Introduction
With advances in medical science, the population of
elderly cancer patients has been increasing. This elderly
population has a lack of vital capacity, so they have a higher
incidence of serious side effects from conventional
chemotherapeutic agents. Therefore, modern chemotherapy strategy is
emphasizing quality of life. Recently much interest has been drawn
to the possibility of controlling cancer with minimal toxicity
using flavonoids from fruit because the flavonoids in fruit can
safely enhance anticancer effects (1–3).
Citrus fruits are edible and their dried peels have
been used as a component of traditional medicine for various
diseases including cancer since ancient times. Citrus unshiu
Marc. called ‘Kamgyul’ in Korean is widely cultivated in Jeju
island, Korea. The peels of Citrus unshiu Marc. are used in
traditional herbal medicine and frequently prescribed in concert
with other herbs for many kinds of disease including cancer. The
flavoids isolated from Citrus fruit are mainly composed of
hesperidin, naringenin, and nobiletin (4). These compounds have been reported to
have some anticancer properties (5). We isolated the flavonoids from the
Citrus unshiu Marc. fruit peel (FCM).
Most of cancer patients eventually die of cancer
metastasis. Inhibition of the metastatic activity is one of the
cancer treatment strategies. Metastasis consists of a series of
sequential complicated steps. Initially, cancer cells have to
detach from the primary lesion, and invade the basal membrane and
move to other lesions though lymphatic system or blood vessels.
Before invading the underlying basement membrane, the cancer cells
temporarily adhere to endothelial cells, and then extravasate. The
ability to adhere to the endothelial cell correlates with the
capacity of cancer cells to form metastatic lesions (6,7).
Cell adhesion molecules (CAMs) are involved in a
broad range of normal physiological processes, but the pivotal role
of CAMs is emphasized by the fact that CAMs are involved in a
variety of pathologies, especially cancer (8). Most of the cell adhesion molecules
(CAMs) belong to four protein families: Ig (immunoglobulin)
superfamily (IgSF CAMs), the integrins, the cadherins, and the
selectins. Among IgSF CAMs, vascular cell adhesion molecule-1
(VCAM-1) and intracellular adhesion molecule-1 (ICAM-1) are
regulated by NF-κB, closely related in structure, well known
endothelial surface adhesion molecules involved in cancer cells
(7). However, it has been reported
that some highly metastatic cancer cells migrate and adhere to
VCAM-1 rather than to ICAM-1 (9),
suggesting that VCAM-1 may relate more to cancer metastasis by
enhancing the adherence of cancer to endothelial cells. Therefore,
it is important to discover that FCM have inhibitory effects on
cancer adhesion and the expression of VCAM-1 and ICAM-1. We
investigated the anticancer activity of FCM on the ability of
cancer to adhere to the endothelial cells and elucidated the
mechanism regarding the expression of VCAM-1 and ICAM-1.
Materials and methods
Preparation of FCM
The flavonoids were isolated from the Citrus
unshiu Marc. fruit peel by extraction with 70% aqueous methanol
followed by ethyl acetate elution over a silica gel cartridge. The
isolated flavonoids were identified by HPLC using a C18 column. We
identified 16 flavonoid components but 2 compounds (hesperetin
7-O-glucoside and hesperetin) were not quantified; a) naringin; b)
hesperidin; c) poncirin; d) isosinensetin; e) hexamethoxyflavone;
f) sinensetin; g) hexamethoxyflavone; h)
tetramethyl-O-isoscutellarein; i) nobiletin; j)
3,30,40,5,6,7,8-heptamethoxyflavone; k)
3-hydroxyhexamethoxyflavone; l) tangeretin; m)
3-hydroxypentamethoxy-flavone; n) hexamethoxyflavone = 54:32:2:0.4:
0.2:1.03:0.2:3.7:1.6:0.4:2.2:0.4:0.2. Among FCM, naringin and
hesperidin were the major compounds (1).
Cells and reagents
Human umbilical vein endothelial cells (HUVECs)
(EA.hy 926 cells) were obtained from ATCC and grown in medium 199
supplemented with 20% FBS, 2 mM L-glutamine, 5 U/ml heparin, 100
IU/ml penicillin, 10 μg/ml streptomycin and 50 μg/ml EC growth
supplements. Cells were cultured in 100-mm dishes and grown in a
humidified 5% CO2 incubator. HUVECs were used between
passage number 3 and 6. The human breast cancer cell line
MDA-MB-231, was obtained from the Korea Cell Line Bank (Seoul,
Korea) and grown in RPMI-1640 supplemented with 10% FBS, 2 mM
L-glutamine, 25 mM N-2-hydroxyethylpiperazine-N′-2-ethanesulfonic
acid, 25 mM NaHCO3, 100 IU/ml penicillin and 10 μg/ml
streptomycin at 37°C in a humidified atmosphere of 95% air and 5%
CO2. 3-(4,5-Dimethylthiazole-2-yl)-2,5-biphenyl
tetrazolium bromide (MTT), 4′,6-diamidino-2-phenyindole, dilactate
(DAPI), anti-β-actin antibody were obtained from Sigma-Aldrich Co.
(St. Louis, MO, USA). The polyclonal anti-ICAM-1 and VCAM-1
antibodies were purchased from Santa Cruz Biotechnology, Inc.
(Santa Cruz, CA, USA). Recombinant human tumor necrosis factor
(TNF) was obtained from R&D Systems (Minneapolis, MN, USA). BD
Matrigel™ basement membrane matrix was supplied by BD Biosciences
(San Diego, CA, USA).
Cell viability assay
Cell viability was determined colorimetrically using
MTT reagent. Cells were seeded at 104 cells/well in
24-well plates. After treatments, 50 μl of 5 mg/ml MTT solution was
added to each well and incubated for 3 h. The supernatants were
aspirated and the formazan crystals were dissolved with 200 μl of 4
N HCl-isopropanol in each well. The optical density of the colored
product was measured at 570 nm, as suggested by the manufacturer,
using an Infinite 200 microplate reader (Tecan Austria GmbH,
Grödig, Austria).
Western blot analysis
The cells were washed in ice-cold PBS and lysed in
PRO-PREP protein extraction solution (iNtRON Biotechnology, Seoul,
Korea). The samples were centrifugated at 13,000 rpm, for 15 min at
4°C. An aliquot of the whole cell lysate was subjected to sodium
dodecyl sulfate (SDS)-polyacrylamide gel electrophoresis and
transferred onto polyvinylidene difluoride membrane. Membranes,
blocked with 5% nonfat milk in Tris-buffered saline (TBS)
containing 0.05% Tween-20 for 2 h at room temperature, were
incubated with anti-ICAM-1 and VCAM-1 antibodies at 1:1,000 in TBS
containing 0.05% Tween-20 and 3% bovine serum albumin (BSA) for
overnight at 4°C. The membranes were then incubated with
horseradish peroxidase-conjugated anti-rabbit IgG (1:5,000)
antibody for 1 h at room temperature. After washing, the membranes
were developed using the ECL reagent (Bionote, Gyeonggi-do,
Korea)
Adhesion assay
HUVECs were seeded into a 6-well plate and treated
with the reagents indicated. MDA-MB-231 cells were pelleted and
resuspended (7.5×105 cells/ml) in RPMI-1640 medium.
HUVECs were washed with serum-free medium, and MDA-MB-231 cells
were applied onto HUVECs at 37°C. After 30 min, cell suspensions
were withdrawn, and the HUVECs were gently washed with PBS. Adhered
MDA-MB-231 cells to HUVECs were counted under a light microscope.
Analyses were repeated three times, over the same region, and the
results of the three independent experiments were similar.
Matrigel invasion assay
MDA-MB-231 cells were cultured for 3 days. The upper
chamber of 24-well cell culture inserts (8 μm pore size, Falcon,
Franklin Lakes, NJ, USA) were washed with a serum-free medium,
coated with 100 μl of Matrigel (1 mg/ml) and dried for 30 min at
37°C. MDA-MB-231 cells treated with FCM were collected;
2×105 cells loaded to the upper chambers filled
serum-free media, and 500 μl of RPMI media containing 10% FBS was
added to the lower chambers. The invasion chambers were incubated
for 24 h in a 37°C cell culture incubator. The non-invasive cells
that remained on the upper surface of the insert membranes were
removed by scrubbing. The cells on the lower insert membranes were
stained with DAPI, and cells were counted under the fluorescence
microscope. Each sample was measured in triplicate, and each
experiment was repeated three times.
Gelatin zymography
Conditioned media collected from MDA-MB-231 cells
were concentrated approximately 10-fold with a protein concentrator
(Thermo Pierce, Rockford, IL, USA). Concentrated media were mixed
with 2X sample loading buffer (2% Glycerol, 0.4% SDS, 0.05%
Bromophenol blue) and subjected to 0.1% gelatin contained gel
electrophoresis without heating. After electrophoresis, gels were
incubated with renaturation buffer (2.5% Triton X-100) for 1 h at
room temperature, and then washed with distilled water three times.
Gels were applied with developing buffer (50 mM Tris (pH 7.6), 20
mM NaCl, 5 mM CaCl2, 0.02% Brij35) overnight at 37°C,
and then washed with distilled water three times. After washing,
gels were stained with coomassie blue solution (0.2% coomassie
brilliant blue, 50% methanol, 10% acetic acid) for 1 h and
destained with destaining buffer (50% methanol, 10% acetic acid).
Gelatinolytic activity was detected as clear bands in the
background of blue staining.
Statistical analysis
Each experiment was performed in triplicate. The
results were expressed as the means ± SE. Significant differences
were determined using the one-way analysis of variance (ANOVA) with
post-test Neuman-Keuls for more than two groups and Student’s
t-test for two groups. Statistical significance was defined as
p<0.05.
Results
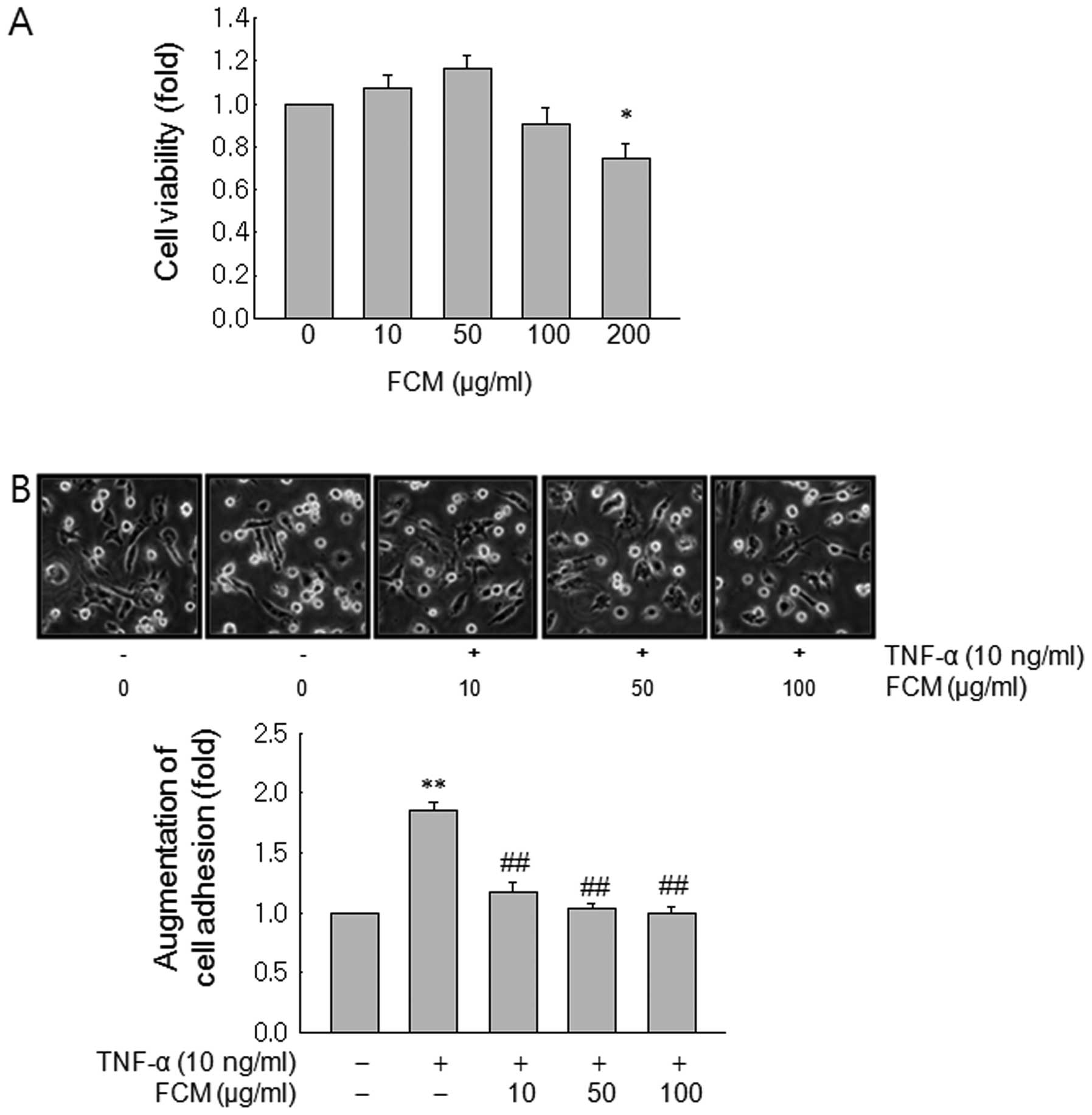
FCM inhibited TNF-induced cancer cell
adhesion to human umbilical vein endothelial cells (HUVECs)
At first, we assessed anti-proliferative effects of
FCM on MDA-MB-231 cells. MTT test revealed that the growth of
MDA-MB-231 cells was inhibited by FCM treatment only at a dose of
200 μg/ml (Fig. 1A). Next we
performed adhesion assay to test the inhibitory effects on cancer
cell adhesion to endothelial cells at the concentrations (10–100
μg/ml) where FCM did not show anti-proliferative effects. The
adhesion assay revealed that FCM significantly inhibited
TNF-induced cancer cell adhesion to HUVECs from the low dose of 10
μg/ml (Fig. 1B).
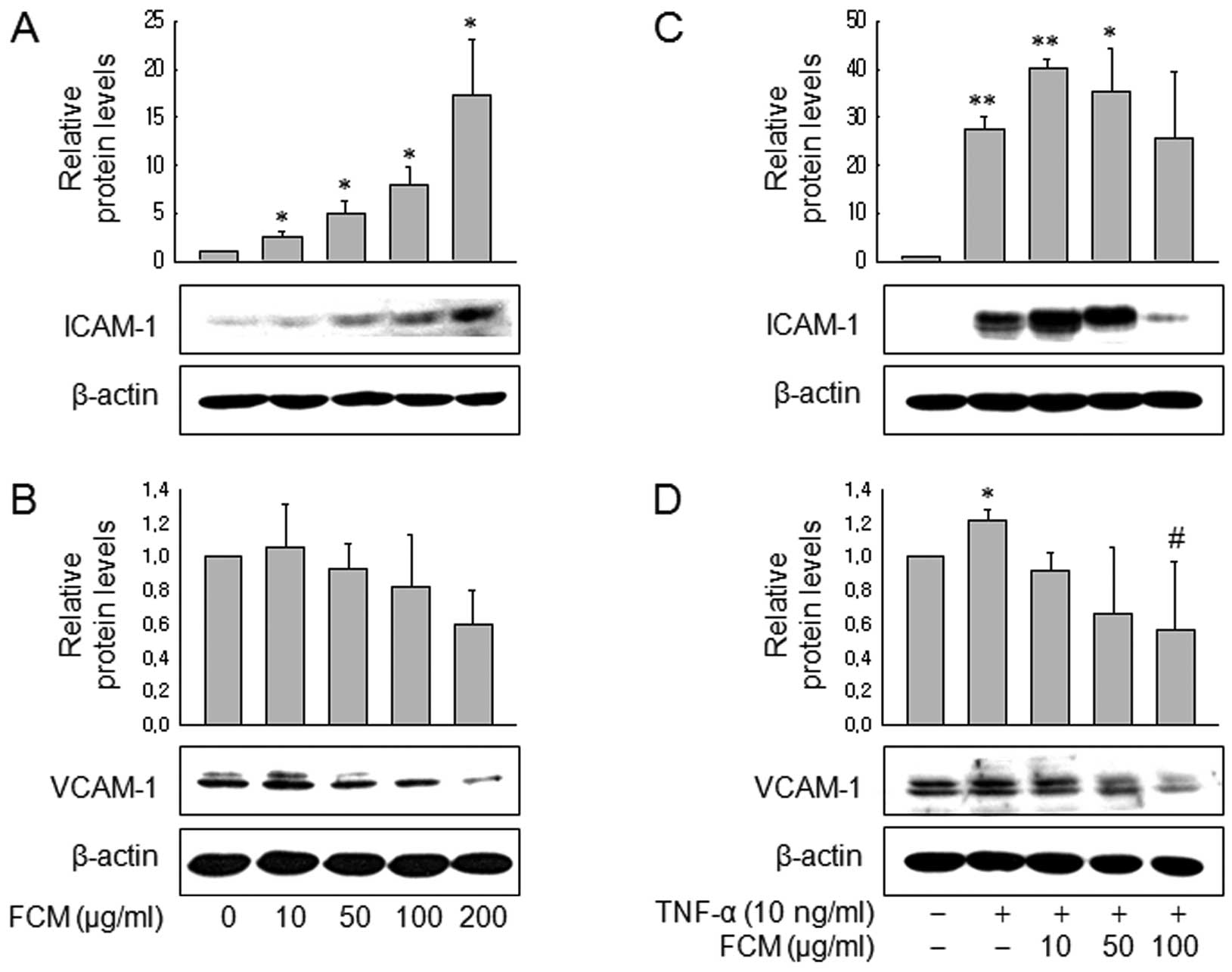
FCM inhibited VCAM-1 expression, but not
ICAM-1 expression
We then assessed the effects of FCM on the
expression of VCAM-1 and ICAM-1 to further investigate this finding
at the molecular level. Western blot analysis revealed that FCM
significantly inhibited VCAM-1 expression, not only in control
MDA-MB-231 cells (Fig. 2A and B)
but also in TNF-treated MDA-MB-231 cells (Fig. 2C and D). Of note, FCM increased
ICAM-1 expression in both the control and TNF-treated MDA-MB-231
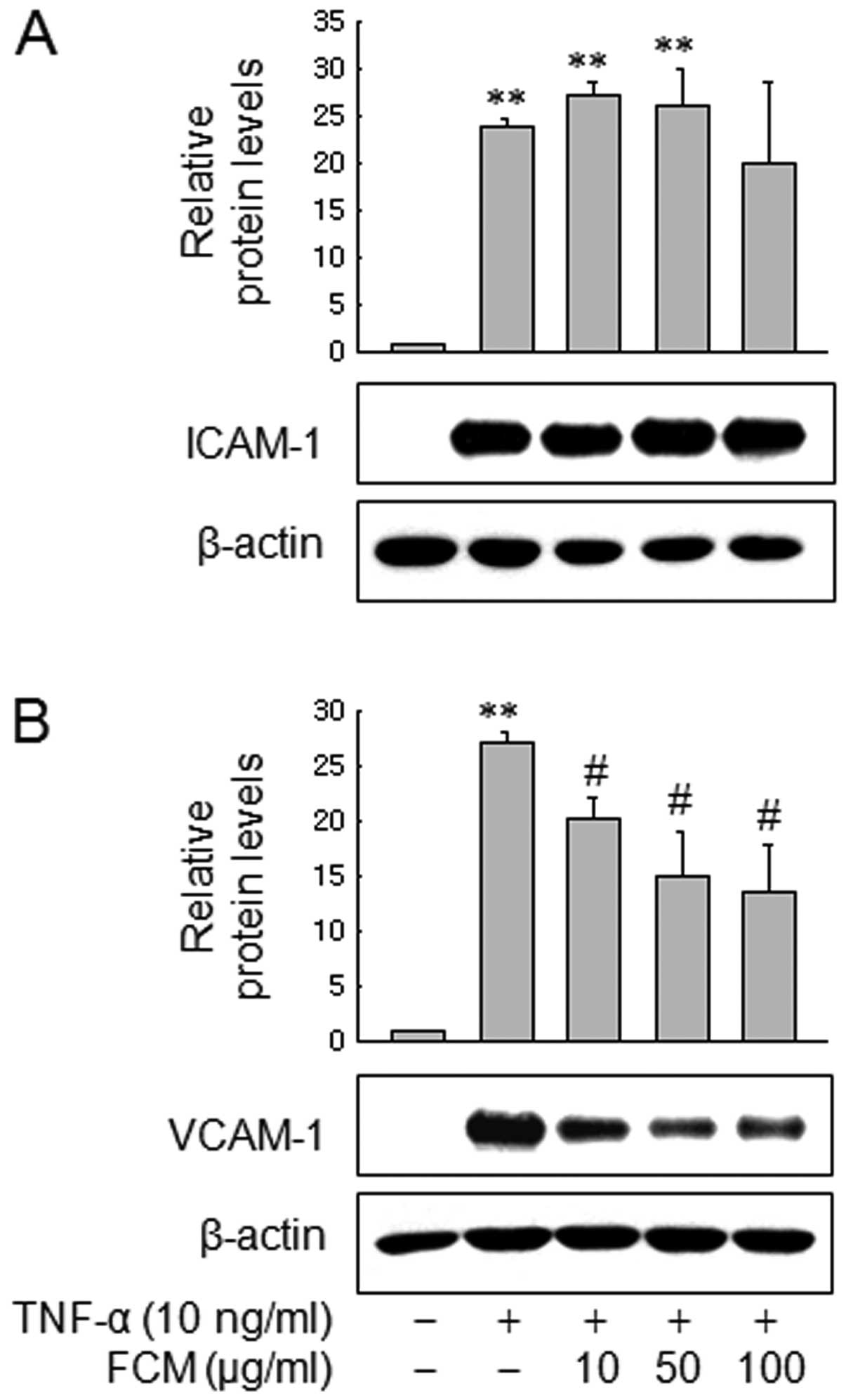
cells. Next we also assessed expression of VCAM-1 and ICAM-1 in
HUVECs. Consistent with the data in MDA-MB-231 cells, western blot
analysis revealed that FCM inhibited VCAM-1 expression in a
dose-dependent manner, but not ICAM-1 expression (Fig. 3A and B). These findings suggest that
FCM may inhibit TNF-induced cancer cell adhesion to HUVECs through
inhibiting VCAM-1 expression.
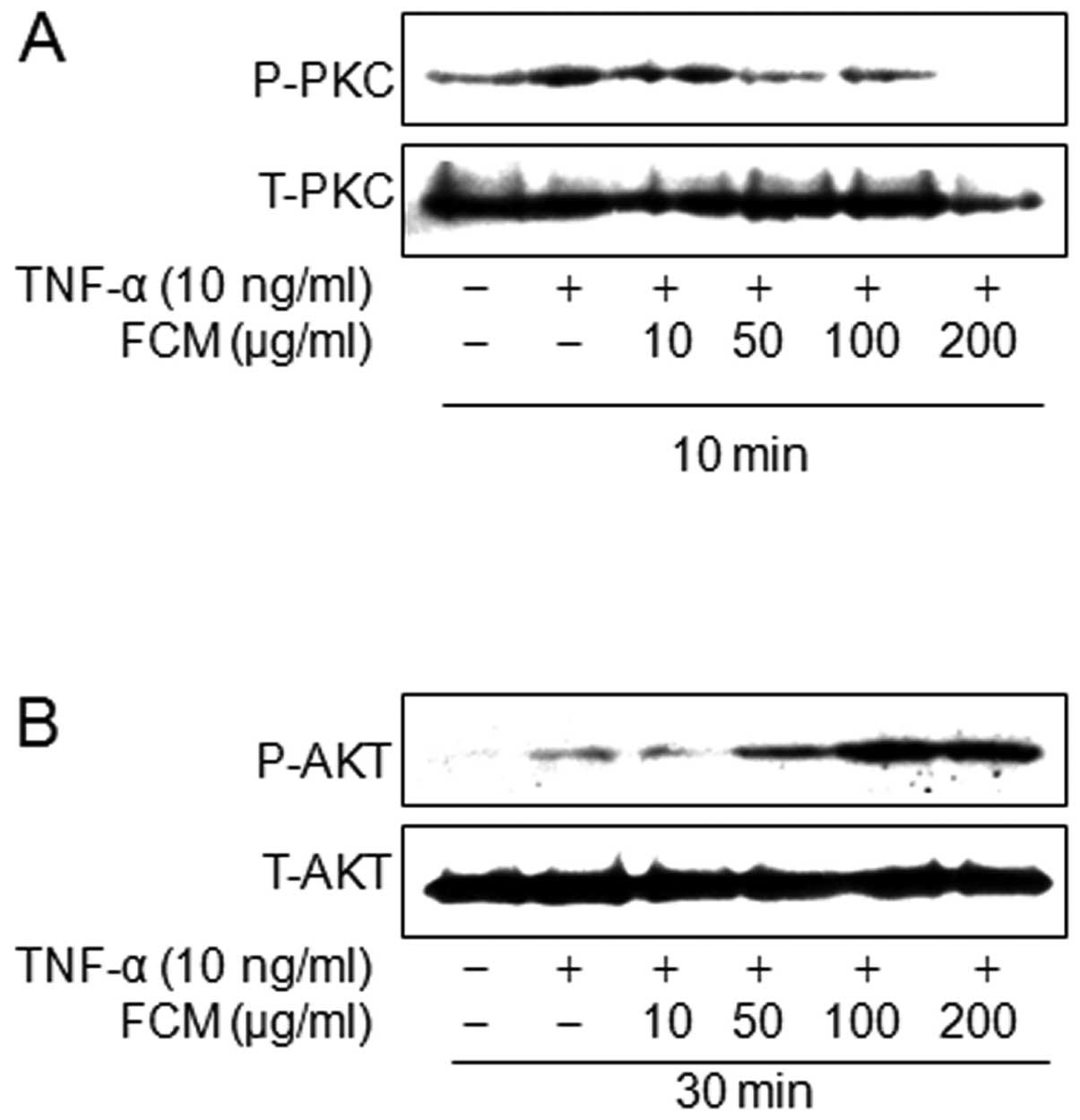
FCM inhibited protein kinase C (PKC)
phosphorylation, but not Akt phosphorylation
Of TNF-mediated signaling, PI3K/Akt and PKC
signaling pathways are involved in VCAM-1 expression rather than
ICAM-1 expression (10). Herein, we
investigated the effects of FCM on PI3K/Akt and PKC phosphrylation
activated by TNF. Western blotting revealed that FCM inhibited the
TNF-induced PKC phosphorylation, but that FCM increased Akt
phosphorylation (Fig. 4A and B).
These findings suggest that the inhibitory effects of FCM on VCAM-1
expression may be linked to inhibition of TNF-induced PKC
phosphorylation, but not Akt phosphorylation.
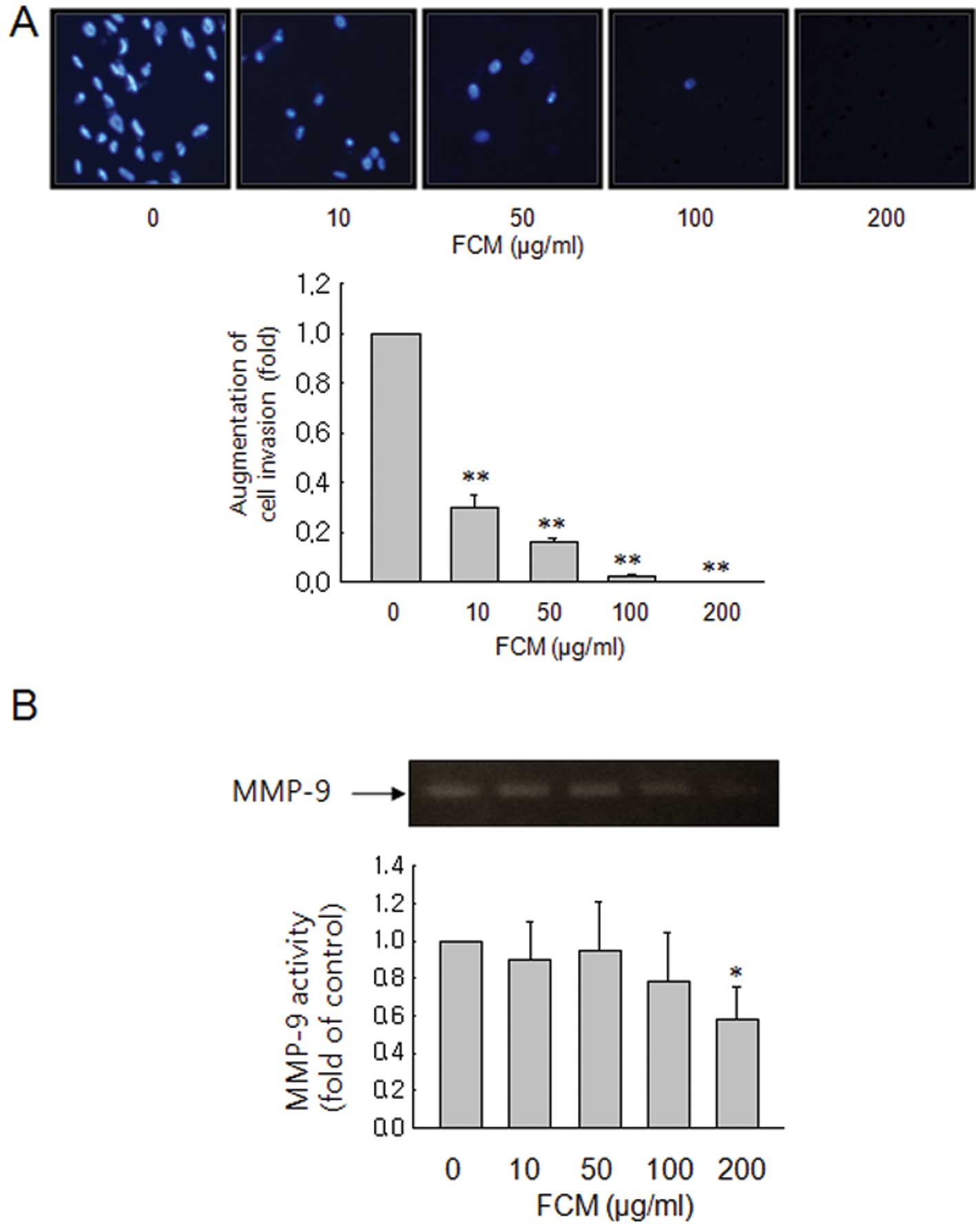
FCM inhibits cell invasion, but not
through inhibition of MMP expression
Activation of PKC induces rapid changes in cell
morphology, cell-cell adhesion, and cell migration (11). Therefore, we also performed Matrigel
invasion assays to assess the effects of FCM on cancer cell
invasion. FCM markedly inhibited cell invasion in a dose-dependent
manner (Fig. 5A). To verify the
molecular mechanisms, we also measured by gelatin zymographic
analyses the activities of the secreted MMP-9 from FCM-treated
cancer cells. As shown in Fig. 5B,
FCM barely suppressed the gelatinolytic activities of secreted
MMP-9, indicating that other mechanisms such as suppression of
motility are involved in the inhibitory effects of FCM on cancer
invasion.
Discussion
Adhesive interaction of cancer cells out of blood
vessels is an important step in cancer metastasis, because cancer
cells are rapidly eliminated from the circulation unless they can
adhere to new vasculature to establish new metastatic colonies.
Therefore, the adhesion molecules such as ICAM-1 and VCAM-1 are
important in the cancer metastasis. This study was designed to
investigate the effects of FCM on cancer metastasis. Accordingly we
examined the effect of FCM on cancer invasion and adherence in the
endothelial cells, and elucidated its mechanism regarding the
regulation of VCAM-1 and ICAM-1 in MDA-MB 231 human breast cancer
cells. MDA-MB 231 cells are a proper model to study cancer
metastasis, since the cells are more aggressive, possessing high
metastatic potential, and are unresponsive to anti-estrogens due to
estrogen receptor negativity (12).
In this study, we found that FCM inhibited TNF-induced cancer cell
adhesion to human umbilical vein endothelial cells (HUVECs) through
inhibiting VCAM-1 expression, and that the inhibitory effects of
FCM on VCAM-1 expression appear to be connected to inhibition of
protein kinase C (PKC) phosphorylation induced by TNF.
Previous studies demonstrated TNF induced both
ICAM-1 and VCAM-1 proteins on vascular endothelial cells (13). ICAM-1 and VCAM-1 are both regulated
by NF-κB. Since TNF is an NF-κB stimulator and naringin and
hesperidin that are the major components of FCM have some
inhibitory effects on NF-κB (14,15),
we expected that FCM may inhibit both ICAM-1 and VCAM-1 expression
induced by TNF. Of note, however, FCM suppressed the TNF-α-induced
expression of VCAM-1, but not of ICAM-1. Relating to this result,
there is a previous study demonstrating that hesperidin selectively
inhibited TNF-induced VCAM-1 expression through suppressing PKC
pathway, leading to inhibition of the adhesion of U937 leukemic
cells to HUVECs (16). This result
is consistent with our results showing the preferential inhibition
of VCAM-1 expression over ICAM-1 by FCM. In addition, this study
demonstrated that the suppression of VCAM-1 expression was
effective in blocking cancer cell adhesion to HUVECs in
TNF-stimulated condition. These results were also similar to those
of a previous study (17). VCAM-1
and ICAM-1 can be regulated by different signaling pathways, and
VCAM-1 is more important in cancer cell adhesion than other
adhesion molecules especially for highly metastatic cancer cells
(9). Consequently, the suppressive
effects of FCM on VCAM-1 expression of endothelial cells as well as
cancer cells may be valuable in interfering with cancer metastasis.
In this selective suppression of VCAM-1 expression, Akt and PKC are
involved (17), and our result
showed that FCM inhibit VCAM-1 expression by TNF through regulation
of PKC pathway. PKC pathways is well-known to be involved in cancer
cell mobility (11). In the process
of cancer invasion, cell migration is also required to pass though
the basement membrane.
Proteolytic digestion of the extracellular matrix
(ECM) by secreted MMPs is one of major steps in cancer invasion
(18,19). MMP-9 is also a biomarker for
epithelial mesenchymal transition (EMT) (20). When we measured the activities of
the secreted MMP-9 from FCM-treated cancer cell by gelatin
zymographic analysis, FCM barely suppressed the gelatinolytic
activity of secreted MMP-9. This finding suggests that inhibitory
effects of FCM on cancer cell invasion may relate to the other
mechanisms such as suppression of motility. Besides MMP-9, we also
assessed the changes in EMT biomarkers to confirm that FCM have
inhibitory effects on EMT (data not shown). Western blotting showed
that FCM did not suppress the mesenchymal markers vimentin, and
N-cadherin, and that the expression of E-cadherin epithelial marker
was not detected in MDA-MB-231 cells (data not shown). In this
study, TNF was used to clearly demonstrate the effects of FCM on
cancer cell adhesion to HUVECs. Therefore, the pathophysiological
relevance that TNF is usually increased in patients with advanced
cancers (21) is supporting that
TNF-augmented cancer cell adhesion to HUVECs shown in this study is
not an artificial in vitro experiment.
In conclusion, this study demonstrated that FCM
suppressed the cancer cell adhesion to HUVECs through selective
suppression of VCAM-1. The inhibitory effects of FCM on VCAM-1
expression may be linked to inhibition of PKC phosphorylation. This
study provides evidence that FCM may have anti-metastatic activity
by inhibiting adhesion molecules and invasion on human breast
cancer cells.
Acknowledgements
This study was supported by grants from the National
R&D Program for Cancer Control, Ministry of Health &
Welfare, Republic of Korea (0820050).
References
|
1
|
de Sousa RR, Queiroz KC, Souza AC,
Gurgueira SA, Augusto AC, Miranda MA, Peppelenbosch MP, Ferreira CV
and Aoyama H: Phosphoprotein levels, MAPK activities and NFkappaB
expression are affected by fisetin. J Enzyme Inhib Med Chem.
22:439–444. 2007.PubMed/NCBI
|
|
2
|
Liu BL, Zhang X, Zhang W and Zhen HN: New
enlightenment of French Paradox: resveratrol’s potential for cancer
chemoprevention and anti-cancer therapy. Cancer Biol Ther.
6:1833–1836. 2007.PubMed/NCBI
|
|
3
|
Chun KH, Kosmeder JW, Sun S, Pezzuto JM,
Lotan R, Hong WK and Lee HY: Effects of deguelin on the
phosphatidylinositol 3-kinase/Akt pathway and apoptosis in
premalignant human bronchial epithelial cells. J Natl Cancer Inst.
95:291–302. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Nogata Y, Sakamoto K, Shiratsuchi H, Ishii
T, Yano M and Ohta H: Flavonoid composition of fruit tissues of
citrus species. Biosci Biotechnol Biochem. 70:178–192. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Manthey JA, Grohmann K and Guthrie N:
Biological properties of citrus flavonoids pertaining to cancer and
inflammation. Curr Med Chem. 8:135–153. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Nicolson GL: Organ specificity of tumor
metastasis: role of preferential adhesion, invasion and growth of
malignant cells at specific secondary sites. Cancer Metastasis Rev.
7:143–188. 1988. View Article : Google Scholar
|
|
7
|
Orr FW, Wang HH, Lafrenie RM, Scherbarth S
and Nance DM: Interactions between cancer cells and the endothelium
in metastasis. J Pathol. 190:310–329. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Okegawa T, Pong RC, Li Y and Hsieh JT: The
role of cell adhesion molecule in cancer progression and its
application in cancer therapy. Acta Biochim Pol. 51:445–457.
2004.PubMed/NCBI
|
|
9
|
Klemke M, Weschenfelder T, Konstandin MH
and Samstag Y: High affinity interaction of integrin alpha4beta1
(VLA-4) and vascular cell adhesion molecule 1 (VCAM-1) enhances
migration of human melanoma cells across activated endothelial cell
layers. J Cell Physiol. 212:368–374. 2007. View Article : Google Scholar
|
|
10
|
Mun L, Jun MS, Kim YM, Lee YS, Kim HJ, Seo
HG, Lee JH, Son KH, Lee DH, Kim YS, Park K and Chang KC:
7,8-Didehydrocimigenol from Cimicifugae rhizoma inhibits
TNF-alpha-induced VCAM-1 but not ICAM-1expression through
upregulation of PPAR-gamma in human endothelial cells. Food Chem
Toxicol. 49:166–172. 2011.
|
|
11
|
Weinstein IB, Lee LS, Fisher PB, Mufson A
and Yamasaki H: Action of phorbol esters in cell culture: mimicry
of transformation, altered differentiation, and effects on cell
membranes. J Supramol Struct. 12:195–208. 1979. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Anandappa SY, Sibson R, Platt-Higgins A,
Winstanley JH, Rudland PS and Barraclough R: Variant estrogen
receptor alpha mRNAs in human breast cancer specimens. Int J
Cancer. 88:209–216. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ahmad M, Zhang Y, Papharalambus C and
Alexander RW: Role of isoprenylcysteine carboxyl methyltransferase
in tumor necrosis factor-alpha stimulation of expression of
vascular cell adhesion molecule-1 in endothelial cells.
Arterioscler Thromb Vasc Biol. 22:759–764. 2002. View Article : Google Scholar
|
|
14
|
Ang ES, Yang X, Chen H, Liu Q, Zheng MH
and Xu J: Naringin abrogates osteoclastogenesis and bone resorption
via the inhibition of RANKL-induced NF-kappaB and ERK activation.
FEBS Lett. 585:2755–2762. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ghorbani A, Nazari M, Jeddi-Tehrani M and
Zand H: The citrus flavonoid hesperidin induces p53 and inhibits
NF-kappaB activation in order to trigger apoptosis in NALM-6 cells:
involvement of PPARgamma-dependent mechanism. Eur J Nutr. 51:39–46.
2012. View Article : Google Scholar
|
|
16
|
Nizamutdinova IT, Jeong JJ, Xu GH, Lee SH,
Kang SS, Kim YS, Chang KC and Kim HJ: Hesperidin, hesperidin methyl
chalone and phellopterin from Poncirus trifoliata (Rutaceae)
differentially regulate the expression of adhesion molecules in
tumor necrosis factor-alpha-stimulated human umbilical vein
endothelial cells. Int Immunopharmacol. 8:670–678. 2008.PubMed/NCBI
|
|
17
|
Sun DI, Nizamutdinova IT, Kim YM, Cai XF,
Lee JJ, Kang SS, Kim YS, Kang KM, Chai GY, Chang KC and Kim HJ:
Bisacurone inhibits adhesion of inflammatory monocytes or cancer
cells to endothelial cells through down-regulation of VCAM-1
expression. Int Immunopharmacol. 8:1272–1281. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Vihinen P and Kahari VM: Matrix
metalloproteinases in cancer: prognostic markers and therapeutic
targets. Int J Cancer. 99:157–166. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Deryugina EI and Quigley JP: Matrix
metalloproteinases and tumor metastasis. Cancer Metastasis Rev.
25:9–34. 2006. View Article : Google Scholar
|
|
20
|
Radisky ES and Radisky DC: Matrix
metalloproteinase-induced epithelial-mesenchymal transition in
breast cancer. J Mammary Gland Biol Neoplasia. 15:201–212. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Correia M, Cravo M, Marques-Vidal P,
Grimble R, Dias-Pereira A, Faias S and Nobre-Leitao C: Serum
concentrations of TNF-alpha as a surrogate marker for malnutrition
and worse quality of life in patients with gastric cancer. Clin
Nutr. 26:728–735. 2007. View Article : Google Scholar : PubMed/NCBI
|