Introduction
Metastasis is the spread of cancer in the body, by
which cancer cells in the primary tumor travel to distant sites,
and form metastatic colonies. Most cancer deaths are caused by the
growth of metastases to form clinically harmful tumors, rather than
primary tumor growth. Since current cancer therapies are
inefficient in eradicating metastases, any strategy that can
prevent metastatic progression or inhibits the growth of metastases
in distant sites benefit patients (1). An agonistic monoclonal antibody (DR5
mAb) targeting a receptor of endogenous death ligand (DR5 TRAIL
receptor) can reduce the number of metastatic colonies in mouse
models (2). However, the effect of
combining DR5 mAb with other antimetastatic natural products has
not been investigated.
A unique feature of solid tumors is the occurrence
of low oxygen levels (hypoxia) in some regions within the tumor as
it grows. This has received much attention as an attractive target
for anticancer drug development (3). Clinical studies indicate correlation
between tumor hypoxia, tumor progression and development of distant
metastases (4,5). The presence of hypoxia is not
restricted to the primary tumor, but also appears in metastatic
sites. Increasing evidence obtained from various animal models
indicates avascular micrometastases (<1 mm in diameter) are
hypoxic, while macrometastases (1–4 mm in diameter) are
vascularized and not hypoxic (6–8).
Tumor-induced angiogenesis is known to contribute to metastasis
promotion and growth of micrometastases (9). Vascular endothelial growth factor
(VEGF) is one of the most potent angiogenic factors, and its
production is upregulated in hypoxic cells, further inducing
angiogenesis in hypoxic tissues (10). Hypoxic adaptation is mediated by
hypoxia-inducible factor (HIF)-1 which is a heterodimeric
transcription factor composed of HIF-1α and HIF-1β subunits.
Although HIF-1 is the master regulator of hypoxic response, other
signaling pathways including STAT3, Akt, ERK, and NF-κB, are also
activated by hypoxia and participate in hypoxia-induced VEGF gene
expression (10,11). VEGF also acts as an autocrine
survival factor for breast cancer cells under hypoxia (12). Moreover, inhibition of VEGF gene
expression in breast cancer cells can suppress metastasis in a
mouse model (13).
Propolis (bee glue) is a resinous material that
honeybees collect from various plants, and is a rich source of
bioactive compounds. Several HIF-1 modulating compounds have been
isolated from Brazilian green propolis, indicating the potential of
propolis as a source of hypoxic modulating compounds (3). We have isolated a flavonoid chrysin
(5,7-dihydroxyflavone) from Thai propolis extract as a major
constituent. Chrysin has been shown to possess multiple
bioactivities. Inhibition of subcutaneous tumor growth by chrysin
has recently been demonstrated in mouse models of breast cancer and
hepatoma (14,15). Chrysin has also been shown to
suppress hypoxia-induced VEGF expression in human prostate cancer
cells in vitro(16), and
abrogates tumor-induced angiogenesis in vivo(15,16).
However, until now, there has been no report on the effect of
chrysin on hypoxic survival and metastatic progression of cancer
cells.
In this study, we demonstrate the inhibitory effect
of chrysin on hypoxic survival of mouse breast cancer cells in
vitro, and also investigate its mechanism of action. We also
studied the in vivo effect of chrysin in a spontaneous
metastasis model of breast cancer, both as a single agent and in
combination with DR5 mAb.
Materials and methods
Chemicals and antibodies
Protease inhibitor cocktail for use with mammalian
cells and tissue extracts was obtained from Sigma-Aldrich (St.
Louis, MO, USA). Chrysin was purchased from Alexis Biochemicals
(San Diago, CA, USA). Specific inhibitors of STAT3
(Cucurbitacin-I), PI3K (LY294002), ERK (PD98059), and NF-κB (IKK-2
inhibitor IV) were obtained from Calbiochem (San Diego, CA, USA).
For in vitro experiments, test compounds were dissolved in
dimethyl sulfoxide (DMSO) and kept as a stock solution at −20°C.
The final concentration of DMSO was kept below 0.2% throughout the
study. Primary antibodies specific to Tubulin, STAT3, Akt, ERK1/2,
and p65 NF-κB and phosphorylated forms of STAT3 (Tyr705), Akt
(Ser473), ERK1/2 (Thr202/Tyr204),
p65 NF-κB (Ser536) were purchased from Cell Signaling
Technology (Beverly, MA, USA). Primary antibody specific to HIF-1α
(H1α67) was obtained from Novus Biologicals (Littleton, CO, USA).
Agonistic monoclonal antibody targeting mouse DR5 TRAIL receptor
(DR5 mAb or MD5-1) was obtained from the Department of Immunology,
Juntendo University School of Medicine, Japan.
Propolis extraction and isolation of
compounds
Thai propolis was harvested from a hive of Apis
mellifera collected in Chiangmai province, Thailand, in August
2006. A voucher specimen (SWU 0212) was deposited at the Faculty of
Pharmacy, Srinakharinwirot University, Nakhon Nayok province,
Thailand. Two major compounds, chrysin and tectochrysin, were
isolated. Propolis (1 kg) was extracted with methanol (MeOH) under
sonication yielding 517 g extract. Part of the extract (140 g) was
fractionated by silica gel column chromatography using
dichloromethane (CH2Cl2), with increasing
concentration of MeOH, to obtain 16 fractions. Fraction 5 (3.0 g,
1–1.5% MeOH-CH2Cl2 eluate) and fraction 8
(9.5 g, 2% MeOH-CH2Cl2 eluate) was left
overnight to obtain the crystals of tectochrysin (314 mg) and
chrysin (732 mg), respectively. Their structures were identified by
comparison of physical and spectroscopic data with authentic
standard and the published data (17,18).
Cell culture
4T1 murine mammary carcinoma cell line was obtained
from the American Type Culture Collection (ATCC, Rockville, MD,
USA). Cells were cultured in DMEM supplemented with 2 mM
L-glutamine, 10% fetal bovine serum (FBS) and antibiotics, and
maintained at 37°C in a humidified atmosphere of 5% CO2:
these conditions were used as normoxic condition (21%
O2). For hypoxic condition (1% O2), the cells
were incubated at 37°C, in a humidified atmosphere of 1%
O2 and 5% CO2 maintained by a regulated
supply of N2 and CO2 gas inside a hypoxic
chamber (C-Chamber with ProOX and ProCO2 controllers,
Biospherix, NY, USA).
Hypoxic cell survival assay
Cells were seeded into 96-well plates
(5×103 cells/well) and left overnight in a
CO2 incubator. The cells were pretreated by adding
medium containing test compound to the wells, and incubated for 30
min, then the plates were further incubated under hypoxia (1%
O2) for 48 h. Number of surviving cells after treatment
was determined by crystal violet staining method as previously
described (19). Briefly, the cells
were washed three times with phosphate buffer saline (PBS), fixed
with 95% ethanol, and stained with crystal violet solution (0.5%
w/v in 25% MeOH). After extensive washing by tap water, the stained
cells were lysed with 0.1 N HCl in MeOH, after which absorbance was
determined at 550 nm using a microplate reader. Assay was performed
in triplicate wells, and results were expressed as percent survival
compared with control (DMSO).
Immunoblot analysis
After treatment, cells were rinsed with cold PBS,
scraped and lysed in RIPA lysis buffer containing phosphatase
inhibitors (20 mM β-glycerophosphate, 1 mM
Na3VO4, 10 mM NaF) and protease inhibitor
cocktail. Cell lysates were fractionated by electrophoresis in 7.5%
or 10% SDS-PAGE, and electrophoretically transferred to PVDF
membrane (Immobilon-P nylon, Millipore, Bedford, MA, USA). After
blocking with BlockAce (Dainippon Pharmaceutical, Co. Ltd., Osaka,
Japan), the membranes were probed with the indicated primary
antibodies overnight, followed by horseradish peroxidase-conjugated
secondary antibodies. Bands were visualized using WesternBright ECL
reagent (Advansta, Menlo Park, CA, USA).
Quantitative reverse
transcriptase-polymerase chain reaction (qRT-PCR)
RNA extraction and first-strand cDNA synthesis were
performed as previously described (20). Real-time PCR was run on a
LightCycler® 2.0 Instrument (Roche), using QuantiTect™
SYBR Green PCR Master mix (Qiagen, Valencia, CA, USA). Sequences of
PCR primers were obtained from following references; mouse VEGF
(21); mouse β-actin (22). VEGF primer (fwd)
5′-GAGATGAGCTTCCTACAGCAC-3′; and VEGF primer (rev)
5′-TCACCGCCTTGGCTTGTCAC-3′. β-actin primer (fwd) 5′-AAG TGT GAC GTT
GAC ATC CGT AA-3′; and β-actin primer (rev) 5′-TGC CTG GGT ACA TGG
TGG TA-3′. PCR reaction was started by an initial activation step
at 95°C for 15 min, followed by 40 cycles comprising denaturation
at 94°C/15 sec, annealing at 55°C/30 sec, and extension at 72°C/30
sec. Expression level of the VEGF gene was normalized to β-actin
gene, and calculated from crossing point (Cp) value of the sample,
relative gene expression level =
2(Cpactin−
Cptarget). The relative change
in gene expression compared with untreated normoxic cells was
expressed as fold change, calculated by 2−ΔΔCp
method.
Spontaneous metastasis mouse model
Balb/c mice (6-week-old female) were purchased from
Sankyo Lab Service Co. (Hamamatsu, Japan). Experiments were
conducted in accordance with institutional guidelines and approval
of University of Toyama Animal Experimental Ethics Committee. Cell
suspensions (5×105 cells) were injected into mammary fat
pads on day 0. Tumors were measured every other day (length and
width) using a caliper. Daily oral administration (p.o.) of chrysin
suspended in 0.5% carboxymethyl cellulose began on day 1 after
tumor inoculation. DR5 mAb (10 μg/mouse) was intraperitoneally
(i.p.) injected on day 10 and day 14 after tumor inoculation.
Primary tumors were excised on day 18, weighed and fixed in 10%
formalin. After sacrificing the mice on day 32, lungs were
collected and fixed in Bouin’s solution (0.9% picric acid, 9%
formaldehyde, 5% acetic acid). Number of metastatic colonies on
surface of the lungs was counted. Size of primary tumors and
metastatic colonies was measured using a caliper square along the
longer axes (a) and shorter axes (b), and tumor
volume was calculated by the following formula; tumor volume
(mm3) = ab2/2. The volume of metastatic
colonies was categorized into 3 sizes; small (<0.5
mm3), medium (0.5–1.5 mm3), and large
(>1.5 mm3). Metastatic burden was determined from
total volume of lung metastatic colonies. Data are expressed as
mean ± SD of 10 mice per group.
Statistical analysis
For in vitro experiments, data are expressed
as mean ± SD, and analyzed by Student’s two-tailed t-test to
determine the significance of differences between groups. For in
vivo experiments, the significance of differences between
groups was analyzed by analysis of variance (ANOVA), and the means
were compared by using Tukey, with a p-value <0.05 being
considered significant.
Results
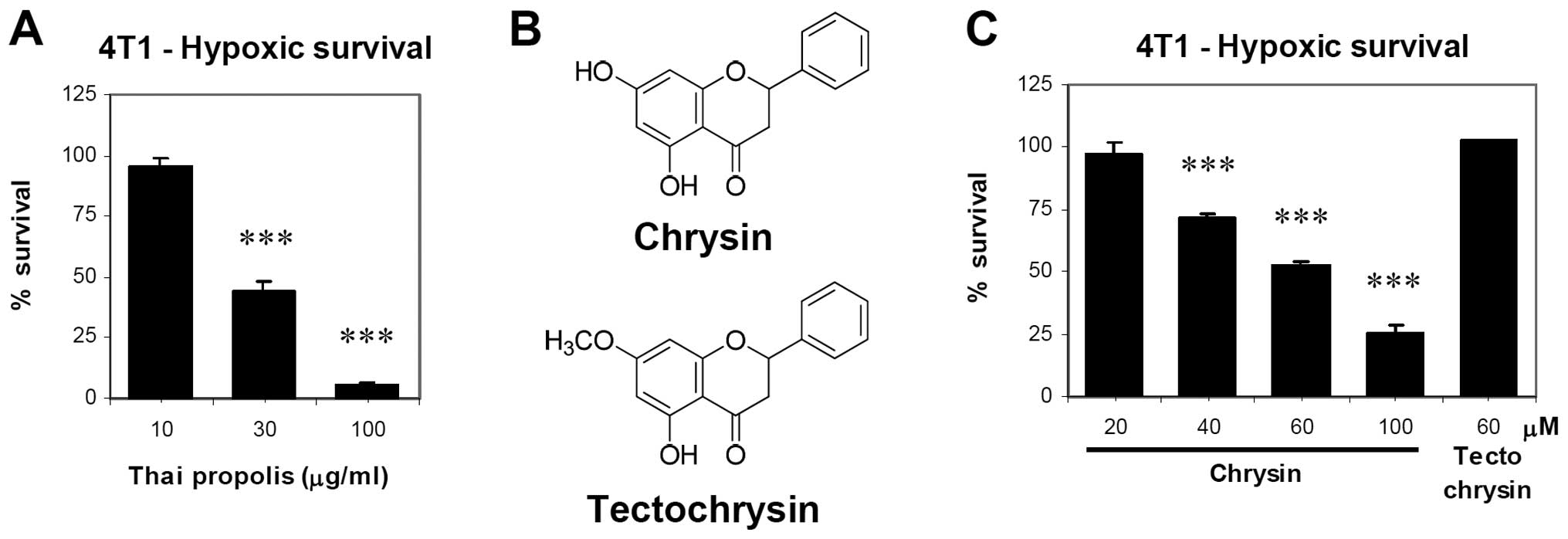
Inhibitory effect of Thai propolis
extract and chrysin on hypoxic survival of breast cancer cells
We first examined the effect of Thai propolis
extract against hypoxic survival of 4T1 cells. Pretreatment with
propolis extract (10–100 μg/ml) for 30 min before exposure to
hypoxia (1% O2) for 2 days decreased hypoxic cell
survival in a concentration-dependent manner (Fig. 1A). Next, the two major constituents
present in Thai propolis extract, chrysin and tectochrysin
(Fig. 1B) were examined. Chrysin
treatment showed a concentration-dependent decrease in hypoxic cell
survival, with 28–74% inhibition observed at 40–100 μM chrysin,
while tectochrysin did not show inhibition at the highest soluble
concentration (60 μM) (Fig. 1C).
These results indicate that chrysin contributes to the inhibitory
effect of Thai propolis extract on hypoxic cell survival.
Effect of chrysin on hypoxia-induced
activation of multiple cell signaling pathways and VEGF
expression
We further explored effect of chrysin on
hypoxia-induced activation of multiple cell signaling pathways that
contribute to cell survival, including HIF-1, STAT3, Akt, ERK, and
NF-κB. 4T1 cells were pretreated with 60–100 μM chrysin for 30 min
before exposure to hypoxia for 6–24 h. HIF-1α protein stabilization
and hypoxia-induced phosphorylation of ERK and p65 NF-κB were not
inhibited by chrysin treatment (Fig.
2A). In contrast, hypoxia-induced STAT3 tyrosine
phosphorylation was markedly inhibited by chrysin, in a dose- and
time-dependent manner, while marginal decrease in Akt
phosphorylation was observed only at the highest concentration test
(100 μM) (Fig. 2A), suggesting that
STAT3 was primary target of chrysin rather than Akt.
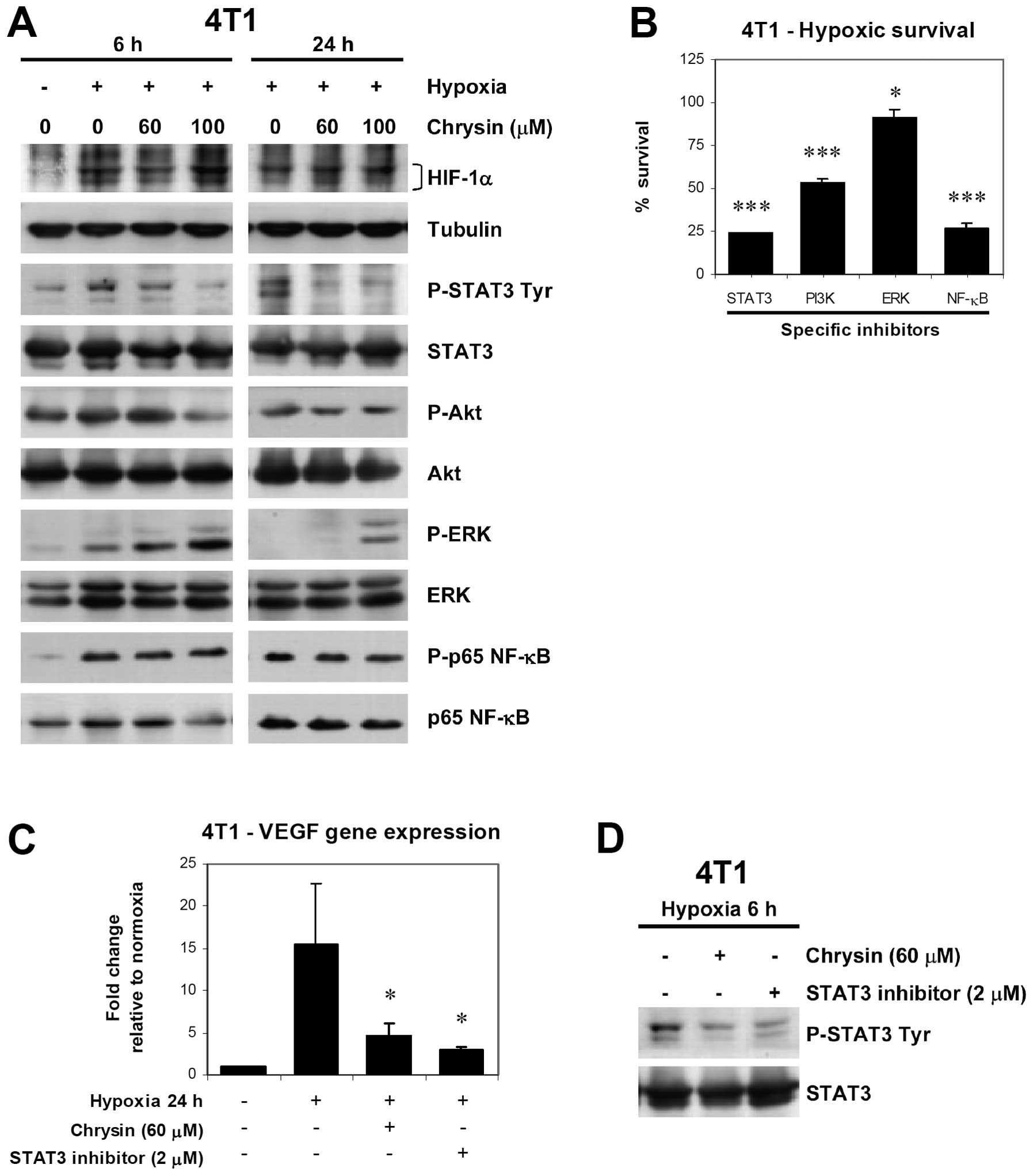 | Figure 2Effect of chrysin on hypoxia-induced
activation of multiple signaling pathways and VEGF gene expression.
(A) Immunoblot analysis of HIF-1α stabilization and phosphorylation
of multiple signaling pathways in 4T1 under hypoxia (1%
O2). Cells were treated with chrysin at indicated
concentrations for 30 min before incubation under hypoxic condition
for 6 or 24 h, after which whole cell lysates were analyzed by
immunoblotting. (B) Effect of specific inhibitors of signaling
pathways, including STAT3 (cucurbitacin-I, 2 μM), PI3K (LY294002,
25 μM), ERK (PD98059, 25 μM), and NF-κB (IKK-2 inhibitor IV, 10
μM), on hypoxic survival of 4T1 cells. The cells were treated with
test inhibitor for 30 min before incubation under hypoxic condition
for 2 days. Number of surviving cells after treatment was
determined by crystal violet staining method. (C) 4T1 cells were
treated with chrysin for 30 min before incubation under hypoxia (1%
O2) for 24 h, VEGF mRNA level was determined by qRT-PCR,
and normalized to β-actin. Change in gene expression compared with
normoxic control is expressed as fold change. Data are expressed as
mean ± SD from three independent experiments, and significant
difference from hypoxic control are shown by *p<0.05,
***p<0.001. (D) Inmmunoblot analysis of STAT3
phosphorylation confirms inhibitory effect of chrysin and STAT3
inhibitor on hypoxia-induced STAT3 phosphorylation in 4T1 cells.
Cells were treated with test compound for 30 min before incubation
under hypoxic condition for 6 h, after which whole cell lysates
were analyzed by immunoblotting. |
Specific inhibitors of STAT3 (cucurbitacin-I, 2 μM),
PI3K (LY294002, 25 μM), ERK (PD98059, 25 μM), and NF-κB (IKK-2
inhibitor IV, 10 μM), were used to evaluate the contribution of the
signaling pathways on hypoxic survival of 4T1 cells. Pretreatment
with inhibitors for 30 min before hypoxic exposure for 2 days
revealed that hypoxic cell survival was inhibited by inhibitors of
STAT3, Akt, and NF-κB pathways, while ERK inhibitor showed marginal
effect (Fig. 2B). These results
indicate the important role of STAT3, Akt, and NF-κB pathways in
hypoxic survival of 4T1 cells. Moreover, STAT3 inhibitor treatment
mimics the effect of chrysin, suggesting that STAT3 inhibition is
the mechanism underlying inhibitory effect of chrysin on hypoxic
survival of 4T1 cells.
VEGF has been shown to be an autocrine survival
factor for breast cancer cells under hypoxia (12). We examined the effect of chrysin on
hypoxia-induced VEGF gene expression in 4T1 cells, in comparison to
a STAT3 inhibitor (cucurbitacin-I). The cells were pretreated with
chrysin (60 μM) or STAT3 inhibitor (2 μM) for 30 min before
exposure to hypoxia for 24 h. qRT-PCR analysis revealed that
hypoxia increased VEGF mRNA levels in 4T1 cells by 16-fold higher
than normoxic cells (Fig. 2C).
Chrysin decreased VEGF mRNA levels in hypoxic 4T1 cells, similar to
that observed with STAT3 inhibitor treatment, by 71 and 81%
inhibition, respectively, compared with control hypoxic cells
(Fig. 2C). The reduction of VEGF
expression by chrysin and STAT3 inhibitor was correlated with
decrease in STAT3 tyrosine phosphorylation levels under hypoxia
(Fig. 2D). Taken together, the
results indicate that STAT3 inhibition is likely to be a mechanism
of action of chrysin for decreasing hypoxia-induced VEGF expression
in 4T1 cells, and this might contribute to the effect of chrysin on
hypoxic survival.
Chrysin suppresses growth of lung
metastatic colonies in vivo
Since hypoxia and angiogenesis contribute to tumor
progression and metastatic formation (7), chrysin was further studied in 4T1
spontaneous metastasis model. The experimental treatment protocol
(Fig. 3A) did not affect body
weight of the mice (data not shown). The effect of treatments was
evaluated from both primary tumor growth and metastatic burden.
Chrysin (100 and 250 mg/kg daily) had no significant effect on
primary tumor growth (Fig. 3B and
C). However, metastatic burden was largely decreased by
chrysin, in a dose-dependent manner, and significant suppression
was observed with the highest dose of chrysin treatment (Fig. 3D). Reduction of metastatic burden
might result from either decrease in metastatic rate (number of
metastatic colonies) or metastatic growth (size of metastatic
colonies), or both. Chrysin tended to decrease metastatic rate but
without statistical significance (Fig.
3E). Suppressive effect of chrysin on metastatic growth was
observed by decreased number of medium and large size metastatic
colonies, in a dose-dependent manner, and significant reduction was
clearly observed with large size metastatic colonies (Fig. 3F). These results reveal the
antimetastatic activity of chrysin in vivo by inhibiting the
growth of metastatic colonies.
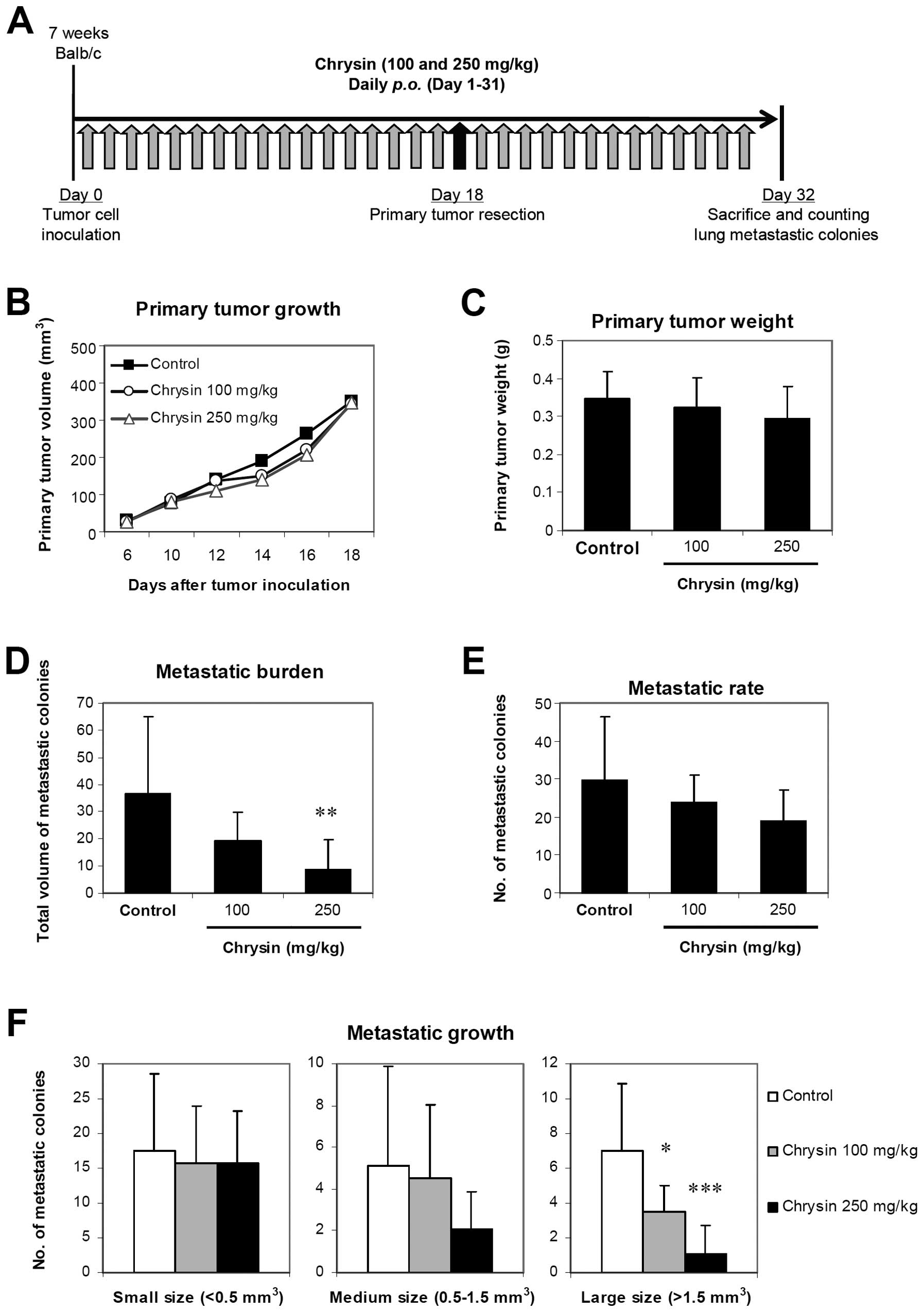 | Figure 3Antimetastatic effect of chrysin
in vivo. (A) Experimental treatment protocol for 4T1
spontaneous lung metastasis. 4T1 cells were inoculated into mammary
fat pad of Balb/c mice. Chrysin (100 and 250 mg/kg) was orally
administered daily until day 31. Primary tumors were resected on
day 18, and mice were sacrificed on day 32. (B and C) Effect of
chrysin on primary tumor growth and weight. Size of primary tumors
was periodically measured and weight was determined after tumor
resection. (D–F) Effect of chrysin on metastatic burden (total
volume of metastatic colonies), metastatic rate (total number of
metastases), and metastatic growth (size of metastases). Lung
metastatic colonies were counted and categorized, according to
their volume, into small (<0.5 mm3), medium (0.5–1.5
mm3), and large (>1.5 mm3). Data are
expressed as mean ± SD (n = 10), and significant differences from
control are shown by *p<0.05, **p<0.01,
***p<0.001. |
Chrysin potentiates antimetastatic
activity of the DR5 agonistic mAb
In addition to therapeutic effect on primary tumor,
DR5 mAb also expresses antimetastatic activity in mouse models
(2). We evaluated the potential of
chrysin as an adjuvant for antimetastatic activity of DR5 mAb.
Since 4T1 cells are sensitive to apoptosis induced by DR5 mAb, low
dose treatment of DR5 mAb (10 μg/mouse, ×2) was used instead of
therapeutic doses (200 μg/mouse, ×3) used previously (2), in order to avoid metastasis reduction
by massive elimination. DR5 mAb alone significantly suppressed
primary tumor growth as shown by reduction in primary tumor size
and weight, compared with control (Fig.
4B and C). Combination of DR5 mAb + chrysin (250 mg/kg/d)
tended to be more suppressive, but no statistical difference
between DR5 mAb alone and the combination treatment was observed
(Fig. 4C). Antimetastatic activity
of DR5 mAb was revealed by significant decrease in metastatic
burden and metastatic rate, and the combination treatment resulted
in a significantly greater effect (Fig.
4D and E). DR5 mAb also expressed suppressive effect on
metastatic growth, as shown by significant decrease in number of
large size metastatic colonies, with greater effect being observed
in combination treatment (Fig. 4F).
Moreover, the effect of DR5 mAb on the number of small size
metastatic colonies was significantly potentiated when DR5 mAb was
combined with chrysin (Fig. 4F).
Collectively, the results suggested that chrysin per se had
a suppressive effect on metastatic growth, and could enhance
antimetastatic activity of DR5 mAb in combination treatment.
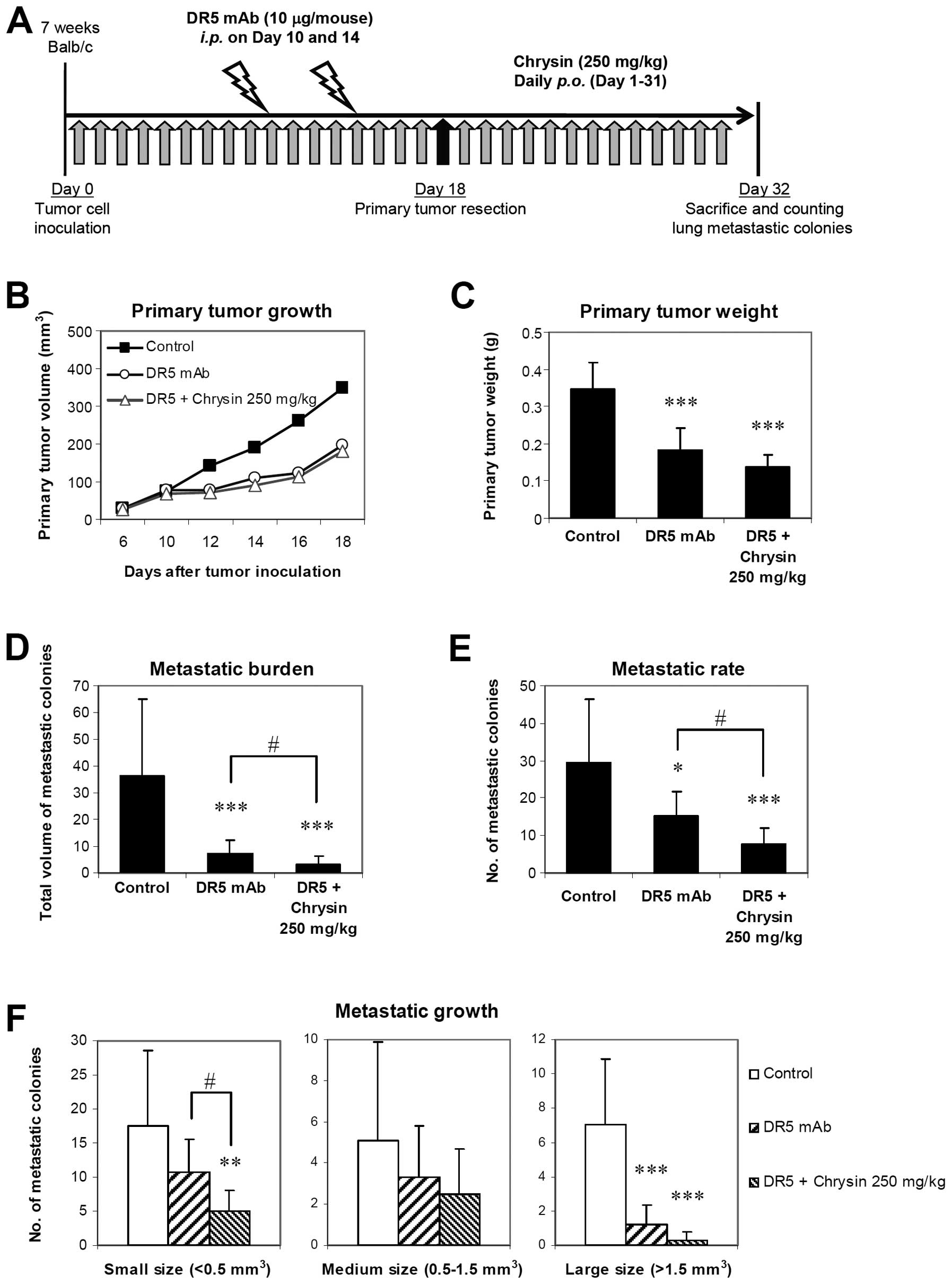 | Figure 4Effect of chrysin and DR5 agonistic
mAb combination treatment in vivo. (A) Experimental
treatment protocol for 4T1 spontaneous lung metastasis. 4T1 cells
were inoculated into mammary fat pad of Balb/c mice. Chrysin (250
mg/kg) was orally administered daily until day 31. DR5 agonistic
mAb (10 μg/mouse) was i.p. administered on day 10 and 14. Primary
tumors were resected on day 18, and mice were sacrificed on day 32.
(B and C) Effect of combination treatment on primary tumor growth
and weight. Size of primary tumors was periodically measured and
weight was determined after tumor resection. (D–F) Effect of
combination treatment on metastatic burden (total volume of
metastatic colonies), metastatic rate (total number of metastatic
colonies), and metastatic growth (size of metastatic colonies).
Lung metastatic colonies were counted and categorized, according to
their volume, into small (<0.5 mm3), medium (0.5–1.5
mm3), and large (>1.5 mm3). Data are
expressed as mean ± SD (n = 10), and significant differences from
control are shown by *p<0.05,
**p<0.01), ***p<0.001. Significant
differences between DR5 mAb alone and DR5 + chrysin are shown by
#p<0.05. |
Discussion
In the present study, we demonstrate the ability of
chrysin to decrease hypoxic survival of cancer cells, by inhibiting
hypoxia-induced STAT3 activation and VEGF expression. We also
provide in vivo evidence of the suppressive effect of
chrysin on metastatic growth, and a potentiating effect of chrysin
when combined with another antimetastatic agent, DR5 mAb. To our
knowledge, this is the first study on the antimetastatic effect of
chrysin in a spontaneous metastasis mouse model.
Increased VEGF production has been shown to play a
role in hypoxic survival of 4T1 cells, and treatment with VEGF
neutralizing antibody can decrease hypoxic survival rate of cells
(12). Consistent with previous
studies, our results showed that treatment with either chrysin or
STAT3 inhibitor (cucurbitacin-I), which inhibited hypoxia-induced
VEGF expression, could decrease hypoxic survival of 4T1 cells.
In hypoxic cells, tyrosine phosphorylated STAT3
forms a complex with HIF-1α at the VEGF promoter, and this complex
is important for upregulation of VEGF gene expression in hypoxic
cells (11). Our results clearly
show that chrysin, as well as a STAT3 inhibitor, could abrogate
hypoxia-induced VEGF gene expression of 4T1 cells, through
inhibition of STAT3 tyrosine phosphorylation, without decreasing
HIF-1α protein levels. Chrysin has been shown to decrease HIF-1α
protein levels in hypoxic DU145 human prostate cancer cells, thus
abrogating insulin-induced HIF-1α protein stabilization in normoxic
DU145 cells, via inhibition of Akt phosphorylation (16).
In a previous study, STAT3 signaling was not
investigated, and the inhibitory effect of chrysin on Akt-modulated
HIF-1α protein stability was shown only in the insulin-stimulated
normoxic cells, but the mechanism was not tested in hypoxic cells
(16). In 4T1 cells, the inhibition
of hypoxia-induced VEGF expression and STAT3 activation by chrysin
were observed at a concentration lower than that affecting Akt
phosphorylation (Fig. 2A),
suggesting that STAT3 was the primary target rather than Akt in our
model. Additionally, a recent study shows that chrysin inhibits
STAT3-mediated VEGF expression, which contributes to suppression of
IL-6-induced angiogenesis by chrysin in human endothelial cells, as
well as in ovo model (23).
Collectively, evidence indicates that chrysin possesses ability to
inhibit angiogenesis, at least partly, through inhibiting
STAT3-mediated VEGF expression.
In our metastasis mouse model, daily oral
administration of chrysin predominantly inhibited metastatic
growth, rather than primary tumor growth. Chrysin has a low oral
bioavailability in humans and rats, due to extensive metabolism to
chrysin sulphate and chrysin glucuronide, and efflux into intestine
(24). Thus, the concentration of
chrysin within primary tumors might be low, especially during early
treatment time, when the primary tumor is growing. Another low oral
bioavailability flavonoid, genistein, also shows a significant
suppressive effect on lung metastatic burden, while marginally
decreasing primary tumor growth at mammary fat pads, in breast
cancer-harboring mice receiving genistein-supplemented diet
(25), similar to that observed in
our chrysin treatment.
Our results revealed that chrysin significantly
suppressed metastatic growth (size of metastatic colonies), while
rate of metastasis (total number of metastatic colonies) was
marginally decreased, suggesting that chrysin interfered with
growth after the tumor cells reached their metastatic sites. Since
progression of micrometastases to macrometastases requires
angiogenesis (9), blocking
VEGF-mediated angiogenesis may suppress metastatic growth. This is
supported by results showing that treatment with a soluble VEGF
decoy receptor (VEGF-Trap) leads to decrease in the size of
established lung metastases of Wilms tumor cells, without changing
the number of metastases (26). In
another study, liver metastatic growth of colon adenocarcinoma was
shown to be suppressed by rapamycin treatment, which affects both
VEGF production of tumor cells and VEGF-response of endothelial
cells (27). Taken together, we
suggest that the suppression of metastatic growth by chrysin was,
at least in part, due to interruption of angiogenesis in hypoxic
micrometastases, by inhibiting hypoxia-induced STAT3 activation and
VEGF expression.
Control of metastatic growth using hypoxia-selective
chemotherapy as adjuvant appears to be an effective strategy, since
a marked reduction of metastatic burden was found in lungs of KHT
sarcoma-harboring mice receiving post-radiotherapy treatment with a
hypoxia-activated cytotoxin NLCQ-1, when hypoxic micrometastases
were evident in lungs (28). In our
study, combination treatment of chrysin and DR5 mAb was more
effective than each single agent in reducing both metastatic rate
and metastatic growth, suggesting that chrysin might be useful as
an adjuvant in controlling metastatic progression.
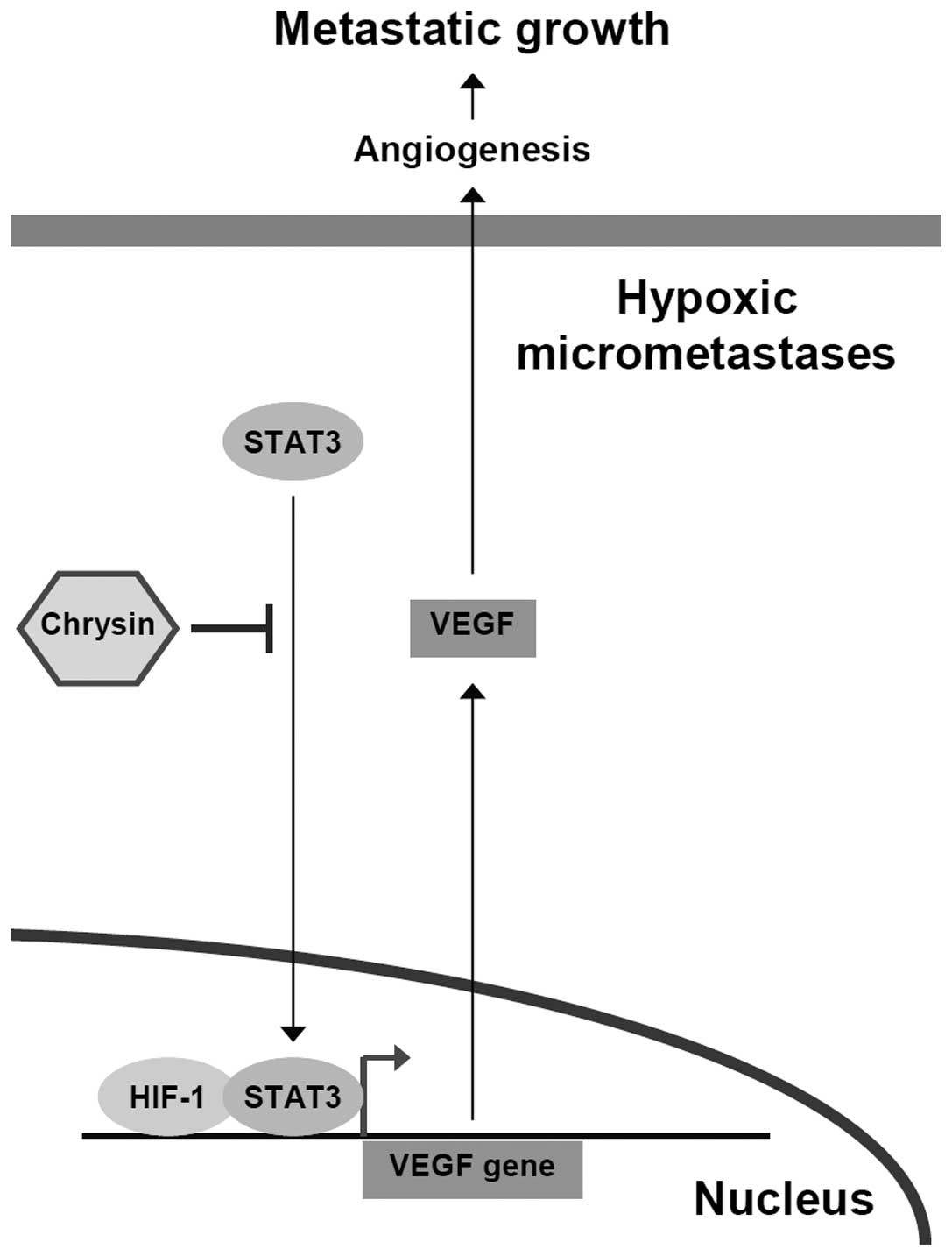
In conclusion, chrysin decreased hypoxic survival,
inhibited STAT3 activation, and diminished VEGF expression in
hypoxic cancer cells, leading to abrogation of hypoxia-induced
angiogenesis and resulting in suppression of metastatic growth
(Fig. 5). This study also suggests
that chrysin is a potential chemopreventive as well as an adjuvant
agent, that can be used in combination with other antimetastatic
agents to control metastatic progression of tumors.
Acknowledgements
This work was supported by the Thailand Research
Fund, TRG5380026 (K. Lirdprapamongkol), a Japanese-Thai
Collaborative Scientific Research Fellowship (JSPS-NRCT), and a
research grant from the Chulabhorn Research Institute. We would
like to thank Ms. Yue Zhou, Dr Orawin Prangsaengtong, Dr Rudee
Surarit, Ms. Korakot Atjanasuppat and Ms. Khajeelak Chiablaem for
their assistance.
References
|
1
|
Chambers AF, Groom AC and MacDonald IC:
Dissemination and growth of cancer cells in metastatic sites. Nat
Rev Cancer. 2:563–572. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Takeda K, Yamaguchi N, Akiba H, et al:
Induction of tumor-specific T cell immunity by anti-DR5 antibody
therapy. J Exp Med. 199:437–448. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hattori H, Okuda K, Murase T, et al:
Isolation, identification, and biological evaluation of
HIF-1-modulating compounds from Brazilian green propolis. Bioorg
Med Chem. 19:5392–5401. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Brizel DM, Scully SP, Harrelson JM, et al:
Tumor oxygenation predicts for the likelihood of distant metastases
in human soft tissue sarcoma. Cancer Res. 56:941–943.
1996.PubMed/NCBI
|
|
5
|
Hockel M, Schlenger K, Aral B, Mitze M,
Schaffer U and Vaupel P: Association between tumor hypoxia and
malignant progression in advanced cancer of the uterine cervix.
Cancer Res. 56:4509–4515. 1996.PubMed/NCBI
|
|
6
|
Li XF, Carlin S, Urano M, Russell J, Ling
CC and O’Donoghue JA: Visualization of hypoxia in microscopic
tumors by immunofluorescent microscopy. Cancer Res. 67:7646–7653.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Li XF and O’Donoghue JA: Hypoxia in
microscopic tumors. Cancer Lett. 264:172–180. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Iglesias P, Fraga M and Costoya JA:
Defining hypoxic microenvironments by non-invasive functional
optical imaging. Eur J Cancer. 49:264–271. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
McDonnell CO, Hill AD, McNamara DA, Walsh
TN and Bouchier-Hayes DJ: Tumour micrometastases: the influence of
angiogenesis. Eur J Surg Oncol. 26:105–115. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Minet E, Michel G, Mottet D, Raes M and
Michiels C: Transduction pathways involved in Hypoxia-Inducible
Factor-1 phosphorylation and activation. Free Radic Biol Med.
31:847–855. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Jung JE, Lee HG, Cho IH, et al: STAT3 is a
potential modulator of HIF-1-mediated VEGF expression in human
renal carcinoma cells. FASEB J. 19:1296–1298. 2005.PubMed/NCBI
|
|
12
|
Barr MP, Bouchier-Hayes DJ and Harmey JJ:
Vascular endothelial growth factor is an autocrine survival factor
for breast tumour cells under hypoxia. Int J Oncol. 32:41–48.
2008.PubMed/NCBI
|
|
13
|
Zhang J, Lu A, Beech D, Jiang B and Lu Y:
Suppression of breast cancer metastasis through the inhibition of
VEGF-mediated tumor angiogenesis. Cancer Ther. 5:273–286.
2007.PubMed/NCBI
|
|
14
|
Sun LP, Chen AL, Hung HC, et al: Chrysin:
a histone deacetylase 8 inhibitor with anticancer activity and a
suitable candidate for the standardization of Chinese propolis. J
Agric Food Chem. 60:11748–11758. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhao X, Shu G, Chen L, Mi X, Mei Z and
Deng X: A flavonoid component from Docynia delavayi
(Franch.) Schneid represses transplanted H22 hepatoma growth and
exhibits low toxic effect on tumor-bearing mice. Food Chem Toxicol.
50:3166–3173. 2012.PubMed/NCBI
|
|
16
|
Fu B, Xue J, Li Z, Shi X, Jiang BH and
Fang J: Chrysin inhibits expression of hypoxia-inducible
factor-1alpha through reducing hypoxia-inducible factor-1alpha
stability and inhibiting its protein synthesis. Mol Cancer Ther.
6:220–226. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wagner H, Chari VM and Sonnenbichler J:
13C-NMR-spektren natürlich vorkommender flavonoide. Tetrahedron
Lett. 17:1799–1802. 1976.(In German).
|
|
18
|
Gaydou E and Bianchini J: Etudes de
composés flavoniques. I. Syntheses et propriétés (UV, RMN du 13C)
de quelques flavones. Bull Soc Chim Fr II. 43–47. 1978.(In
French).
|
|
19
|
Lirdprapamongkol K, Mahidol C, Thongnest
S, et al: Anti-metastatic effects of aqueous extract of
Helixanthera parasitica. J Ethnopharmacol. 86:253–256. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lirdprapamongkol K, Chiablaem K, Sila-Asna
M, Surarit R, Bunyaratvej A and Svasti J: Exploring stemness gene
expression and vasculogenic mimicry capacity in well- and
poorly-differentiated hepatocellular carcinoma cell lines. Biochem
Biophys Res Commun. 422:429–435. 2012. View Article : Google Scholar
|
|
21
|
Mitra SK, Mikolon D, Molina JE, et al:
Intrinsic FAK activity and Y925 phosphorylation facilitate an
angiogenic switch in tumors. Oncogene. 25:5969–5984. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yang H, Xu Z, Iuvone PM and Grossniklaus
HE: Angiostatin decreases cell migration and vascular endothelium
growth factor (VEGF) to pigment epithelium derived factor (PEDF)
RNA ratio in vitro and in a murine ocular melanoma model. Mol Vis.
12:511–517. 2006.
|
|
23
|
Lin CM, Shyu KG, Wang BW, Chang H, Chen YH
and Chiu JH: Chrysin suppresses IL-6-induced angiogenesis via
down-regulation of JAK1/STAT3 and VEGF: an in vitro and in ovo
approach. J Agric Food Chem. 58:7082–7087. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Walle T, Otake Y, Brubaker JA, Walle UK
and Halushka PV: Disposition and metabolism of the flavonoid
chrysin in normal volunteers. Br J Clin Pharmacol. 51:143–146.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Vantyghem SA, Wilson SM, Postenka CO,
Al-Katib W, Tuck AB and Chambers AF: Dietary genistein reduces
metastasis in a postsurgical orthotopic breast cancer model. Cancer
Res. 65:3396–3403. 2005.PubMed/NCBI
|
|
26
|
Huang J, Frischer JS, Serur A, et al:
Regression of established tumors and metastases by potent vascular
endothelial growth factor blockade. Proc Natl Acad Sci USA.
100:7785–7790. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Guba M, von Breitenbuch P, Steinbauer M,
et al: Rapamycin inhibits primary and metastatic tumor growth by
antiangiogenesis: involvement of vascular endothelial growth
factor. Nat Med. 8:128–135. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lunt SJ, Cawthorne C, Ali M, et al: The
hypoxia-selective cytotoxin NLCQ-1 (NSC 709257) controls metastatic
disease when used as an adjuvant to radiotherapy. Br J Cancer.
103:201–208. 2010. View Article : Google Scholar : PubMed/NCBI
|