Introduction
Cervical cancer is the third most common cancer
worldwide, and more than 85% of cervical cancer cases occur in
developing countries (1). It is
well known that infection of the oncogenic type of HPVs,
particularly HPV 16, is an etiologic factor of cervical cancers
(2,3). Although the two HPV major capsid
protein L1 virus-like particle-based preventive vaccines have a
remarkable safety profile and clinical efficacy against the HPV
genotypes from which they were derived, they are not effective in
the elimination of pre-existing infection and HPV-associated
diseases (4,5). Thus, it is urgent to develop
therapeutic HPV vaccines. Since the HPV oncoprotein E7 is
constitutively expressed in HPV-infected cells and cervical
cancers, it has become an attractive target for the development of
HPV therapeutic vaccines (6–9).
DNA vaccines have become an attractive approach for
generating antigen-specific immunotherapy. Naked plasmid DNA can
generate effective cytotoxic T lymphocytes (CTLs) and antibody
responses by delivering foreign antigens to antigen-presenting
cells (APCs) that stimulate CD4+ and CD8+ T
cells. They are easily prepared with high purity and stability and
can be repeatedly administered (10). Several versatile immune stimulatory
molecules have been used to overcome the weak immunogenicity of DNA
vaccines, one of which is HSP70, a promising molecule due to its
attractive adjuvant activity in enhancing antigen-specific immunity
(11–13). Immunological functions of HSP70 can
be categorized into chaperoning properties, cross-priming abilities
and linking danger and pathogen recognition activities (14–20).
Many HSP70-based therapeutic cancer vaccines have been reported
(11,13,21,22).
We previously used human HSP70 or Mycobacterium tuberculosis
HSP70 as a dendritic cell (DC)-targeting molecule to construct two
HPV 16 mE7-based fusion DNA vaccines with a leader peptide gene
sequence of CD33 (Sig) at the upstream of the fusion gene,
SigmE7/MtHSP70 and SigmE7/HuHSP70 DNA. We found that vaccination of
both the fusion DNA vaccines effectively enhanced the antitumor
responses (23). The reason for
choosing the leader sequence of the human CD33 as the signal
peptide was that it has been used in tumor cell vaccines
transfected with secretable HSP70 and displays enhanced antitumor
efficacy (21). Currently, some
HSP70-based HPV 16 E7 fusion DNA vaccines with or without an
additional leading peptide sequence have shown promise in further
clinical trials (11,13,24).
To investigate which strategy may facilitate HSP70 to function more
efficiently in therapeutic fusion DNA vaccines, we considered two
options: a strategy which constructs an antigen-HSP70 fusion gene
alone or as an antigen-HSP70 fusion gene with a leading
peptide.
In the present study, we constructed mE7/MtHSP70
fused tumor DNA vaccine and compared its potency in mice with
pre-constructed SigmE7/MtHSP70 fusion DNA vaccine with a leader
peptide gene sequence of the CD33 molecule at the upstream of the
fusion gene. Our results showed that without the help of the CD33
signal peptide, the mE7/MtHSP70 fusion protein was efficiently
release from cells, and vaccination of mE7/MtHSP70 fusion DNA
vaccine induced more effective CD8+ T cell responses and
antitumor effects than did SigmE7/MtHSP70 DNA. Our data suggest
that without the help of a signal peptide, HSP70 can play a more
powerful adjuvant role in antigen and HSP70 fused tumor DNA
vaccine.
Materials and methods
Plasmid DNA constructs and
preparation
We previously reported the modified HPV 16 E7 gene
with abolishment of its potential transformation activity and
enhanced immunogenicity by a combination of gene shuffling,
site-directed mutagenesis and codon optimization methods (23,25).
mE7 gene and SigmE7 containing SigCD33 were amplified by overlap
PCR with the primers:
5′-CGAGTCGTGCGGCCGCCACCATGCCGCTGCTGCTACTGCTGCCCCTGCTGTGG GCAG-3′
(NotI),
5′-CTGCCCCTGCTGTGGGCAGGGGCCCTGGCTATGATGGATCTGCTCATGGGCAC-3′ and
5′-GCTCTAGAGCGGTAGTCTCGGGCTGCAG-3′ (XbaI). We digested Sig
mE7 with NotI and XbaI and ligated it to
NotI/XbaI-digested pVR1012 to generate
pVR1012-SigmE7. To generate pVR1012-SigmE7/MtHSP70, MtHSP70
digested with XbaI and BamHI from pVR1012-mE7/MtHSP70
(constructed by our laboratory) was ligated into
XbaI/BamHI-digested pVR1012-SigmE7. All constructs
were validated by restriction enzyme digestion and DNA sequencing.
Plasmid DNA was prepared with EndoFree Plasmid Purification kits
from Qiagen Inc. (Valencia, CA, USA) resuspended in endotoxin-free
normal saline at a concentration of 1 μg/μl. The integrity of the
DNA plasmids was verified by electrophoresis on a 1% agarose gel.
DNA concentration was determined by absorbance measured at 260
nm.
Western blot analysis
COS-7 cells with 70% confluence in a 6-well plate
were transfected with 10 μg plasmid DNA using Lipofectamine 2000
(Invitrogen). Supernatants and cells were harvested 48 h after
transfection. The cells were lysed in 50 mM Tris HCl, 150 mM NaCl,
1 mM EDTA, 1% NP-40 and protease inhibitors. The protein
concentration was determined using the BCA protein assay kit
(Pierce). Each lysate (60 μg) or 40 μl of 4-fold concentrated
supernatants was denatured at 100°C for 5 min, loaded on a 10%
SDS-PAGE gel separated under reducing conditions, and transferred
to polyvinylidene difluoride membranes (Bio-Rad Laboratories,
Hercules, CA, USA). Membranes were blocked overnight with 4% BSA
and incubated with polyclonal rabbit anti-HPV 16 E7 antibody
(1:2,000) followed by horseradish peroxidase-conjugated goat
anti-rabbit IgG (1:10,000; Beijing Zhongshan Golden Bridge
Biotechnology Co., Ltd., Beijing, China). Blots were developed by
chemiluminescence reagent (ECL kit; Pierce).
Mice and tumor cell line
Six- to 8-week-old female C57BL/6 mice were
purchased from the Institute of Zoology, Chinese Academy Sciences,
and were maintained at the animal facility of the Institute of
Laboratory Animal Sciences, Chinese Academy of Medical Sciences and
Peking Union Medical College. All experimental protocols were
approved by the Institutional Animal Care and Use Committee. Care
was taken to minimize pain and discomfort to all animals during the
procedures in the present study. TC-1 cells were generated by
co-transfection of primary pulmonary epithelial cells from C57BL/6
mice with HPV16 E6 and E7 and activated c-Ha-ras oncogenes. The
cells were grown in RPMI-1640 supplemented with 10% fetal calf
serum, 2 mM L-glutamine, 1 mM sodium pyruvate, 2 mM non-essential
amino acids and 0.4 mg/ml G418 and antibiotics.
DNA vaccination
The mice were divided into three groups (n=7 per
group). The mice were injected intramuscularly (i.m.) with 125 μg
bupivacaine hydrochloride into each side of the M. quadriceps. One
day later, the mice were inoculated with 50 μg of DNA at the same
site on each side of the M. quadriceps. One week later, the mice
received the DNA constructs similar to the priming. All DNA
constructs for injection were prepared with EndoFree Plasmid
Purification kits from Qiagen.
ELISPOT assay
The ELISPOT assay was performed as described in our
previous study (23). Briefly,
96-well ELISPOT plates (BD Pharmingen, San Diego, CA, USA) were
coated with 5 μg/ml rat anti-mouse IFN-γ antibody in 100 μl of PBS.
After overnight incubation at 4°C, the wells were washed and
blocked with RPMI-1640 culture medium containing 10% fetal calf
serum. Different concentrations of freshly isolated splenocytes
from each vaccinated mouse group (from 1×106 to
1.25×105/well) were added to the wells along with 50
IU/ml IL-2 and 1 μg/ml E7 peptide containing CTL epitope
(H-2Db, aa 49–57) (26).
After a 24-h culture, the plate was washed followed by incubation
with 2.5 μg/ml biotinylated IFN-γ antibodies in 100 μl in PBS
containing 10% fetal calf serum at 4°C overnight. After washing,
avidin-HRP in 100 μl of PBS was added and incubated for 1 h at room
temperature. After washing five times, spots were developed by
adding 100 μl AEC (3-amino-9-ethylcarbazole) solution. The spots
were counted using an ELISPOT Reader system.
Intracytoplasmic cytokine staining and
flow cytometric analysis
To detect E7-specific CD8+ T cell
precursors and E7-specific CD4+ T helper cell responses,
splenocytes from each vaccinated mouse group were incubated either
with 2 μg/ml of E7 peptide (aa 49–57) or with 2 μg/ml of E7 peptide
(aa 30–67) containing MHC class II epitope (27) for 20 h. Golgistop (BD Pharmingen)
was added 6 h before harvesting the cells from the culture. Cells
were then washed once in FACScan buffer and stained with
phycoerythrin (PE)-conjugated monoclonal rat anti-mouse CD8 or CD4
antibody (BD Pharmingen). Cells were subjected to intracellular
cytokine staining using the Cytofix/Cytoperm kit according to the
manufacturer’s instructions (BD Pharmingen). PE-conjugated
anti-IFN-γ or anti-IL-4 antibodies and the FITC-conjugated rat
IgG2a, k or PE-conjugated at IgG1 isotype control antibody were all
purchased from Pharmingen. Analyses were performed on a Beckman
Coulter EPICS XL (Beckman Coulter Inc., Fullerton, CA, USA).
Anti-E7 ELISA
The anti-HPV16 E7 antibodies in the sera were
determined by a direct ELISA as previously described (23,25).
Serially diluted sera collected from mice on day 10
post-immunization were incubated at 4°C overnight with 100 ng of
bacteria-derived HPV16 E7 protein in an ELISA plate. A 1:3,000
dilution of HRP-conjugated goat anti-mouse IgG antibody (Beijing
Zhongshan Golden Bridge Biotechnology) was used. The ELISA plate
was read using a standard ELISA reader at 490 nm.
In vivo tumor protection experiments
For the tumor protection experiment, C57BL/6 mice (7
per group) were vaccinated i.m. with 100 μg of pVR1012-mE7/MtHSP70,
pVR1012-SigmE7/MtHSP70 or pVR1012 vector control twice with a
1-week interval. One week after the last vaccination, mice were
challenged s.c. with 7.5xl04 TC-1 cells per mouse in the
right flank and then monitored twice a week for tumor growth.
In vivo tumor treatment experiments
To test the ability of the DNA vaccination to
inhibit the growth of established tumors, C57BL/6 mice (7 per
group) were s.c. challenged with 7.5xl04 TC-1 cells per
mouse in the right flank. Three days later, mice were immunized
with 100 μg of each plasmid of pVR1012-mE7/MtHSP70,
pVR1012-SigmE7/MtHSP70 and pVR1012 i.m., and the mice were boosted
1 week after the first immunization. Mice were monitored twice a
week for tumor growth.
Data analyses
ELISPOT and FACS data were analyzed using the mean
of two sample comparison of Poisson distribution. ELISA data were
analyzed using the Student’s t-test, and tumor incidence data were
analyzed by the Fisher’s exact probabilities in a 2×2 table. Values
of P<0.05 were considered to indicate a statistically
significant result.
Results
Detection of fusion proteins secreted
from transfected cells in vitro
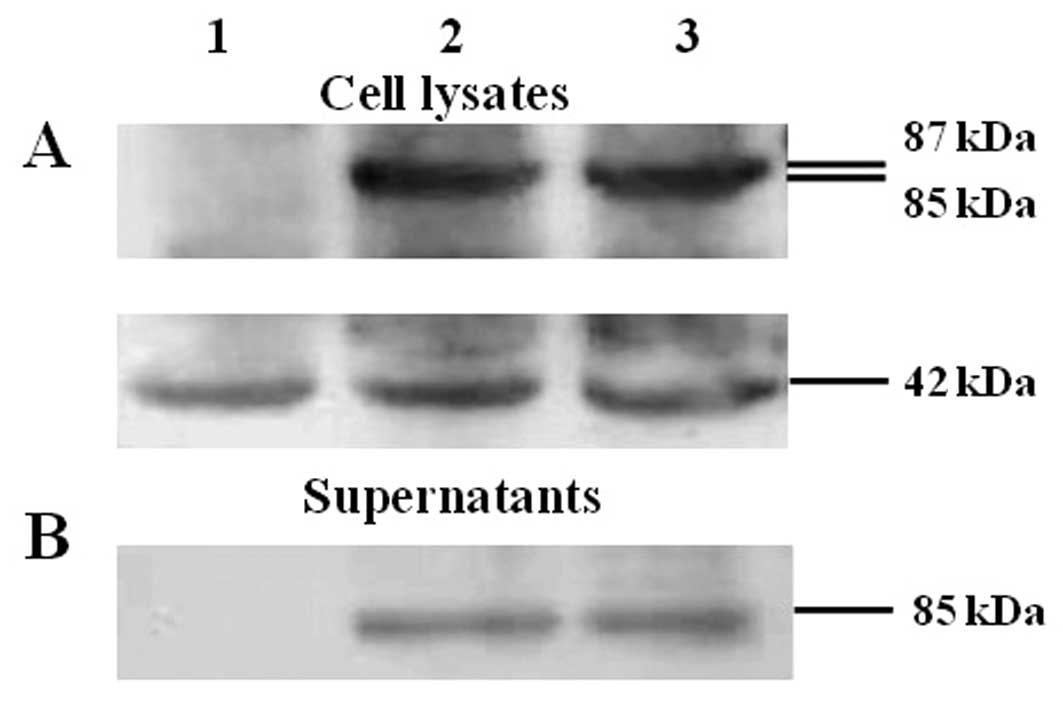
As determined by the western blot results (Fig. 1A and B) mE7 and HSP70 fusion
proteins were confirmed to be expressed both in the culture
supernatants and in the cell lysates of cells after transfection
with mE7/MtHSP70 or SigmE7/MtHSP70 fusion DNA constructs, whereas
cells transfected with pVR1012 showed no expression signal in the
supernatants and in the lysates. β-actin was detected as the
internal loading control (Fig. 1A).
The ratio of mE7/MtHSP70 or SigmE7/MtHSP70 fusion protein vs.
β-actin was 1.21 and 1.12 in the cell lysates as determined by gray
scale scanning analysis from a gel imaging system (Fig. 1C). No difference was observed in the
mE7/MtHSP70 fusion protein expression level between the cell
lysates of cells transfected with the SigmE7/MtHSP70 and
mE7/MtHSP70 fusion DNA constructs (P>0.05). The ratios of
mE7/MtHSP70 or SigmE7/MtHSP70 fusion protein to β-actin were 0.69
and 0.75 in the culture supernatants (Fig. 1C); no significant difference was
observed between the values (P>0.05). These results indicate
that addition of a signal peptide at the upstream of the
mE7/MtHSP70 fusion gene did enhance the secretary expression of the
mE7 and MtHSP70 fusion protein.
mE7/MtHSP70 fusion DNA induces a higher
level of E7-specific CD8+ T cells than SigmE7/MtHSP70
DNA
CD8+ T lymphocytes are one of the most
critical components among antitumor effectors in tumor immunity.
Thus, we examined E7-specific CD8+ T-cell precursor
frequency induced by DNA vaccination. ELISPOT results are shown in
Fig. 2A and B. The number of
E7-specific IFN-γ-producing CD8+ T cells in splenocytes
from the mE7/MtHSP70 fusion DNA immunized mice was greater than 4
times that from the SigmE7/MtHSP70 fusion DNA immunized mice (503
vs. 100 per 5×105 splenocytes; P<0.01). Results of
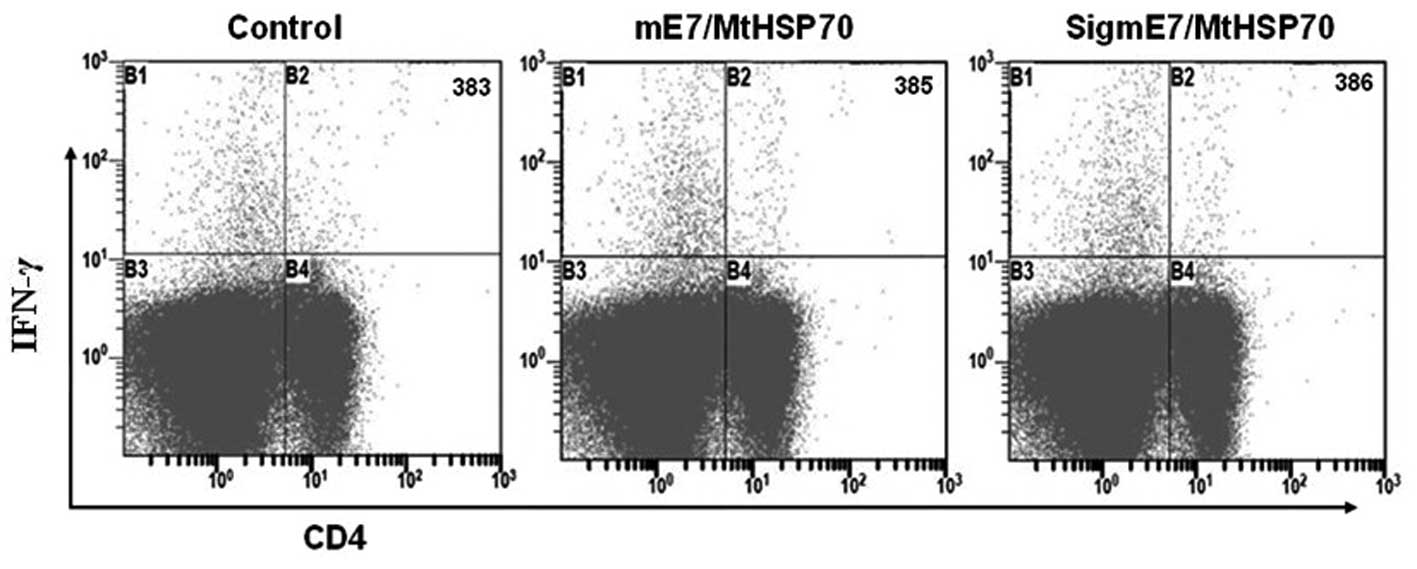
the flow cytometric analysis of E7-specific IFN-γ-producing
CD8+ T cells are shown in Fig. 2C. Subtracting the background
produced by the control (280 cells/3×105 splenocytes),
mice vaccinated with mE7/MtHSP70 fusion DNA generated the highest
number of IFN-γ-secreting CD8+ T-cell precursors (217
cells/3×105 splenocytes), whereas mice vaccinated with
SigmE7/MtHSP70 fusion DNA generated only ~109 IFN-γ-producing
CD8+ T-cell precursors per 3×105 splenocytes
(P<0.01). The results were correlated closely with that of the
ELISPOT results (Fig. 2A and B).
The results indicated that while MtHSP70 or Sig/MtHSP70 linked to
mE7 could both induce the activation of antigen-specific
CD8+ T cells, mE7/MtHSP70 induced a higher level of
E7-specific CD8+ T cell response than SigmE7/MtHSP70
DNA.
Neither the mE7/MtHSP70 nor
SigmE7/MtHSP70 fusion DNA vaccine elicits E7-specific
CD4+ T cell-mediated immune responses
To determine the E7-specific CD4+
T-precursors activated by the vaccines, we performed double
staining for the CD4 surface marker and the intracellular IFN-γ or
IL-4 in the splenocytes after incubation with the E7 peptide (aa
30–67). Results are shown in Fig.
3. There was no significant difference in the number of
E7-specific IFN-γ-secreting CD4 cells as determined using flow
cytometry staining among the various vaccination groups, and no
significant CD4+/IL-4+ double-positive cells
were identified in mice receiving mE7/MtHSP70, SigmE7/MtHSP70 and
control DNA (data not shown). These data indicated that neither
mE7/MtHSP70 nor SigmE7/MtHSP70 fusion DNA vaccine activated
E7-specific Th cell responses.
Neither mE7/MtHSP70 nor SigmE7/MtHSP70
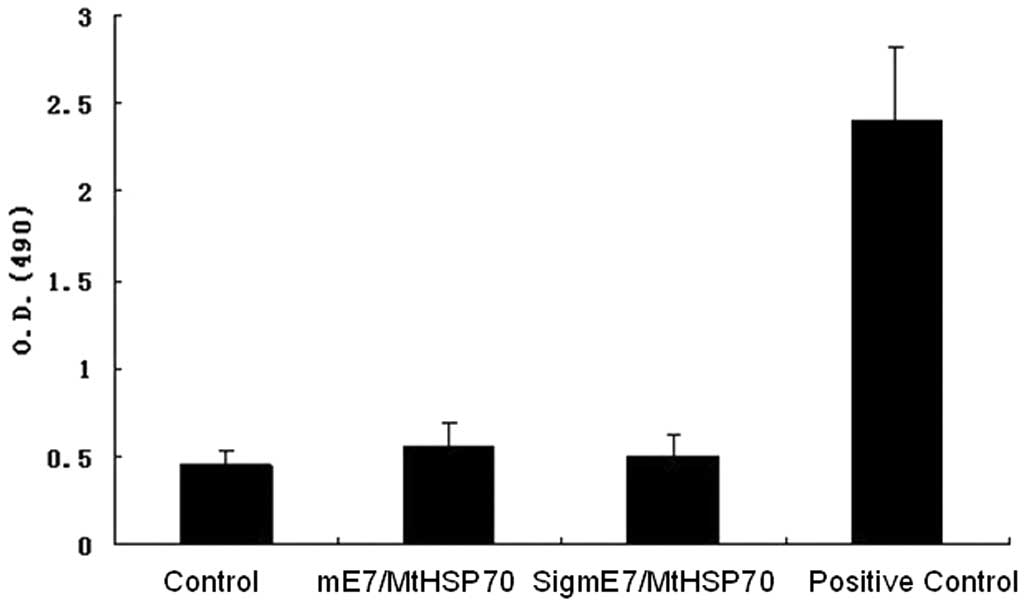
fusion DNA vaccine induces E7-specific antibodies
No E7-specific antibodies were detected in the sera
of mice in any of the vaccinated groups (Fig. 4). The results suggest that the
modifications introduced in the construction of the two DNA
vaccines could not elicit antibody responses.
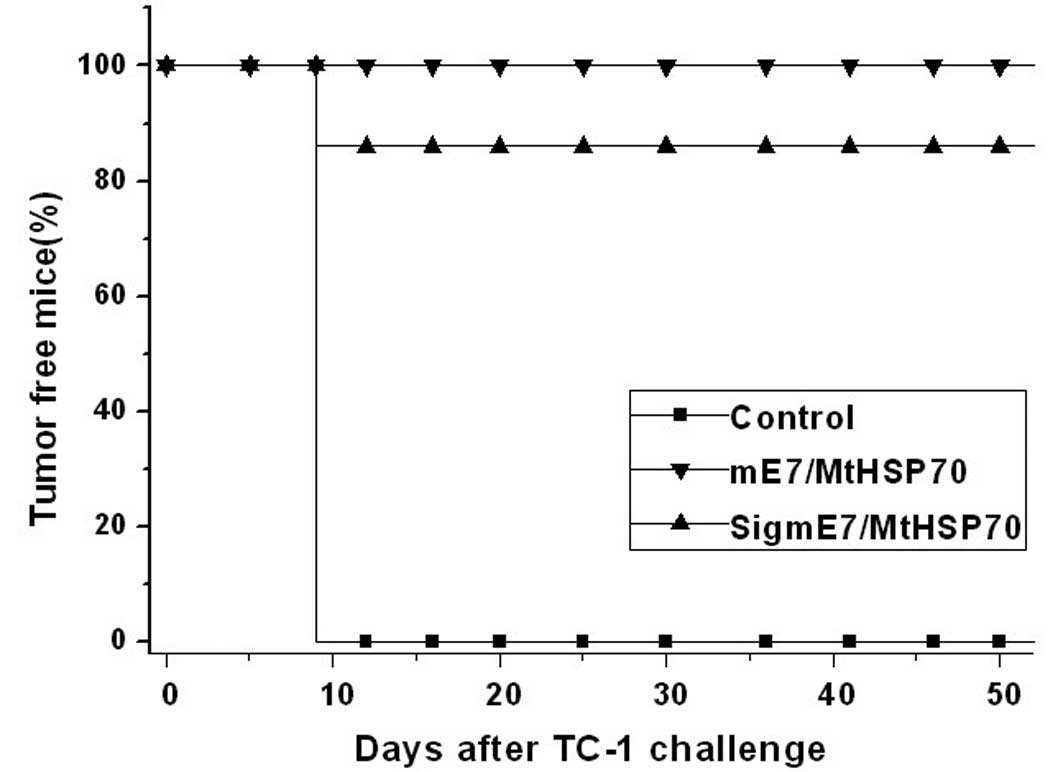
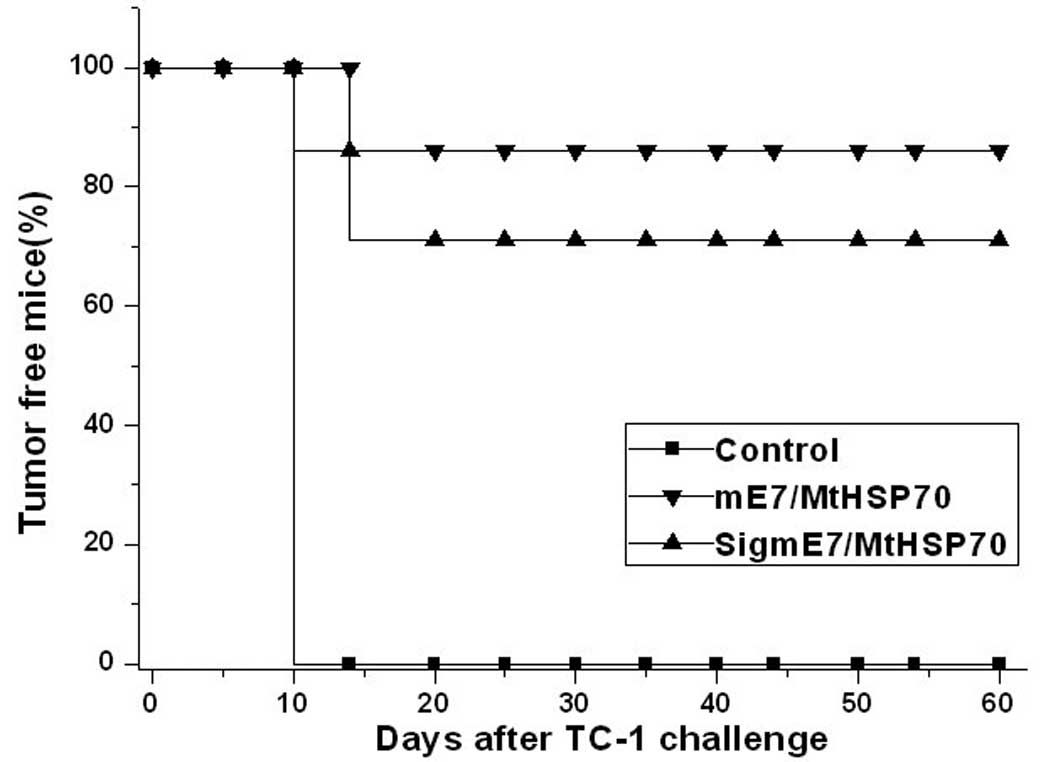
mE7/MtHSP70 and SigmE7/MtHSP70 fusion DNA
vaccines prevent tumors in vivo
As shown Fig. 5,
both mE7/MtHSP70 and SigmE7/MtHSP70 fusion DNA vaccines induced
effective immunity against TC-1 tumors. At ~2 months after TC-1
tumor challenge, 100% of the mice receiving mE7/MtHSP70 fusion DNA
remained tumor free, whereas only 86% of mice receiving
SigmE7/MtHSP70 fusion DNA remained tumor free. In contrast, all of
the mice receiving the empty vector developed a tumor growth on day
12 after tumor challenge. The results demonstrated that the
mE7/MtHSP70 fusion DNA vaccine induced a much stronger antitumor
immune effect, indicating that addition of a secretary signal
peptide at the N terminal of the mE7/MtHSP70 fusion protein did not
enhance the immune effect of the mE7/MtHSP70 antigen.
mE7/MtHSP70 and SigmE7/MtHSP70 fusion DNA
vaccines eradicate tumors in vivo
The therapeutic effects of the two DNA vaccines in
eradicating the established TC-1 tumors are shown in Fig. 6. After immunization, 100% mice
receiving the empty vector developed tumors soon after TC-1 cell
challenge. In contrast, 71% mice receiving the SigmE7/MtHSP70
fusion DNA vaccine were tumor free, and the percentage of
tumor-free mice receiving the mE7/MtHSP70 fusion DNA vaccine was up
to 86% for at least 2 months. The results showed that the
vaccination with either forms of mE7 and MtHSP70 fusion DNA
vaccines could eradicate the established tumors, and the
mE7/MtHSP70 fusion DNA vaccine induced stronger antitumor activity
than the SigmE7/MtHSP70 fusion DNA vaccine. The above results
indicate that addition of a secretary signal peptide at the N
terminal of the mE7/MtHSP70 fusion protein did not enhance the
immune effect of the mE7/MtHSP70 fusion DNA vaccine.
Discussion
Our results showed that antitumor activity of the
two mE7 and MtHSP70 fusion DNA vaccines was mainly dependent on
E7-specific CD8+ T cell responses, whereas
CD4+ T cells and the E7-specific antibody were not
detected, indicating that the antigen presentation pathway involved
in the CD8+ T cell responses are mainly mediated by MHC
class I molecule participating in the antigen presentation pathway.
Our results were consistent with previous reports that specific
CD8+ T cell responses generated by antigen and the HSP70
fusion gene were independent of the help of CD4+ T cells
(11,28). HSP70 plays an important role, not
only in the process of protein folding, transport and degradation,
but also in the participation of directing more efficient antigen
presentation to CD8+ T cells through the MHC class I
pathway (20,29–31).
The possible mechanisms of HSP70 enhancement of CD8+ T
cell responses independent of the help of CD4+ T cells
may involve activation of DCs directly and indirectly to release
proinflammatory cytokines, cross presentation and the intrinsic
molecular chaperone activity of HSP70.
HSP70 was first considered as a cytoplasmic protein.
Afterwards, it was found that HSP70 can also be released from cells
and become an extracellular protein, playing versatile biological
functions. In the present study, expression of the mE7/MtHSP70
fusion protein was observed both in the cell lysates and in culture
supernatants after transfection with either mE7/MtHSP70 or
SigmE7/MtHSP70 fusion DNA constructs. To our surprise, mE7/MtHSP70
fusion proteins can be secreted from cells efficiently even without
the guidance of a signal peptide. Why could HSP70 be released from
cells? At first, release of HSP70 was considered to be a pathologic
phenomenon, i.e. it can be released from cells during cellular
necrosis and cytolysis (32).
Further studies found that several human cancer cell lines could
also secrete HSP70, and the secretion can be increased when these
cells are transfected with HSP70 DNA (33). Some specific cell membrane
microdomains, for example, exosome and endosome lysosomes, may play
important roles in HSP70 exocytosis (34–36).
Our results showed that HSP70 and the antigen fusion protein can be
secreted from cells, and addition of a heterogeneous signal peptide
at the N terminal of mE7/MtHSP70 did not enhance secretion of the
fusion protein, indicating that HSP70 has potent intrinsic
secretion activity and can direct HSP70 and antigen fusion protein
out of the cells via several types of secretary pathways.
The reason why the mE7/MtHSP70 fusion DNA vaccine
produced a more effective CD8+ T cell response and
antitumor activity than the SigmE7/MtHSP70 vaccine can be explained
by the following three reasons. Firstly, in the antigen direct
presentation pathway, DCs intake DNA construct and express
mE7/MtHSP70 fusion protein, providing a favorable advantage for
HSP70 and the antigen to function in the same DC. Thus, HSP70 may
have an opportunity to chaperone E7 peptide cross-presentation to
CD8+ T cells by the MHC class I pathway. HSPs have also
been proposed to be involved in processing MHC class I restricted
antigens (29,37,38).
How HSP70 takes chaperone effects in the form of the mE7/HSP70
fusion protein, deserves further study. In this case, the existence
of a signal peptide in the fusion protein may influence the
chaperone activity of HSP70. This may explain the reason why the
SigmE7/MtHSP70 fusion DNA vaccine had decreased potency compared
with the mE7/MtHSP70 fusion DNA vaccine. Secondly, in the antigen
cross-presentation pathway, when muscle cells and/or DCs intake the
DNA construct, then the synthesize and release the coded protein
out of cells. The secretory protein is then taken up by DCs via
mediation of the endocytic receptors, and present the antigen
peptide to CD8+ T cells. When then mE7/MtHSP70 fusion
protein is secreted from the cells transfected with the
SigmE7/MtHSP70 fusion DNA vaccine, the signal peptide will be cut
off from the fusion protein, so the secretory fusion proteins
produced by the mE7/MtHSP70 or SigmE7/MtHSP70 fusion DNA vaccines
are the same, both in the form of mE7/MtHSP70. As the secretory
levels of mE7/HSP70 fusion proteins from cells transfected with
SigmE7/MtHSP70 and mE7/MtHSP70 fusion DNA vaccines were similar, we
could not explain the different CD8+ T cell responses of
these two DNA vaccines from this aspect. Lastly, another possible
condition, is that when the muscle cells which take up the two DNA
vaccines are dead, the fusion proteins are released out of the
cells in the form of either SigmE7/MtHSP70 or mE7/MtHSP70. In this
case, the existence of the signal peptide may interfere with the
binding of HSP70 to its endocytic receptor on DCs resulting in the
decreased immunogenicity of the SigmE7/MtHSP70 fusion DNA
vaccine.
In summary, we constructed a more effective and
simple HPV 16 therapeutic DNA vaccine that is capable of generating
significantly high levels of antigen-specific antitumor activity
without the addition of a signal peptide gene sequence. Our
observations may serve as an important foundation and significant
reference for future basic research and clinical trials.
Acknowledgements
The present study was supported by a grant from the
National Natural Science Foundation of China (no. 30271355), the
National Natural Science Foundation of China (no. 31070 813) and
the Nature Science Foundation for Young Scholars of Shandong
Province (ZR2010HQ009).
Abbreviations:
|
HPV
|
human papillomavirus
|
|
CTL
|
cytotoxic T lymphocyte
|
|
MHC
|
major histocompatibility complex
|
|
HSP
|
heat shock protein
|
|
APC
|
antigen-presenting cell
|
|
DC
|
dendritic cell
|
|
mE7
|
modified and optimized HPV16 E7
gene
|
|
mE7/MtHSP70
|
modified E7 linked with
Mycobacterium tuberculosis HSP70
|
|
SigmE7/MtHSP70
|
modified E7 linked with
Mycobacterium tuberculosis HSP70 attached with CD33 signal
peptide
|
|
FCS
|
fetal calf serum
|
|
OPD
|
o-phenylenediamine
|
|
TLR
|
toll-like receptor
|
References
|
1
|
Jemal A, Bray F, Center MM, Ferlay J, et
al: Global cancer statistics. CA Cancer J Clin. 61:69–90. 2011.
View Article : Google Scholar
|
|
2
|
Clifford GM, Smith JS, Plummer M, et al:
Human papillomavirus types in invasive cervical cancer worldwide: a
meta-analysis. Br J Cancer. 88:63–73. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Quek SC, Lim BK, Domingo E, et al: Human
papillomavirus type distribution in invasive cervical cancer and
high-grade cervical intraepithelial neoplasia across 5 countries in
Asia. Int J Gynecol Cancer. 23:148–156. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Brown DR, Kjaer SK, Sigurdsson K, et al:
The impact of quadrivalent human papillomavirus (HPV; types 6, 11,
16, and 18) L1 virus-like particle vaccine on infection and disease
due to oncogenic nonvaccine HPV types in generally HPV-naive women
aged 16–26 years. J Infect Dis. 199:926–935. 2009.PubMed/NCBI
|
|
5
|
Kemp TJ, Hildesheim A, Safaeian M, et al:
HPV16/18 L1 VLP vaccine induces cross-neutralizing antibodies that
may mediate cross-protection. Vaccine. 29:2011–2014. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chen CH, Wang TL, Hung CF, et al: Boosting
with recombinant vaccinia increases HPV-16 E7-specific T cell
precursor frequencies of HPV-16 E7-expressing DNA vaccines.
Vaccine. 18:2015–2022. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Brinkman JA, Xu X and Kast WM: The
efficacy of a DNA vaccine containing inserted and replicated
regions of the E7 gene for treatment of HPV-16 induced tumors.
Vaccine. 25:3437–3444. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Eiben GL, Velders MP, Schreiber H, et al:
Establishment of an HLA-A*0201 human papillomavirus type 16 tumor
model to determine the efficacy of vaccination strategies in
HLA-A*0201 transgenic mice. Cancer Res. 62:5792–5799. 2002.
|
|
9
|
Ohlschlager P, Pes M, Osen W, et al: An
improved rearranged human papillomavirus type 16 E7 DNA vaccine
candidate (HPV-16 E7SH) induces an E7 wildtype-specific T cell
response. Vaccine. 24:2880–2893. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Gurunathan S, Klinman DM and Seder RA: DNA
vaccines: immunology, application, and optimization. Annu Rev
Immunol. 18:927–974. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chen CH, Wang TL, Hung CF, et al:
Enhancement of DNA vaccine potency by linkage of antigen gene to an
HSP70 gene. Cancer Res. 60:1035–1042. 2000.PubMed/NCBI
|
|
12
|
Li Y, Subjeck J, Yang G, et al: Generation
of anti-tumor immunity using mammalian heat shock protein 70 DNA
vaccines for cancer immunotherapy. Vaccine. 24:5360–5370. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hauser H, Shen L, Gu QL, et al: Secretory
heat-shock protein as a dendritic cell-targeting molecule: a new
strategy to enhance the potency of genetic vaccines. Gene Ther.
11:924–932. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Milani V, Noessner E, Ghose S, et al: Heat
shock protein 70: role in antigen presentation and immune
stimulation. Int J Hyperthermia. 18:563–575. 2002.PubMed/NCBI
|
|
15
|
Li Z, Menoret A and Srivastava P: Roles of
heat-shock proteins in antigen presentation and cross-presentation.
Curr Opin Immunol. 14:45–51. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Bendz H, Ruhland SC, Pandya MJ, et al:
Human heat shock protein 70 enhances tumor antigen presentation
through complex formation and intracellular antigen delivery
without innate immune signaling. J Biol Chem. 282:31688–31702.
2007. View Article : Google Scholar
|
|
17
|
Udono H, Ichiyanagi T, Mizukami S, et al:
Heat shock proteins in antigen trafficking - implications on
antigen presentation to T cells. Int J Hyperthermia. 25:617–625.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Srivastava P: Interaction of heat shock
proteins with peptides and antigen presenting cells: chaperoning of
the innate and adaptive immune responses. Annu Rev Immunol.
20:395–425. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Osterloh A and Breloer M: Heat shock
proteins: linking danger and pathogen recognition. Med Microbiol
Immunol. 197:1–8. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Binder RJ and Srivastava PK: Peptides
chaperoned by heat-shock proteins are a necessary and sufficient
source of antigen in the cross-priming of CD8+ T cells.
Nat Immunol. 6:593–599. 2005.PubMed/NCBI
|
|
21
|
Massa C, Guiducci C, Arioli I, et al:
Enhanced efficacy of tumor cell vaccines transfected with
secretable hsp70. Cancer Res. 64:1502–1508. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Murshid A, Gong J, Stevenson MA, et al:
Heat shock proteins and cancer vaccines: developments in the past
decade and chaperoning in the decade to come. Expert Rev Vaccines.
10:1553–1568. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zong J, Peng Q, Wang Q, et al: Human HSP70
and modified HPV16 E7 fusion DNA vaccine induces enhanced specific
CD8+ T cell responses and anti-tumor effects. Oncol Rep.
22:953–961. 2009.PubMed/NCBI
|
|
24
|
Trimble CL, Peng S, Kos F, et al: A phase
I trial of a human papillomavirus DNA vaccine for HPV16+
cervical intraepithelial neoplasia 2/3. Clin Cancer Res.
15:361–367. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang QY, Xu YF, Fan DS, et al: Linkage of
modified human papillomavirus type 16 E7 to CD40 ligand
enhances specific CD8+T-lymphocyte induction and
anti-tumour activity of DNA vaccine. Zhongguo Yi Xue Ke Xue Yuan
Xue Bao. 29:584–591. 2007.(In Chinese).
|
|
26
|
Feltkamp MC, Smits HL, Vierboom MP, et al:
Vaccination with cytotoxic T lymphocyte epitope-containing peptide
protects against a tumor induced by human papillomavirus type
16-transformed cells. Eur J Immunol. 23:2242–2249. 1993. View Article : Google Scholar
|
|
27
|
Tindle RW, Fernando GJ, Sterling JC, et
al: A ‘public’ T-helper epitope of the E7 transforming protein of
human papillomavirus 16 provides cognate help for several E7 B-cell
epitopes from cervical cancer-associated human papillomavirus
genotypes. Proc Natl Acad Sci USA. 88:5887–5891. 1991.
|
|
28
|
Hsu KF, Hung CF, Cheng WF, et al:
Enhancement of suicidal DNA vaccine potency by linking
Mycobacterium tuberculosis heat shock protein 70 to an
antigen. Gene Ther. 8:376–383. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Liu H, Wu BH, Rowse GJ, et al: Induction
of CD4-independent E7-specific CD8+ memory response by
heat shock fusion protein. Clin Vaccine Immunol. 14:1013–1023.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Huang Q, Richmond JF, Suzue K, et al: In
vivo cytotoxic T lymphocyte elicitation by mycobacterial heat shock
protein 70 fusion proteins maps to a discrete domain and is
CD4+ T cell independent. J Exp Med. 191:403–408. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Delneste Y, Magistrelli G, Gauchat J, et
al: Involvement of LOX-1 in dendritic cell-mediated antigen
cross-presentation. Immunity. 17:353–362. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Saito K, Dai Y and Ohtsuka K: Enhanced
expression of heat shock proteins in gradually dying cells and
their release from necrotically dead cells. Exp Cell Res.
310:229–236. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wang MH, Grossmann ME and Young CY: Forced
expression of heat-shock protein 70 increases the secretion of
Hsp70 and provides protection against tumour growth. Br J Cancer.
90:926–931. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Mambula SS, Stevenson MA, Ogawa K, et al:
Mechanisms for Hsp70 secretion: crossing membranes without a
leader. Methods. 43:168–175. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Broquet AH, Thomas G, Masliah J, et al:
Expression of the molecular chaperone Hsp70 in detergent-resistant
microdomains correlates with its membrane delivery and release. J
Biol Chem. 278:21601–21606. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Lancaster GI and Febbraio MA:
Exosome-dependent trafficking of HSP70: a novel secretory pathway
for cellular stress proteins. J Biol Chem. 280:23349–23355. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Srivastava PK, Udono H, Blachere NE, et
al: Heat shock proteins transfer peptides during antigen processing
and CTL priming. Immunogenetics. 39:93–98. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Srivastava PK and Udono H: Heat shock
protein-peptide complexes in cancer immunotherapy. Curr Opin
Immunol. 6:728–732. 1994. View Article : Google Scholar : PubMed/NCBI
|