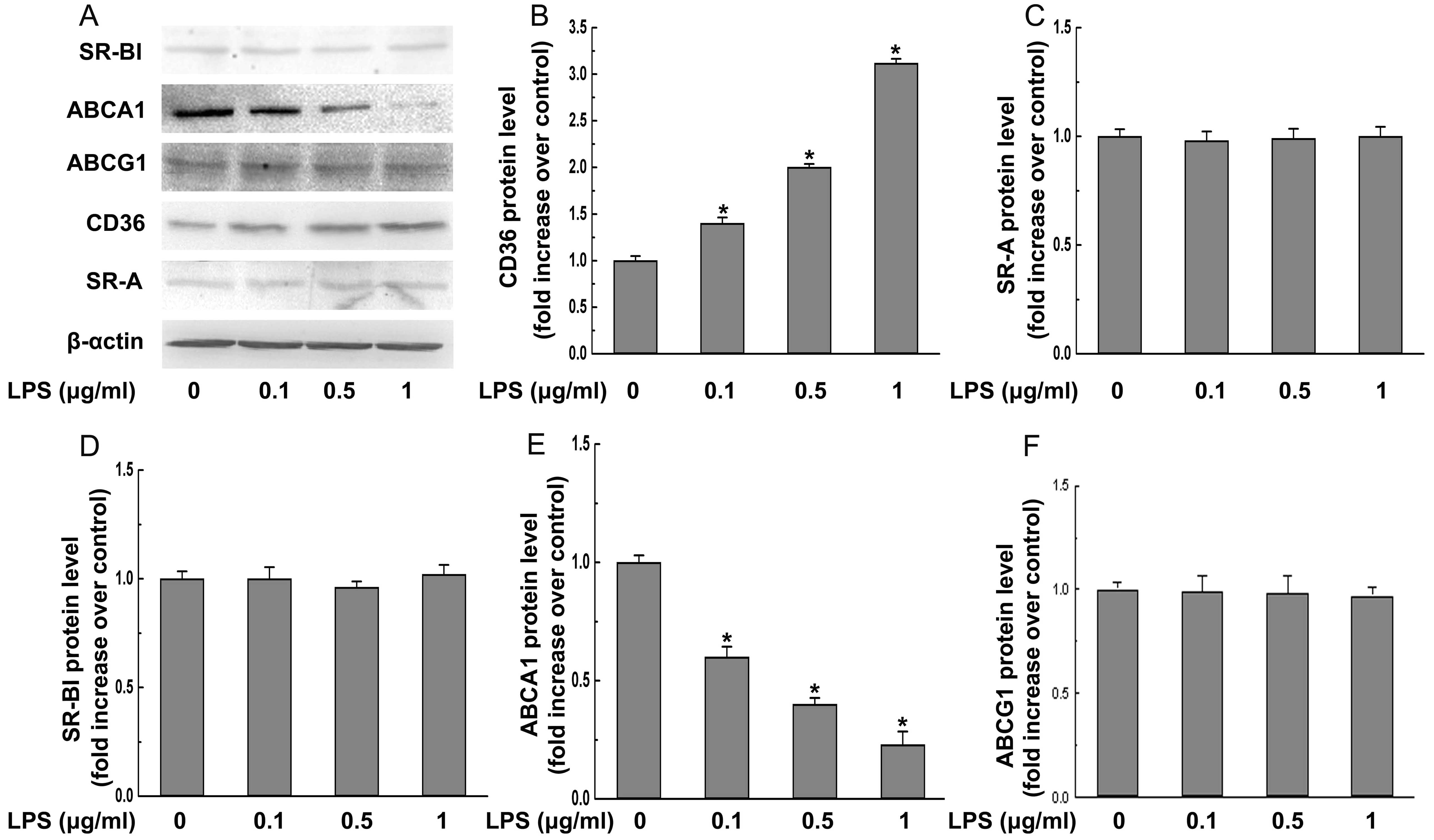 | Figure 2P. gingivalis LPS (LPS)
decreases protein expression of ABCA1, but increases protein
expression of CD36. (A) THP-1-derived macrophages were treated with
the indicated concentrations (0, 0.1, 0.5, 1 μg/ml) of P.
gingivalis LPS for 24 h and the protein levels of SR-A, CD36,
SR-BI, ABCA1, ABCG1, or β-actin were determined by western
blotting. The relative protein levels of (B) CD36, (C) SR-A, (D)
SR-BI, (E) ABCA1, (F) ABCG1 are presented as means ± SEM of the
optical density from 3 separated experiments, *P<0.05
compared with the control. |
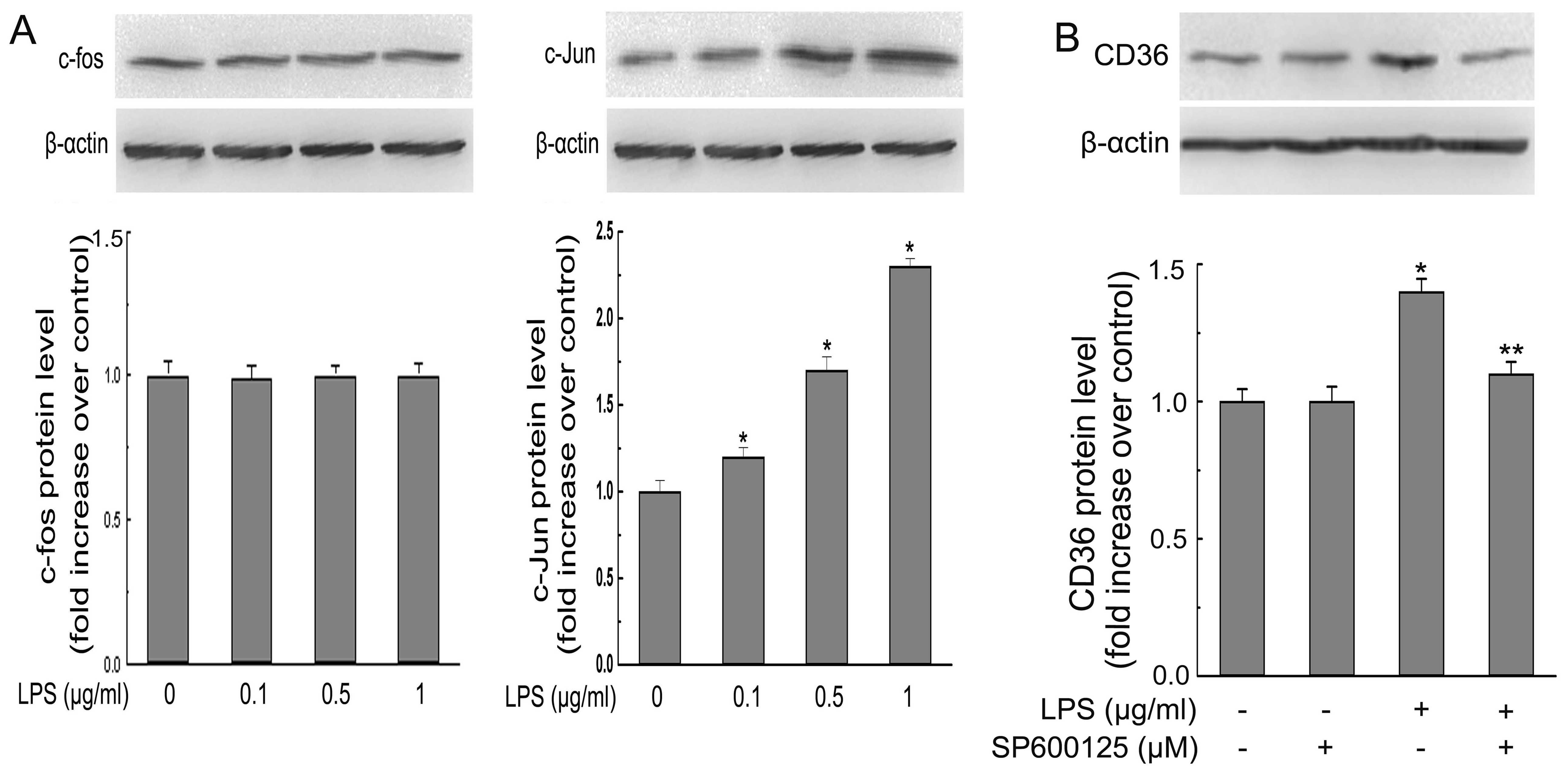
P. gingivalis LPS-induced CD36 expression
is mediated by the c-Jun-AP-1 pathway
It has been reported that AP-1 (c-Jun and c-Fos, 2
key subunits of AP-1) activity contributes to the fate of the cell
after P. gingivalis LPS treatment (23). Thus, we detected the possibility
that P. gingivalis LPS-induced CD36 expression in
macrophages is mediated by the AP-1 pathway. Our results revealed
that P. gingivalis LPS treatment had no effect on the
nuclear expression of c-Fos, while P. gingivalis LPS
treatment of macrophages caused dose-dependent increases in nuclear
c-Jun expression (Fig. 4A).
Additionally, c-Jun N-terminal kinase (JNK) inhibitor, SP600125,
which is a potent, cell-permeable selective and reversible
inhibitor of JNK1, JNK2 and JNK3, blocked P. gingivalis
LPS-induced CD36 expression (Fig.
4B). Collectively, these results suggest that induction of CD36
expression and subsequent exacerbation of foam cell formation by
P. gingivalis LPS are partly c-Jun-AP-1-dependent.
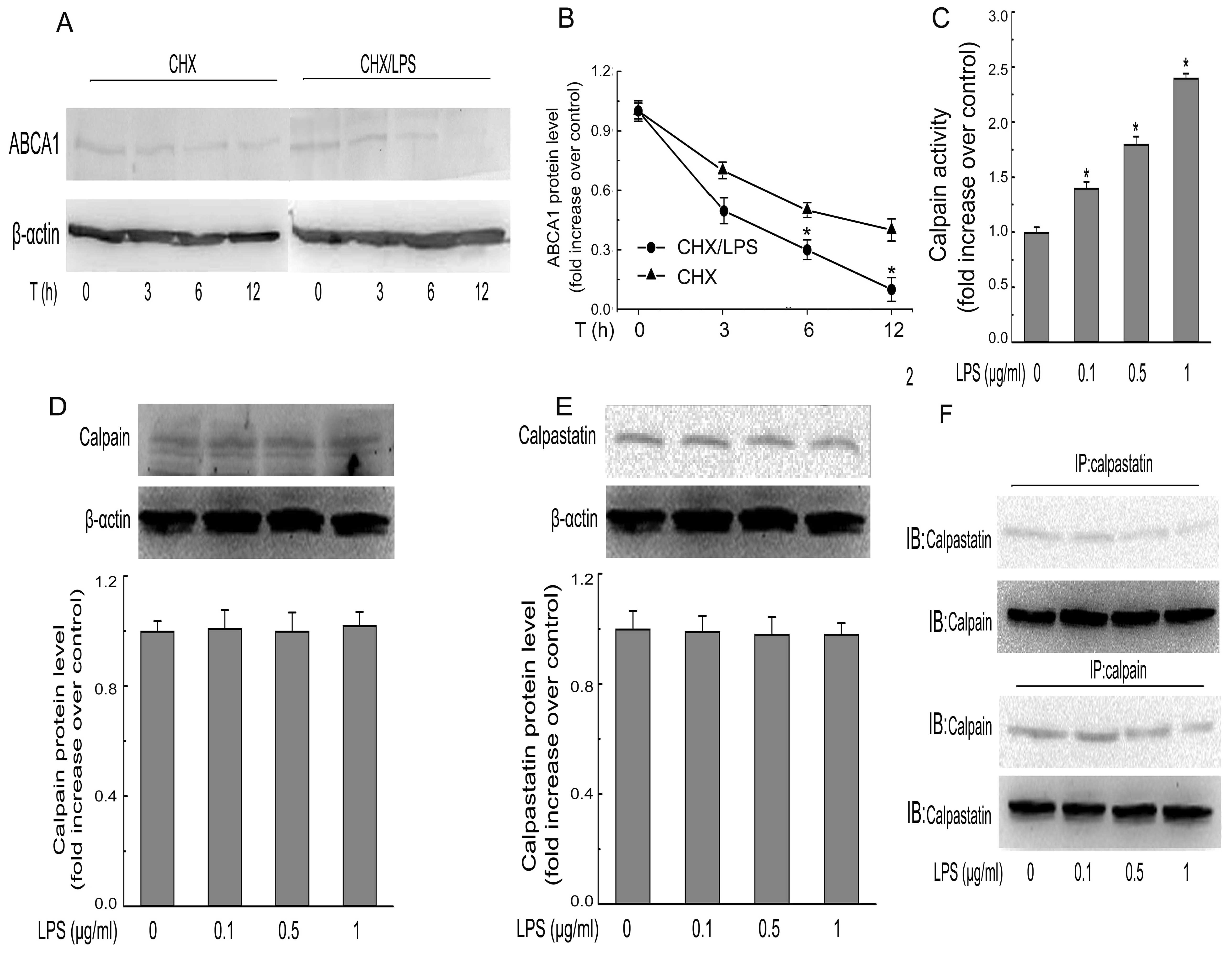
P. gingivalis LPS decreases the stability
of ABCA1 protein by increasing calpain activity
Further analysis of the protein stability of ABCA1
showed that, in the presence of CHX (an inhibitor of de novo
protein synthesis), the degradation profile of ABCA1 protein during
a 12-h treatment with P. gingivalis LPS was more rapid than
in the group without P. gingivalis LPS (Fig. 5A and B). We further investigated the
involvement of calpain, a protease involved in ABCA1 proteolysis
(20). As shown in Fig. 5C, P. gingivalis LPS
incubation enhanced the calpain activity in a dose-dependent
manner. However, the expression of calpain and calpastatin (the
endogenous inhibitor for calpain) were not altered by P.
gingivalis LPS (Fig. 5D and E).
Obviously, the enhanced calpain activity resulted from a decrease
in the protein interaction between calpain and calpastatin
(Fig. 5F).
The enhanced effect of P. gingivalis LPS
on foam cell formation is mediated by HO-1
We determined the role of HO-1 in P.
gingivalis LPS-mediated exacerbation in foam cells. The protein
level of HO-1 in macrophages was dose-dependently decreased in
response to P. gingivalis LPS as measured by western blot
analysis (Fig. 6A). Moreover,
transfection of the HO-1 shRNA for gene knockdown resulted in
promotion of the P. gingivalis LPS-induced effect on HO-1
expression (Fig. 6B), whereas
transfection with the corresponding scrambled shRNA failed to do
so. Additionally, transfection with HO-1 shRNA augmented the P.
gingivalis LPS-mediated effects on the upregulation of c-Jun
(Fig. 6C), and CD36 protein
expression (Fig. 6D),
downregulation of ABCA1 protein expression (Fig. 6E), promotion of calpain activity
(Fig. 6F), and augmentation of
lipid accumulation (Fig. 6G) in
THP-1-derived macrophages, indicating the crucial role of HO-1 in
the exacerbating effects by P. gingivalis LPS.
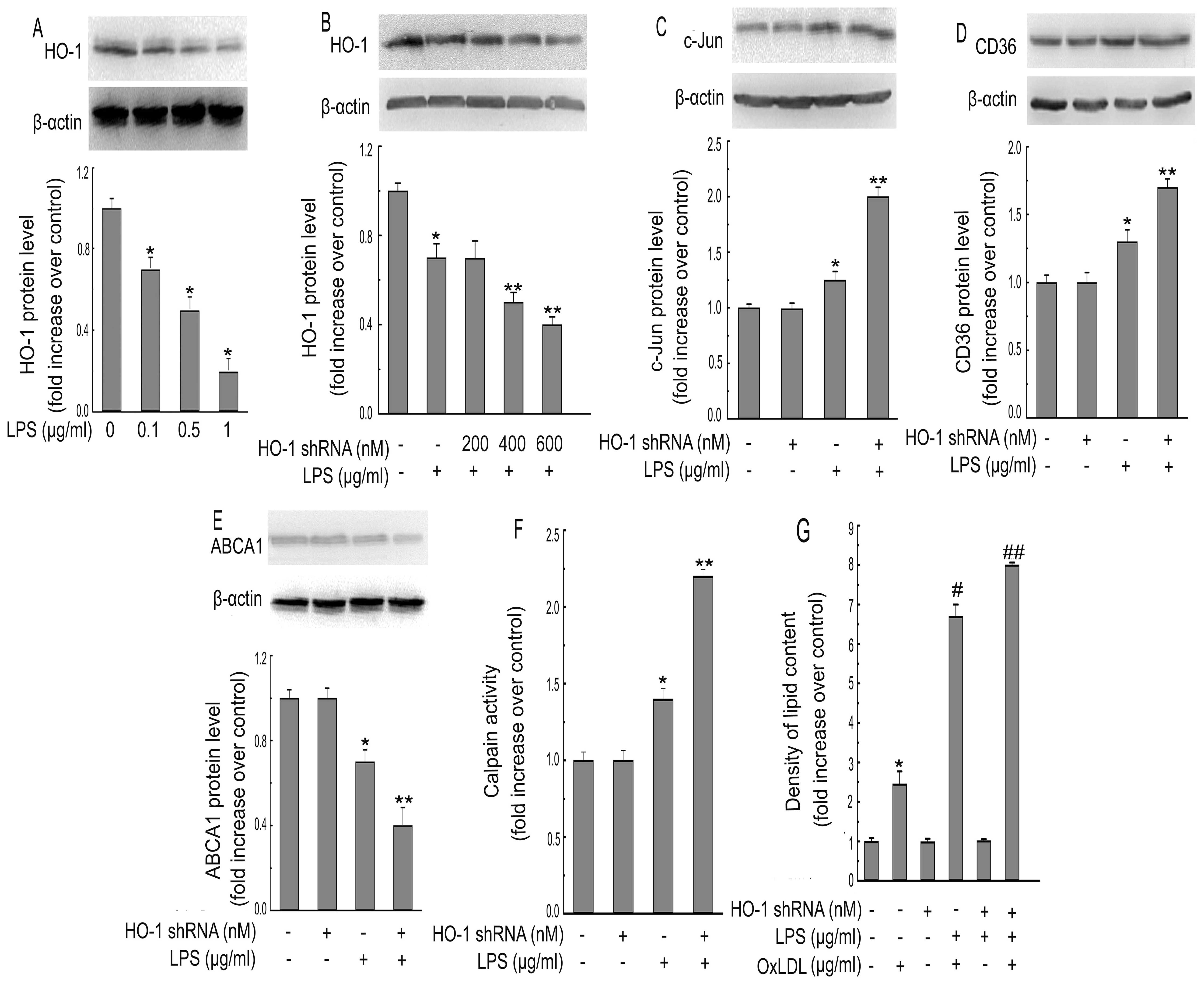 | Figure 6P. gingivalis LPS
(LPS)-induced atherogenic effects in macrophages are mediated by
HO-1. (A) THP-1-derived macrophages were incubated with various
concentrations (0, 0.1, 0.5, 1 μg/ml) of P. gingivalis LPS
for 12 h. Western blotting was used to detect the protein
expression of HO-1 following treatment. (B) THP-1-derived
macrophages were transfected with various concentrations of HO-1
shRNA (200, 400, 600 nM) for 24 h, followed by P. gingivalis
LPS treatment (0.1 μg/ml) for an additional 12 h. Protein
expression of HO-1 and β-actin was measured by western blotting.
(C–E) Macrophages were pre-treated with HO-1 shRNA (600 nM) for 24
h, followed by P. gingivalis LPS for an additional 6 h (C)
or 24 h (D and E). Protein levels of c-Jun, β-actin, CD36 and ABCA1
were determined by western blotting. (F) Calpain activity was
measured by using an enzymatic method. (G) OxLDL-induced lipid
accumulation was measured by alcohol extraction. The data are
representative of 3 independent experiments (means ± SEM).
*P<0.05 compared with the control,
**P<0.05 compared with the P. gingivalis
LPS-treated group. #P<0.05 compared with the
OxLDL-treated group. ##P<0.05 compared with the P.
gingivalis LPS plus OxLDL group. |
Discussion
Emerging data reveal that P. gingivalis LPS
may have an atherogenic effect on promoting intracellular lipid
accumulation during the formation of macrophage-derived foam cells
(12,17). In the present study, our results
indicated that P. gingivalis LPS indeed augmented the uptake
of oxLDL in THP-1-derived macrophages, consistent with previous
studies (12,17). We then used this experimental model
to explore the molecular mechanisms underlying P. gingivalis
LPS-induced promotion of foam cell formation. Our data showed that
P. gingivalis LPS markedly increased the protein and mRNA
expression of CD36, without changing the expression of SR-A. The
importance of CD36 and SR-A in atherogenesis and foam cell
formation is well established (24). The genetic inactivation of CD36 has
previously been shown to reduce oxLDL uptake in vitro and in
atherosclerotic lesions in mice (25), strongly supporting a pro-atherogenic
role for CD36. The expression of CD36 in macrophages is also known
to be upregulated by inflammatory cytokines or stimuli that have an
atherogenic role (26). In view of
CD36 function, it may be concluded that the increase in CD36
expression followed by P. gingivalis LPS treatment may
contribute to the exacerbated lipid accumulation and subsequent
promotion of foam cell formation.
In the present study, we demonstrated for the first
time that c-Jun-AP-1 is the key transcriptional factor for P.
gingivalis LPS-induced upregulation of CD36. This notion is
also supported by the finding that inhibition of AP-1 suppressed
the upregulation of CD36 by P. gingivalis LPS. A recent
study (23) reported that P.
gingivalis LPS, at a similar concentration, activated both
c-Jun and c-Fos, two subunit of AP-1 in RAW 264.7 cells, a mouse
cell line. The discrepancy between this and that study (23) could be due to the differences in
cell types.
In addition to its effects on CD36 expression, our
results further demonstrated that P. gingivalis LPS
decreased the mRNA and protein levels of ABCA1 without affecting
the protein and mRNA expression of ABCG1 or SR-BI. Cholesterol
efflux in macrophages is mainly regulated by ABCA1 (27). The results of the present study
confirmed a recent in vivo study that the ABCA1 mRNA level
in the aorta was significantly reduced in long-term P.
gingivalis-infected mice (28).
The importance of ABCA1 in maintaining cholesterol homeostasis in
macrophages is well established (29). Additionally, the expression of ABCA1
is well known to be downregulated by various inflammatory cytokines
(30). In view of the function of
ABCA1 function, the decreased expression of ABCA1 by P.
gingivalis LPS detected in the present study is also likely to
contribute to foam cell formation.
Furthermore, we provide evidence for the involvement
of calpain protease in P. gingivalis LPS-decreased
post-transcriptional regulation of ABCA1, as revealed by increased
calpain activity and decreased interaction of calpain and
calpastatin, in response to P. gingivalis LPS treatment. In
fact, the critical role of calpain in the stabilization of ABCA1
protein is well established (31,32).
Moreover, protein kinase C (PKC) phosphorylates and stabilizes
ABCA1 through inhibition of its degradation mediated by calpain
(33). The involvement of this
pathway in the P. gingivalis LPS-induced downregulation of
ABCA1 protein degradation warrants further investigation.
More importantly, our results suggest that the
decreased expression of ABCA1, upregulated expression of CD36 and
increased c-Jun/AP-1 nuclear translocation induced by P.
gingivalis LPS is accompanied by decreased HO-1 expression.
This finding was further supported by the results from the western
blot analysis, in which the P. gingivalis LPS-mediated
upregulation of CD36, downregulation of ABCA1, and promotion of
c-Jun/AP-1 nuclear translocation was augmented by the knockdown of
the HO-1 gene using shRNA. In addition, the inhibition of HO-1
exacerbated the P. gingivalis LPS effects on the
upregulation of lipid accumulation and calpain activity in the
macrophages. These results suggest that the decreased expression of
HO-1 is required for the decreased expression of ABCA1, upregulated
expression of CD36 and increased c-Jun/AP-1 nuclear translocation
induced by P. gingivalis LPS. Our findings are in agreement
with reports that deletion of HO-1 leads to the exacerbation of
atherosclerosis and foam cell formation (34). A number of studies have reported
that bilirubin or carbon monoxide regulates the antioxidative or
anti-inflammatory action of HO-1 (35,36).
The involvement of this pathway in the P. gingivalis
LPS-induced promotion of foam cell formation warrants further
investigation.
In summary, our study provides new insight into the
crucial role of HO-1 in the P. gingivalis LPS-mediated
atherogenic effects in macrophages, which exacerbates lipid
accumulation in oxLDL-induced foam cells by a decrease in
cholesterol efflux. The cholesterol efflux regulated by P.
gingivalis LPS is via transcriptional upregulation of CD36
expression and transcriptional and post-transcriptional
downregulation of ABCA1 expression. The results of the present
study provide a potential mechanism for the contribution of P.
gingivalis LPS to atherogenesis and shed light on the
underlying mechanism by which periodontitis affects atherosclerosis
and subsequent coronary heart disease in humans.
Acknowledgements
This investigation was supported by a grant from the
Public Health Bureau of Chongqing China (grant no. 2012-2-137).
References
|
1
|
Xue JH, Yuan Z, Wu Y, et al: High glucose
promotes intracellular lipid accumulation in vascular smooth muscle
cells by impairing cholesterol influx and efflux balance.
Cardiovasc Res. 86:141–150. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Li AC and Glass CK: The macrophage foam
cell as a target for therapeutic intervention. Nat Med.
8:1235–1242. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kleemann R, Zadelaar S and Kooistra T:
Cytokines and atherosclerosis: a comprehensive review of studies in
mice. Cardiovasc Res. 79:360–376. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lin CY, Lee TS, Chen CC, Chang CA, Lin YJ,
Hsu YP and Ho LT: Endothelin-1 exacerbates lipid accumulation by
increasing the protein degradation of the ATP-binding cassette
transporter G1 in macrophages. J Cell Physiol. 226:2198–2205. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Cheng LC, Su KH, Kou YR, Shyue SK, et al:
α-Lipoic acid ameliorates foam cell formation via liver X receptor
α-dependent upregulation of ATP-binding cassette transporters A1
and G1. Free Radic Biol Med. 50:47–54. 2011.
|
|
6
|
Park YM, Kashyap SR, Major J and
Silverstein RL: Insulin promotes macrophage foam cell formation:
potential implications in diabetes-related atherosclerosis. Lab
Invest. 92:1171–1180. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Morimoto Y, Kikuchi K, Ito T, et al: MK615
attenuates Porphyromonas gingivalis
lipopolysaccharide-induced pro-inflammatory cytokine release via
MAPK inactivation in murine macrophage-like RAW264.7 cells. Biochem
Biophys Res Commun. 389:90–94. 2009.
|
|
8
|
Triantafilou M, Gamper FG, Lepper PM, et
al: Lipopolysaccharides from atherosclerosis-associated bacteria
antagonize TLR4, induce formation of TLR2/1/CD36 complexes in lipid
rafts and trigger TLR2-induced inflammatory responses in human
vascular endothelial cells. Cell Microbiol. 9:2030–2039. 2007.
View Article : Google Scholar
|
|
9
|
Nakamura N, Yoshida M, Umeda M, et al:
Extended exposure of lipopolysaccharide fraction from
Porphyromonas gingivalis facilitates mononuclear cell
adhesion to vascular endothelium via Toll-like receptor-2 dependent
mechanism. Atherosclerosis. 196:59–67. 2008.PubMed/NCBI
|
|
10
|
Kiechl S, Egger G, Mayr M, et al: Chronic
infections and the risk of carotid atherosclerosis: prospective
results from a large population study. Circulation. 103:1064–1070.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Gitlin JM and Loftin CD: Cyclooxygenase-2
inhibition increases lipopolysaccharide-induced atherosclerosis in
mice. Cardiovasc Res. 81:400–407. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lei L, Li H, Yan F, Li Y and Xiao Y:
Porphyromonas gingivalis lipopolysaccharide alters
atherosclerotic-related gene expression in oxidized
low-density-lipoprotein-induced macrophages and foam cells. J
Periodontal Res. 46:427–437. 2011. View Article : Google Scholar
|
|
13
|
Pussinen PJ and Mattila K: Periodontal
infections and atherosclerosis: mere associations? Curr Opin
Lipidol. 15:583–588. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Qi M, Miyakawa H and Kuramitsu HK:
Porphyromonas gingivalis induces murine macrophage foam cell
formation. Microb Pathog. 35:259–267. 2003. View Article : Google Scholar
|
|
15
|
Kuramitsu HK, Kang IC and Qi M:
Interactions of Porphyromonas gingivalis with host cells:
implications for cardiovascular diseases. J Periodontol. 74:85–89.
2003.
|
|
16
|
Kuramitsu HK, Qi M, Kang IC and Chen W:
Role for periodontal bacteria in cardiovascular diseases. Ann
Periodontol. 6:41–47. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kuramitsu HK, Miyakawa H, Qi M and Kang
IC: Cellular responses to oral pathogens. Ann Periodontol. 7:90–94.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Katayama I, Hotokezaka Y, Matsuyama T,
Sumi T and Nakamura T: Ionizing radiation induces macrophage foam
cell formation and aggregation through JNK-dependent activation of
CD36 scavenger receptors. Int J Radiat Oncol Biol Phys. 70:835–846.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li XY, Kong LX, Li J, He HX and Zhou YD:
Kaempferol suppresses lipid accumulation in macrophages through the
downregulation of cluster of differentiation 36 and the
upregulation of scavenger receptor class B type I and ATP-binding
cassette transporters A1 and G1. Int J Mol Med. 31:331–338.
2013.
|
|
20
|
Tsai JY, Su KH, Shyue SK, et al: EGb761
ameliorates the formation of foam cells by regulating the
expression of SR-A and ABCA1: role of haem oxygenase-1. Cardiovasc
Res. 88:415–423. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Feng B and Tabas I: ABCA1-mediated
cholesterol efflux is defective in free cholesterol-loaded
macrophages. Mechanism involves enhanced ABCA1 degradation in a
process requiring full NPC1 activity. J Biol Chem. 277:43271–43280.
2002. View Article : Google Scholar
|
|
22
|
Wang Y and Oram JF: Unsaturated fatty
acids inhibit cholesterol efflux from macrophages by increasing
degradation of ATP-binding cassette transporter A1. J Biol Chem.
277:5692–5697. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kim HS, Park SY, Kim EK, Ryu EY, Kim YH,
Park G and Lee SJ: Acanthopanax senticosus has a heme
oxygenase-1 signaling-dependent effect on Porphyromonas
gingivalis lipopolysaccharide-stimulated macrophages. J
Ethnopharmacol. 142:819–828. 2012. View Article : Google Scholar
|
|
24
|
de Winther MP and Hofker MH: Scavenging
new insights into atherogenesis. J Clin Invest. 105:1039–1041.
2000.PubMed/NCBI
|
|
25
|
Kunjathoor VV, Febbraio M, Podrez EA, et
al: Scavenger receptors class A-I/II and CD36 are the principal
receptors responsible for the uptake of modified low density
lipoprotein leading to lipid loading in macrophages. J Biol Chem.
277:49982–49988. 2002. View Article : Google Scholar
|
|
26
|
Ortiz-Masià D, Díez I, Calatayud S, et al:
Induction of CD36 and thrombospondin-1 in macrophages by
hypoxia-inducible factor 1 and its relevance in the inflammatory
process. PLoS One. 7:e485352012.PubMed/NCBI
|
|
27
|
Brooks-Wilson A, Marcil M, Clee SM, et al:
Mutations in ABC1 in Tangier disease and familial high-density
lipoprotein deficiency. Nat Genet. 22:336–345. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Maekawa T, Takahashi N, Tabeta K, et al:
Chronic oral infection with Porphyromonas gingivalis
accelerates atheroma formation by shifting the lipid profile. PLoS
One. 6:e202402011.
|
|
29
|
Oram JF and Heinecke JW: ATP-binding
cassette transporter A1: a cell cholesterol exporter that protects
against cardiovascular disease. Physiol Rev. 85:1343–1372. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yin K, Liao DF and Tang CK: ATP-binding
membrane cassette transporter A1 (ABCA1): a possible link between
inflammation and reverse cholesterol transport. Mol Med.
16:438–449. 2010.PubMed/NCBI
|
|
31
|
Martinez LO, Agerholm-Larsen B, Wang N,
Chen W and Tall AR: Phosphorylation of a pest sequence in ABCA1
promotes calpain degradation and is reversed by ApoA-I. J Biol
Chem. 278:37368–37374. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Arakawa R and Yokoyama S: Helical
apolipoproteins stabilize ATP-binding cassette transporter A1 by
protecting it from thiol protease-mediated degradation. J Biol
Chem. 277:22426–22429. 2002. View Article : Google Scholar
|
|
33
|
Liu XY, Lu Q, Ouyang XP, et al: Apelin-13
increases expression of ATP-binding cassette transporter A1 via
activating protein kinase C α signaling in THP-1 macrophage-derived
foam cells. Atherosclerosis. 226:398–407. 2013.PubMed/NCBI
|
|
34
|
Orozco LD, Kapturczak MH, Barajas B, et
al: Heme oxygenase-1 expression in macrophages plays a beneficial
role in atherosclerosis. Circ Res. 100:1703–1711. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Idriss NK, Blann AD and Lip GY:
Hemoxygenase-1 in cardiovascular disease. J Am Coll Cardiol.
52:971–978. 2008. View Article : Google Scholar
|
|
36
|
Ishikawa K, Sugawara D, Wang Xp, Suzuki K,
Itabe H, Maruyama Y and Lusis AJ: Heme oxygenase-1 inhibits
atherosclerotic lesion formation in ldl-receptor knockout mice.
Circ Res. 88:506–512. 2001. View Article : Google Scholar : PubMed/NCBI
|